
Thrombin Receptor-Activating Protein (TRAP)-Activated Akt Is Involved in the Release of Phosphorylated-HSP27 (HSPB1) from Platelets in DM Patients

Abstract
:1. Introduction
2. Results
2.1. Characterization of the Subjects
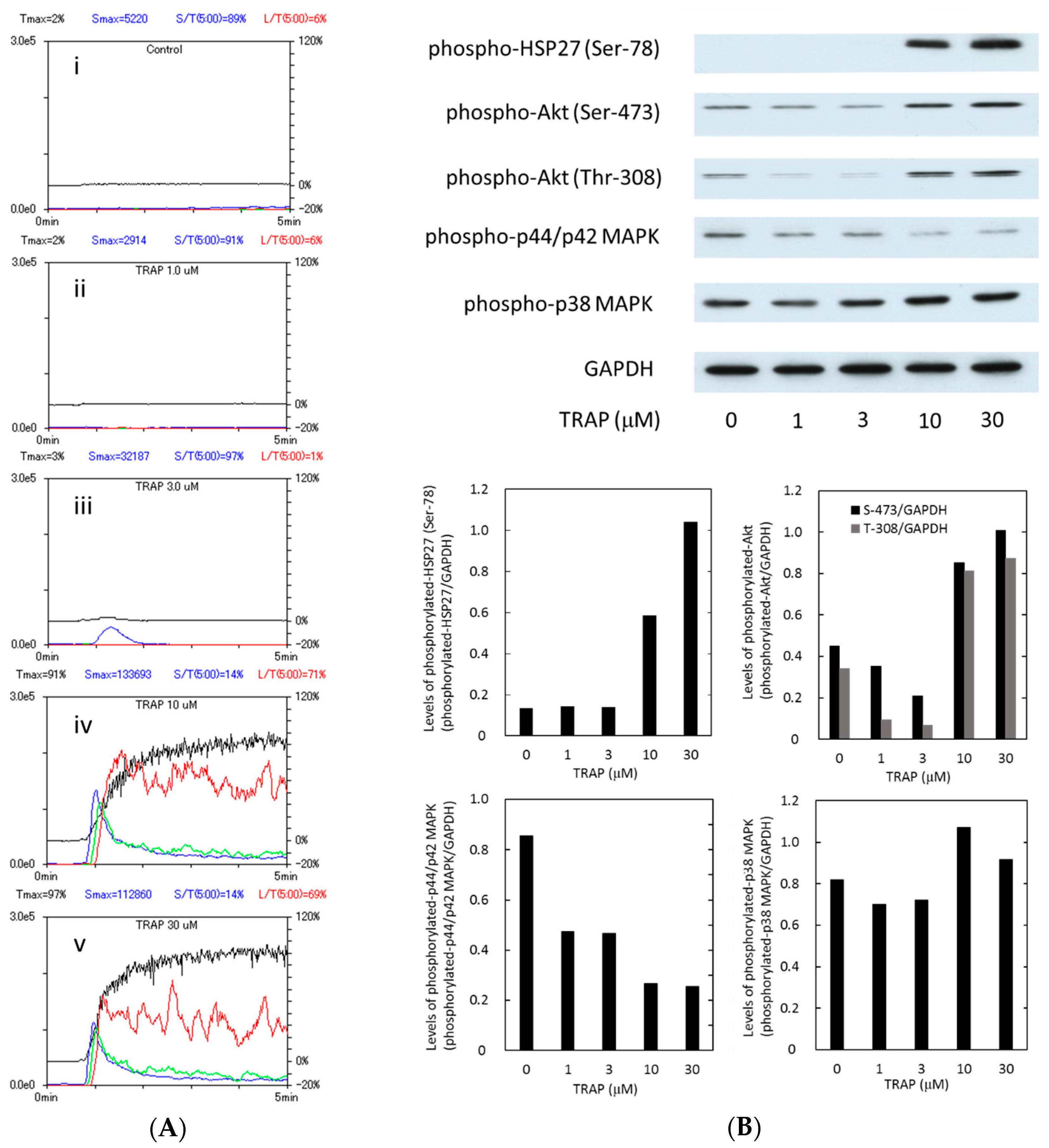
2.2. Platelet Aggregation and the Phosphorylation of Heat Shock Protein 27 (HSP27), Akt, p44/p42 Mitogen-Activated Protein (MAP) Kinase and p38 MAP Kinase in the Subjects
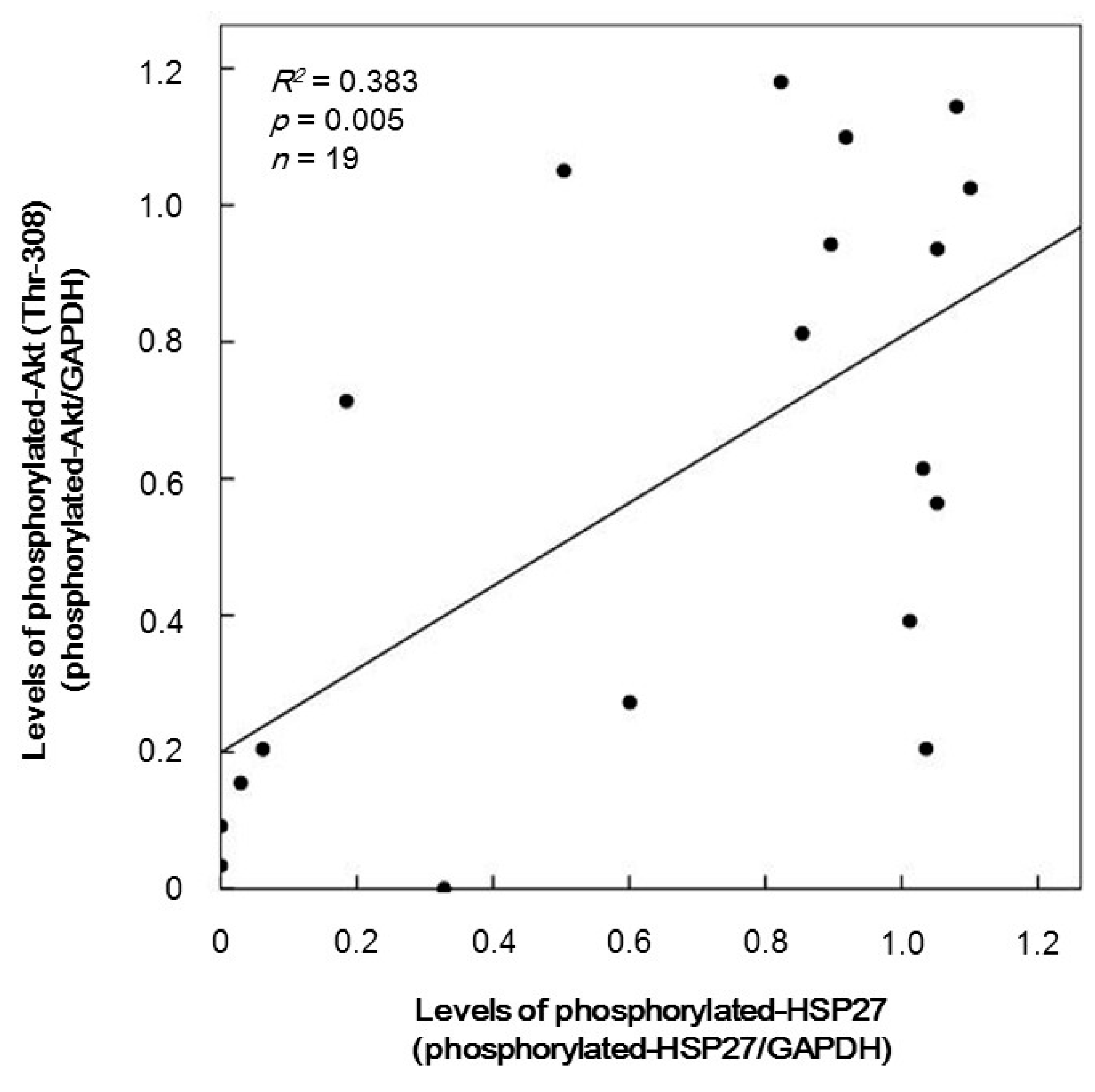
2.3. The Relationship between Levels of Phosphorylated-HSP27 and Those of Phosphorylated-Akt (Thr-308) Induced by TRAP in the Platelets of Type 2 Diabetes Mellitus (DM) Patients
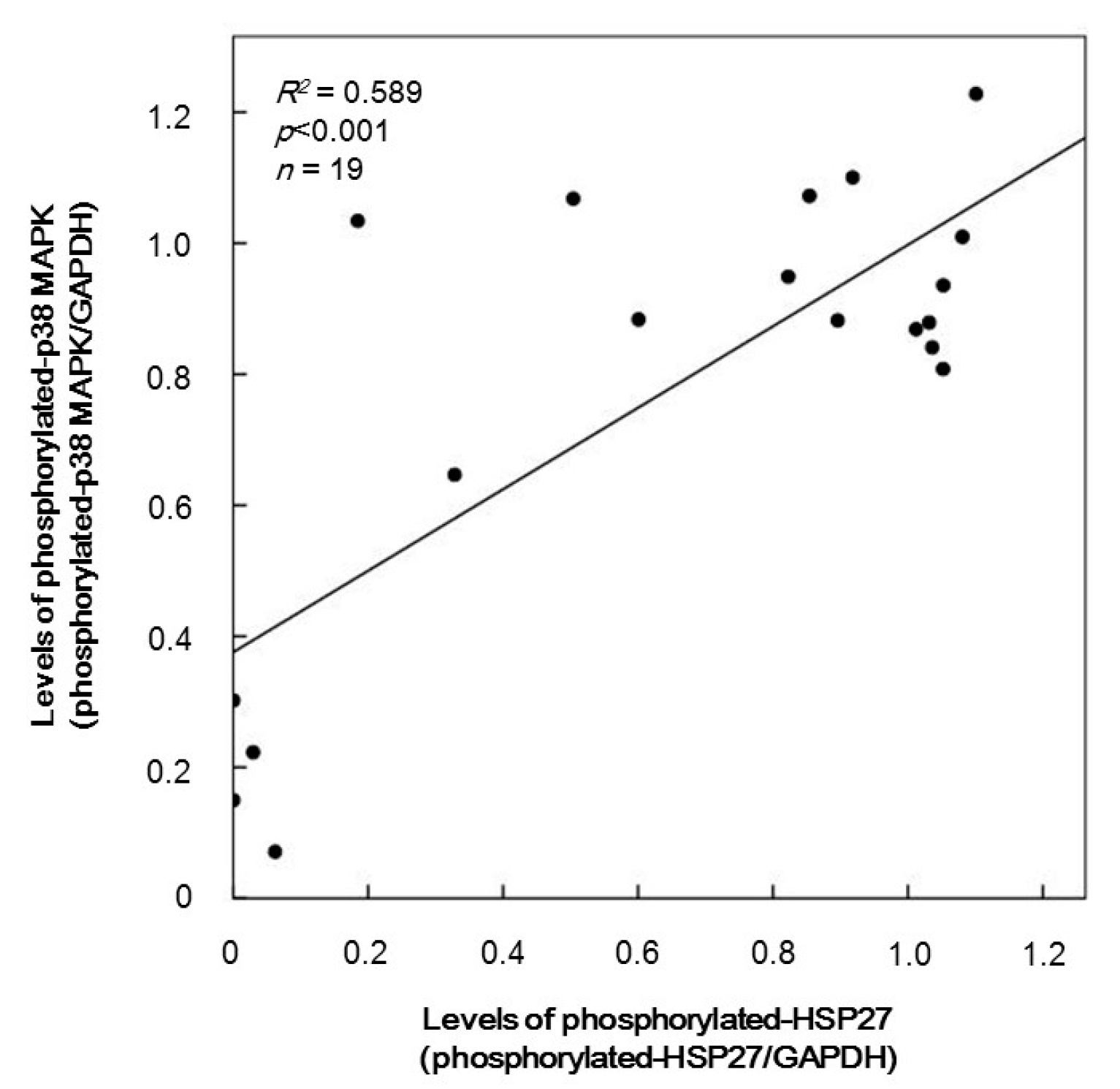
2.4. The Relationship between Levels of Phosphorylated-HSP27 and Those of Phosphorylated-p38 MAP Kinase Induced by TRAP in the Platelets from Type 2 DM Patients
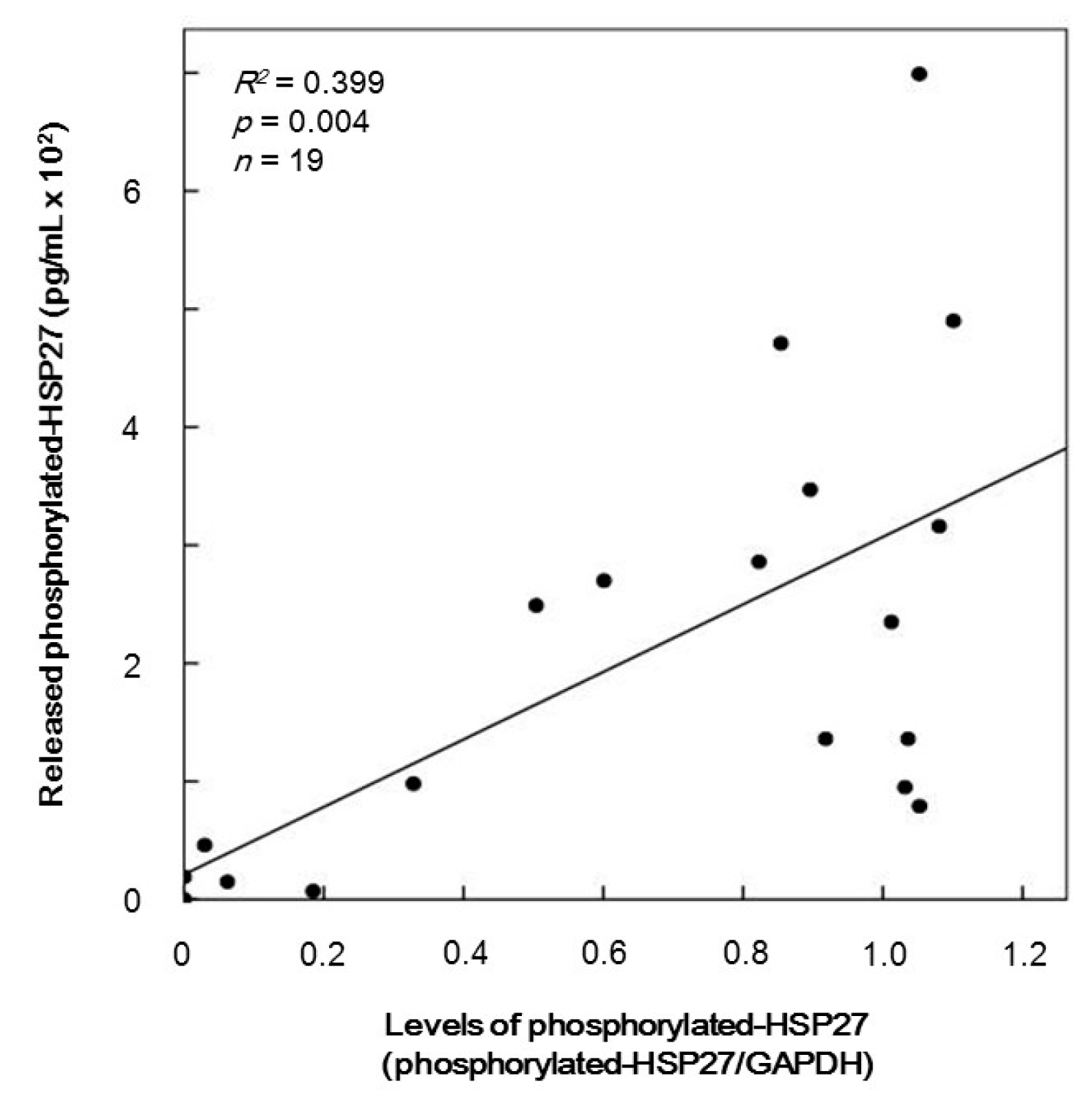
2.5. The Relationship between Levels of HSP27 Phosphorylation and Levels of Released Phosphorylated-HSP27 Induced by TRAP in the Platelets of Type 2 DM Patients
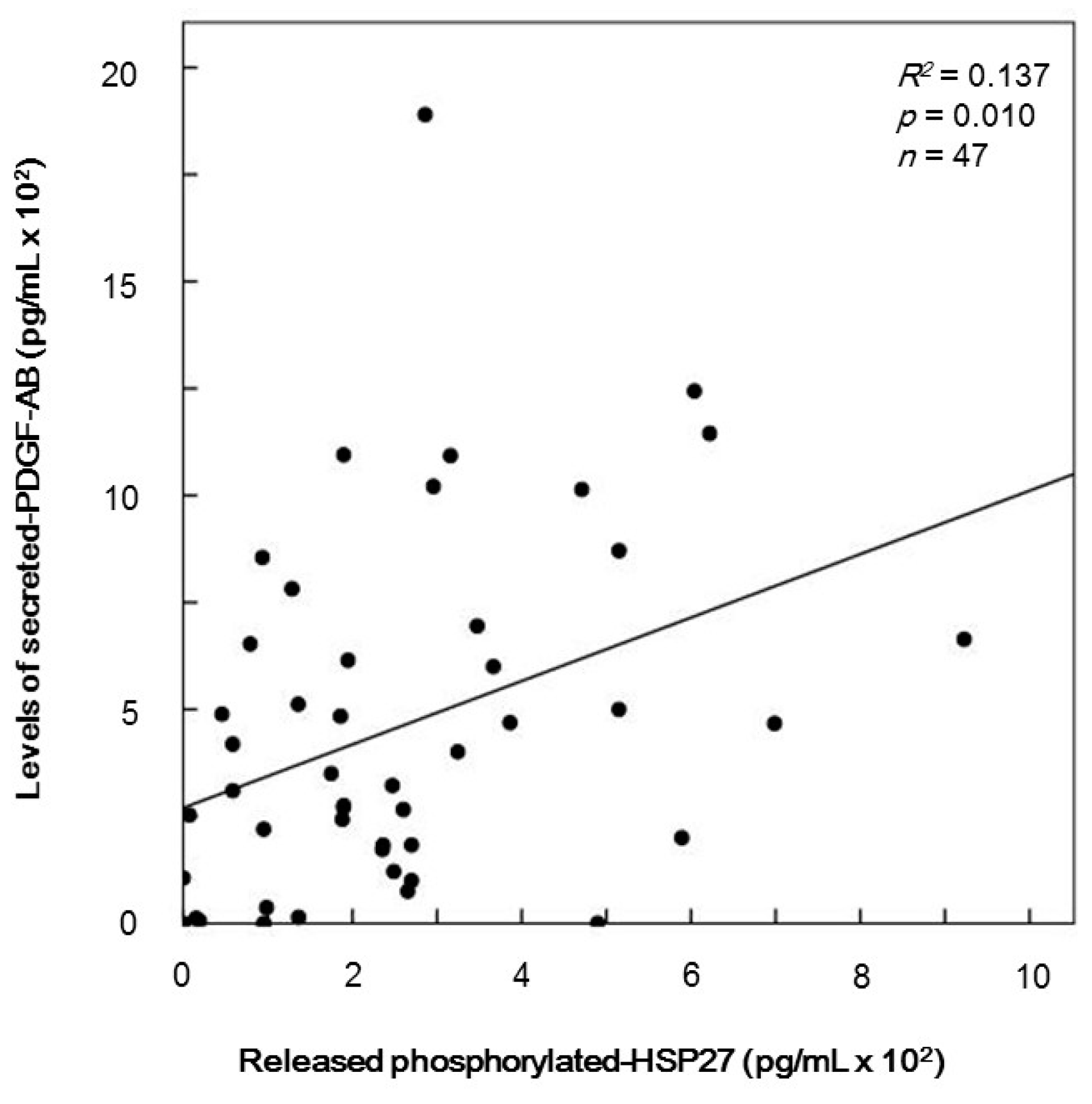
2.6. The Relationship between Levels of Secreted Platelet-Derived Growth Factor-AB (PDGF-AB) and Levels of Release of Phosphorylated-HSP27 Induced by TRAP from the Platelets of Type 2 DM Patients
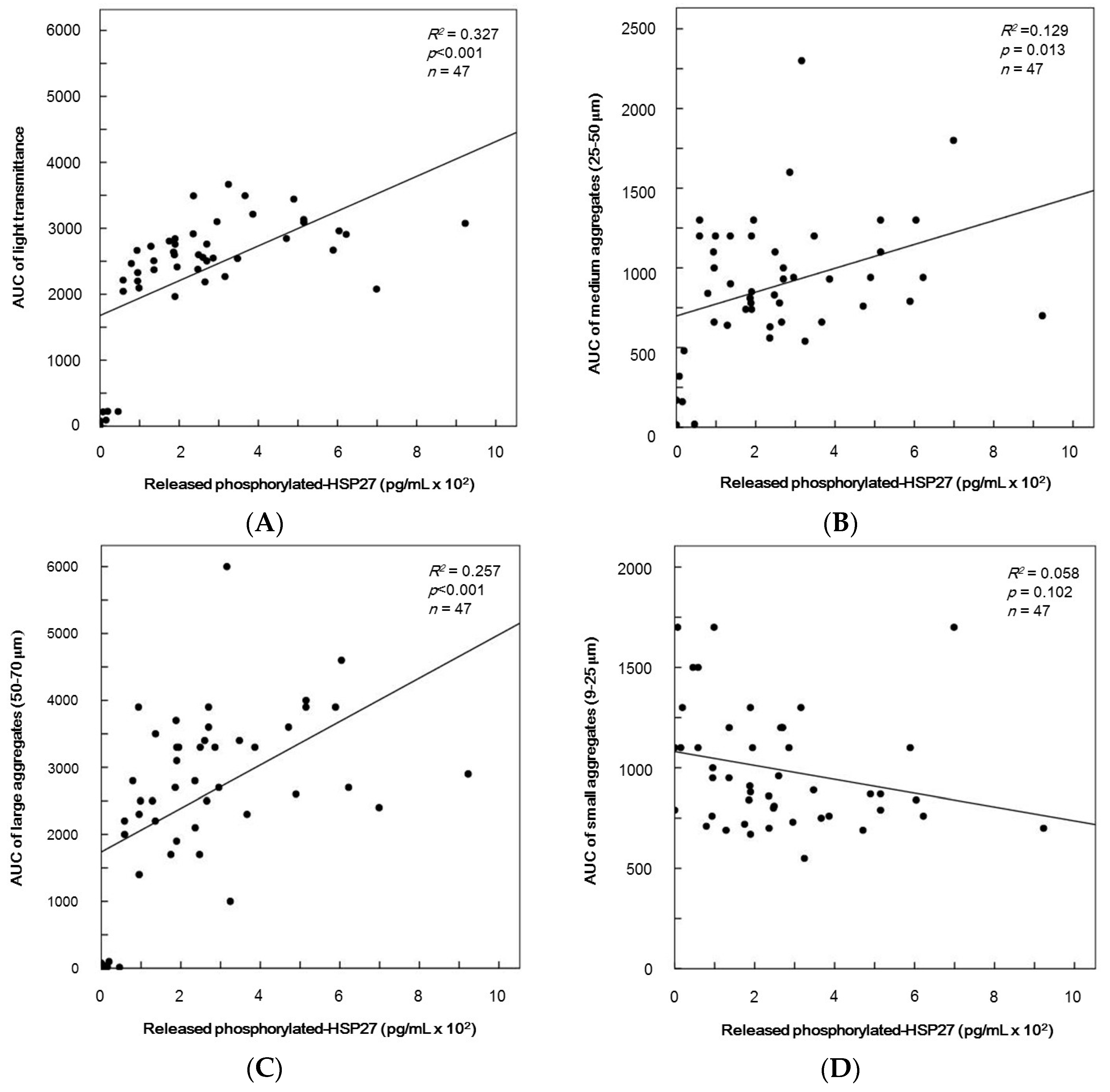
2.7. The Relationship between Parameters for Platelet Aggregation and the Levels of Release of Phosphorylated-HSP27 Induced by TRAP in Type 2 DM Patients
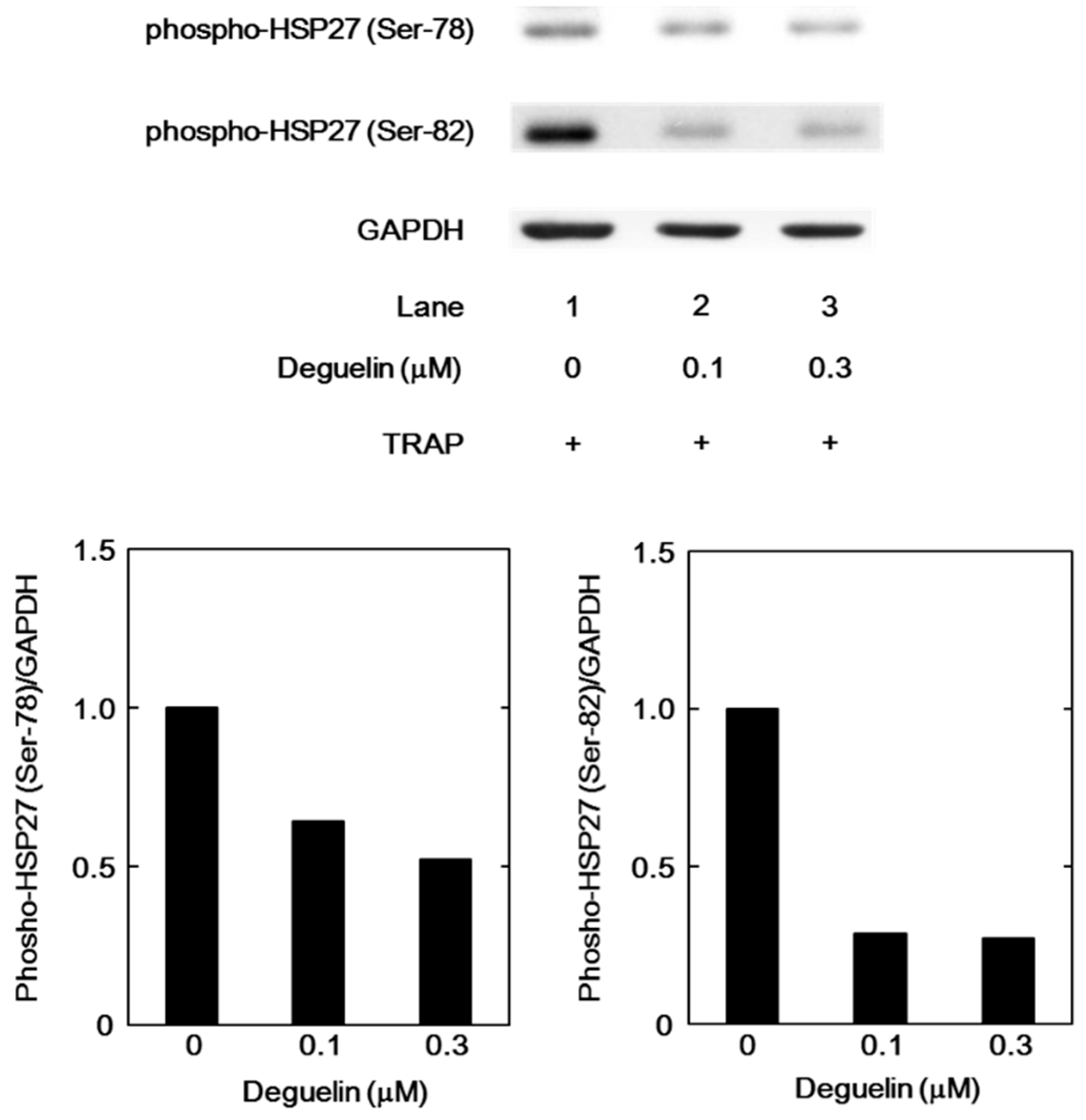
2.8. The Effect of Deguelin on the TRAP-Induced Phosphorylation of HSP27 in Human Platelets
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Subjects
4.3. Blood Sampling
4.4. Platelet Aggregation
4.5. Western Blot Analysis
4.6. ELISA for PDGF-AB and Phosphorylated-HSP27
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HSP27 | heat shock protein 27 |
| MAP | mitogen-activated protein |
| TRAP | thrombin-activating protein |
| DM | diabetes mellitus |
| AUC | area under the curve |
| GP | glycoprotein |
| ADP | adenosine diphosphate |
| PDGF-AB | platelet-derived growth factor-AB |
| PAR | protease-activated receptor |
| PDK1 | 3-phosphoinositide-dependent protein kinase-1 |
| PRP | platelet-rich plasma |
| PPP | platelet-poor plasma |
| PAGE | polyacrylamide gel electrophoresis |
| PVDF | polyvinylidene fluoride |
References
- Davi, G.; Patrono, C. Platelet activation and atherothrombosis. N. Engl. J. Med. 2007, 357, 2482–2494. [Google Scholar] [CrossRef] [PubMed]
- Furie, B.; Furie, B.C. Mechanism of thrombus formation. N. Engl. J. Med. 2008, 359, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, Y.; Maruyama, I.; Soejima, Y. Thrombin receptor agonist peptide decreases thrombomodulin activity in cultured umblical endothelial cells. Biochem. Biophys. Res. Commun. 1994, 199, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.R.; Seatter, M.J.; Kanke, T.; Hunter, G.D.; Plevin, R. Protease-activated receptors. Pharmacol. Rev. 2001, 53, 245–282. [Google Scholar] [PubMed]
- Zimmet, P.; Albert, K.G.; Shaw, J. Global and social implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Benjamin, I.J.; Brurke, G.L.; Chait, A.; Eckel, R.H.; Howard, B.V.; Mitch, W.; Smith, S.C., Jr.; Sowers, J.R. Diabetes and cardiovasucular disease: A statement for healthcare professionals from the American Heart Association. Circulation 1999, 100, 1134–1146. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, H.; Tokuda, H.; Ishisaki, A.; Zhou, Y.; Kitajima, Y.; Kozawa, O. P2Y12 receptors play a significant role in the development of platelet microaggregation in patients with diabetes. J. Clin. Endocrinol. Metab. 2005, 90, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Hanai, Y.; Adachi, S.; Yasuda, I.; Takai, S.; Matsushima-Nishiwaki, R.; Kato, H.; Enomoto, Y.; Akamatsu, S.; Sakakibara, S.; Ogura, S.; et al. Collagen-induced p38 MAP kinase activation is a biomarker of platelet hyper-aggregation in patients with diabetes mellitus. Life Sci. 2009, 85, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Mymrikov, E.V.; Seir-Nebi, A.S.; Gusev, N.B. Large potentials of small heat shock proteins. Physiol. Rev. 2011, 91, 1123–1159. [Google Scholar] [CrossRef] [PubMed]
- Bruey, J.M.; Ducasse, C.; Bonniaud, P.; Ravagnan, L.; Susin, S.A.; Diaz-Latoud, C.; Gurbuxani, S.; Arrigo, A.P.; Kroemer, G.; Solary, E.; et al. Hsp27 negatively regulates cell death by interacting with cytochrome c. Nat. Cell. Biol. 2000, 2, 645–652. [Google Scholar] [PubMed]
- Arrigo, A.P. The cellular “networking” of mammalian Hsp27 and its functions in the control of protein folding, redox state and apoptosis. Adv. Exp. Med. Biol. 2007, 94, 14–26. [Google Scholar]
- Pockley, A.G. Heat shock proteins, inflammation, and cardiovascular disease. Circulation 2002, 105, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Landry, J.; Lambert, H.; Zhou, M.; Lavoie, J.N.; Hickey, E.; Weber, L.A.; Anderson, C.W. Human HSP27 is phosphorylated at serines 78 and 82 by heat shock and mitogen-activated kinases that recognize the same amino acid motif as S6 kinase II. J. Biol. Chem. 1992, 267, 794–803. [Google Scholar] [PubMed]
- Acunzo, J.; Katsogiannou, M.; Rocchi, P. Small heat shock proteins HSP27 (HspB1), αB-crystallin (HspB5) and HSP22 (HspB8) as regulators of cell death. Int. J. Biochem. Cell Biol. 2012, 44, 1622–1631. [Google Scholar] [CrossRef] [PubMed]
- Mendelshon, M.E.; Zhu, Y.; O’Neill, S. The 29-kDa proteins phosphorylated in thrombin-activated human platelets are forms of the estrogen receptor-related 27-kDa heat shock proten. Proc. Natl. Acad. Sci. USA 1991, 88, 11212–11216. [Google Scholar] [CrossRef]
- Saklatvala, J.; Rawlinson, L.; Waller, R.J.; Starsfield, S.; Lee, J.C.; Morton, L.F.; Barnes, M.J.; Farndale, R.W. Role of p38 mitogen-activated protein kinase in platelet aggregation caused by collagen or a thromboxane analogue. J. Biol. Chem. 1996, 271, 6586–6589. [Google Scholar] [PubMed]
- Kageyama, Y.; Doi, T.; Akamatsu, S.; Kuroyanagi, G.; Kondo, A.; Mizutani, J.; Otsuka, T.; Tokuda, H.; Kozawa, O.; Ogura, S. Rac regulates collagen-induced HSP27 phosphorylation via p44/p42 MAP kinase in human platelets. Int. J. Mol. Med. 2013, 32, 813–818. [Google Scholar] [PubMed]
- Tokuda, H.; Kuroyanagi, G.; Tsujimoto, M.; Enomoto, Y.; Matsushima-Nishiwaki, R.; Onuma, T.; Kojima, A.; Doi, T.; Tanabe, K.; Akamatsu, S.; et al. Release of phosphorylated HSP27 (HSPB1) from platelets is accompanied with the accerelation of aggregation in diabetic patients. PLoS ONE 2015, 10, e0128977. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.; Li, D.; Zhang, L.; Wang, K.; Luo, K.; Yang, Z.; Wu, Y.; Liu, J. PDK1 regulates platelet activation and arterial thrombosis. Blood 2013, 121, 3718–3726. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Takai, S.; Matsushima-Nishiwaki, R.; Adachi, S.; Minamitani, C.; Otsuka, T.; Tokuda, H.; Akamatsu, S.; Doi, T.; Ogura, S.; et al. HSP27 phosphorylation is correlated with ADP-induced platelet granule secretion. Arch. Biochem. Biophys. 2008, 475, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Toker, A.; Newton, A.C. Cellular signaling: Pivoting around PDK-1. Cell 2000, 103, 185–188. [Google Scholar] [CrossRef]
- Nakajima, K.; Hirade, K.; Ishisaki, A.; Matsuno, H.; Suga, H.; Kanno, Y.; Shu, E.; Kitajima, Y.; Katagiri, Y.; Kozawa, O. Akt regulates thrombin-induced HSP27 phosphorylation in aortic smooth muscle cells: Function at a point downstream from p38 MAP kinase. Life Sci. 2005, 77, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Widmann, C.; Gibson, S.; Jarpe, M.B.; Johnson, G.L. Mitogen-activated protein kinase: Conservation of a three-kinase module from yeast to human. Physiol. Rev. 1999, 79, 143–180. [Google Scholar] [PubMed]
- Chun, K.H.; Kosmeder, J.W., 2nd; Sun, S.; Pezzuto, J.M.; Lotan, R.; Hong, W.K.; Lee, H.Y. Effects of deguelin on the phosphatidylinositol 3-kinase/Akt pathway and apoptosis in premalignant human bronchial epithelial cells. J. Natl. Cancer Inst. 2003, 95, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.; Chen, Y.-K.; Mcnulty, M.; Simard, T.; Zhao, X.; Wells, D.J.; de Belleroche, J.; O’Brien, E.R. Extracellular release of the atheroprotectrive heat shock protein 27 is mediated by estrogen and competitively inhibits acLDL binding to scavenger receptor-A. Circ. Res. 2008, 103, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Thuringer, D.; Jego, G.; Wettstein, G.; Terrier, O.; Cronier, L.; Yousfi, N.; Hebrard, S.; Bouchot, A.; Hazoume, A.; Joly, A.L.; et al. Extracellular HSP27 mediates angiogenesis through Toll-like receptor 3. FASEB J. 2013, 27, 4169–4183. [Google Scholar] [CrossRef] [PubMed]
- Salari, S.; Seibert, T.; Chen, Y.X.; Hu, T.; Shi, C.; Zhao, X.; Cuerrier, C.M.; Raizman, J.E.; O’Brien, E.R. Extracellular HSP27 acts as a signaling molecule to activate NF-κB in macrophages. Cell Stress Chaperones 2013, 18, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Cleveland, J.C.; Ao, L.; Zeng, Q.; Fullerton, D.A.; Meng, X. Human myocardium releases heat shock protein 27 (HSP27) after global ischemia: The proinflammatory effect of extracellular HSP through toll-like receptor (TLR)-2 and TLR4. Mol. Med. 2014, 20, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Lundstöm, A.; Laska, A.C.; von Arbin, M.; Jörneskog, G.; Wallėn, H. Glucose intolerance and insulin resistance as predictors of low platelet response to clopidogrel in patients with minor ischemic stroke or TIA. Platelets 2014, 25, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Ito, H.; Hasegawa, K.; Inaguma, Y.; Kozawa, O.; Asano, T. Modulation of the stress-induced synthesis of hsp27 and αB crystallin by cyclic AMP in C6 rat glioma cells. J. Neurochem. 1996, 66, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | For Western Blotting | For ELISA |
|---|---|---|
| Total number | 19 | 47 |
| Gender (F/M) | (12/7) | (22/25) |
| Age (years) | 71.5 ± 4.9 | 72.0 ± 6.6 |
| DM duration (years) | 15.1 ± 8.3 | 14.1 ± 7.6 |
| Height (cm) | 157.3 ± 7.7 | 157.1 ± 9.8 |
| Weight (kg) | 59.9 ± 8.5 | 59.5 ± 12.8 |
| BMI | 24.1 ± 2.7 | 23.9 ± 3.5 |
| sBP (mmHg) | 115.2 ± 17.3 | 119.8 ± 18.9 |
| dBP (mmHg) | 65.3 ± 11.5 | 67.1 ± 11.3 |
| HbA1c (%) | 8.4 ± 1.2 | 8.5 ± 1.3 |
| Glu (mg/dL) | 167.0 ± 57.5 | 165.3 ± 56.0 |
| TC (mg/dL) | 189.4 ± 42.2 | 184.1 ± 37.9 |
| TG (mg/dL) | 146.1 ± 109.3 | 130.3 ± 77.7 |
| HDL (mg/dL) | 49.0 ± 16.6 | 49.5 ± 13.7 |
| Plt (×104/µL) | 19.1 ± 4.3 | 21.0 ± 4.9 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokuda, H.; Kuroyanagi, G.; Tsujimoto, M.; Matsushima-Nishiwaki, R.; Akamatsu, S.; Enomoto, Y.; Iida, H.; Otsuka, T.; Ogura, S.; Iwama, T.; et al. Thrombin Receptor-Activating Protein (TRAP)-Activated Akt Is Involved in the Release of Phosphorylated-HSP27 (HSPB1) from Platelets in DM Patients. Int. J. Mol. Sci. 2016, 17, 737. https://doi.org/10.3390/ijms17050737
Tokuda H, Kuroyanagi G, Tsujimoto M, Matsushima-Nishiwaki R, Akamatsu S, Enomoto Y, Iida H, Otsuka T, Ogura S, Iwama T, et al. Thrombin Receptor-Activating Protein (TRAP)-Activated Akt Is Involved in the Release of Phosphorylated-HSP27 (HSPB1) from Platelets in DM Patients. International Journal of Molecular Sciences. 2016; 17(5):737. https://doi.org/10.3390/ijms17050737
Chicago/Turabian StyleTokuda, Haruhiko, Gen Kuroyanagi, Masanori Tsujimoto, Rie Matsushima-Nishiwaki, Shigeru Akamatsu, Yukiko Enomoto, Hiroki Iida, Takanobu Otsuka, Shinji Ogura, Toru Iwama, and et al. 2016. "Thrombin Receptor-Activating Protein (TRAP)-Activated Akt Is Involved in the Release of Phosphorylated-HSP27 (HSPB1) from Platelets in DM Patients" International Journal of Molecular Sciences 17, no. 5: 737. https://doi.org/10.3390/ijms17050737