
The Stable Level of Glutamine synthetase 2 Plays an Important Role in Rice Growth and in Carbon-Nitrogen Metabolic Balance


Abstract
:1. Introduction
2. Results
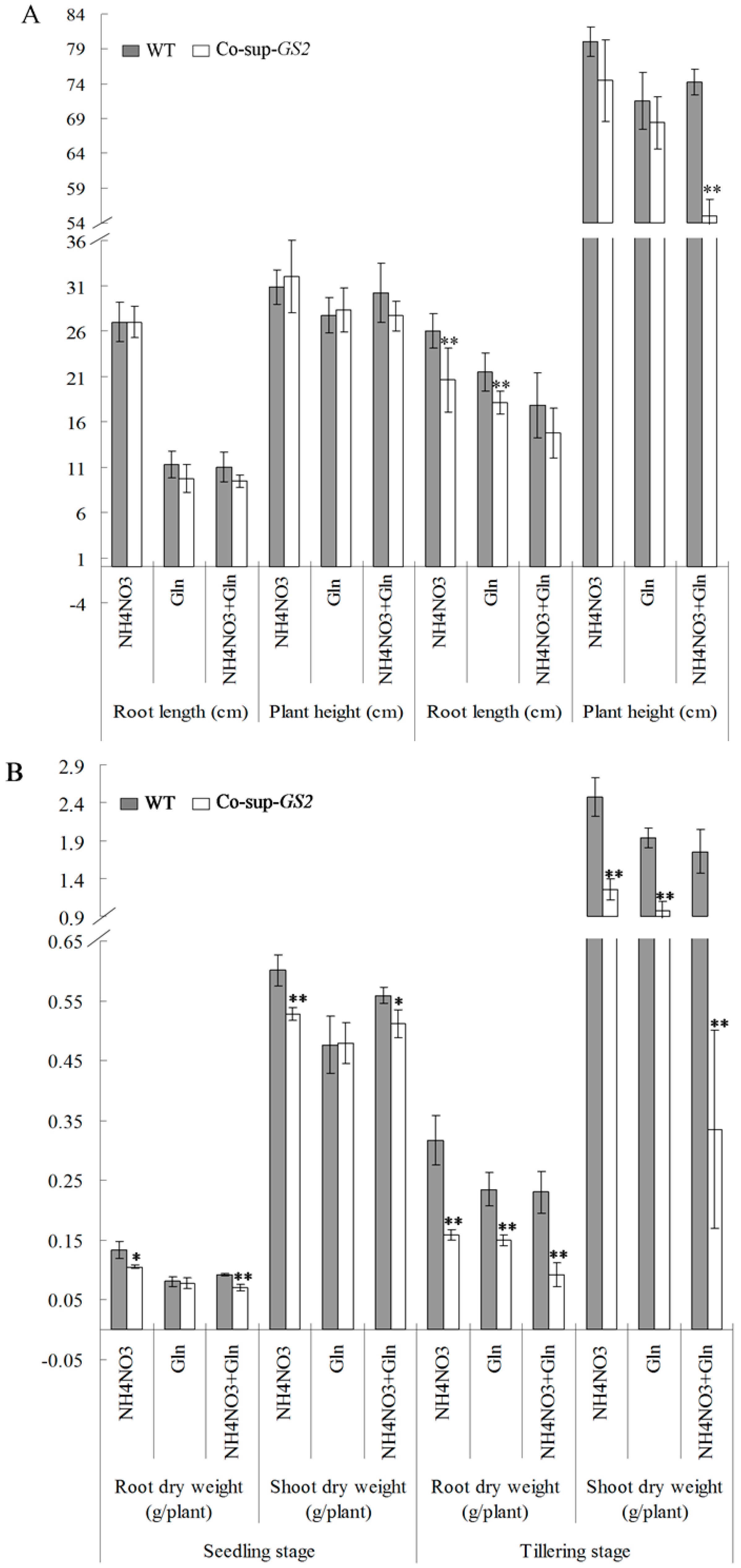
2.1. Growth Phenotype of GS2-Cosuppressed Plants at the Seedling and Tillering Stages


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | SPAD at Tillering Stage | Photosynthesis Parameters at Tillering Stage | |||
|---|---|---|---|---|---|
| Photosynthetic Rate (μmol CO2 m−2·s−2) | Stomatal Conductance (mmol m−2·s−1) | Intercellular CO2 Concentration (μL·L−1) | Transpiration Rate (mmol H2O m−2·S−1) | ||
| NH4NO3 | |||||
| WT | 46.1 ± 2.9 | 28.24 ± 3.32 | 0.82 ± 0.06 | 287.75 ± 9.62 | 15.06 ± 1.10 |
| Co-sup-GS2 | 38.3 ± 6.3 ** | 29.76 ± 3.24 | 1.01 ± 0.26 | 292.91 ± 8.36 | 15.99 ± 1.36 |
| Gln | |||||
| WT | 46.0 ± 2.2 | 33.24 ± 2.10 | 0.90 ± 0.12 | 278.39 ± 7.79 | 16.36 ± 1.56 |
| Co-sup-GS2 | 41.1 ± 3.2 ** | 28.39 ± 1.54 ** | 0.71 ± 0.05 | 277.77 ± 1.32 | 12.99 ± 0.61 ** |
| NH4NO3 + Gln | |||||
| WT | 44.5 ± 2.4 | 30.83 ± 2.03 | 0.76 ± 0.10 | 274.75 ± 9.98 | 14.70 ± 1.61 |
| Co-sup-GS2 | 42.3 ± 2.0 * | 29.53 ± 2.36 | 0.64 ± 0.02 | 268.03 ± 5.93 | 13.32 ± 0.74 |
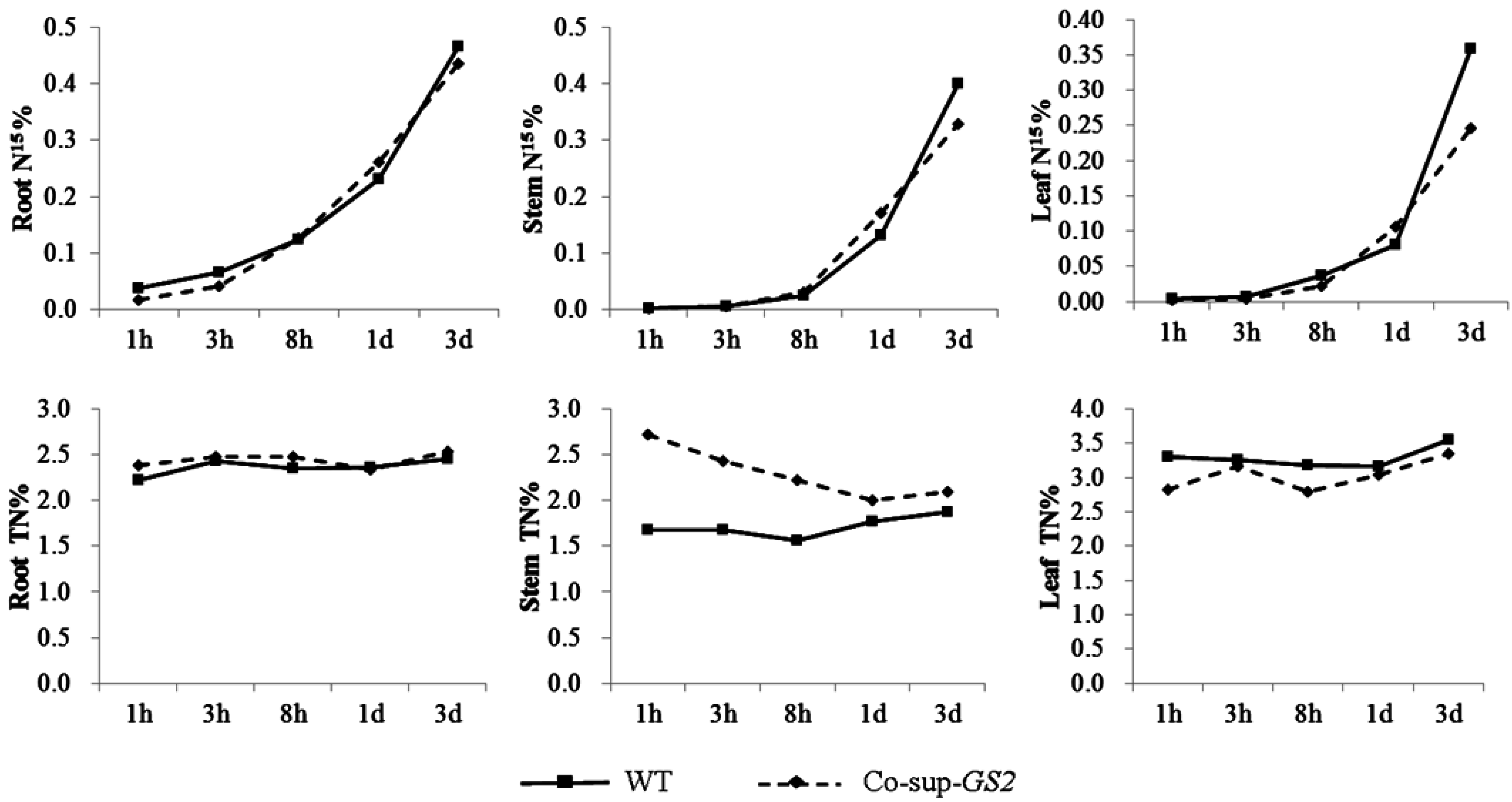
2.2. Nitrogen Uptake by GS2-Cosuppressed Plants

| Treatment | C% | N% | C/N | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Leaf | Root | Stem | Leaf | |
| NH4NO3 | |||||||||
| WT | 38.64 ± 0.23 | 35.02 ± 0.52 | 38.81 ± 0.39 | 2.81 ± 0.13 | 2.50 ± 0.09 | 3.19 ± 0.05 | 13.75 ± 0.59 | 14.00 ± 0.41 | 12.16 ± 0.10 |
| Co-sup-GS2 | 38.49 ± 0.49 | 35.11 ± 0.47 | 38.97 ± 0.80 | 3.16 ± 0.10 | 3.15 ± 0.15 ** | 3.07 ± 0.17 | 12.18 ± 0.53 | 11.17 ± 0.69 ** | 12.69 ± 0.41 |
| Gln | |||||||||
| WT | 41.18 ± 0.16 | 36.59 ± 0.30 | 39.78 ± 0.35 | 3.29 ± 0.20 | 2.84 ± 0.22 | 3.41 ± 0.10 | 12.53 ± 0.79 | 12.92 ± 0.86 | 11.68 ± 0.24 |
| Co-sup-GS2 | 42.41 ± 0.30 | 37.00 ± 0.66 | 40.31 ± 0.73 | 3.21 ± 0.07 | 3.77 ± 0.07 ** | 3.23 ± 0.10 | 13.20 ± 0.40 | 9.82 ± 0.10 * | 12.48 ± 0.23 |
| NH4NO3 + Gln | |||||||||
| WT | 41.92 ± ND | 38.67 ± 0.36 | 42.64 ± 0.26 | 3.86 ± ND | 3.70 ± 0.07 | 4.16 ± 0.06 | 10.86 ± ND | 10.45 ± 0.17 | 10.25 ± 0.08 |
| Co-sup-GS2 | ND | 37.54 ± 0.57 | 40.67 ± 1.14 | ND | 4.37 ± 0.13 ** | 3.75 ± 0.17 | ND | 8.58 ± 0.13 ** | 10.84 ± 0.19 |
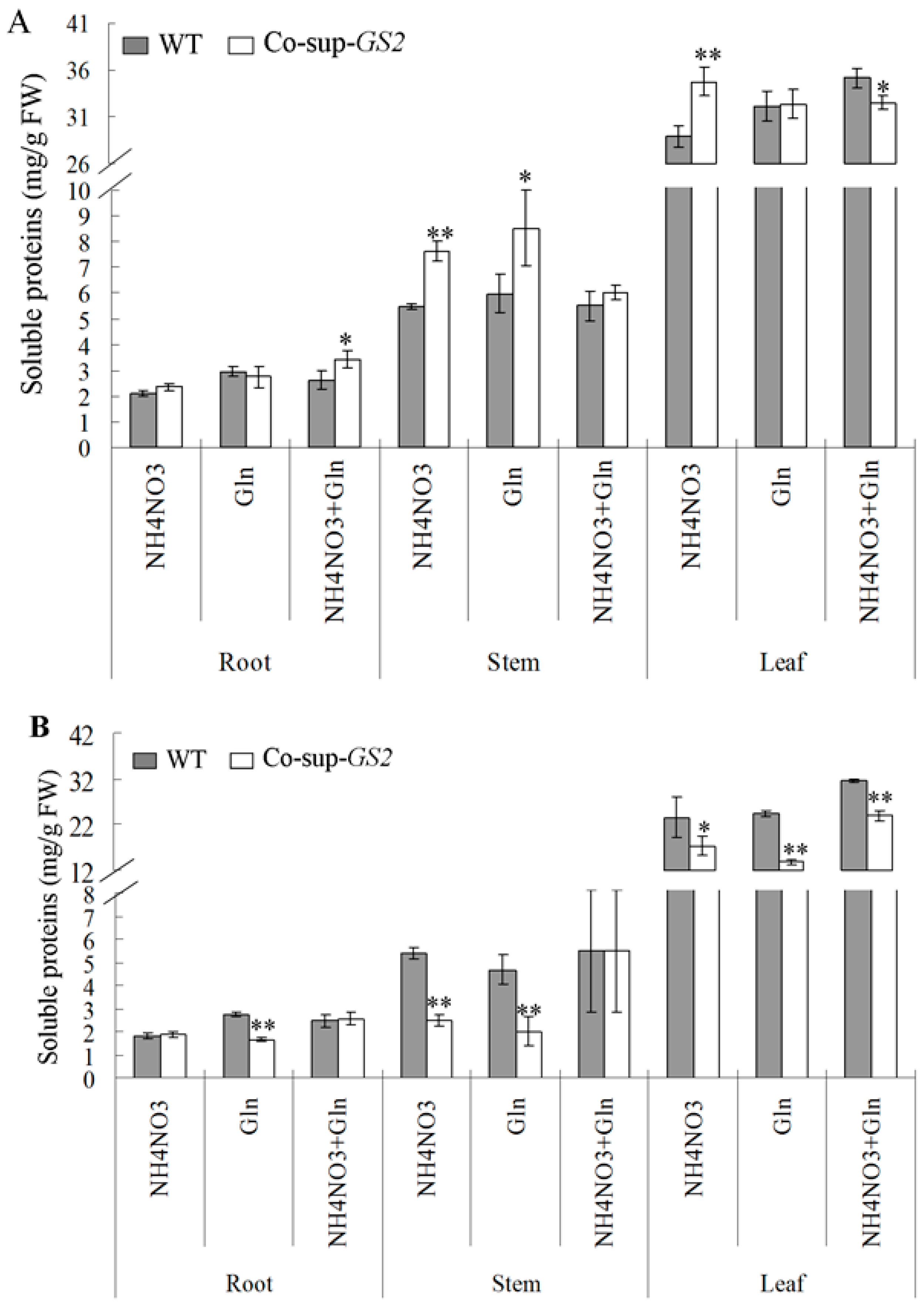
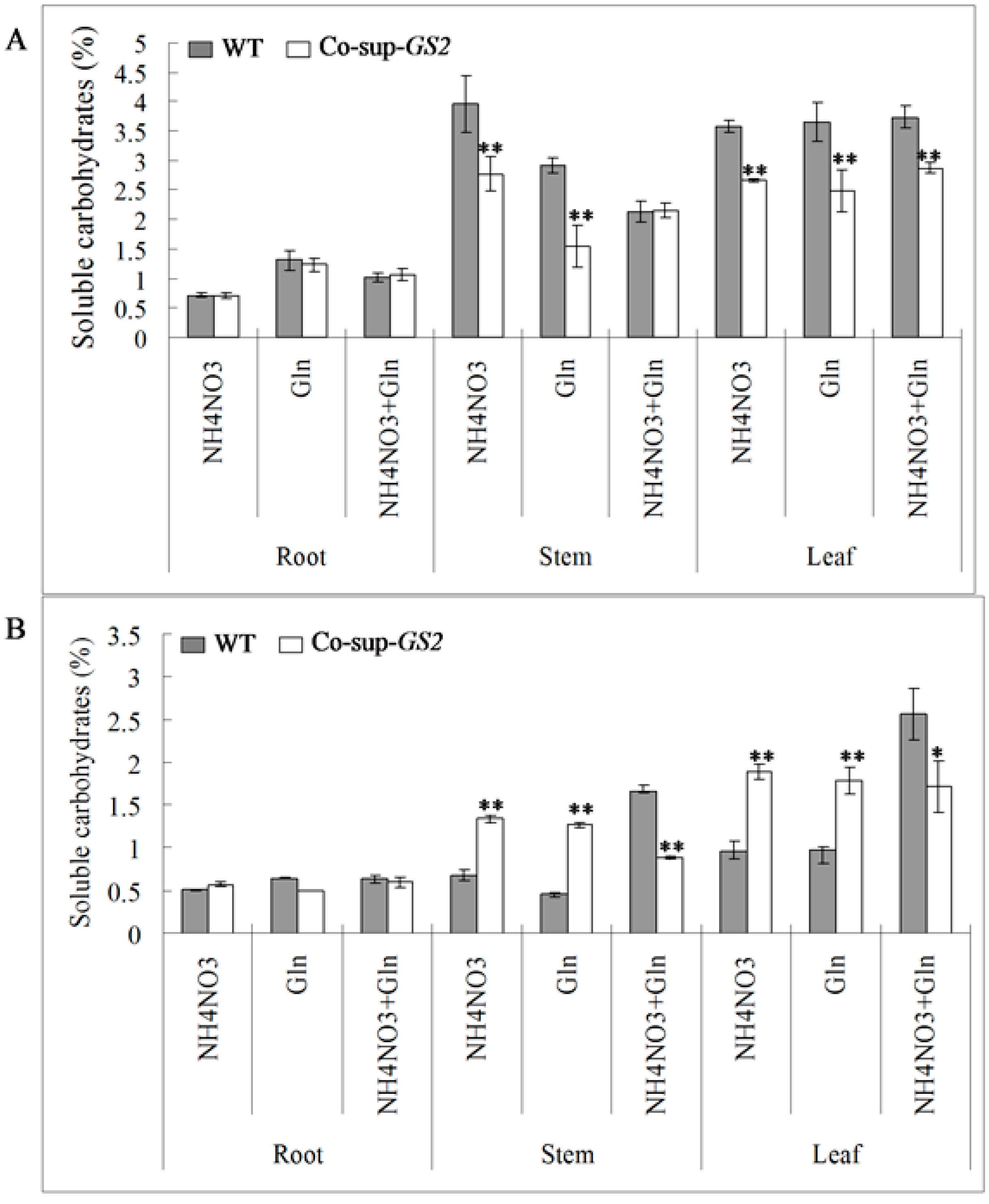
2.3. Soluble Proteins and Carbohydrates Determination in the GS2-Cosuppressed Plants


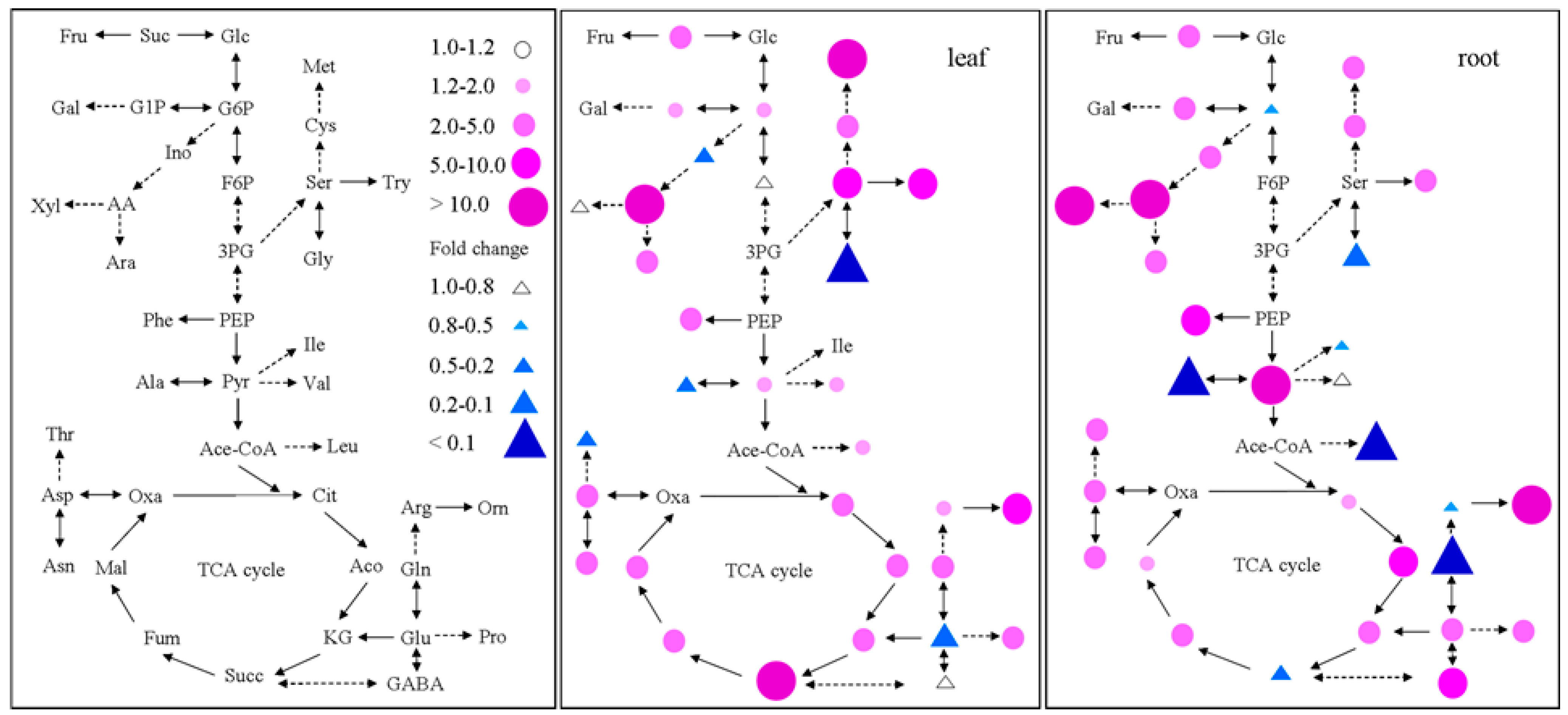
2.4. Metabolite Profiling Analysis in GS2-Cosuppressed Plants

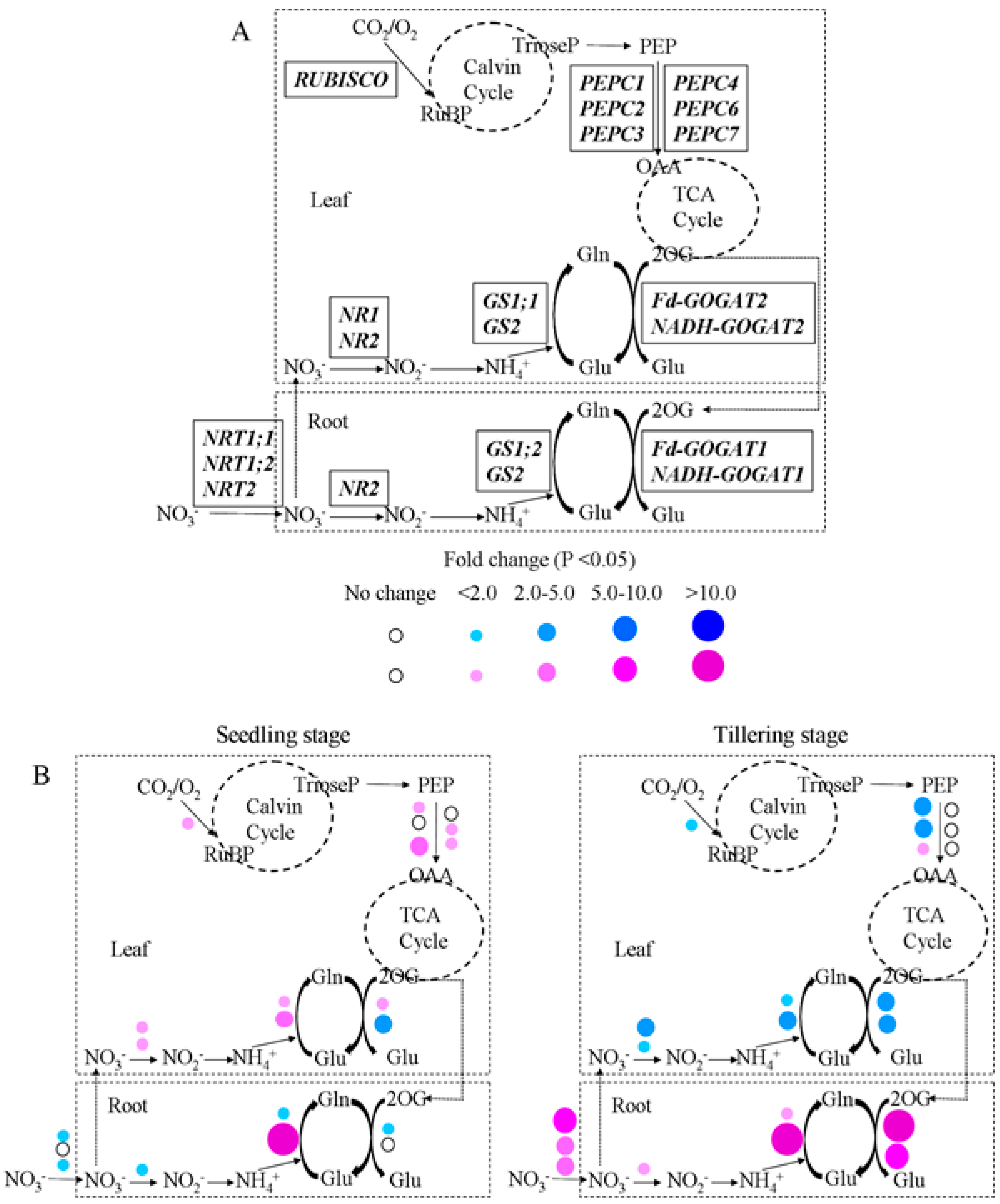
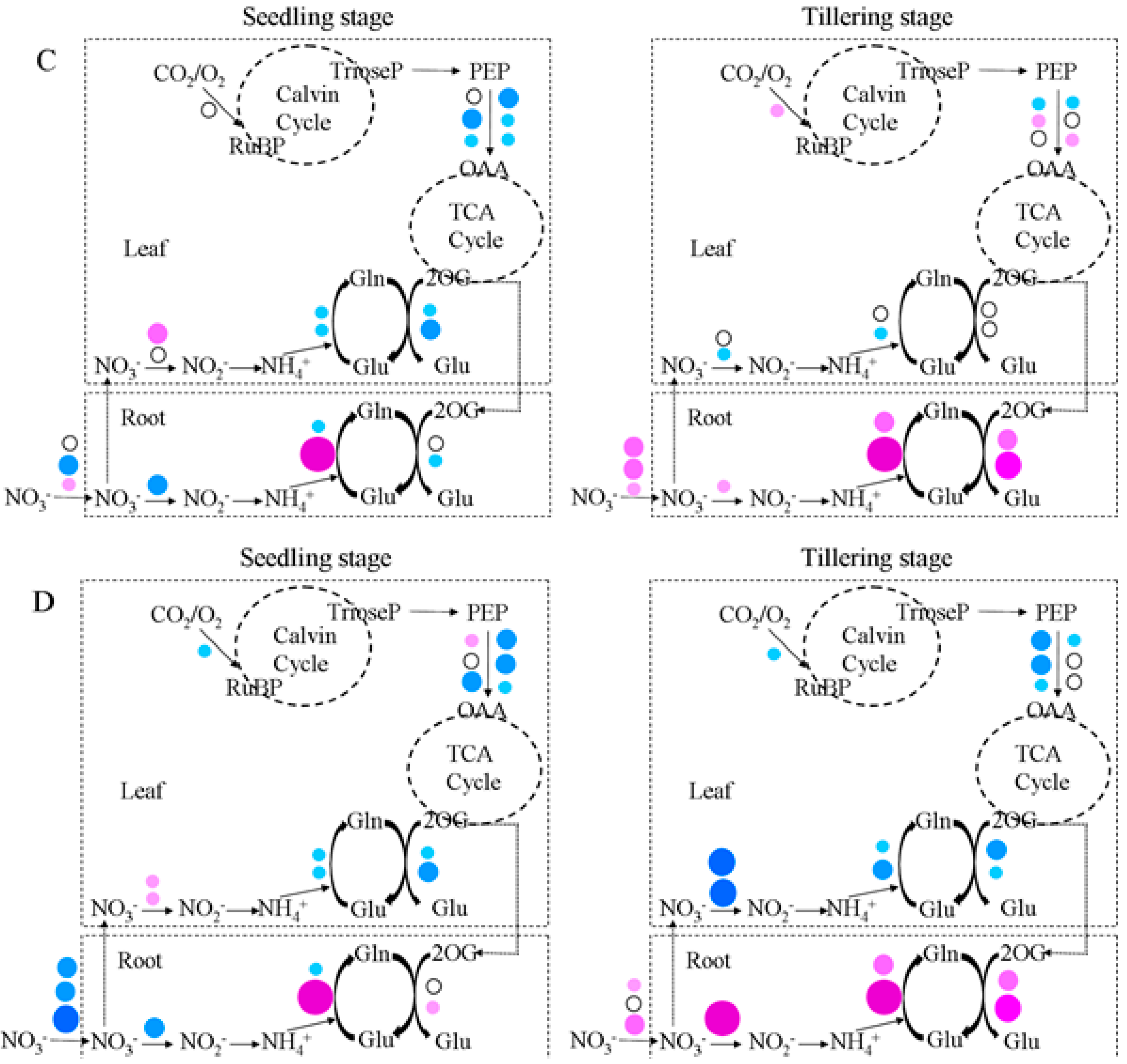
2.5. Gene Expression Analysis in GS2-Cosuppressed Plants


3. Discussion
3.1. The Carbon-Nitrogen Metabolic Imbalance Is One of the Possible Reasons for the Chlorosis Phenotype in the GS2-Cosuppressed Plants
3.2. The Incoordination between the Root and Leaf Tissues Is Another Possible Reason for the Chlorosis Phenotype in GS2-Cosuppressed Plants
3.3. The Impact of Gln on Plant Growth and Carbon-Nitrogen Metabolism
4. Experimental Section
4.1. Plant Materials and Growth Conditions
4.2. Nitrogen Uptake Assay
4.3. Determination of the Leaf SPAD Value and Photosynthetic Parameters
4.4. Determination of the Physiological Parameters
4.5. Gene Expression Analysis
4.6. Statistical Analysis
5. Conclusions

Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Frink, C.R.; Waggoner, P.E.; Ausubel, J.H. Nitrogen fertilizer: Retrospect and prospect. Proc. Natl. Acad. Sci. USA 1999, 96, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Forde, B.G. Molecular and developmental biology of inorganic nitrogen nutrition. In The Arabidopsis Book; Meyerowitz, E., Somerville, C., Eds.; American Society of Plant Biologists: Rockville, MD, USA, 2002. [Google Scholar]
- Zheng, Z. Carbon and nitrogen nutrient balance signaling in plants. Plant Signal. Behav. 2009, 4, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A.; Ferrario-Méry, S.; Touraine, B. Nitrogen and signalling. In Advances in Photosynthesis; Foyer, C., Noctor, G., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2002; Volume 12, pp. 205–225. [Google Scholar]
- Krapp, A.; Truong, H.N. Regulation of C/N interaction in model plant species. In The Efficiency of Nitrogen Utilization in Plants; Goyal, S., Tischner, R., Basra, A., Eds.; Enhancing Haworth Press: New York, NY, USA, 2005; pp. 127–173. [Google Scholar]
- Coruzzi, G.M.; Zhou, L. Carbon and nitrogen sensing and signaling in plants: Emerging “matrix effects”. Curr. Opin. Plant Biol. 2001, 4, 247–253. [Google Scholar] [CrossRef]
- Martin, T.; Oswald, O.; Graham, I.A. Arabidopsis seedling growth, storage lipid mobilization and photosynthetic gene expression are regulated by carbon: nitrogen availability. Plant Physiol. 2002, 128, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef] [PubMed]
- Miflin, B.J.; Lea, P.J. Ammonia assimilation. In The Biochemistry of Plants; Miflin, B.J., Ed.; Academic Press: New York, NY, USA, 1980; Volume 5, pp. 169–202. [Google Scholar]
- Hirel, B.; Lea, P.J. Ammonium assimilation. In Plant Nitrogen; Lea, P.J., Morof Gaudry, J.F., Eds.; Springer-Verlag: Berlin, Germany, 2001; pp. 79–99. [Google Scholar]
- Tobin, A.K.; Yamaya, T. Cellular compartmentation of ammonium assimilation in rice and barley. J. Exp. Bot. 2001, 53, 591–604. [Google Scholar] [CrossRef]
- Ireland, R.J.; Lea, P.J. The enzymes of glutamine, glutamate, asparagines and aspirate metabolism. In Plant Amino Acids: Biochemistry and Biotechnology; Singh, B.K., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 49–109. [Google Scholar]
- Mann, A.F.; Fenten, P.A.; Stewart, G.R. Identification of two forms of glutamine synthetase in barley (Hordeum vulgare L.). Biochem. Biophys. Res. Commun. 1979, 88, 515–521. [Google Scholar] [CrossRef]
- Hirel, B.; Gadal, P. Glutamine synthetase in rice: A comparative study of the enzymes from roots and leaves. Plant Physiol. 1980, 66, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Kamachi, K.; Oikawa, M.; Ojima, K.; Yamaya, T. Response of glutamine synthetase and glutamate synthase isoforms to nitrogen sources in rice cell cultures. Plant Cell Physiol. 1990, 31, 1071–1077. [Google Scholar]
- Kamachi, K.; Yamaya, T.; Maie, T.; Ojima, K. A role for glutamine synthetase in the remobilization of leaf nitrogen during natural senescence in rice leaves. Plant Physiol. 1991, 96, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Zozaya-Garza, M.; Sengupta-Gopalan, C. Glutamine synthetase gene isolation from an alfalfa leaf cDNA library. Plant Physiol. 1999, 119, 1568. [Google Scholar]
- Ishiyama, K.; Inoue, E.; Tabuchi, M.; Yamaya, T.; Takahashi, H. Biochemical background and compartmentalized functions of cytosolic glutamine synthetase for active ammonium assimilation in rice roots. Plant Cell Physiol. 2004, 45, 1640–1647. [Google Scholar] [CrossRef] [PubMed]
- Bernard, S.M.; Moller, A.L.; Dionisio, G.; Kichey, T.; Jahn, T.P.; Dubois, F.; Baudo, M.; Lopes, M.S.; Tercé-Laforgue, T.; Foyer, C.H.; et al. Gene expression, cellular localisation and function of glutamine synthetase isozymes in wheat (Triticum aestivum L.). Plant Mol. Biol. 2008, 67, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.-W.; Zhang, Y.-D.; Chen, Y.; Wang, S.; Tang, D.-M.; Huang, D.-F. Isolation and characterization of a GS2 gene in melon (Cucumis melo L.) and its expression patterns under the fertilization of different forms of N. Mol. Biotechnol. 2010, 44, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, A.; Nigro, D.; Giancaspro, A.; Blanco, A. The glutamine synthetase (GS2) gene is in relation to grain protein content of durum wheat. Funct. Integr. Genomics 2011, 11, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Seabra, A.; Vieira, C.P.; Cullimore, J.V.; Garvalho, H.G. Medicago truncatula contains a second gene encoding a plastid located glutamine synthetase exclusively expressed in developing seeds. BMC Plant Biol. 2010, 10, 183. [Google Scholar] [PubMed]
- Tabuchi, M.; Sugiyama, K.; Ishiyama, K.; Inoue, E.; Sato, T.; Takahashi, H.; Yamaya, T. Severe reduction in growth rate and grain filling of rice mutants lacking OsGS1;1, a cytosolic glutamine synthetase1;1. Plant J. 2005, 42, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, I.C.; Brears, T.; Knight, T.J.; Clark, A.; Coruzzi, G.M. Overexpression of cytosolic glutamine synthetase: Relation to nitrogen, light and photorespiration. Plant Physiol. 2002, 129, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Hoshida, H.; Tanaka, Y.; Hibino, T.; Hayashi, Y.; Tanaka, A.; Takabe, T.; Takabe, T. Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase. Plant Mol. Biol. 2000, 43, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Husted, S.; Mattsson, M.; Möllers, C.; Wallbraun, M.; Schjoerring, J.K. Photorespiratory NH4+ production in leaves of wild-type and glutamine synthetase 2 antisense oilseed rape. Plant Physiol. 2002, 130, 989–998. [Google Scholar] [CrossRef] [PubMed]
- García-Calderón, M.; Chiurazzi, M.; Espuny, M.R.; Márquez, A.J. Photorespiratory metabolism and nodule function: Behavior of Lotus japonicas mutants deficient in plastid glutamine synthetase. Mol. Plant. Microbe Interact. 2012, 25, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, R.D.; Murray, A.J.S.; Lea, P.J. Inhibition of photosynthesis in barley with decreased levels of chloroplastic glutamine synthetase activity. J. Exp. Bot. 1987, 38, 1799–1809. [Google Scholar] [CrossRef]
- Blackwell, R.D.; Murray, A.J.S.; Lea, P.J.; Joy, K.W. Photorespiratory amino donors, sucrose synthesis and the induction of CO2 fixation in barley deficient in glutamine synthetase and/or glutamate synthase. J. Exp. Bot. 1988, 39, 845–858. [Google Scholar] [CrossRef]
- Wallsgrove, R.M.; Turner, J.C.; Hall, N.P.; Kendall, A.C.; Bright, S.W.J. Barley mutants lacking chloroplast glutamine synthetase: Biochemical and genetic analysis. Plant Physiol. 1987, 83, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Socolow, R. Nitrogen management and the future of food: Lessons from the management of energy and carbon. Proc. Natl. Acad. Sci. USA 1999, 96, 6001–6008. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F.S. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.F. Strategies for developing green super rice. Proc. Natl. Acad. Sci. USA 2007, 104, 16402–16409. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Xiao, J.; Zhang, Q.; Lian, X. Co-suppressed glutamine synthetase 2 gene modifies nitrogen metabolism and plant growth in rice. Chin. Sci. Bull. 2010, 55, 823–833. [Google Scholar] [CrossRef]
- Gutiérrez, R.A.; Lejay, L.; Dean, A.; Chiaromonte, F.; Shasha, D.E.; Coruzzi, G.M. Qualitative network models and genome wide expression data define carbon/nitrogen-responsive molecular machines in Arabidopsis. Genome Biol. 2007, 8. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Gutiérrez, R.A. A systems view of nitrogen nutrient and metabolite responses in Arabidopsis. Curr. Opin. Plant Biol. 2008, 11, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Pant, B.D.; Musialak-Lange, M.; Nuc, P.; May, P.; Buhtz, A.; Kehr, J.; Walther, D.; Scheible, W.R. Identification of nutrient responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time PCR profiling and small RNA sequencing. Plant Physiol. 2009, 150, 1541–1555. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Araus, V.; Lu, C.; Parry, G.; Green, P.J.; Coruzzi, G.M.; Gutiérrez, R.A. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 4477–4482. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, U.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Weber, A.P.; Zellerhoff, N.; Bucher, M.; Fahnenstich, H.; Sonnewald, U. Adaptation of maize source leaf metabolism to stress related disturbances in carbon, nitrogen and phosphorus balance. BMC Genomics 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Bao, A.; Zhao, Z.; Ding, G.; Shi, L.; Xu, F.; Cai, H. Accumulated expression level of cytosolic glutamine synthetase 1 gene (OsGS1;1 or OsGS1;2) alter plant development and the carbon-nitrogen metabolic status in rice. PLoS ONE 2014, 9, e95581. [Google Scholar] [CrossRef] [PubMed]
- Kusano, M.; Tabuchi, M.; Fukushima, A.; Funayama, K.; Diaz, C.; Kobayashi, M.; Hayashi, N.; Tsuchiya, Y.N.; Takahashi, H.; Kamata, A.; et al. Metabolomics data reveal a crucial role of cytosolic glutamine synthetase 1;1 in coordinating metabolic balance in rice. Plant J. 2011, 66, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Delgado, C.M.; García-Calderón, M.; Sánchez, D.H.; Udvardi, M.K.; Kopka, J.; Márquez, A.J.; Betti, M. Transcriptomic and metabolic changes associated with photorespiratory ammonium accumulation in the model legume Lotus japonicas. Plant Physiol. 2013, 162, 1834–1848. [Google Scholar] [CrossRef] [PubMed]
- Näsholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Kielland, K. Amino-acid-absorption by arctic plants-implications for plant nutrition and nitrogen cycling. Ecology 1994, 75, 2373–2383. [Google Scholar] [CrossRef]
- Lipson, D.; Näsholm, T. The unexpected versatility of plants: Organic nitrogen use and availability in terrestrial ecosystems. Oecologia 2001, 128, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Forsum, O.; Svennerstam, H.; Ganeteg, U.; Nasholm, T. Capacities and constraints of amino acid utilization in Arabidopsis. New Phytol. 2008, 179, 1058–1069. [Google Scholar] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd Ed. ed; Academic Press Limited: San Diego, CA, USA, 1997. [Google Scholar]
- Cambui, C.A.; Svennerstam, H.; Gruffman, L.; Nordin, A.; Ganeteg, U.; Nasholm, T. Patterns of plant biomass partitioning depend on nitrogen source. PLoS ONE 2011, 6, e19211. [Google Scholar] [CrossRef] [PubMed]
- Thornton, B.; Robinson, D. Uptake and assimilation of nitrogen from solutions containing multiple sources. Plant Cell Environ. 2005, 28, 813–821. [Google Scholar] [CrossRef]
- Wang, H.-J.; Wu, L.-H.; Wang, M.-Y.; Zhu, Y.-H.; Tao, Q.-N.; Zhang, F.-S. Effects of amino acids replacing nitrate on growth, nitrate accumulation, and macroelement concentrations in pak-choi (Brassica chinensis L.). Pedosphere 2007, 17, 595–600. [Google Scholar] [CrossRef]
- Henry, H.A.L.; Jefferies, R.L. Free amino acid, ammonium and nitrate concentrations in soil solutions of a grazed coastal marsh in relation to plant growth. Plant Cell Environ. 2002, 25, 665–675. [Google Scholar] [CrossRef]
- Bollard, E.G. A comparative study of the ability of organic nitrogenous compounds to serve as sole nitrogen sources of nitrogen for the growth of plants. Plant Soil 1966, 25, 153–166. [Google Scholar] [CrossRef]
- Yoshida, S.; Forno, D.A.; Cook, J.H.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice, 3rd ed.; International Rice Research Institute: Manila, Philippine, 1976. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Morris, D.L. Quantitative determination of carbohydrates with Dreywood’s anthrone reagent. Science 1948, 107, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Maness, N. Extraction and analysis of soluble carbohydrates. Methods Mol. Biol. 2010, 639, 341–370. [Google Scholar] [PubMed]
- Kusano, M.; Fukushima, A.; Arita, M.; Jonsson, P.; Moritz, T.; Kobayashi, M.; Hayashi, N.; Tohge, T.; Saito, K. Unbiased characterization of genotype-dependent metabolic regulations by metabolomic approach in Arabidopsis thaliana. BMC Syst. Biol. 2007, 1. [Google Scholar] [CrossRef] [PubMed]
- Kusano, M.; Fukushima, A.; Kobayashi, M.; Hayashi, N.; Jonsson, P.; Moritz, T.; Ebana, K.; Saito, K. Application of a metabolomic method combining one-dimensional and two-dimensional gas chromatographytime-of-flight/mass spectrometry to metabolic phenotyping of natural variants in rice. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 855, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Redestig, H.; Fukushima, A.; Stenlund, H.; Moritz, T.; Arita, M.; Saito, K.; Kusano, M. Compensation for systematic cross-contribution improves normalization of mass spectrometry based metabolomics data. Anal. Chem. 2009, 81, 7974–7980. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, A.; Zhao, Z.; Ding, G.; Shi, L.; Xu, F.; Cai, H. The Stable Level of Glutamine synthetase 2 Plays an Important Role in Rice Growth and in Carbon-Nitrogen Metabolic Balance. Int. J. Mol. Sci. 2015, 16, 12713-12736. https://doi.org/10.3390/ijms160612713
Bao A, Zhao Z, Ding G, Shi L, Xu F, Cai H. The Stable Level of Glutamine synthetase 2 Plays an Important Role in Rice Growth and in Carbon-Nitrogen Metabolic Balance. International Journal of Molecular Sciences. 2015; 16(6):12713-12736. https://doi.org/10.3390/ijms160612713
Chicago/Turabian StyleBao, Aili, Zhuqing Zhao, Guangda Ding, Lei Shi, Fangsen Xu, and Hongmei Cai. 2015. "The Stable Level of Glutamine synthetase 2 Plays an Important Role in Rice Growth and in Carbon-Nitrogen Metabolic Balance" International Journal of Molecular Sciences 16, no. 6: 12713-12736. https://doi.org/10.3390/ijms160612713