
Identification and Characterization of the Gene CYP340W1 from Plutella xylostella and Its Possible Involvement in Resistance to Abamectin



Abstract
:1. Introduction
2. Results
2.1. Cloning and Characterization of the CYP340W1 Gene
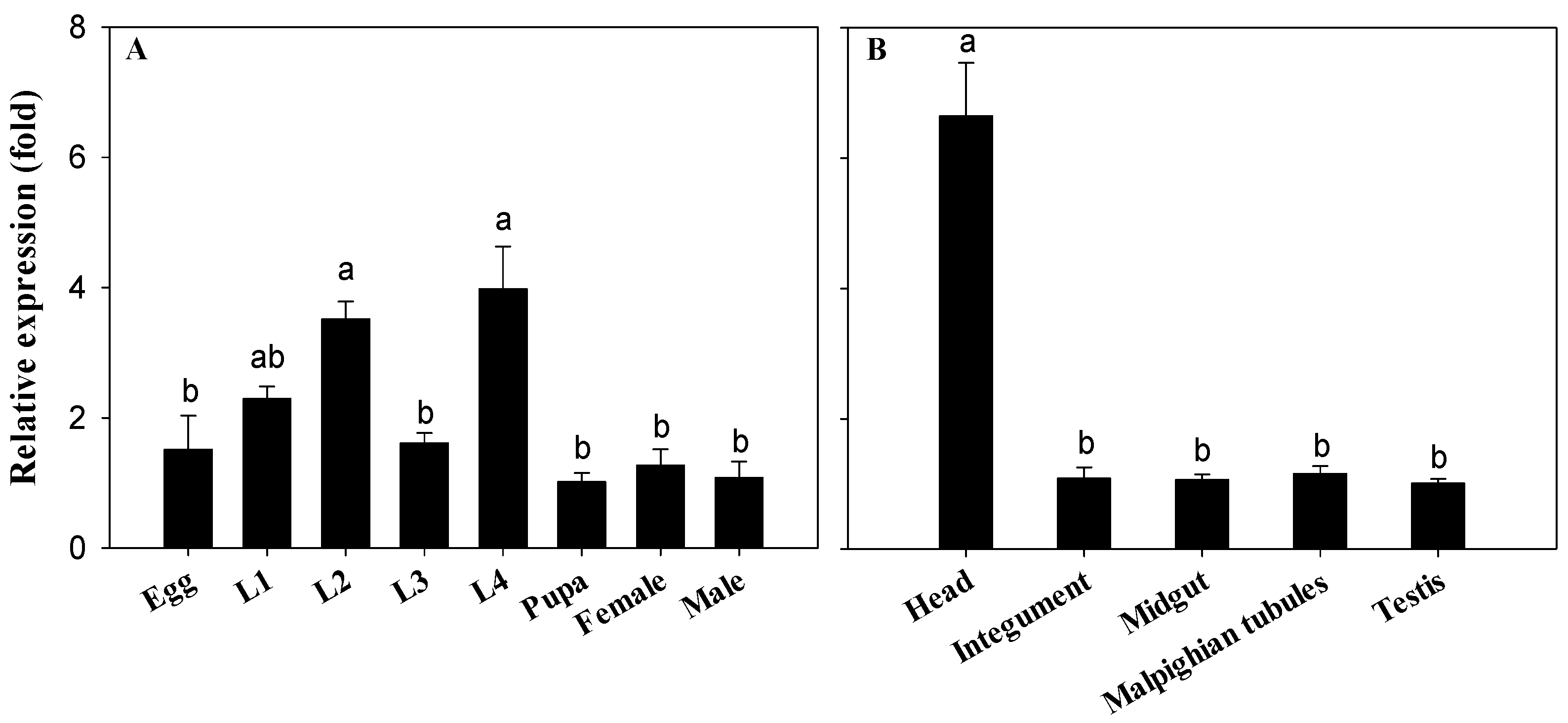
2.2. Expression Profile of CYP340W1 in Different Developmental Stages and Tissues of P. xylostella
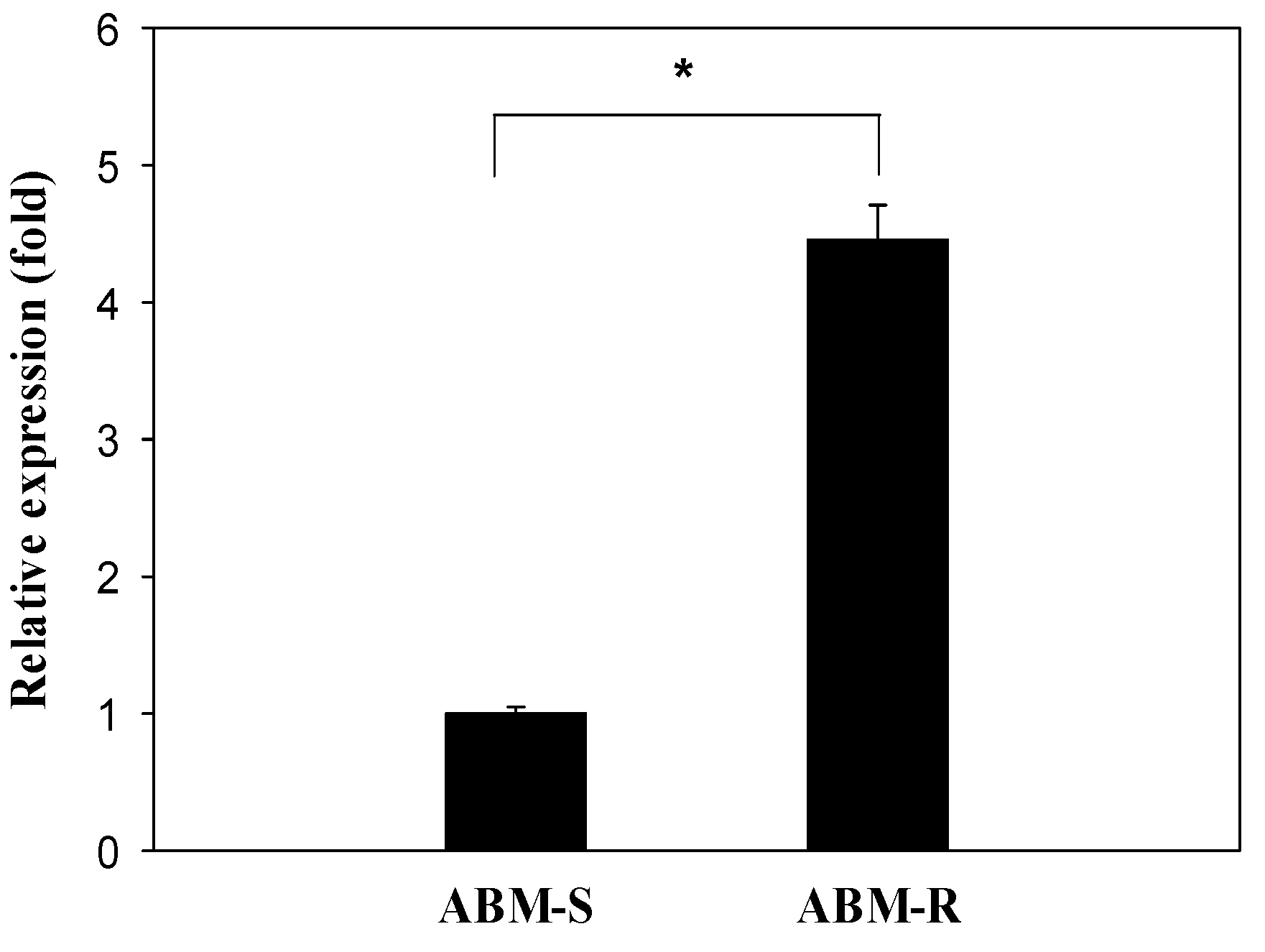
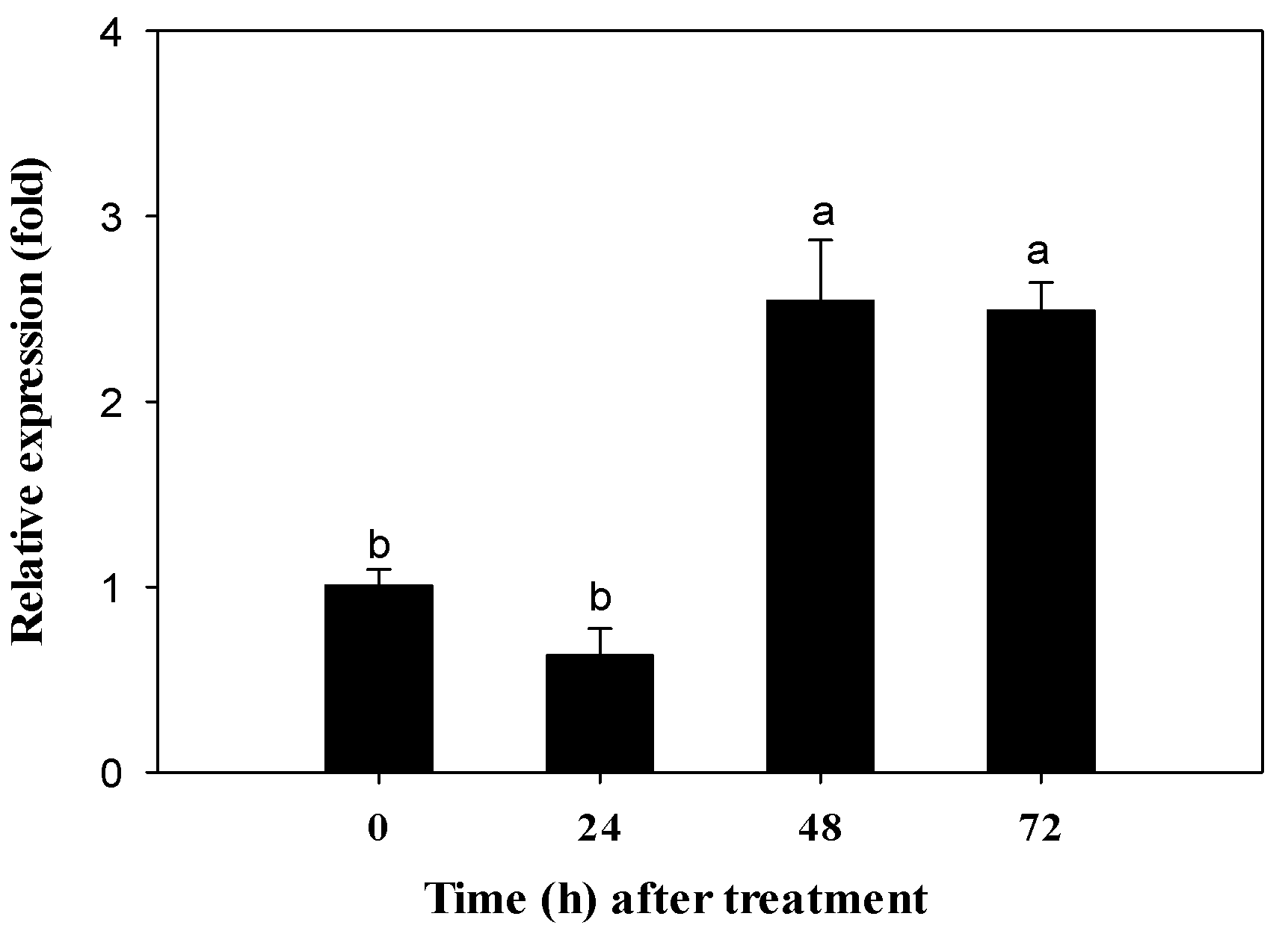
2.3. Transcriptional Response of CYP340W1 to Abamectin Exposure
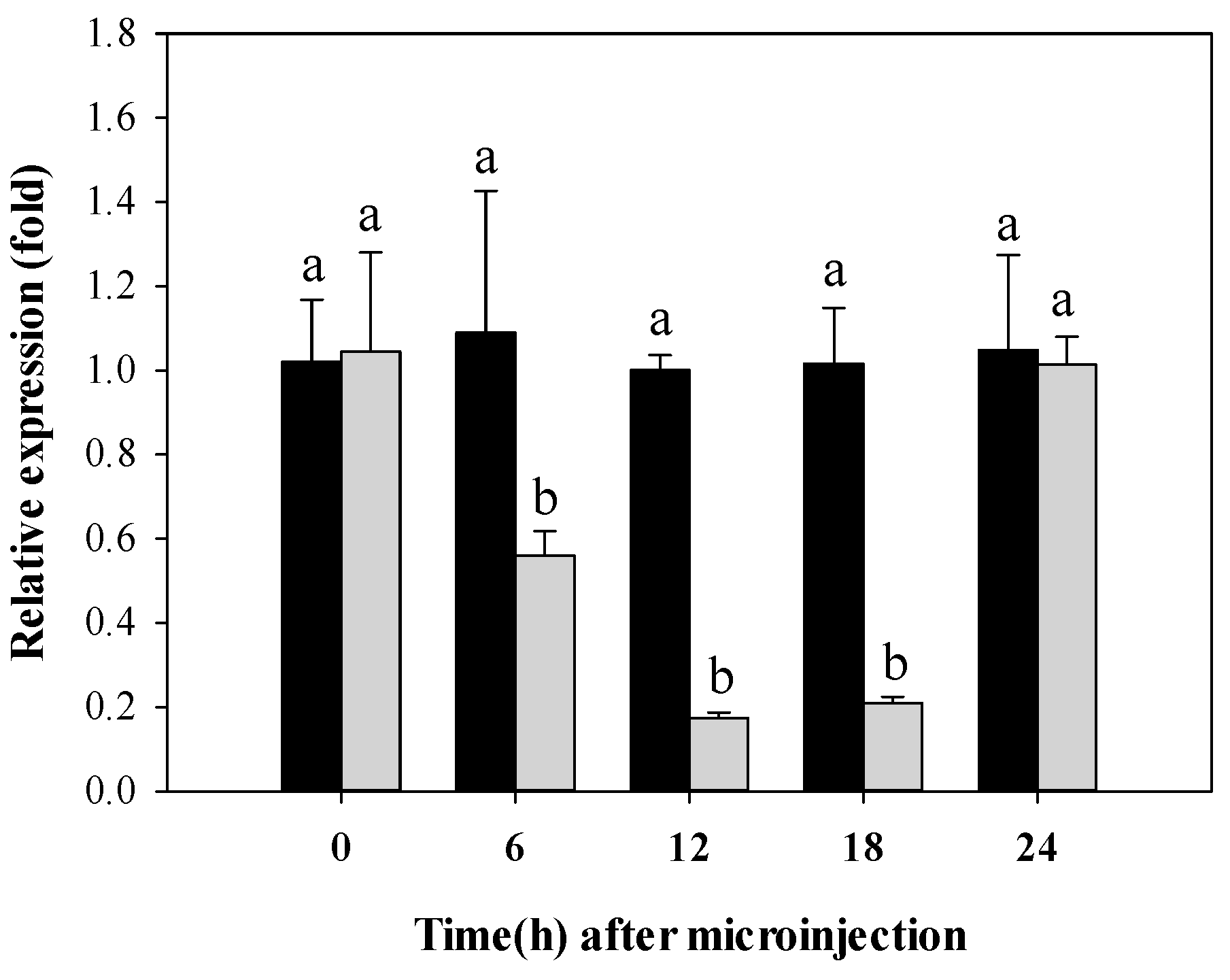
2.4. Expression of CYP340W1 after RNAi
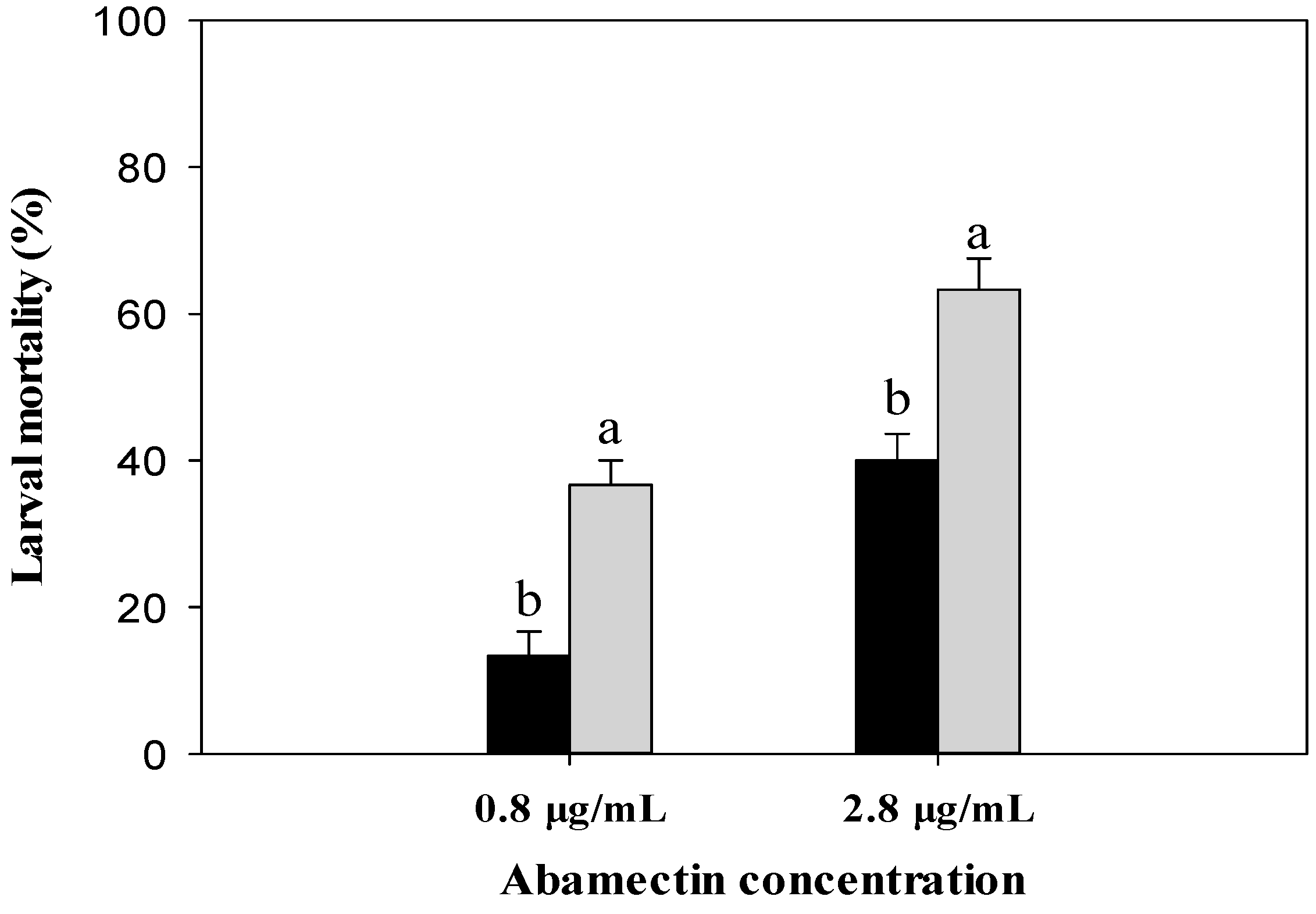
2.5. Effect of CYP340W1 Silencing on Larval Mortality
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Chemicals and Bioassay
4.3. Molecular Cloning of CYP340W1
4.4. Quantitative RT-PCR
4.5. dsRNA Synthesis
4.6. RNAi Assays
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Burg, R.W.; Miller, B.M.; Baker, E.E.; Birnbaum, J.; Currie, S.A.; Hartman, R.; Kong, Y.L.; Monaghan, R.L.; Olson, G.; Putter, I.; et al. Avermectins, new family of potent anthelmintic agents: Producing organism and fermentation. Antimicrob. Agents Chemother. 1979, 15, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Putter, I.; Mac Connell, J.G.; Preiser, F.A.; Haidri, A.A.; Ristich, S.S.; Dybas, R.A. Avermectins: Novel insecticides, acaricides and nematicides from a soil microorganism. Cell. Mol. Life Sci. 1981, 37, 963–964. [Google Scholar] [CrossRef]
- Iqbal, M.; Wright, D.J. Evaluation of resistance, cross-resistance and synergism of abamectin and teflubenzuron in a multi-resistant field population of Plutella xylostella (Lepidoptera: Plutellidae). Bull. Entomol. Res. 1997, 87, 481–486. [Google Scholar] [CrossRef]
- De Oliveira, A.C.; Abreu de Siqueira, H.A.; de Oliveira, J.V.; Da Silva, J.E.; Michereff Filho, M. Resistance of brazilian diamondback moth populations to insecticides. Sci. Agricola 2011, 68, 154–159. [Google Scholar]
- Attique, M.N.R.; Khaliq, A.; Sayyed, A.H. Could resistance to insecticides in Plutella xylostella (Lep., Plutellidae) be overcome by insecticide mixtures? J. Appl. Entomol. 2006, 130, 122–127. [Google Scholar] [CrossRef]
- Pu, X.; Yang, Y.; Wu, S.; Wu, Y. Characterisation of abamectin resistance in a field-evolved multiresistant population of Plutella xylostella. Pest Manag. Sci. 2010, 66, 371–378. [Google Scholar] [PubMed]
- Abro, G.H.; Syed, T.S.; Kalhoro, A.N.; Sheikh, G.H.; Awan, M.S.; Jessar, R.D.; Shelton, A.M. Insecticides for control of the diamondback moth, Plutella xylostella (L.)(Lepidoptera: Plutellidae) in Pakistan and factors that affect their toxicity. Crop Prot. 2013, 52, 91–96. [Google Scholar] [CrossRef]
- Lai, K.P.; Liao, P.F.; Mo, Y.B.; Fang, F.; Ye, Y.Q. Toxicity and field efficacy of abamectin 0.8% CS against main vegetable pests. Plant Prot. 2011, 37, 157–160. [Google Scholar]
- Liang, P.; Gao, X.W.; Zheng, B.Z. Genetic basis of resistance and studieson cross-resistance in a population of diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2003, 59, 1232–1236. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.J.; Zhang, W.J.; Zhang, Y.J.; Xu, B.Y.; Zhu, G.R. Cuticular penetration and desensitivity of GABA_ (A) receptor in abamectin resistant Plutella xylostella. L. Acta Entomol. Sin. 2002, 45, 336–340. [Google Scholar]
- Qian, L.; Cao, G.C.; Song, J.X.; Yin, Q.; Han, Z. Biochemical mechanisms conferring cross-resistance between tebufenozide and abamectin in Plutella xylostella. Pestic. Biochem. Physiol. 2008, 91, 175–179. [Google Scholar] [CrossRef]
- Huang, H.S.; Hu, N.T.; Yao, Y.E.; Wu, C.Y.; Chiang, S.W.; Sun, C.N. Molecular cloning and heterologous expression of a glutathione S-transferase involved in insecticide resistance from the diamondback moth, Plutella xylostella. Insect Biochem. Mol. Biol. 1998, 28, 651–658. [Google Scholar] [CrossRef]
- Zhou, X.M.; Wu, Q.J.; Zhang, Y.J.; Bai, L.Y.; Huang, X.Y. Cloning and characterization of a GABA receptor from Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2008, 101, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.X.; Yang, J.Q.; Hou, W.J.; Xu, B.Y.; Xie, W.; Wang, S.L.; Zhang, Y.J.; Zhou, X.G.; Wu, Q.J. Molecular cloning and characterization of a P-glycoprotein from the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Mol. Sci. 2013, 14, 22891–22905. [Google Scholar] [CrossRef] [PubMed]
- Eldefrawi, M.E.; Miskus, R.; Sutcher, V. Methylenedioxyphenyl derivatives as synergists for carbamate insecticides in susceptible, DDT-and parathion-resistant house flies. J. Econ. Entomol. 1960, 53, 231–234. [Google Scholar] [CrossRef]
- Demaeght, P.; Dermauw, W.; Tsakireli, D.; Khajehali, J.; Nauen, R.; Tirry, L.; Vontas, J.; Lümmen, P.; Leeuwen, T.V. Molecular analysis of resistance to acaricidal spirocyclic tetronic acids in Tetranychus urticae: CYP392E10 metabolizes spirodiclofen. Insect Biochem. Mol. Biol. 2013, 43, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.J.; Ma, C.X.; Li, M.; Sheng, C.F.; Liu, H.X.; Qiu, X.H. CYP9A12 and CYP9A17 in the cotton bollworm Helicoverp armigera: Sequence similarity, expression profile and xenobiotic response. Pest Manag. Sci. 2010, 66, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Karatolos, N.; Williamson, M.S.; Denholm, I.; Gorman, K.; Richard, H.; Ffrench-Constant, R.H.; Bass, C. Over-expression of a cytochrome P450 is associated with resistance to pyriproxyfen in the greenhouse whitefly Trialeurodes vaporariorum. PLoS ONE 2012, 7, e31077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, M.A.M.; Miyata, T.; Miura, K.; Tanaka, T. RNA interference-mediated knockdown of a cytochrome P450, CYP6BG1, from the diamondback moth, Plutella xylostella, reduces larval resistance to permethrin. Insect Biochem. Mol. Biol. 2009, 39, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Riga, M.; Tsakireli, D.; Ilias, A.; Morou, E.; Myridakis, A.; Stephanou, E.G.; Nauen, R.; Dermauw, W. Abamectin is metabolized by CYP392A16, a cytochrome P450 associated with high levels of acaricide resistance in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 46, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Bogwitz, M.R.; Chung, H.; Magoc, L.; Rigby, S.; Wong, W.; Keefe, M.O.; Mckenzie, J.A.; Batterham, P.; Daborn, P.J. Cyp12a4 confers lufenuron resistance in a natural population of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2005, 102, 12807–12812. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Yue, L.; Chen, S.; Wu, Y.D. Functional expression of Helicoverpa Armigera CYP9A12 and CYP9A14 in Saccharomyces cerevisiae. Pestic. Biochem. Physiol. 2008, 92, 101–105. [Google Scholar] [CrossRef]
- Yu, L.Y.; Tang, W.Q.; He, W.Y.; Ma, X.L.; Vasseur, L.; Baxter, S.W.; Yang, G.; Huang, S.G.; Song, F.Q.; You, M.S. Characterization and expression of the cytochrome P450 gene family in diamondback moth, Plutella xylostella (L.). Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.H.; Cheng, E.Y. Insecticide resistance in Plutella xylostella L. XI. Resistance to newly introduced insecticides in Taiwan (1990–2001). Agric. Res. China 2001, 50, 80–89. [Google Scholar]
- Santos, V.C.; de Siqueira, H.A.A.; da Silva, J.E.; de Farias, M.J.D.C. Insecticide resistance in populations of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), from the state of Pernambuco, Brazil. Neotrop. Entomol. 2011, 40, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Lin, Q.; Chen, H.; Li, Z.; Yin, F.; Feng, X. Identification of a novel cytochrome P450 gene, CYP321E1 from the diamondback moth, Plutella Xylostella (L.) and RNA interference to evaluate its role in chlorantraniliprole resistance. Bull. Entomol. Res. 2014, 104, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Puinean, A.M.; Foster, S.P.; Oliphant, L.; Denholm, L.; Field, L.M.; Millar, N.S.; Williamson, M.S.; Bass, C. Amplification of a cytochrome P450 gene is associated with resistance to neonicotinoid insecticides in the aphid Myzus persicae. PLoS Genet. 2010, 6, e1000999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carino, F.A.; Koener, J.F.; Plapp, F.W.; Feyereisen, R. Constitutive overexpression of the cytochrome P450 gene CYP6A1 in a house fly strain with metabolic resistance to insecticides. Insect Biochem. Mol. Biol. 1994, 24, 411–418. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Kulye, M.; Yang, F.S.; Xiao, L.; Zhang, Y.T.; Zeng, H.M.; Wang, J.H.; Liu, Z.X. Identification, characterization, and expression of a novel P450 gene encoding CYP6AE25 from the Asian corn borer, Ostrinia. furnacalis. J. Insect Sci. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect cytochrome P450. Compr. Mol. Insect Sci. 2005, 4, 1–77. [Google Scholar]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Snyder, M.J.; Stevens, J.L.; Andersen, J.F.; Feyereisen, R. Expression of cytochrome P450 genes of the CYP4 family in midgut and fat body of the tobacco hornworm, Manduca. sexta. Arch. Biochem. Biophys. 1995, 321, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Maıbeche-Coisne, M.; Monti-Dedieu, L.; Aragon, S.; Dauphin-Villemant, C. A new cytochrome P450 from Drosophila melanogaster, CYP4G15, expressed in the nervous system. Biochem. Biophys. Res. Commun. 2000, 273, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Andersen, E.C.; Shapiro, J.A.; Gerke, J.P.; Kruglyak, L. Natural variation in a chloride channel subunit confers avermectin resistance in C. elegans. Science 2012, 335, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Korytko, P.J.; Scott, J.G. CYP6D1 protects thoracic ganglia of houseflies from the neurotoxic insecticide cypermethrin. Arch. Insect Biochem. Physiol. 1998, 37, 57–63. [Google Scholar] [CrossRef]
- Zhu, F.; Parthasarathy, R.; Bai, H.; Woithe, K.; Kaussmann, M.; Nauen, R.; Harrison, D.A.; Palli, S.R. A brain-specific cytochrome P450 responsible for the majority of deltamethrin resistance in the QTC279 strain of Tribolium castaneum. Proc. Natl. Acad. Sci. USA 2010, 107, 8557–8562. [Google Scholar] [CrossRef] [PubMed]
- Festucci-Buselli, R.A.; Carvalho-Dias, A.S.; Oliveira-Andrade, D.; Caixeta-Nunes, C.; Li, H.M.; Stuart, J.J.; Muir, W.; Scharf, M.E.; Pittendrigh, B.R. Expression of Cyp6g1 and Cyp12d1 in DDT resistant and susceptible strains of Drosophila melanogaster. Insect Mol. Biol. 2005, 14, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Liu, N. Differential expression of CYP6A5 and CYP6A5v2 in pyrethroid-resistant house flies, Musca domestica. Arch. Insect Biochem. Physiol. 2008, 34, 147–161. [Google Scholar]
- Le Goff, G.; Hilliou, F.; Siegfried, B.D.; Boundy, S.; Wajnberg, E.; Sofer, L.; Audant, P.; Ffrench-constant, R.H.; Feyereisen, R. Xenobiotic response in Drosophila melanogaster: Sex dependence of P450 and GST gene induction. Insect Biochem. Mol. Biol. 2006, 36, 674–682. [Google Scholar] [CrossRef]
- Bautista, M.A.M.; Tanaka, T.; Miyata, T. Identification of permethrin-inducible cytochrome P450s from the diamondback moth, Plutella xylostella (L.) and the possibility of involvement in permethrin resistance. Pestic. Biochem. Physiol. 2007, 87, 85–93. [Google Scholar] [CrossRef]
- Liu, F.; Shi, X.Z.; Liang, Y.P.; Wu, Q.J.; Xu, B.Y.; Xie, W.; Wang, S.L.; Zhang, Y.J.; Liu, N.N. A 36-bp deletion in the alpha subunit of glutamate-gated chloride channel contributes to abamectin resistance in Plutella xylostella. Entomol. Exp. Appl. 2014, 153, 85–92. [Google Scholar] [CrossRef]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; Roy, N.V.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.J.; Kang, S.; Zhu, X.; Xia, J.X.; Wu, Q.J.; Wang, S.L.; Xie, W.; Zhang, Y.J. The novel ABC transporter ABCH1 is a potential target for RNAi-based insect pest control and resistance management. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.J.; Kang, S.; Chen, D.F.; Wu, Q.J.; Wang, S.L.; Xie, W.; Zhu, X.; Baxter, S.W.; Zhou, X.G.; Jurat-fuentes, J.L.; et al. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in damondback moth. PLoS Genet. 2015, 11, e1005124. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | LC50 (mg/L) (95% FL) | Slope ± SE | Χ2 (df) | RR a | LC10 (95% FL) (mg/L) | LC20 (95% FL) (mg/L) |
|---|---|---|---|---|---|---|
| ABM-S | 0.024 (0.020–0.028) | 1.862 ± 0.335 | 2.856 (3) | 1 | 0.004 (0.001–0.009) | 0.008 (0.013–0.015) |
| ABM-R | 2.822 (2.013–4.920) | 1.485 ± 0.257 | 5.529 (3) | 114.96 | 0.478 (0.136–0.802) | 0.879 (0.399–1.285) |
| Experiment | Primer Name | Primer Sequence (5’-3’) |
|---|---|---|
| 3’RACE | CYP-RACE3’ | CGGCTACGACACGTCTTCCACCACT |
| 5’RACE | CYP-RACE5’ | GAACCGCTGCCGGGGGATGCTGAAC |
| Full-length confirmation | CYP-Full-F | CATCAATGTCTGTATCATTGCTTCT |
| CYP-Full-R | TCTAGAAACATATTAATTAACAGGC | |
| qRT-PCR | CYP340-QF | GTTTTCGGCAACTCGGACAG |
| CYP340-QR | TGGGGATGGTCACGTTTTTC | |
| Reference gene | EF1-F | GCCTCCCTACAGCGAATC |
| EF1-R | CCTTGAACCAGGGCATCT | |
| RPL32-F | CCAATTTACCGCCCTACC | |
| RPL32-R | TACCCTGTTGTCAATACCTCT |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Yang, J.; Xu, B.; Xie, W.; Wang, S.; Zhang, Y.; Yang, F.; Wu, Q. Identification and Characterization of the Gene CYP340W1 from Plutella xylostella and Its Possible Involvement in Resistance to Abamectin. Int. J. Mol. Sci. 2016, 17, 274. https://doi.org/10.3390/ijms17030274
Gao X, Yang J, Xu B, Xie W, Wang S, Zhang Y, Yang F, Wu Q. Identification and Characterization of the Gene CYP340W1 from Plutella xylostella and Its Possible Involvement in Resistance to Abamectin. International Journal of Molecular Sciences. 2016; 17(3):274. https://doi.org/10.3390/ijms17030274
Chicago/Turabian StyleGao, Xue, Jiaqiang Yang, Baoyun Xu, Wen Xie, Shaoli Wang, Youjun Zhang, Fengshan Yang, and Qingjun Wu. 2016. "Identification and Characterization of the Gene CYP340W1 from Plutella xylostella and Its Possible Involvement in Resistance to Abamectin" International Journal of Molecular Sciences 17, no. 3: 274. https://doi.org/10.3390/ijms17030274