
Ellagic Acid Prevents Particulate Matter-Induced Pulmonary Inflammation and Hyperactivity in Mice: A Pilot Study
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
- (1)
- Control (CON): 5% dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO, USA) was administered orally for 14 days, and after the eighth day of DMSO administration, distilled water was instilled for another 7 days.
- (2)
- PM-instilled (PMI): PM (5 mg/kg; standard reference material 2975; National Institute of Standards and Technology, Gaithersburg, MD, USA) was instilled for 7 days.
- (3)
- Low-dose of EA with PMI (EL + PMI): EA (20 mg/kg; Sigma-Aldrich) was administered orally for 14 days, and after the eighth day of EA administration, PM (5 mg/kg) was instilled for another 7 days.
- (4)
- High-dose of EA with PMI (EH + PMI): EA (100 mg/kg) was administered orally for 14 days, and after the eighth day of EA administration, PM (5 mg/kg) was instilled for another 7 days.
2.2. Histological Analysis and Collection of Bronchoalveolar Lavage Fluid (BALF)
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Quantitative Real-Time PCR (qRT-PCR)
2.5. Open Field Test
2.6. Statistical Analysis
3. Results
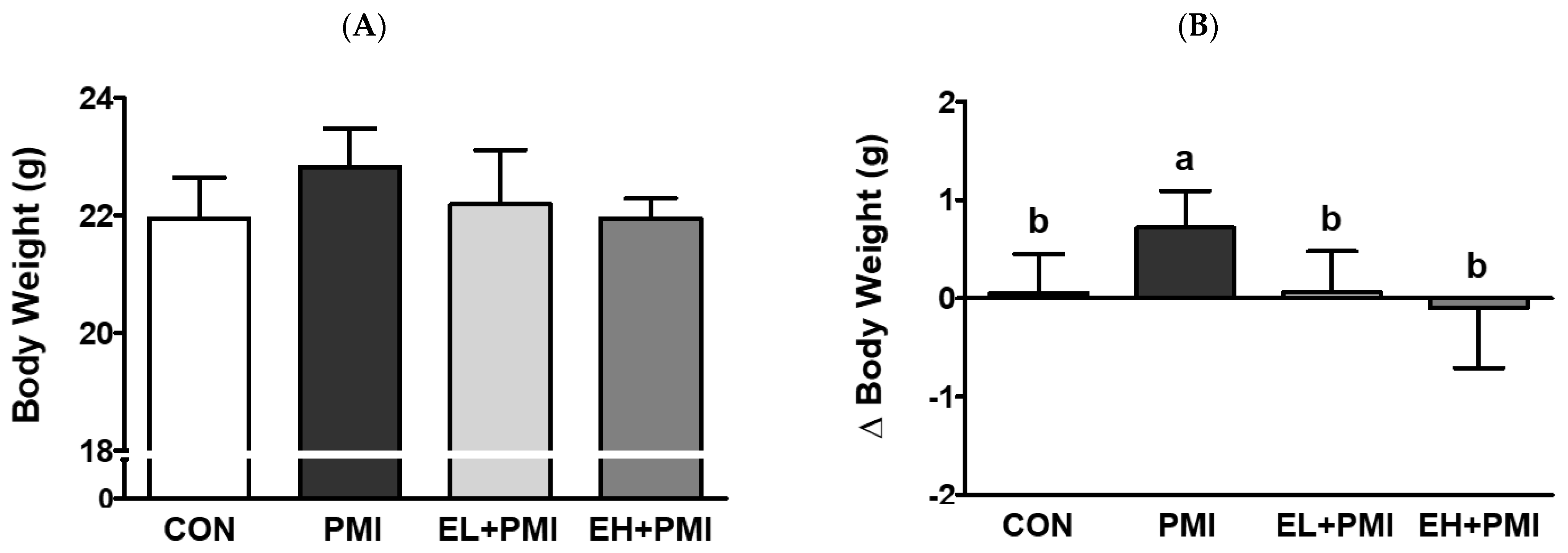
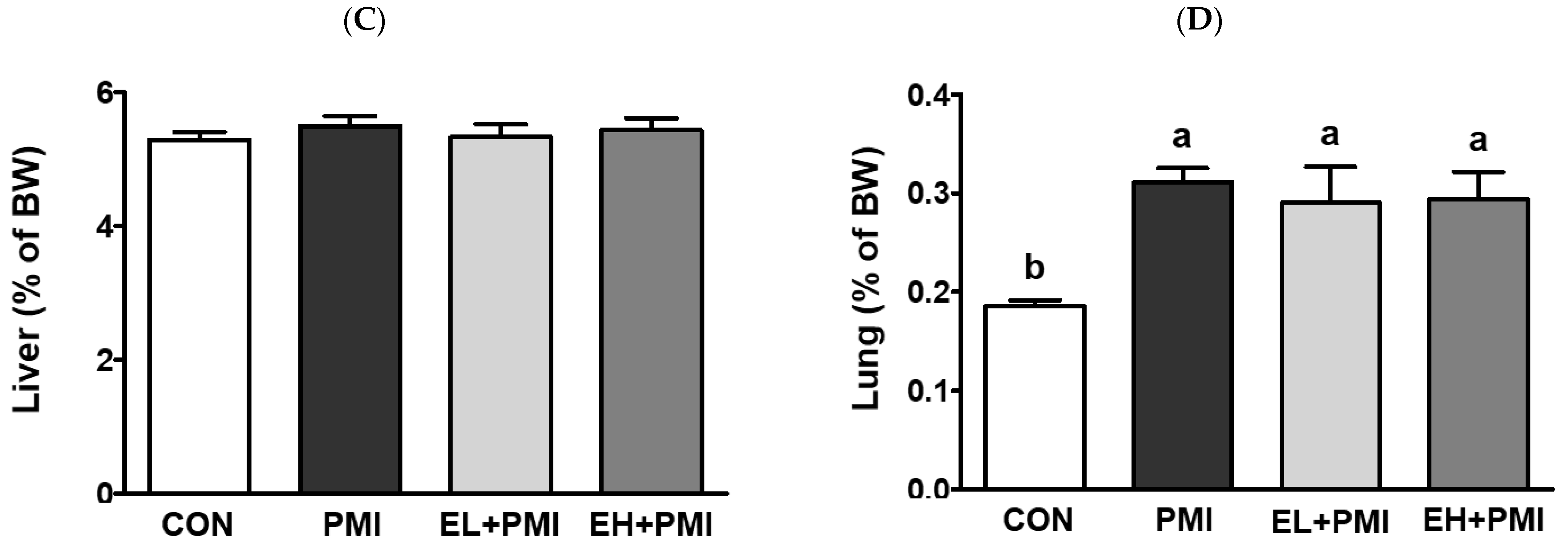
3.1. Alterations in Body and Relative Organ Weights
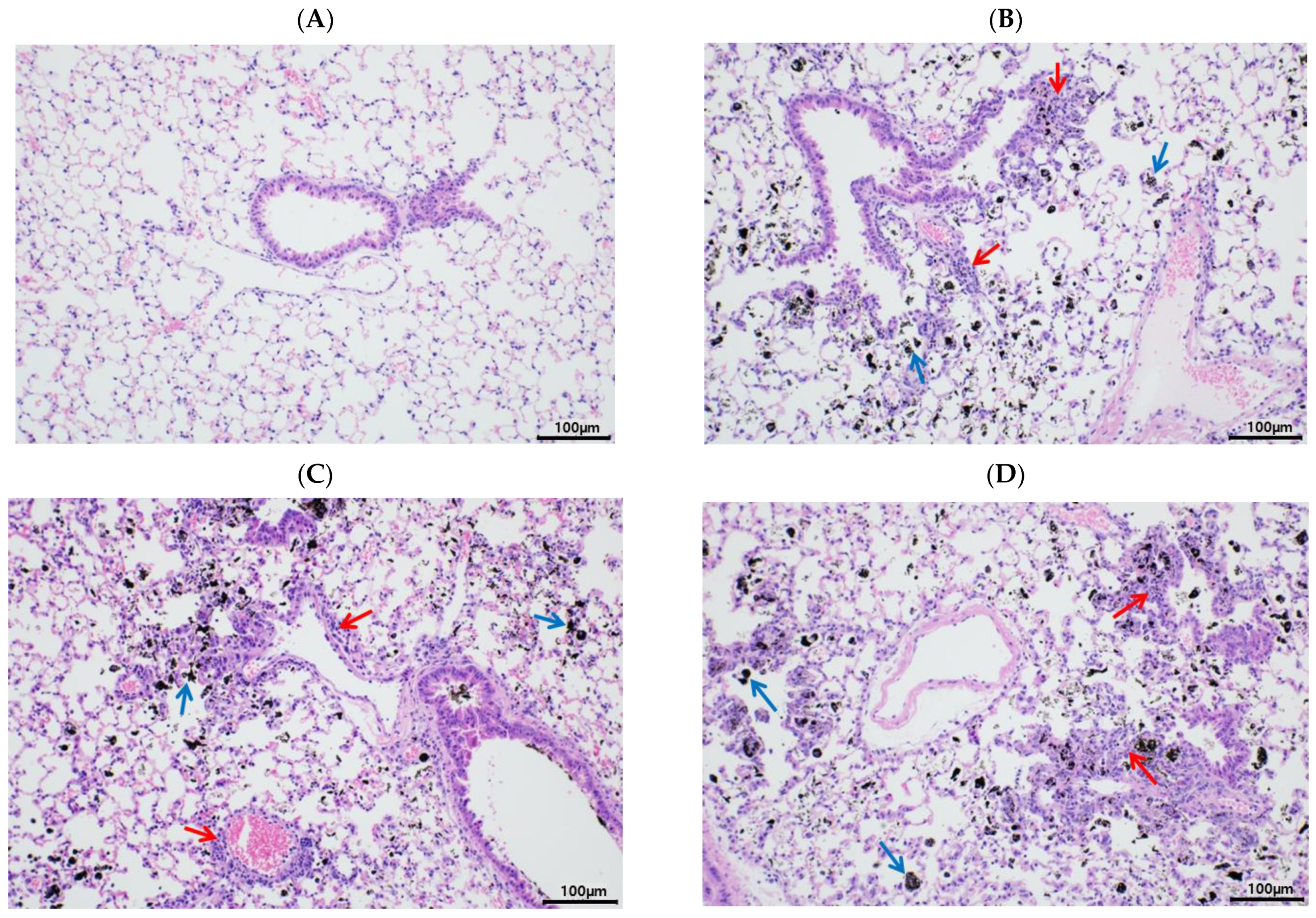
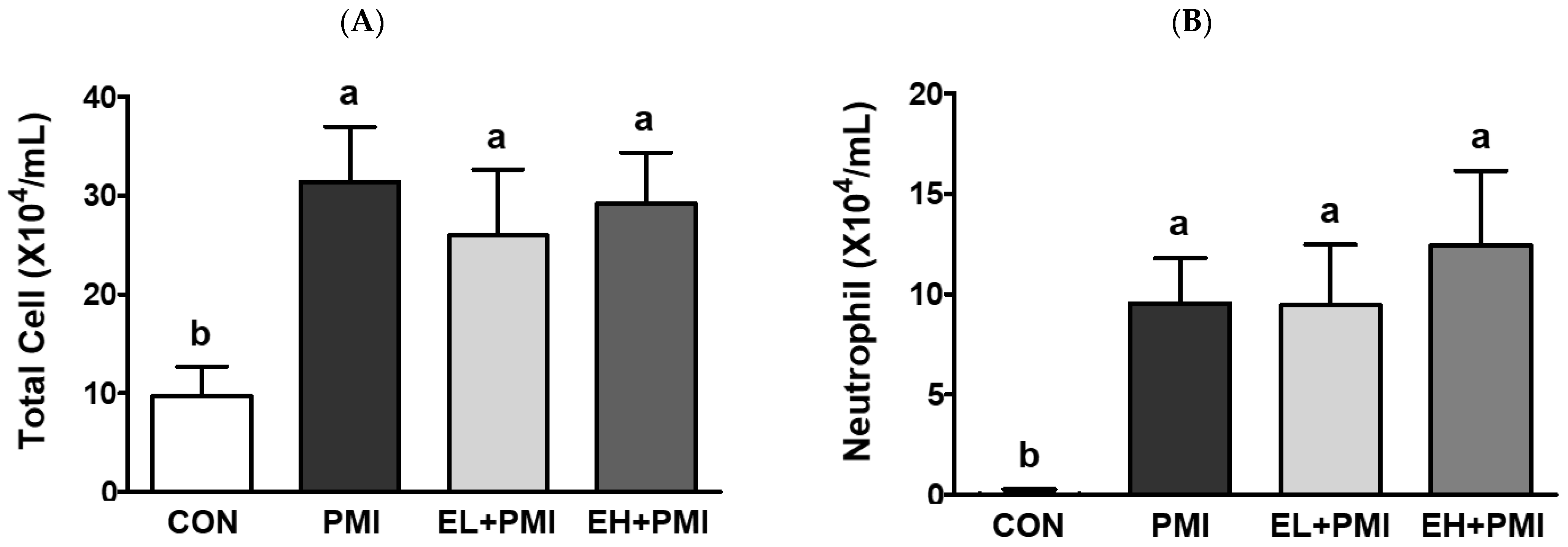
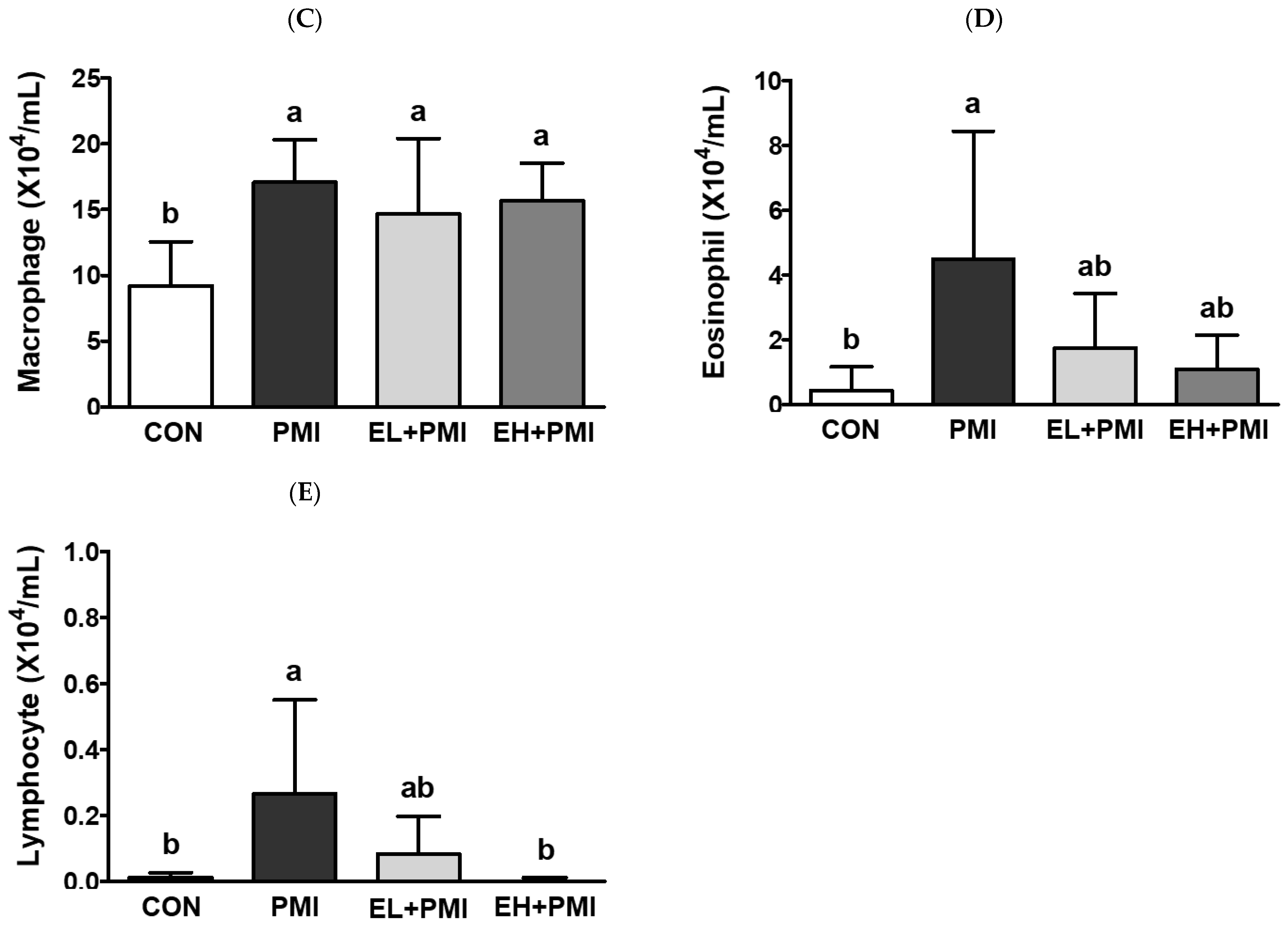
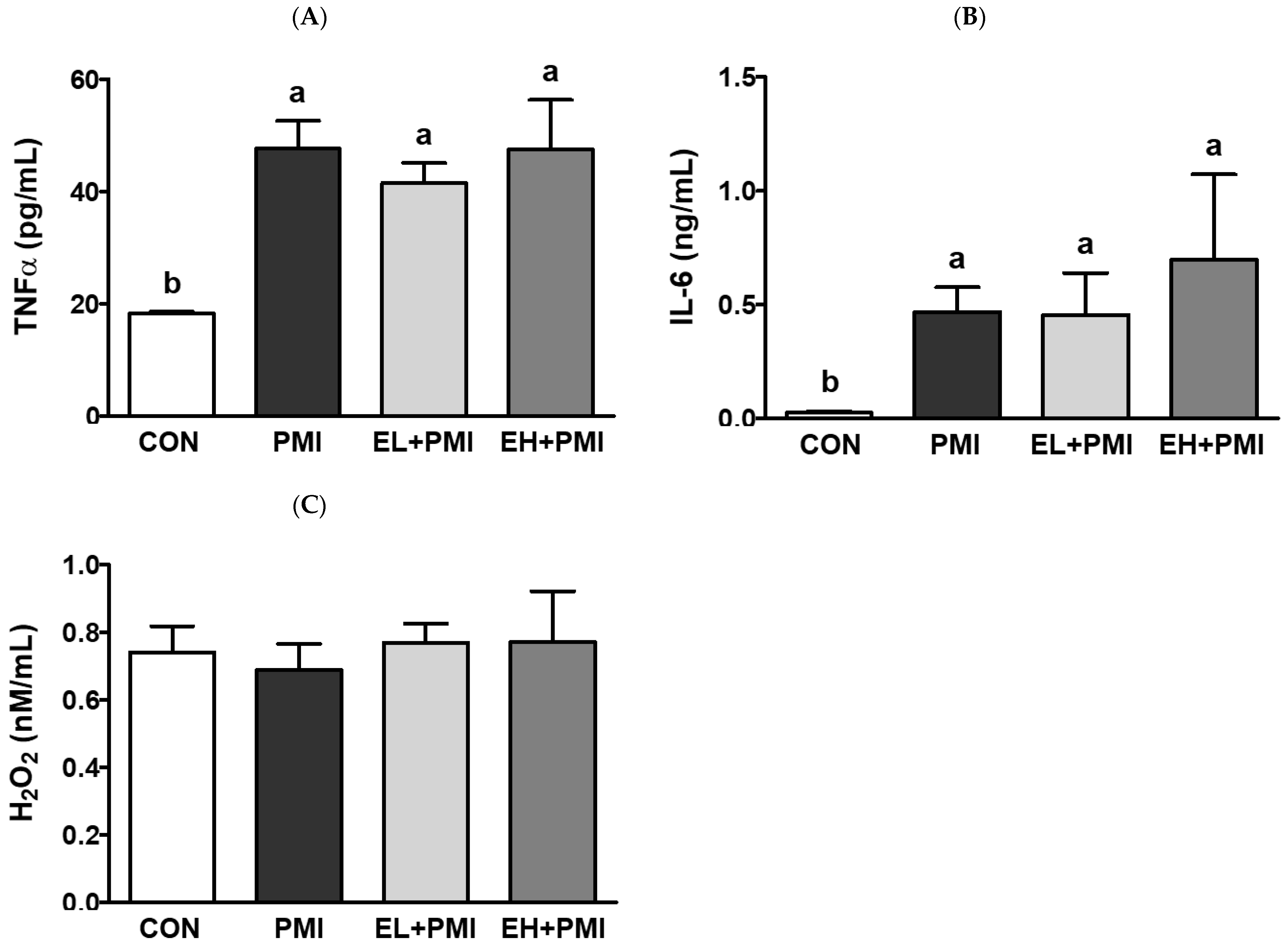
3.2. Pulmonary PM Loading and Inflammatory Cytokine Secretion
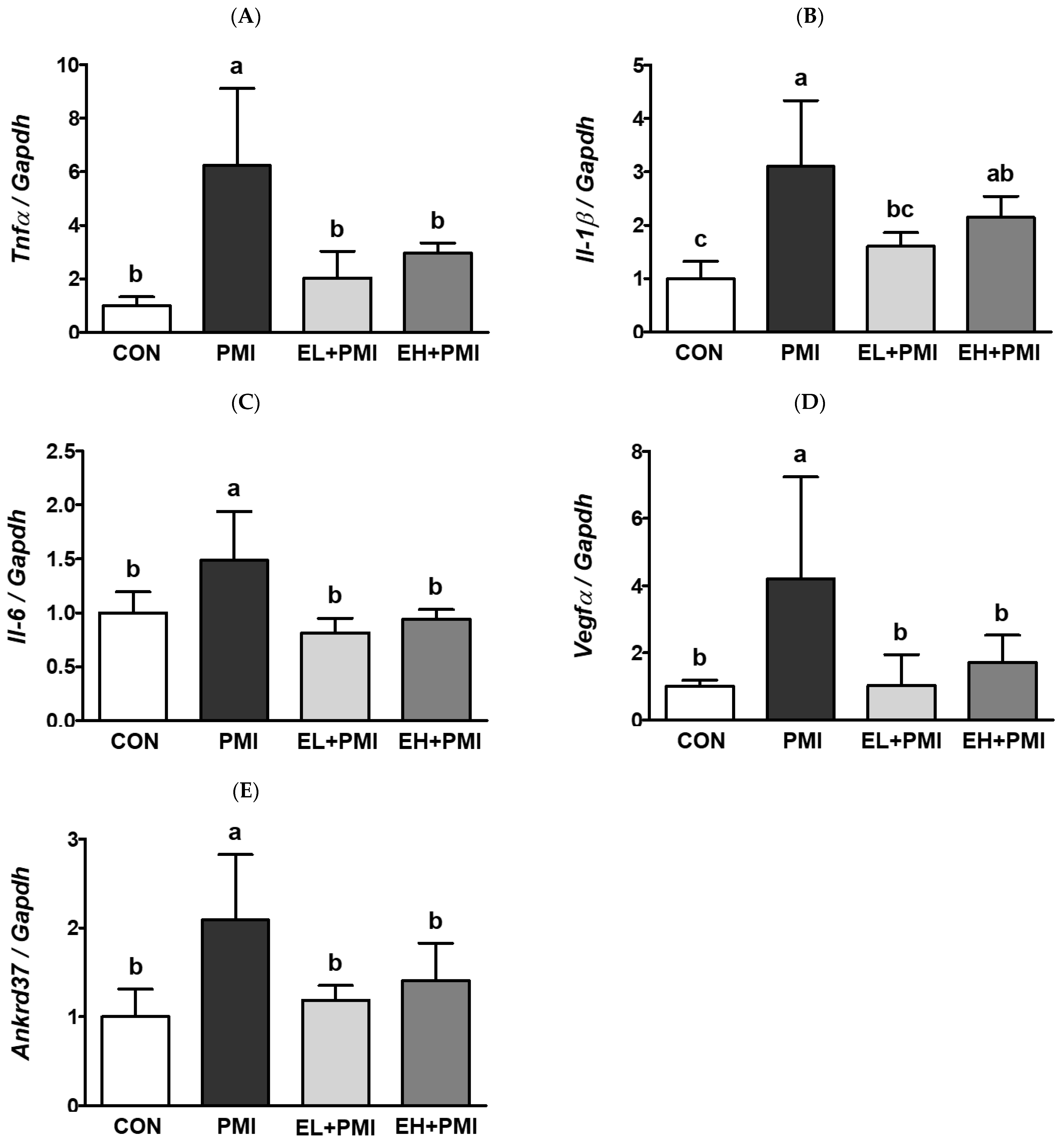
3.3. EA Treatment Prevented PM-Induced Expression of Inflammatory and Hypoxic Response Genes
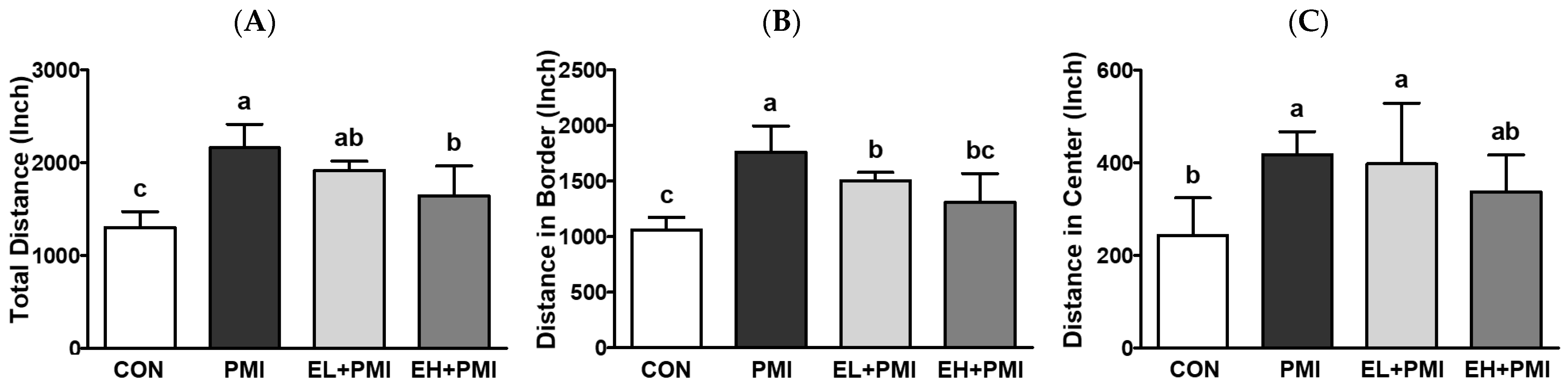
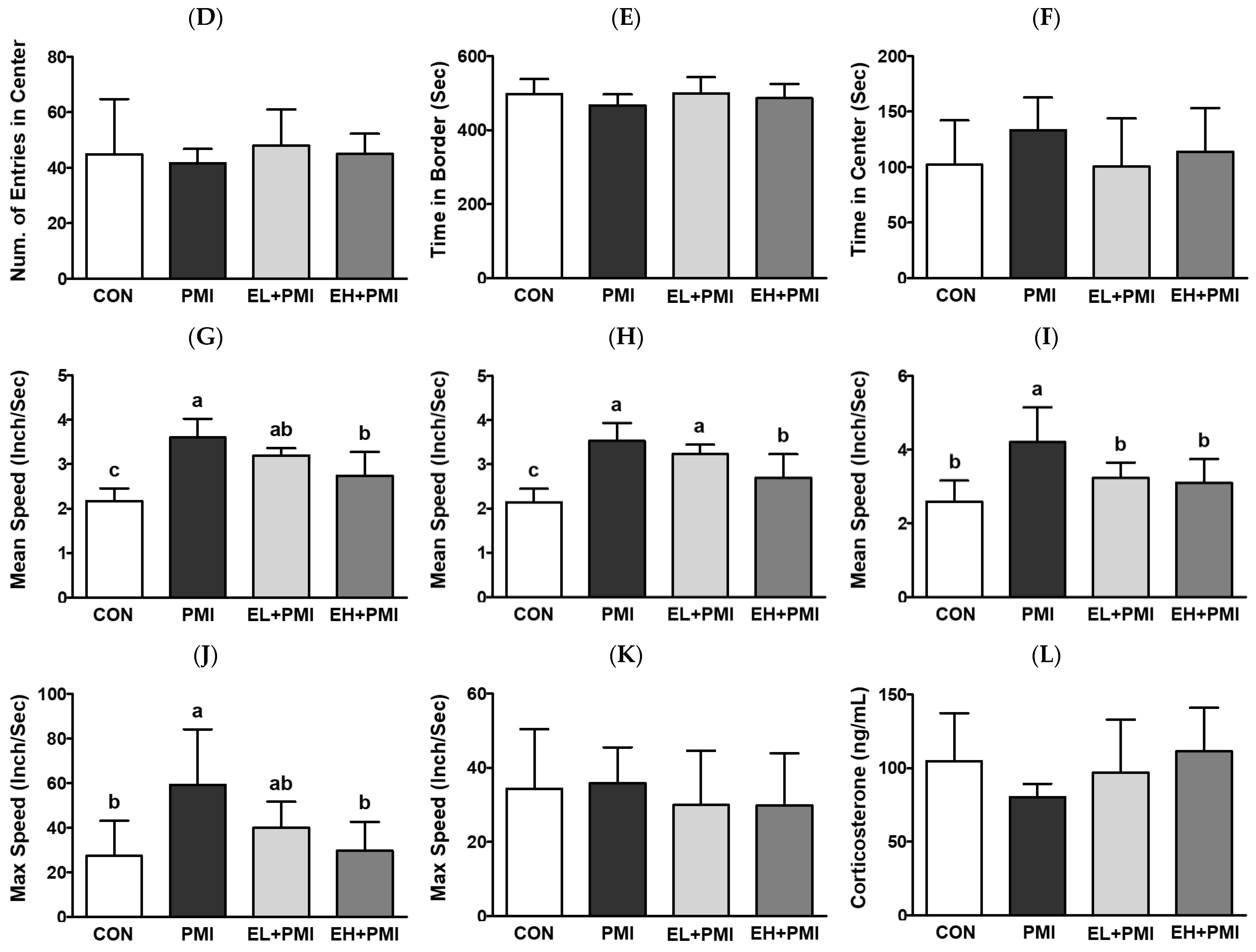
3.4. EA Treatment Prevented PM-Induced Hyperactivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wichmann, H.E. Diesel exhaust particles. Inhal. Toxicol. 2007, 19 (Suppl. 1), 241–244. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, T.; Furuyama, A.; Sagai, M. Biological effects of diesel exhaust particles (DEP). II. Acute toxicity of DEP introduced into lung by intratracheal instillation. Toxicology 1995, 99, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K.; Adachi, S.; Takahashi, M.; Moller, L.; Udagawa, T.; Mizuno, S.; Sugawara, I. Early oxidative DNA damages and late development of lung cancer in diesel exhaust-exposed rats. Environ. Res. 2000, 84, 255–264. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Zia, S.; Yasin, J.; Al Husseni, I.; Ali, B.H. Diesel exhaust particles in the lung aggravate experimental acute renal failure. Toxicol. Sci. 2010, 113, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morsi, A.A.; Fouad, H.; Alasmari, W.A.; Faruk, E.M. The biomechanistic aspects of renal cortical injury induced by diesel exhaust particles in rats and the renoprotective contribution of quercetin pretreatment: Histological and biochemical study. Environ. Toxicol. 2022, 37, 310–321. [Google Scholar] [CrossRef]
- Ito, Y.; Yanagiba, Y.; Ramdhan, D.H.; Hayashi, Y.; Li, Y.; Suzuki, A.K.; Kamijima, M.; Nakajima, T. Nanoparticle-rich diesel exhaust-induced liver damage via inhibited transactivation of peroxisome proliferator-activated receptor alpha. Environ. Toxicol. 2016, 31, 1985–1995. [Google Scholar] [CrossRef]
- Miller, M.R.; Newby, D.E. Air pollution and cardiovascular disease: Car sick. Cardiovasc. Res. 2020, 116, 279–294. [Google Scholar] [CrossRef]
- Olumegbon, L.T.; Lawal, A.O.; Oluyede, D.M.; Adebimpe, M.O.; Elekofehinti, O.O.; Umar, H.U. Hesperetin protects against diesel exhaust particles-induced cardiovascular oxidative stress and inflammation in Wistar rats. Environ. Sci. Pollut. Res. Int. 2022, 29, 52574–52589. [Google Scholar] [CrossRef]
- Kang, Y.J.; Tan, H.Y.; Lee, C.Y.; Cho, H. An Air Particulate Pollutant Induces Neuroinflammation and Neurodegeneration in Human Brain Models. Adv. Sci. 2021, 8, e2101251. [Google Scholar] [CrossRef]
- Kioumourtzoglou, M.A.; Schwartz, J.D.; Weisskopf, M.G.; Melly, S.J.; Wang, Y.; Dominici, F.; Zanobetti, A. Long-term PM2.5 Exposure and Neurological Hospital Admissions in the Northeastern United States. Environ. Health Perspect. 2016, 124, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Nemmar, A.; Al-Salam, S.; Yuvaraju, P.; Beegam, S.; Ali, B.H. Emodin mitigates diesel exhaust particles-induced increase in airway resistance, inflammation and oxidative stress in mice. Respir. Physiol. Neurobiol. 2015, 215, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Hamadi, N.; Ali, B.H. In Vivo Protective Effects of Nootkatone against Particles-Induced Lung Injury Caused by Diesel Exhaust Is Mediated via the NF-kappaB Pathway. Nutrients 2018, 10, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.C.; Chen, C.M.; Tsai, J.D.; Chiang, K.L.; Tsai, S.C.; Huang, C.Y.; Lin, C.L.; Hsu, C.Y.; Chang, K.H. Association between Exposure to Particulate Matter Air Pollution during Early Childhood and Risk of Attention-Deficit/Hyperactivity Disorder in Taiwan. Int. J. Environ. Res. Public Health 2022, 19, 6138. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chen, W.T.; Su, S.H.; Jung, C.R.; Hwang, B.F. PM2.5 exposure and incident attention-deficit/hyperactivity disorder during the prenatal and postnatal periods: A birth cohort study. Environ. Res. 2022, 214, 113769. [Google Scholar] [CrossRef]
- Khan, A.; Plana-Ripoll, O.; Antonsen, S.; Brandt, J.; Geels, C.; Landecker, H.; Sullivan, P.F.; Pedersen, C.B.; Rzhetsky, A. Environmental pollution is associated with increased risk of psychiatric disorders in the US and Denmark. PLoS Biol. 2019, 17, e3000353. [Google Scholar] [CrossRef] [Green Version]
- Weitekamp, C.A.; Hofmann, H.A. Effects of air pollution exposure on social behavior: A synthesis and call for research. Environ. Health 2021, 20, 72. [Google Scholar] [CrossRef]
- Roberts, S.; Arseneault, L.; Barratt, B.; Beevers, S.; Danese, A.; Odgers, C.L.; Moffitt, T.E.; Reuben, A.; Kelly, F.J.; Fisher, H.L. Exploration of NO2 and PM2.5 air pollution and mental health problems using high-resolution data in London-based children from a UK longitudinal cohort study. Psychiatry Res. 2019, 272, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Thirtamara Rajamani, K.; Doherty-Lyons, S.; Bolden, C.; Willis, D.; Hoffman, C.; Zelikoff, J.; Chen, L.C.; Gu, H. Prenatal and early-life exposure to high-level diesel exhaust particles leads to increased locomotor activity and repetitive behaviors in mice. Autism Res. 2013, 6, 248–257. [Google Scholar] [CrossRef]
- Cui, J.; Fu, Y.; Lu, R.; Bi, Y.; Zhang, L.; Zhang, C.; Aschner, M.; Li, X.; Chen, R. Metabolomics analysis explores the rescue to neurobehavioral disorder induced by maternal PM2.5 exposure in mice. Ecotoxicol. Environ. Saf. 2019, 169, 687–695. [Google Scholar] [CrossRef]
- Jeong, S.; Lee, J.H.; Ha, J.H.; Kim, J.; Kim, I.; Bae, S. An Exploratory Study of the Relationships between Diesel Engine Exhaust Particle Inhalation, Pulmonary Inflammation and Anxious Behavior. Int. J. Environ. Res. Public Health 2021, 18, 1166. [Google Scholar] [CrossRef]
- Jeong, S.; Bae, S.; Yu, D.; Yang, H.S.; Yang, M.J.; Lee, J.H.; Ha, J.H. Dietary Intervention with Quercetin Attenuates Diesel Exhaust Particle-Instilled Pulmonary Inflammation and Behavioral Abnormalities in Mice. J. Med. Food 2023, 26, 2. [Google Scholar] [CrossRef]
- Tong, H. Dietary and pharmacological intervention to mitigate the cardiopulmonary effects of air pollution toxicity. Biochim. Biophys. Acta 2016, 1860, 2891–2898. [Google Scholar] [CrossRef]
- Romieu, I.; Castro-Giner, F.; Kunzli, N.; Sunyer, J. Air pollution, oxidative stress and dietary supplementation: A review. Eur. Respir. J. 2008, 31, 179–197. [Google Scholar] [CrossRef]
- Peter, S.; Holguin, F.; Wood, L.G.; Clougherty, J.E.; Raederstorff, D.; Antal, M.; Weber, P.; Eggersdorfer, M. Nutritional Solutions to Reduce Risks of Negative Health Impacts of Air Pollution. Nutrients 2015, 7, 10398–10416. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.H.; Bae, C.H.; Kim, J.H.; Park, S.D.; Shim, J.J.; Lee, J.L. Lactobacillus casei HY2782 and Pueraria lobata Root Extract Complex Ameliorates Particulate Matter-Induced Airway Inflammation in Mice by Inhibiting Th2 and Th17 Immune Responses. Prev. Nutr. Food Sci. 2022, 27, 188–197. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Gallego, J.; Garcia-Mediavilla, M.V.; Sanchez-Campos, S.; Tunon, M.J. Fruit polyphenols, immunity and inflammation. Br. J. Nutr. 2010, 104 (Suppl. 3), S15–S27. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.J.; Yoon, D.S.; Baek, H.J.; Kang, B.; Jung, U.J. Dietary Sinapic Acid Alleviates Adiposity and Inflammation in Diet-Induced Obese Mice. Prev. Nutr. Food Sci. 2022, 27, 407–413. [Google Scholar] [CrossRef]
- Lee, H.; Im, S.W.; Jung, C.H.; Jang, Y.J.; Ha, T.Y.; Ahn, J. Tyrosol, an olive oil polyphenol, inhibits ER stress-induced apoptosis in pancreatic beta-cell through JNK signaling. Biochem. Biophys. Res. Commun. 2016, 469, 748–752. [Google Scholar] [CrossRef]
- Yan, B.; Chen, L.; Wang, Y.; Zhang, J.; Zhao, H.; Hua, Q.; Pei, S.; Yue, Z.; Liang, H.; Zhang, H. Preventive Effect of Apple Polyphenol Extract on High-Fat Diet-Induced Hepatic Steatosis in Mice through Alleviating Endoplasmic Reticulum Stress. J. Agric. Food Chem. 2022, 70, 3172–3180. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Stagos, D. Antioxidant Activity of Polyphenolic Plant Extracts. Antioxidants 2019, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Park, S.K. Antioxidant Activities of Bioactive Compounds Isolated from Rheum emodi Wall (Himalayan Rhubarb) Based on LC-DAD-ESI/MS and Preparative LC/MS System. Prev. Nutr. Food Sci. 2022, 27, 223–233. [Google Scholar] [CrossRef]
- Evtyugin, D.D.; Magina, S.; Evtuguin, D.V. Recent Advances in the Production and Applications of Ellagic Acid and Its Derivatives. A Review. Molecules 2020, 25, 2745. [Google Scholar] [CrossRef]
- Lee, K.H.; Jeong, E.S.; Jang, G.; Na, J.R.; Park, S.; Kang, W.S.; Kim, E.; Choi, H.; Kim, J.S.; Kim, S. Unripe Rubus coreanus Miquel Extract Containing Ellagic Acid Regulates AMPK, SREBP-2, HMGCR, and INSIG-1 Signaling and Cholesterol Metabolism In Vitro and In Vivo. Nutrients 2020, 12, 610. [Google Scholar] [CrossRef] [Green Version]
- Shiojima, Y.; Takahashi, M.; Kikuchi, M.; Akanuma, M. Effect of ellagic acid on body fat and triglyceride reduction in healthy overweight volunteers: A randomized, double-blind, placebo-controlled parallel group study. Funct. Foods Health Dis. 2020, 10, 180–194. [Google Scholar] [CrossRef]
- Ghadimi, M.; Foroughi, F.; Hashemipour, S.; Nooshabadi, M.R.; Ahmadi, M.H.; Yari, M.G.; Kavianpour, M.; Haghighian, H.K. Decreased insulin resistance in diabetic patients by influencing Sirtuin1 and Fetuin-A following supplementation with ellagic acid: A randomized controlled trial. Diabetol. Metab. Syndr. 2021, 13, 16. [Google Scholar] [CrossRef]
- Umesalma, S.; Sudhandiran, G. Ellagic acid prevents rat colon carcinogenesis induced by 1, 2 dimethyl hydrazine through inhibition of AKT-phosphoinositide-3 kinase pathway. Eur. J. Pharmacol. 2011, 660, 249–258. [Google Scholar] [CrossRef]
- Mohammadinejad, A.; Mohajeri, T.; Aleyaghoob, G.; Heidarian, F.; Kazemi Oskuee, R. Ellagic acid as a potent anticancer drug: A comprehensive review on in vitro, in vivo, in silico, and drug delivery studies. Biotechnol. Appl. Biochem. 2022, 69, 2323–2356. [Google Scholar] [CrossRef]
- Cornelio Favarin, D.; Martins Teixeira, M.; Lemos de Andrade, E.; de Freitas Alves, C.; Lazo Chica, J.E.; Arterio Sorgi, C.; Faccioli, L.H.; Paula Rogerio, A. Anti-inflammatory effects of ellagic acid on acute lung injury induced by acid in mice. Mediat. Inflamm. 2013, 2013, 164202. [Google Scholar] [CrossRef] [Green Version]
- Marin, M.; Maria Giner, R.; Rios, J.L.; Recio, M.C. Intestinal anti-inflammatory activity of ellagic acid in the acute and chronic dextrane sulfate sodium models of mice colitis. J. Ethnopharmacol. 2013, 150, 925–934. [Google Scholar] [CrossRef]
- Zeb, A. Ellagic acid in suppressing in vivo and in vitro oxidative stresses. Mol. Cell. Biochem. 2018, 448, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.Y.; Fatemi, I.; Kalantari, H.; Mombeini, M.A.; Mehrzadi, S.; Goudarzi, M. Ellagic Acid Prevents Oxidative Stress, Inflammation, and Histopathological Alterations in Acrylamide-Induced Hepatotoxicity in Wistar Rats. J. Diet. Suppl. 2020, 17, 651–662. [Google Scholar] [CrossRef]
- Aslan, A.; Hussein, Y.T.; Gok, O.; Beyaz, S.; Erman, O.; Baspinar, S. Ellagic acid ameliorates lung damage in rats via modulating antioxidant activities, inhibitory effects on inflammatory mediators and apoptosis-inducing activities. Environ. Sci. Pollut. Res. Int. 2020, 27, 7526–7537. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, Z.; Dianat, M.; Radan, M.; Badavi, M. Ellagic Acid Ameliorates Lung Inflammation and Heart Oxidative Stress in Elastase-Induced Emphysema Model in Rat. Inflammation 2020, 43, 1143–1156. [Google Scholar] [CrossRef] [PubMed]
- Saba; Khan, S.; Parvez, S.; Chaudhari, B.; Ahmad, F.; Anjum, S.; Raisuddin, S. Ellagic acid attenuates bleomycin and cyclophosphamide-induced pulmonary toxicity in Wistar rats. Food Chem. Toxicol. 2013, 58, 210–219. [Google Scholar] [CrossRef]
- Zhou, E.; Fu, Y.; Wei, Z.; Yang, Z. Inhibition of allergic airway inflammation through the blockage of NF-kappaB activation by ellagic acid in an ovalbumin-induced mouse asthma model. Food Funct. 2014, 5, 2106–2112. [Google Scholar] [CrossRef]
- Guan, S.; Zheng, Y.; Yu, X.; Li, W.; Han, B.; Lu, J. Ellagic acid protects against LPS-induced acute lung injury through inhibition of nuclear factor kappa B, proinflammatory cytokines and enhancement of interleukin-10. Food Agric. Immunol. 2017, 28, 1347–1361. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.T.; Colgan, S.P. Regulation of immunity and inflammation by hypoxia in immunological niches. Nat. Rev. Immunol. 2017, 17, 774–785. [Google Scholar] [CrossRef]
- Pham, K.; Parikh, K.; Heinrich, E.C. Hypoxia and Inflammation: Insights from High-Altitude Physiology. Front. Physiol. 2021, 12, 676782. [Google Scholar] [CrossRef]
- Thygesen, M.; Holst, G.J.; Hansen, B.; Geels, C.; Kalkbrenner, A.; Schendel, D.; Brandt, J.; Pedersen, C.B.; Dalsgaard, S. Exposure to air pollution in early childhood and the association with Attention-Deficit Hyperactivity Disorder. Environ. Res. 2020, 183, 108930. [Google Scholar] [CrossRef] [PubMed]
- McGarry, T.; Biniecka, M.; Veale, D.J.; Fearon, U. Hypoxia, oxidative stress and inflammation. Free Radic. Biol. Med. 2018, 125, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, M.; Benevenuto, S.G.M.; Tomasini, P.P.; Yariwake, V.Y.; de Oliveira Alves, N.; Rahmeier, F.L.; da Cruz Fernandes, M.; Moura, D.J.; Nascimento Saldiva, P.H.; Veras, M.M. Concentrated ambient fine particulate matter (PM2.5) exposure induce brain damage in pre and postnatal exposed mice. Neurotoxicology 2020, 79, 127–141. [Google Scholar] [CrossRef] [PubMed]
- BenSaad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and punicalagin A&B isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- Bains, M.; Kaur, J.; Akhtar, A.; Kuhad, A.; Sah, S.P. Anti-inflammatory effects of ellagic acid and vanillic acid against quinolinic acid-induced rat model of Huntington’s disease by targeting IKK-NF-kappaB pathway. Eur. J. Pharmacol. 2022, 934, 175316. [Google Scholar] [CrossRef]
- Bidanchi, R.M.; Lalrindika, L.; Khushboo, M.; Bhanushree, B.; Dinata, R.; Das, M.; Nisa, N.; Lalrinzuali, S.; Manikandan, B.; Saeed-Ahmed, L.; et al. Antioxidative, anti-inflammatory and anti-apoptotic action of ellagic acid against lead acetate induced testicular and hepato-renal oxidative damages and pathophysiological changes in male Long Evans rats. Environ. Pollut. 2022, 302, 119048. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| Tnfα | GGCTGCCCCGACTACGT | ACTTTCTCCTGGTATGAGATAGCAAAT |
| Il-1β | GTCACAAGAAACCATGGCACAT | GCCCATCAGAGGCAAGGA |
| Il-6 | CTGCAAGAGACTTCCATCCAGTT | AGGGAAGGCCGTGGTTGT |
| Vegfα | GAGCAGAAGTCCCATGAAGTG | TGTCCACCAGGGTCTCAATC |
| Ankrd37 | CGGCCTTGCGTGCTTT | TGGTTGAGGTCAGCACCTGTT |
| Gapdh | CATGGCCTTCCGTGTTCCTA | GCGGCACGTCAGATCCA |
| Strain | Model/Inducer | EA Treatment | Biological Marker | Ref. |
|---|---|---|---|---|
| BALB/c mice | Acute lung injury/hydrochloric acid | 10 mg/kg, p.o. | BALF and lung neutrophil ↓ | [40] |
| BALF IL-6 ↓ | ||||
| BALF IL-10 ↑ | ||||
| BALB/c mice | Acute lung injury/LPS | 5 mg/kg, i.p. | BALF TNFα, IL-1β, IL-6 ↓ | [48] |
| BALF IL-10 ↑ | ||||
| BALF total protein ↓ | ||||
| Lung myeloperoxidase ↓ | ||||
| Sprague Dawley rats | Emphysema/elastase | 30 mg/kg, p.o. | Lung edema ↓ | [45] |
| Lung immune cell infiltration ↓ | ||||
| BALF immune cells ↓ | ||||
| BALF TNFα and IL-6 ↓ | ||||
| BALB/c mice | Asthma/ovalbumin | 10 mg/kg, p.o. | BALF eosinophils ↓ | [47] |
| Mucus production ↓ | ||||
| BALF IL-4, IL-5, IL-13 ↓ | ||||
| Lung NFκB activation ↓ | ||||
| Wistar rats | Oxidative stress/carbon tetrachloride | 10 mg/kg, i.p. | Lung malondialdehyde ↓ | [44] |
| Lung catalase activity, glutathione, Nrf-2 ↑ | ||||
| Lung NFκB, COX-2, TNFα ↓ | ||||
| Wistar rats | Fibrosis-like/bleomycin and cyclophosphamide | 15 mg/kg, p.o. | Lung hydroxyproline ↓ | [46] |
| Lung lipid peroxidation ↓ | ||||
| Lung glutathione, antioxidant enzymes ↑ | ||||
| Lung myeloperoxidase ↓ | ||||
| BALF immune cell, total protein ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Bae, S.; Shin, E.-C.; Lee, J.-H.; Ha, J.-H. Ellagic Acid Prevents Particulate Matter-Induced Pulmonary Inflammation and Hyperactivity in Mice: A Pilot Study. Int. J. Environ. Res. Public Health 2023, 20, 4523. https://doi.org/10.3390/ijerph20054523
Jeong S, Bae S, Shin E-C, Lee J-H, Ha J-H. Ellagic Acid Prevents Particulate Matter-Induced Pulmonary Inflammation and Hyperactivity in Mice: A Pilot Study. International Journal of Environmental Research and Public Health. 2023; 20(5):4523. https://doi.org/10.3390/ijerph20054523
Chicago/Turabian StyleJeong, Sunyoung, Sungryong Bae, Eui-Cheol Shin, Jong-Hwa Lee, and Jung-Heun Ha. 2023. "Ellagic Acid Prevents Particulate Matter-Induced Pulmonary Inflammation and Hyperactivity in Mice: A Pilot Study" International Journal of Environmental Research and Public Health 20, no. 5: 4523. https://doi.org/10.3390/ijerph20054523