Protective Effect of Resveratrol against IL-1β-Induced Inflammatory Response on Human Osteoarthritic Chondrocytes Partly via the TLR4/MyD88/NF-κB Signaling Pathway: An “in Vitro Study”

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
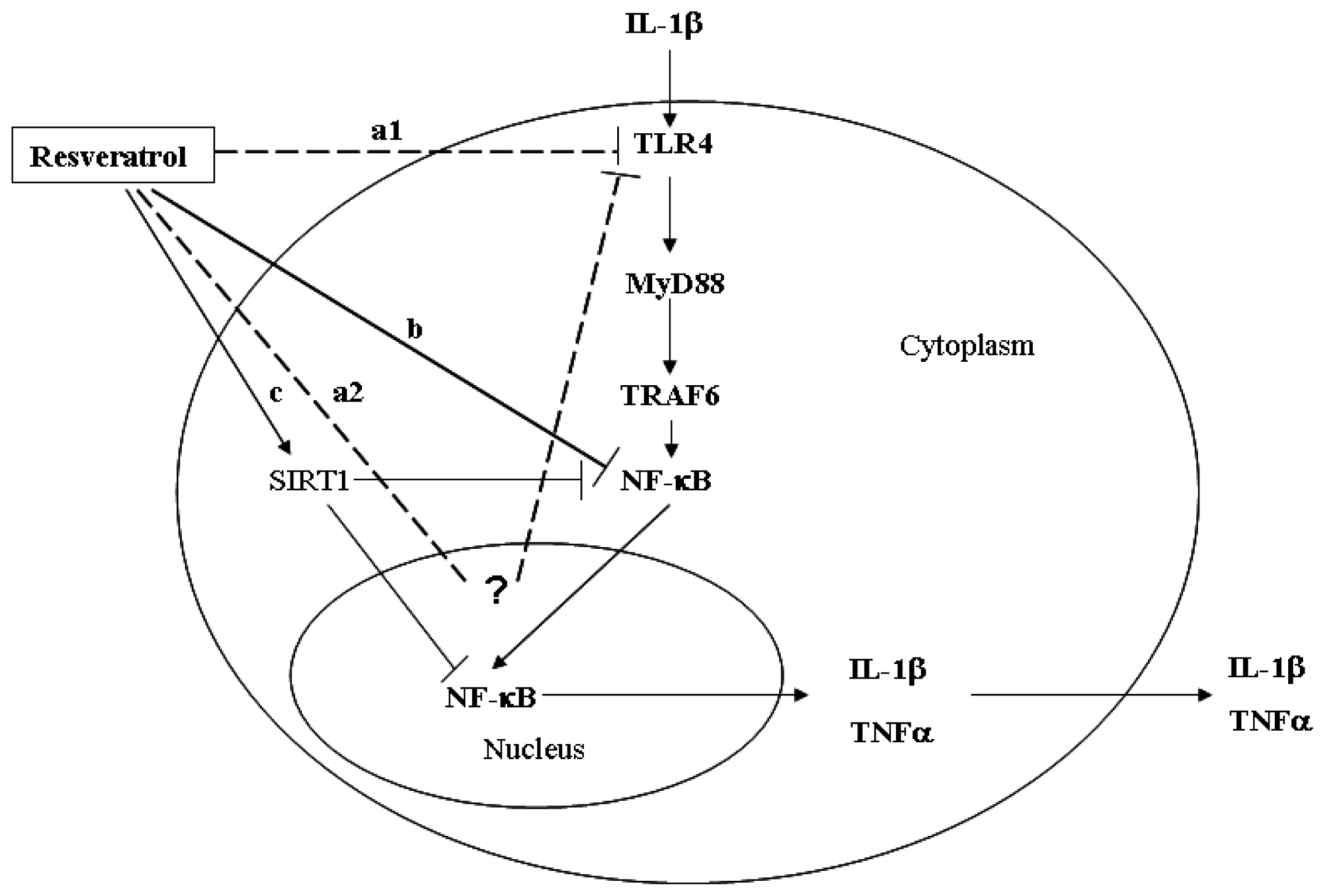
2. Results and Discussion
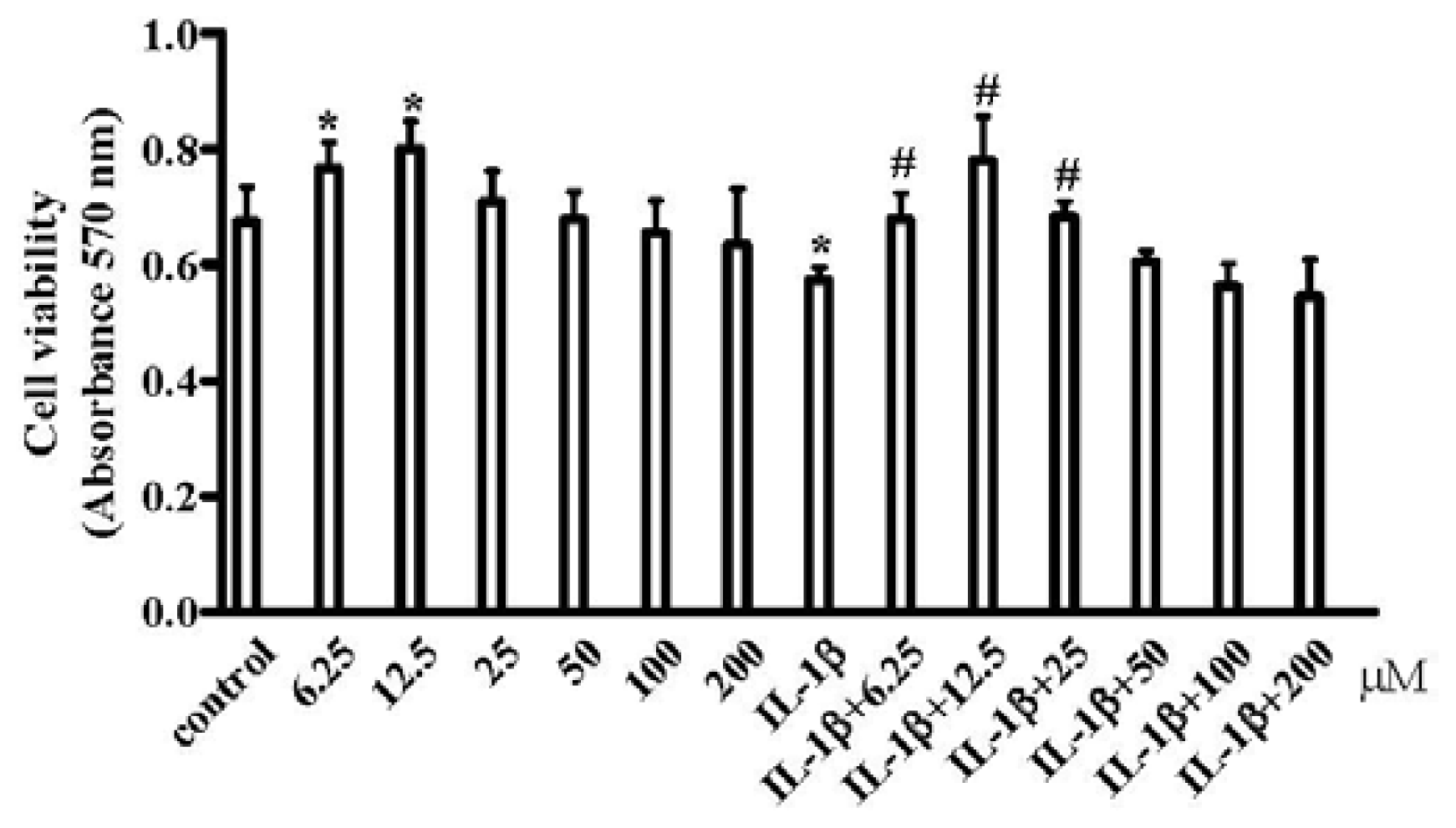
2.1. Resveratrol Improved the Inhibition of IL-1β-Induced Chondrocyte Proliferation
2.2. Resveratrol Suppressed IL-1β-Induced TLR4 Expression and TNFα Production
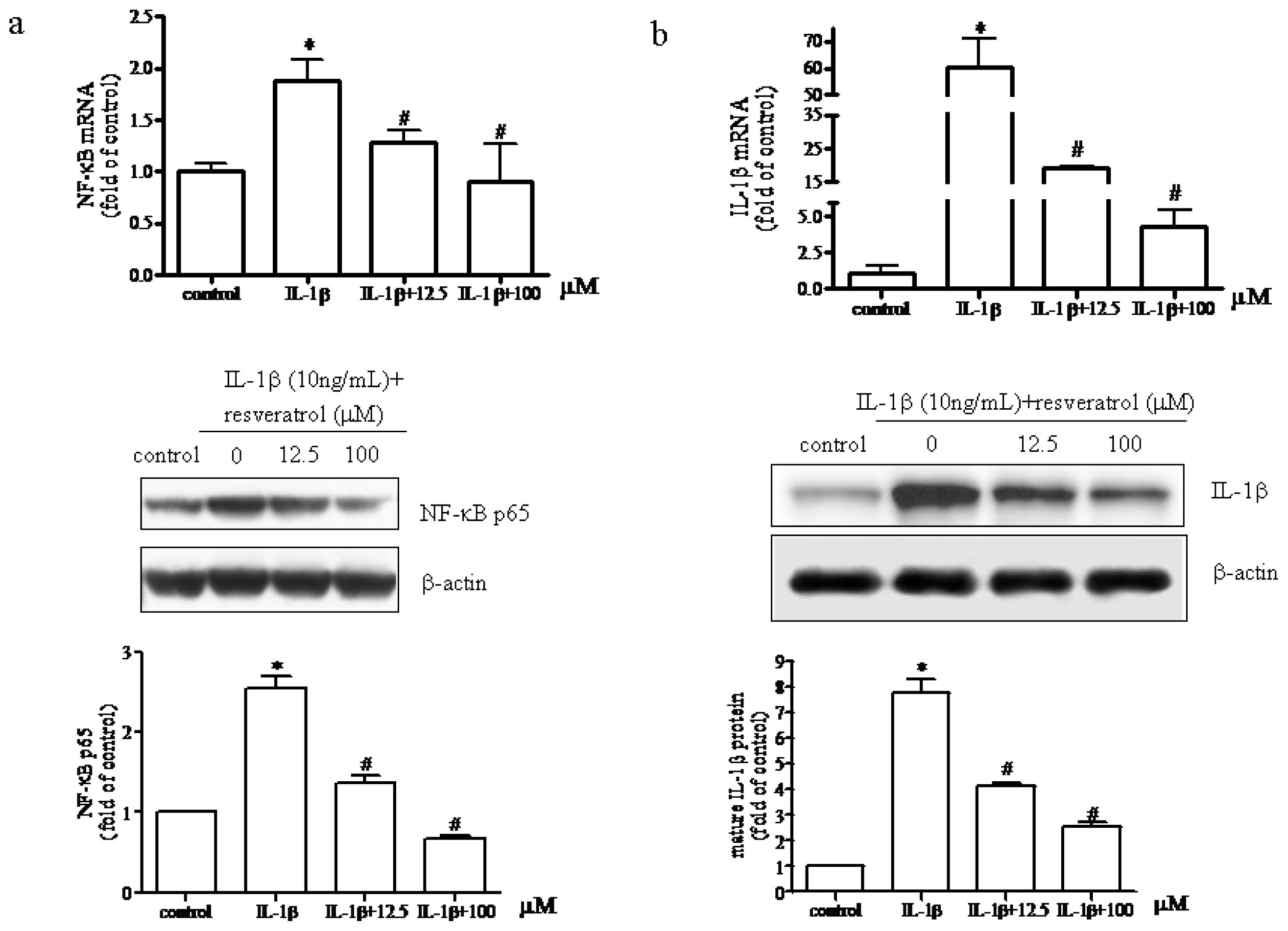
2.3. Resveratrol Suppressed Activation of the TLR4/NF-κB Signaling Pathway and Subsequent Synthesis of IL-1β in IL-1β-Stimulated Human Chondrocytes
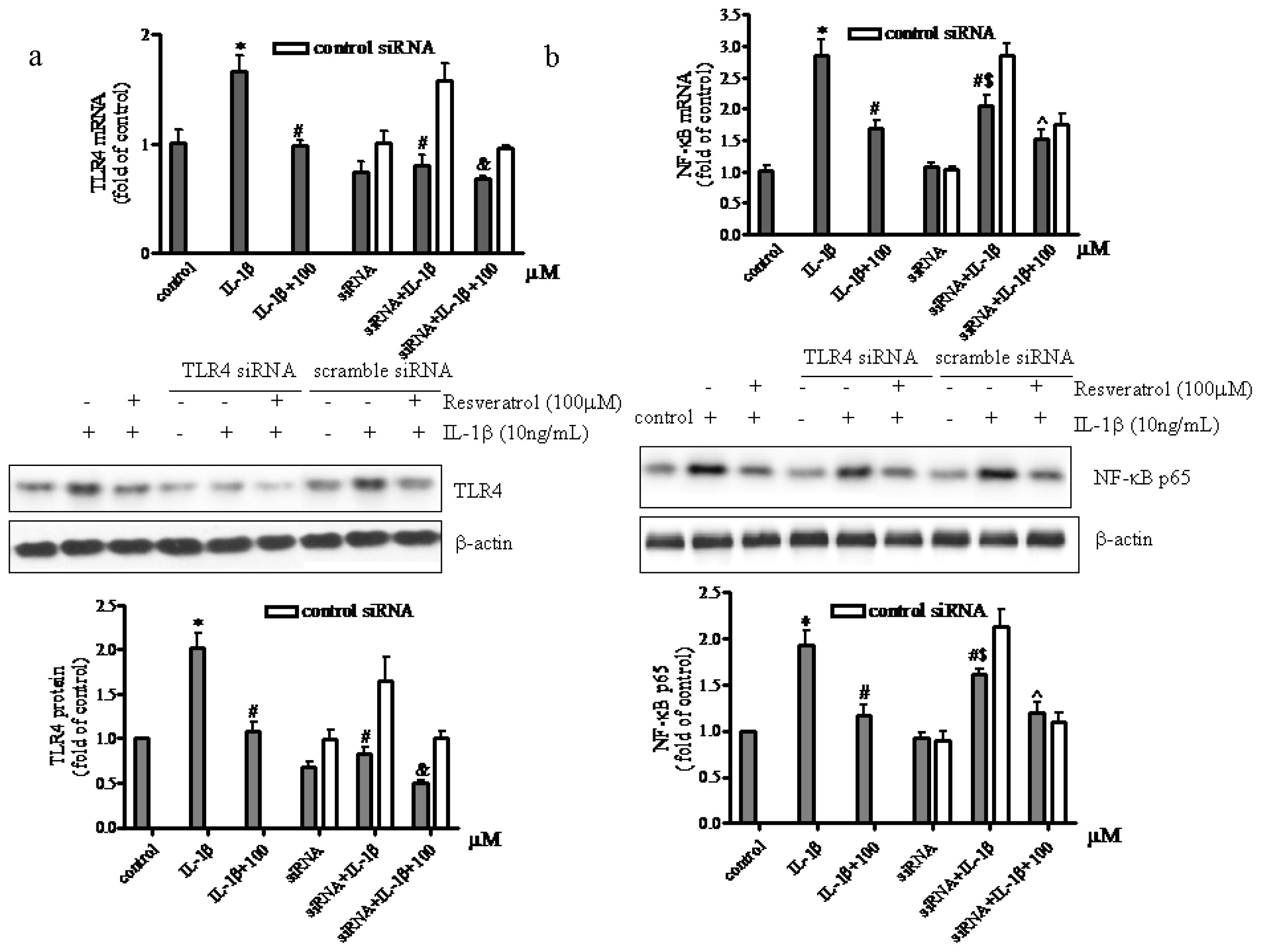
2.4. TLR4-Knockdown Increased the Effects of Resveratrol on IL-1β-Stimulated Human Chondrocytes
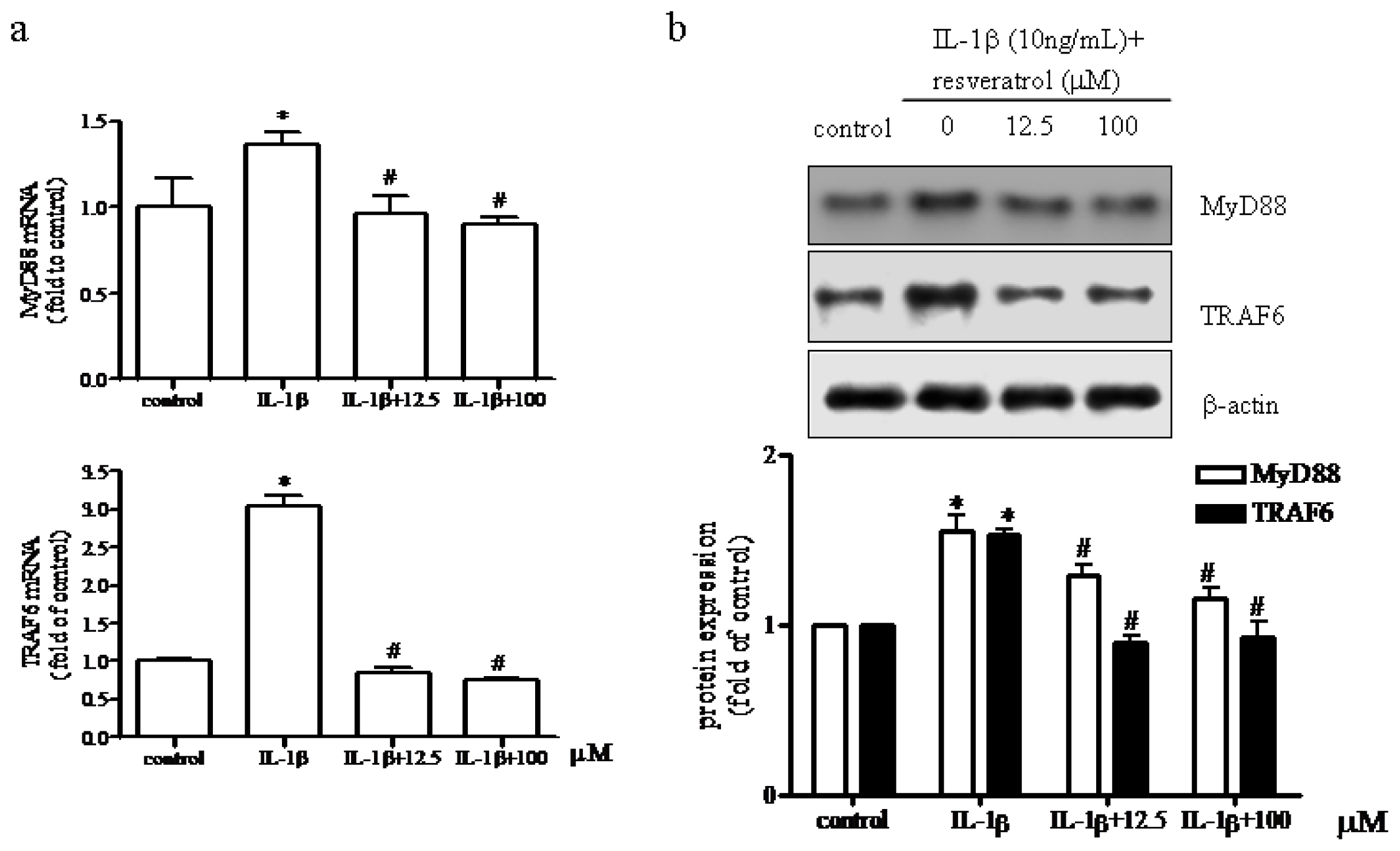
2.5. Effect of Resveratrol on MyD88 and TRAF6 Expression in IL-1β-Stimulated Human Chondrocytes
3. Experimental Section
3.1. Reagents and Antibodies
3.2. Chondrocyte Isolation and Culture
3.3. Cell Viability Assay
3.4. Chondrocyte Treatment
3.5. TLR4 siRNA Treatment
3.6. RNA Extraction and Reverse Transcription Polymerase Chain Reaction (RT-PCR)
3.7. Protein Extraction and Immunoblot Analysis
3.8. ELISA Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsAll authors were involved in the conception and design of the study, and all authors approved the final version to be published. L.L., H.G, H.L., Y.J., K.L. and Y.Z. conducted the experiments. L.L., L.A. and J.Y. analyzed the data. Manuscript was drafted by L.L and edited by H.G.
References
- Busija, L.; Bridgett, L.; Williams, S.R.; Osborne, R.H.; Buchbinder, R.; March, L.; Fransen, M. Osteoarthritis. Best Pract. Res. Clin. Rheumatol 2010, 24, 757–768. [Google Scholar]
- Campo, G.M.; Avenoso, A.; Campo, S.; D’Ascola, A.; Nastasi, G.; Calatroni, A. Molecular size hyaluronan differently modulates toll-like receptor-4 in LPS-induced inflammation in mouse chondrocytes. Biochimie 2010, 92, 204–215. [Google Scholar]
- Chiu, Y.C.; Yang, R.S.; Hsieh, K.H.; Fong, Y.C.; Way, T.D.; Lee, T.S.; Wu, H.C.; Fu, W.M.; Tang, C.H. Stromal cell-derived factor-1 induces matrix metalloprotease-13 expression in human chondrocytes. Mol. Pharmacol 2007, 72, 695–703. [Google Scholar]
- Umlauf, D.; Frank, S.; Pap, T.; Bertrand, J. Cartilage biology, pathology, and repair. Cell Mol. Life Sci 2010, 67, 4197–4211. [Google Scholar]
- Sofat, N.; Ejindu, V.; Kiely, P. What makes osteoarthritis painful? The evidence for local and central pain processing. Rheumatology (Oxford) 2011, 50, 2157–2165. [Google Scholar]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol 2011, 23, 471–478. [Google Scholar]
- Man, G.; Mologhianu, G. Osteoarthritis pathogenesis—A complex process that involves the entire joint. J. Med. Life 2014, 7, 37–41. [Google Scholar]
- Henrotin, Y.; Lambert, C.; Couchourel, D.; Ripoll, C.; Chiotelli, E. Nutraceuticals: Do they represent a new era in the management of osteoarthritis?—A narrative review from the lessons taken with five products. Osteoarthr. Cartil 2011, 19, 1–21. [Google Scholar]
- Sgaglione, N.A. Biologic approaches to articular cartilage surgery: Future trends. Orthop. Clin. N. Am 2005, 36, 485–495. [Google Scholar]
- Henrotin, Y.; Kurz, B.; Aigner, T. Oxygen and reactive oxygen species in cartilage degradation: Friends or foes? Osteoarthr. Cartil 2005, 13, 643–654. [Google Scholar]
- Henrotin, Y.; Kurz, B. Antioxidant to treat osteoarthritis: Dream or reality? Curr. Drug Targets 2007, 8, 347–357. [Google Scholar]
- Musumeci, G.; Trovato, F.M.; Pichler, K.; Weinberg, A.M.; Loreto, C.; Castrogiovanni, P. Extra-virgin olive oil diet and mild physical activity prevent cartilage degeneration in an osteoarthritis model: An in vivo and in vitro study on lubricin expression. J. Nutr. Biochem 2013, 24, 2064–2075. [Google Scholar]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol 2003, 21, 335–376. [Google Scholar]
- Miyake, K. Innate immune sensing of pathogens and danger signals by cell surface Toll-like receptors. Semin. Immunol 2007, 19, 3–10. [Google Scholar]
- Bobacz, K.; Sunk, I.G.; Hofstaetter, J.G.; Amoyo, L.; Toma, C.D.; Akira, S.; Weichhart, T.; Saemann, M.; Smolen, J.S. Toll-like receptors and chondrocytes: The lipopolysaccharide-induced decrease in cartilage matrix synthesis is dependent on the presence of toll-like receptor 4 and antagonized by bone morphogenetic protein 7. Arthritis Rheumatol 2007, 56, 1880–1893. [Google Scholar]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol 1999, 162, 3749–3752. [Google Scholar]
- Loniewski, K.J.; Patial, S.; Parameswaran, N. Sensitivity of TLR4- and -7-induced NF kappa B1 p105-TPL2-ERK pathway to TNF-receptor-associated-factor-6 revealed by RNAi in mouse macrophages. Mol. Immunol 2007, 44, 3715–3723. [Google Scholar]
- Abdollahi-Roodsaz, S.; Joosten, L.A.; Roelofs, M.F.; Radstake, T.R.; Matera, G.; Popa, C.; van der Meer, J.W.; Netea, M.G.; van den Berg, W.B. Inhibition of Toll-like receptor 4 breaks the inflammatory loop in autoimmune destructive arthritis. Arthritis Rheumatol 2007, 56, 2957–2967. [Google Scholar]
- Schelbergen, R.F.; Blom, A.B.; van den Bosch, M.H.; Sloetjes, A.; Abdollahi-Roodsaz, S.; Schreurs, B.W.; Mort, J.S.; Vogl, T.; Roth, J.; van den Berg, W.B.; et al. Alarmins S100A8 and S100A9 elicit a catabolic effect in human osteoarthritic chondrocytes that is dependent on Toll-like receptor 4. Arthritis Rheumatol 2012, 64, 1477–1487. [Google Scholar]
- Kundu, J.K.; Shin, Y.K.; Kim, S.H.; Surh, Y.J. Resveratrol inhibits phorbol ester-induced expression of COX-2 and activation of NF-kappaB in mouse skin by blocking IkappaB kinase activity. Carcinogenesis 2006, 27, 1465–1474. [Google Scholar]
- Dave, M.; Attur, M.; Palmer, G.; Al-Mussawir, H.E.; Kennish, L.; Patel, J.; Abramson, S.B. The antioxidant resveratrol protects against chondrocyte apoptosis via effects on mitochondrial polarization and ATP production. Arthritis Rheumatol 2008, 58, 2786–2797. [Google Scholar]
- Csaki, C.; Mobasheri, A.; Shakibaei, M. Synergistic chondroprotective effects of curcumin and resveratrol in human articular chondrocytes: Inhibition of IL-1beta-induced NF-kappaB-mediated inflammation and apoptosis. Arthritis Res. Ther 2009, 11, R165. [Google Scholar]
- Shakibaei, M.; Csaki, C.; Nebrich, S.; Mobasheri, A. Resveratrol suppresses interleukin-1beta-induced inflammatory signaling and apoptosis in human articular chondrocytes: Potential for use as a novel nutraceutical for the treatment of osteoarthritis. Biochem. Pharmacol 2008, 76, 1426–1439. [Google Scholar]
- Eo, S.H.; Cho, H.; Kim, S.J. Resveratrol inhibits nitric oxide-induced apoptosis via the NF-kappa B pathway in rabbit articular chondrocytes. Biomol. Ther. (Seoul) 2013, 21, 364–370. [Google Scholar]
- Csaki, C.; Keshishzadeh, N.; Fischer, K.; Shakibaei, M. Regulation of inflammation signalling by resveratrol in human chondrocytes in vitro. Biochem. Pharmacol. 2008, 75, 677–687. [Google Scholar]
- Shakibaei, M.; John, T.; Seifarth, C.; Mobasheri, A. Resveratrol inhibits IL-1 beta-induced stimulation of caspase-3 and cleavage of PARP in human articular chondrocytes in vitro. Ann. N. Y. Acad. Sci. 2007, 1095, 554–563. [Google Scholar]
- Elmali, N.; Esenkaya, I.; Harma, A.; Ertem, K.; Turkoz, Y.; Mizrak, B. Effect of resveratrol in experimental osteoarthritis in rabbits. Inflamm. Res 2005, 54, 158–162. [Google Scholar]
- Elmali, N.; Baysal, O.; Harma, A.; Esenkaya, I.; Mizrak, B. Effects of resveratrol in inflammatory arthritis. Inflammation 2007, 30, 1–6. [Google Scholar]
- Im, H.J.; Li, X.; Chen, D.; Yan, D.; Kim, J.; Ellman, M.B.; Stein, G.S.; Cole, B.; Kc, R.; Cs-Szabo, G.; et al. Biological effects of the plant-derived polyphenol resveratrol in human articular cartilage and chondrosarcoma cells. J. Cell Physiol 2012, 227, 3488–3497. [Google Scholar]
- Manna, S.K.; Mukhopadhyay, A.; Aggarwal, B.B. Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-kappa B, activator protein-1, and apoptosis: potential role of reactive oxygen intermediates and lipid peroxidation. J. Immunol 2000, 164, 6509–6519. [Google Scholar]
- Lei, M.; Wang, J.G.; Xiao, D.M.; Fan, M.; Wang, D.P.; Xiong, J.Y.; Chen, Y.; Ding, Y.; Liu, S.L. Resveratrol inhibits interleukin 1beta-mediated inducible nitric oxide synthase expression in articular chondrocytes by activating SIRT1 and thereby suppressing nuclear factor-kappaB activity. Eur. J. Pharmacol 2012, 674, 73–79. [Google Scholar]
- Youn, H.S.; Lee, J.Y.; Fitzgerald, K.A.; Young, H.A.; Akira, S.; Hwang, D.H. Specific inhibition of MyD88-independent signaling pathways of TLR3 and TLR4 by resveratrol: Molecular targets are TBK1 and RIP1 in TRIF complex. J. Immunol 2005, 175, 3339–3346. [Google Scholar]
- Zhang, C.; Lin, G.; Wan, W.; Li, X.; Zeng, B.; Yang, B.; Huang, C. Resveratrol, a polyphenol phytoalexin, protects cardiomyocytes against anoxia/reoxygenation injury via the TLR4/NF-kappaB signaling pathway. Int. J. Mol. Med 2012, 29, 557–563. [Google Scholar]
- Blanco, F.J.; Guitian, R.; Moreno, J.; de Toro, F.J.; Galdo, F. Effect of antiinflammatory drugs on COX-1 and COX-2 activity in human articular chondrocytes. J. Rheumatol 1999, 26, 1366–1373. [Google Scholar]
- Hashimoto, S.; Ochs, R.L.; Komiya, S.; Lotz, M. Linkage of chondrocyte apoptosis and cartilage degradation in human osteoarthritis. Arthritis Rheumatol 1998, 41, 1632–1638. [Google Scholar]
- Heraud, F.; Heraud, A.; Harmand, M.F. Apoptosis in normal and osteoarthritic human articular cartilage. Ann. Rheum. Dis 2000, 59, 959–965. [Google Scholar]
- Aigner, T.; Kim, H.A. Apoptosis and cellular vitality: Issues in osteoarthritic cartilage degeneration. Arthritis Rheumatol 2002, 46, 1986–1996. [Google Scholar]
- Iacono, A.; Gomez, R.; Sperry, J.; Conde, J.; Bianco, G.; Meli, R.; Gomez-Reino, J.J.; Smith, A.B., 3rd; Gualillo, O. Effect of oleocanthal and its derivatives on inflammatory response induced by lipopolysaccharide in a murine chondrocyte cell line. Arthritis Rheumatol 2010, 62, 1675–1682. [Google Scholar]
- De Oliveira, R.M.; Pais, T.F.; Outeiro, T.F. Sirtuins: Common targets in aging and in neurodegeneration. Curr. Drug Targets 2010, 11, 1270–1280. [Google Scholar]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol 2004, 4, 499–511. [Google Scholar]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol 2001, 2, 675–680. [Google Scholar]
- Estrov, Z.; Shishodia, S.; Faderl, S.; Harris, D.; Van, Q.; Kantarjian, H.M.; Talpazm, M.; Aggarwal, B.B. Resveratrol blocks interleukin-1beta-induced activation of the nuclear transcription factor NF-kappaB, inhibits proliferation, causes S-phase arrest, and induces apoptosis of acute myeloid leukemia cells. Blood 2003, 102, 987–995. [Google Scholar]
- Chen, Y.J.; Tsai, K.S.; Chiu, C.Y.; Yang, T.H.; Lin, T.H.; Fu, W.M.; Chen, C.F.; Yang, R.S.; Liu, S.H. EGb761 inhibits inflammatory responses in human chondrocytes and shows chondroprotection in osteoarthritic rat knee. J. Orthop. Res 2013, 31, 1032–1038. [Google Scholar]
- She, Q.B.; Bode, A.M.; Ma, W.Y.; Chen, N.Y.; Dong, Z. Resveratrol-induced activation of p53 and apoptosis is mediated by extracellular-signal-regulated protein kinases and p38 kinase. Cancer Res 2001, 61, 1604–1610. [Google Scholar]
- Jung, D.Y.; Lee, H.; Jung, B.Y.; Ock, J.; Lee, M.S.; Lee, W.H.; Suk, K. TLR4, but not TLR2, signals autoregulatory apoptosis of cultured microglia: A critical role of IFN-beta as a decision maker. J. Immunol 2005, 174, 6467–6476. [Google Scholar]
- Sebai, H.; Ristorcelli, E.; Sbarra, V.; Hovsepian, S.; Fayet, G.; Aouani, E.; Lombardo, D. Protective effect of resveratrol against LPS-induced extracellular lipoperoxidation in AR42J cells partly via a Myd88-dependent signaling pathway. Arch. Biochem. Biophys 2010, 495, 56–61. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar]






© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, L.; Gu, H.; Liu, H.; Jiao, Y.; Li, K.; Zhao, Y.; An, L.; Yang, J. Protective Effect of Resveratrol against IL-1β-Induced Inflammatory Response on Human Osteoarthritic Chondrocytes Partly via the TLR4/MyD88/NF-κB Signaling Pathway: An “in Vitro Study”. Int. J. Mol. Sci. 2014, 15, 6925-6940. https://doi.org/10.3390/ijms15046925
Liu L, Gu H, Liu H, Jiao Y, Li K, Zhao Y, An L, Yang J. Protective Effect of Resveratrol against IL-1β-Induced Inflammatory Response on Human Osteoarthritic Chondrocytes Partly via the TLR4/MyD88/NF-κB Signaling Pathway: An “in Vitro Study”. International Journal of Molecular Sciences. 2014; 15(4):6925-6940. https://doi.org/10.3390/ijms15046925
Chicago/Turabian StyleLiu, Li, Hailun Gu, Huimin Liu, Yongliang Jiao, Keyu Li, Yue Zhao, Li An, and Jun Yang. 2014. "Protective Effect of Resveratrol against IL-1β-Induced Inflammatory Response on Human Osteoarthritic Chondrocytes Partly via the TLR4/MyD88/NF-κB Signaling Pathway: An “in Vitro Study”" International Journal of Molecular Sciences 15, no. 4: 6925-6940. https://doi.org/10.3390/ijms15046925