
Follicle-Stimulating Hormone Promotes the Development of Endometrial Cancer In Vitro and In Vivo

Abstract
:1. Introduction
2. Methods
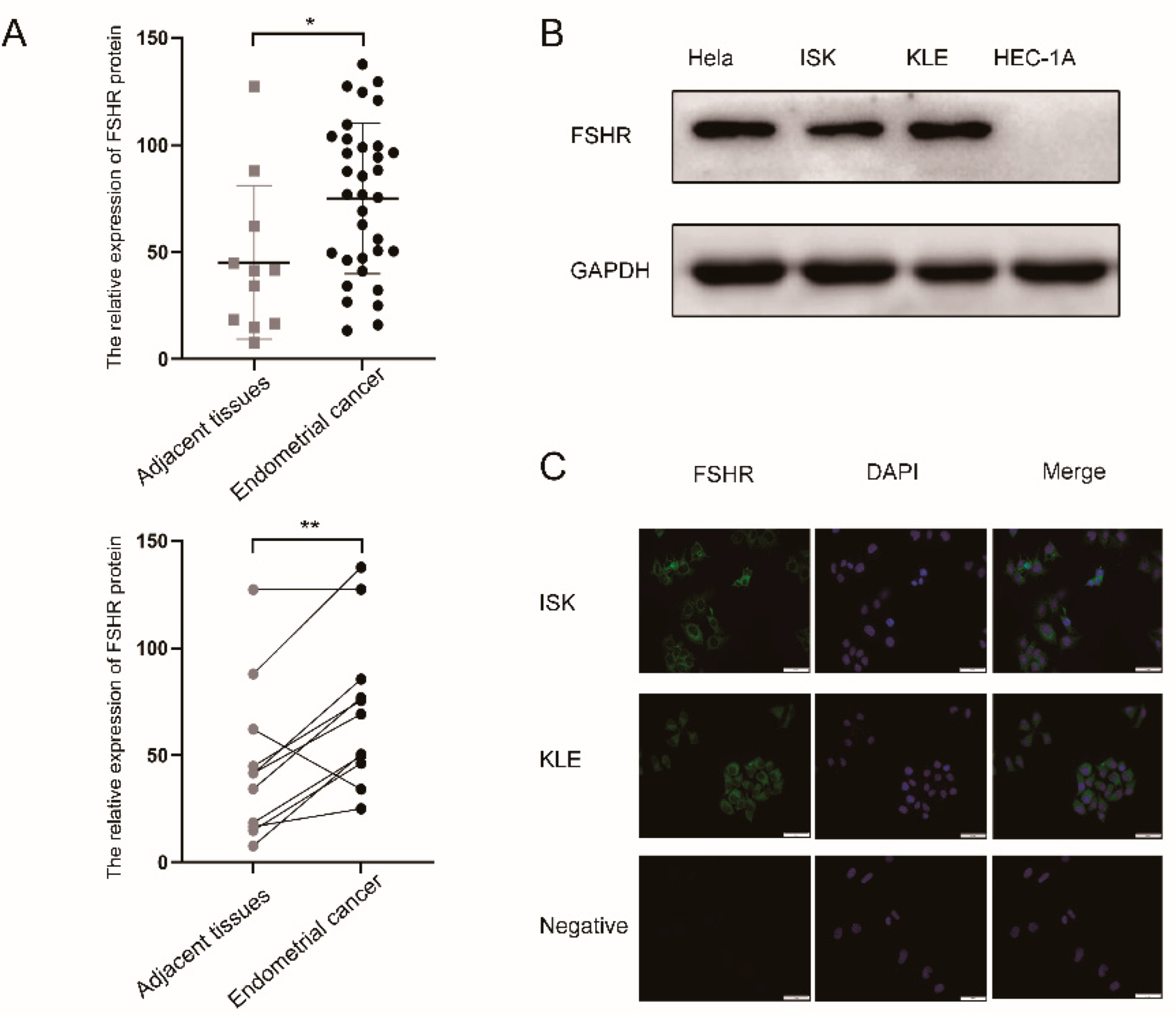
2.1. Human Tissue and Tissue Microarray
2.2. Immunohistochemistry
2.3. Cell Lines and Cell Culture
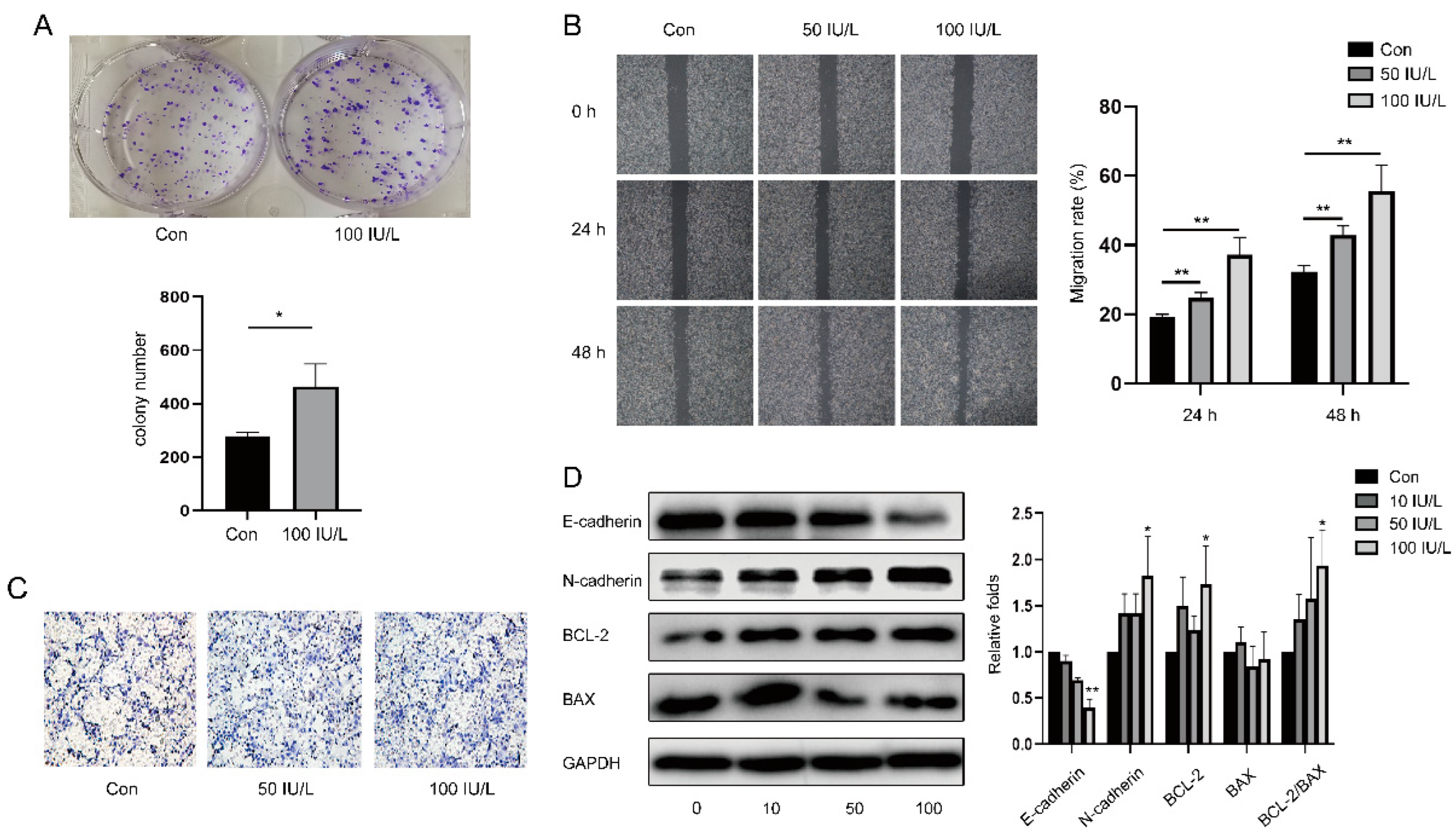
2.4. Colony Formation Assay
2.5. Cell Scratch Assay
2.6. Transwell Migration Assay
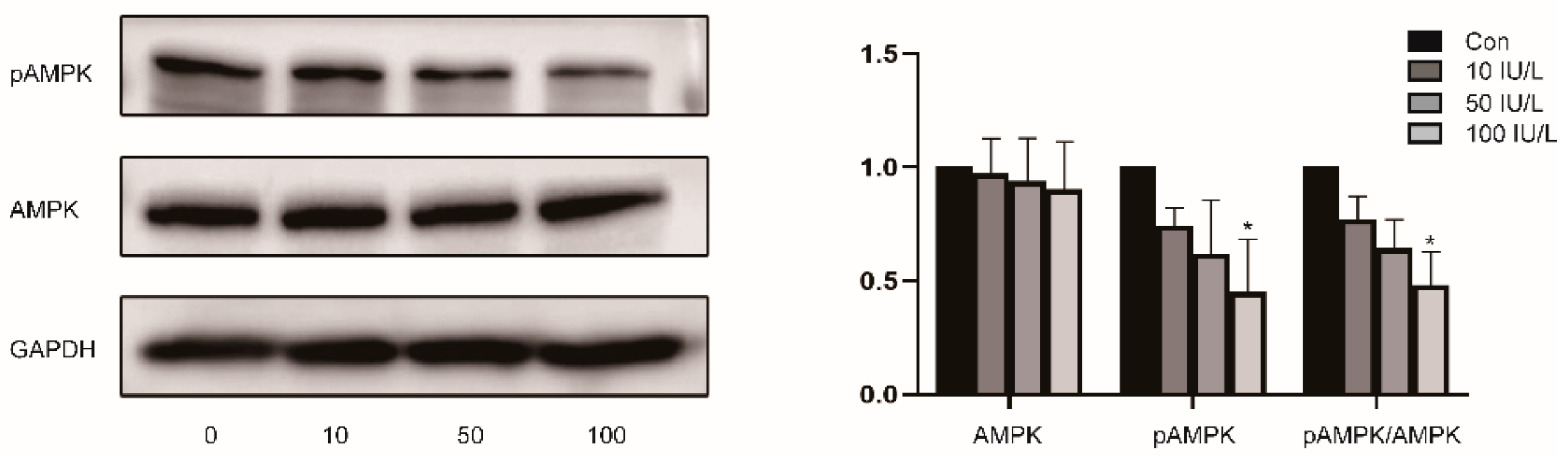
2.7. Western Blot
2.8. Immunofluorescence
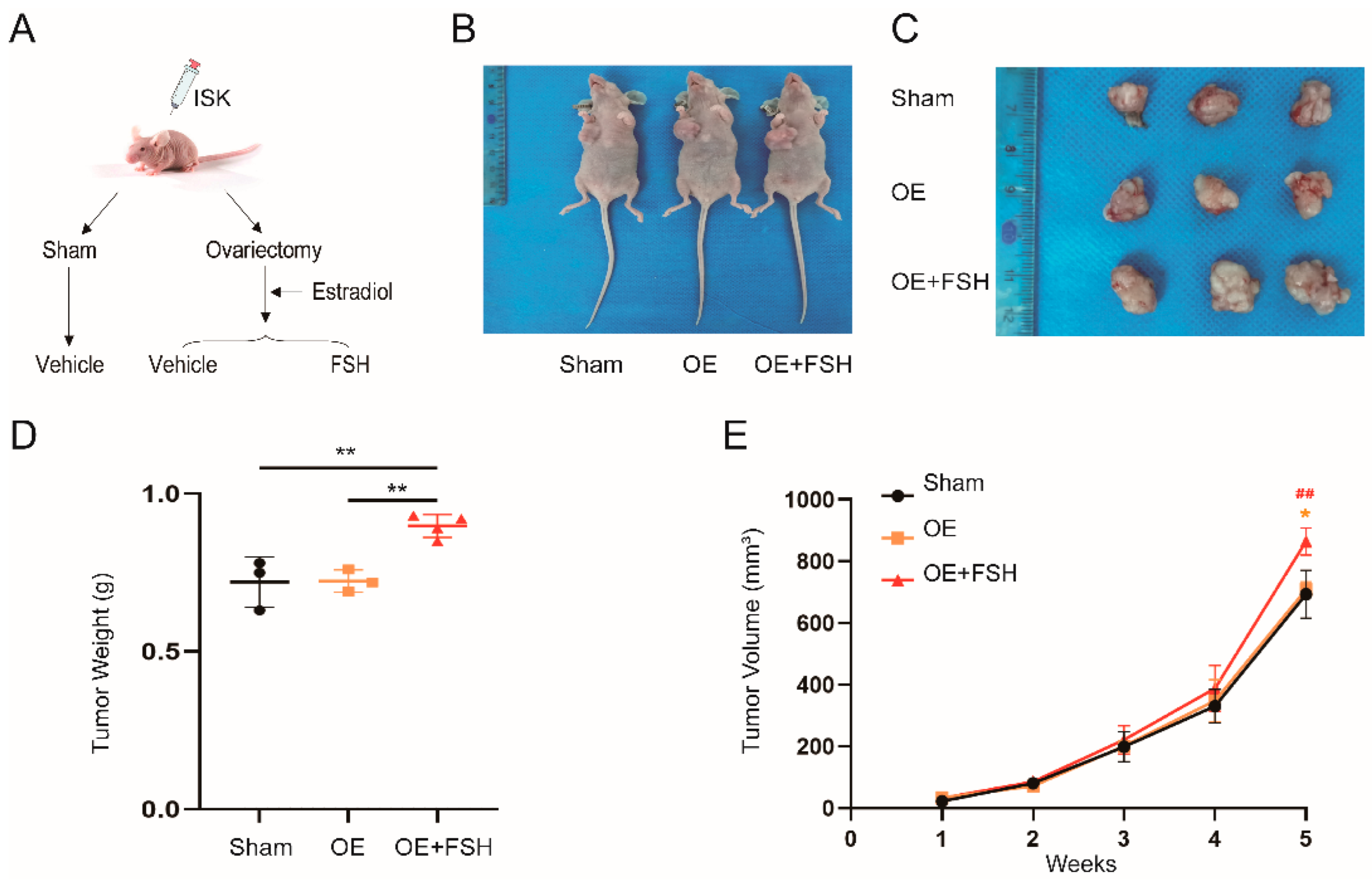
2.9. Animal Study
2.10. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EC | endometrial cancer |
| FSH | follicle-stimulating hormone |
| FSHR | FSH receptor |
| IHC | immunohistochemistry |
| IF | immunofluorescence |
| WB | Western blot |
| ISK | Ishikawa |
| E2 | oestradiol |
| GPCRs | G protein-coupled receptors |
| EDCs | endocrine disruptor compounds |
| AMPK | AMP-activated protein kinase |
| mTOR | mammalian target of the rapamycin |
| TMA | tissue microarray |
| DAB | 3,3′-diaminobenzidine |
| RIPA | radioimmunoprecipitation assay |
| PMSF | phenylmethylsulfonyl fluoride |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| PVDF | polyvinylidene difluoride |
| BCL-2 | B-cell lymphoma-2 |
| BAX | BCL-2-Associated X |
| pAMPK | phospho-AMPK |
References
- Canipari, R.; De Santis, L.; Cecconi, S. Female Fertility and Environmental Pollution. Int. J. Environ. Res. Public Health 2020, 17, 8802. [Google Scholar] [CrossRef] [PubMed]
- Sayers, N.S.; Anujan, P.; Yu, H.N.; Palmer, S.S.; Nautiyal, J.; Franks, S.; Hanyaloglu, A.C. Follicle-Stimulating Hormone Induces Lipid Droplets via Galphai/o and beta-Arrestin in an Endometrial Cancer Cell Line. Front. Endocrinol. 2021, 12, 798866. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Hwang, I.S.; Jung, G.; Kang, H.J.; Chung, Y.H. Relationship between metabolic syndrome and follicle-stimulating hormone in postmenopausal women. Medicine 2022, 101, e29216. [Google Scholar] [CrossRef]
- Mallozzi, M.; Leone, C.; Manurita, F.; Bellati, F.; Caserta, D. Endocrine Disrupting Chemicals and Endometrial Cancer: An Overview of Recent Laboratory Evidence and Epidemiological Studies. Int. J. Environ. Res. Public Health 2017, 14, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raglan, O.; Kalliala, I.; Markozannes, G.; Cividini, S.; Gunter, M.J.; Nautiyal, J.; Gabra, H.; Paraskevaidis, E.; Martin-Hirsch, P.; Tsilidis, K.K.; et al. Risk factors for endometrial cancer: An umbrella review of the literature. Int. J. Cancer 2019, 145, 1719–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumph, J.T.; Stephens, V.R.; Martin, J.L.; Brown, L.K.; Thomas, P.L.; Cooley, A.; Osteen, K.G.; Bruner-Tran, K.L. Uncovering Evidence: Associations between Environmental Contaminants and Disparities in Women’s Health. Int. J. Environ. Res. Public Health 2022, 19, 1257. [Google Scholar] [CrossRef]
- Rutkowska, A.Z.; Szybiak, A.; Serkies, K.; Rachon, D. Endocrine disrupting chemicals as potential risk factor for estrogen-dependent cancers. Pol. Arch. Med. Wewn. 2016, 126, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Hlisnikova, H.; Petrovicova, I.; Kolena, B.; Sidlovska, M.; Sirotkin, A. Effects and Mechanisms of Phthalates’ Action on Reproductive Processes and Reproductive Health: A Literature Review. Int. J. Environ. Res. Public Health 2020, 17, 6811. [Google Scholar] [CrossRef]
- Harlow, S.D.; Hood, M.M.; Ding, N.; Mukherjee, B.; Calafat, A.M.; Randolph, J.F.; Gold, E.B.; Park, S.K. Per- and Polyfluoroalkyl Substances and Hormone Levels During the Menopausal Transition. J. Clin. Endocrinol. Metab. 2021, 106, e4427–e4437. [Google Scholar] [CrossRef]
- Crosbie, E.J.; Kitson, S.J.; McAlpine, J.N.; Mukhopadhyay, A.; Powell, M.E.; Singh, N. Endometrial cancer. Lancet 2022, 399, 1412–1428. [Google Scholar] [CrossRef]
- Gu, B.; Shang, X.; Yan, M.; Li, X.; Wang, W.; Wang, Q.; Zhang, C. Variations in incidence and mortality rates of endometrial cancer at the global, regional, and national levels, 1990–2019. Gynecol. Oncol. 2021, 161, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Makker, V.; Taylor, M.H.; Aghajanian, C.; Oaknin, A.; Mier, J.; Cohn, A.L.; Romeo, M.; Bratos, R.; Brose, M.S.; DiSimone, C.; et al. Lenvatinib Plus Pembrolizumab in Patients With Advanced Endometrial Cancer. J. Clin. Oncol. 2020, 38, 2981–2992. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Sun, C. Association of abnormal glucose metabolism and insulin resistance in patients with atypical and typical endometrial cancer. Oncol. Lett. 2018, 15, 2173–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.Q.; Fei, H.L.; Zhu, H.L. Bu Shen Yang Xue Prescription Has Treating Effect on Endometrial Cancer through FSH/PI3K/AKT/Gankyrin/HIF-alpha/cyclinD 1 Pathway in Ishikawa Cells. Evid. Based Complement Alternat. Med. 2018, 2018, 8412984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.C.; Peng, D.; Cai, Z.; Lin, H.K. AMPK Signaling and Its Targeting in Cancer Progression and Treatment. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Ghose, A.; Gullapalli, S.V.N.; Chohan, N.; Bolina, A.; Moschetta, M.; Rassy, E.; Boussios, S. Applications of Proteomics in Ovarian Cancer: Dawn of a New Era. Proteomes 2022, 10, 16. [Google Scholar] [CrossRef]
- Conza, D.; Mirra, P.; Cali, G.; Insabato, L.; Fiory, F.; Beguinot, F.; Ulianich, L. Metformin Dysregulates the Unfolded Protein Response and the WNT/beta-Catenin Pathway in Endometrial Cancer Cells through an AMPK-Independent Mechanism. Cells 2021, 10, 1067. [Google Scholar] [CrossRef]
- Rho, S.B.; Byun, H.J.; Kim, B.R.; Lee, C.H. Knockdown of LKB1 Sensitizes Endometrial Cancer Cells via AMPK Activation. Biomol. Ther. 2021, 29, 650–657. [Google Scholar] [CrossRef]
- Rice, S.; Elia, A.; Jawad, Z.; Pellatt, L.; Mason, H.D. Metformin inhibits follicle-stimulating hormone (FSH) action in human granulosa cells: Relevance to polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2013, 98, E1491–E1500. [Google Scholar] [CrossRef]
- Onizuka, Y.; Nagai, K.; Ideno, Y.; Kitahara, Y.; Iwase, A.; Yasui, T.; Nakajima-Shimada, J.; Hayashi, K. Association between FSH, E1, and E2 levels in urine and serum in premenopausal and postmenopausal women. Clin. Biochem. 2019, 73, 105–108. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, L.; Sun, X. CircRNA hsa_circ_0002577 accelerates endometrial cancer progression through activating IGF1R/PI3K/Akt pathway. J. Exp. Clin. Cancer Res. 2020, 39, 169. [Google Scholar] [CrossRef]
- Shi, X.; Qiu, S.; Zhuang, W.; Wang, C.; Zhang, S.; Yuan, N.; Yuan, F.; Qiao, Y. Follicle-stimulating hormone inhibits cervical cancer via NF-kappaB pathway. Onco. Targets Ther. 2018, 11, 8107–8115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zisis, T.; Bruckner, D.B.; Brandstatter, T.; Siow, W.X.; d’Alessandro, J.; Vollmar, A.M.; Broedersz, C.P.; Zahler, S. Disentangling cadherin-mediated cell-cell interactions in collective cancer cell migration. Biophys. J. 2022, 121, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Guo, Y.; Song, Y.; Yu, C.; Zhao, L.; Fang, L.; Kong, D.; Zhao, J.; Gao, L. Follicle-stimulating hormone enhances hepatic gluconeogenesis by GRK2-mediated AMPK hyperphosphorylation at Ser485 in mice. Diabetologia 2018, 61, 1180–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Zhao, M.; Bo, T.; Ma, S.; Yuan, Z.; Chen, W.; He, Z.; Hou, X.; Liu, J.; Zhang, Z.; et al. Blocking FSH inhibits hepatic cholesterol biosynthesis and reduces serum cholesterol. Cell Res. 2019, 29, 151–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhartiya, D.; Patel, H. An overview of FSH-FSHR biology and explaining the existing conundrums. J. Ovarian Res. 2021, 14, 144. [Google Scholar] [CrossRef] [PubMed]
- Chrusciel, M.; Ponikwicka-Tyszko, D.; Wolczynski, S.; Huhtaniemi, I.; Rahman, N.A. Extragonadal FSHR Expression and Function-Is It Real? Front. Endocrinol. 2019, 10, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skok, K.; Maver, U.; Gradisnik, L.; Kozar, N.; Takac, I.; Arko, D. Endometrial cancer and its cell lines. Mol. Biol. Rep. 2020, 47, 1399–1411. [Google Scholar] [CrossRef]
- Ghinea, N. Vascular Endothelial FSH Receptor, a Target of Interest for Cancer Therapy. Endocrinology 2018, 159, 3268–3274. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Dai, L.; Long, X.; Wang, W.; Di, W. Follicle-stimulating hormone promotes the proliferation of epithelial ovarian cancer cells by activating sphingosine kinase. Sci. Rep. 2020, 10, 13834. [Google Scholar] [CrossRef]
- Stilley, J.A.; Christensen, D.E.; Dahlem, K.B.; Guan, R.; Santillan, D.A.; England, S.K.; Al-Hendy, A.; Kirby, P.A.; Segaloff, D.L. FSH receptor (FSHR) expression in human extragonadal reproductive tissues and the developing placenta, and the impact of its deletion on pregnancy in mice. Biol. Reprod. 2014, 91, 74. [Google Scholar] [CrossRef]
- Gu, H.; Li, Y.; Cui, X.; Cao, H.; Hou, Z.; Ti, Y.; Liu, D.; Gao, J.; Wang, Y.; Wen, P. MICAL1 inhibits colorectal cancer cell migration and proliferation by regulating the EGR1/beta-catenin signaling pathway. Biochem. Pharmacol. 2022, 195, 114870. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Ladilov, Y. Emerging Role of cAMP/AMPK Signaling. Cells 2022, 11, 308. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Zhou, M.; Chen, C.; Wu, X.; Wang, X. Role of AMPK mediated pathways in autophagy and aging. Biochimie 2022, 195, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhang, L.; Guo, H.; Wysham, W.Z.; Roque, D.R.; Willson, A.K.; Sheng, X.; Zhou, C.; Bae-Jump, V.L. Glucose promotes cell proliferation, glucose uptake and invasion in endometrial cancer cells via AMPK/mTOR/S6 and MAPK signaling. Gynecol. Oncol. 2015, 138, 668–675. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Li, X.W.; Chen, Z.; Hao, F.; Tao, S.X.; Yu, H.Y.; Cheng, R.; Liu, H. Neuro-Protective Role of Metformin in Patients with Acute Stroke and Type 2 Diabetes Mellitus via AMPK/Mammalian Target of Rapamycin (mTOR) Signaling Pathway and Oxidative Stress. Med. Sci. Monit. 2019, 25, 2186–2194. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, W.; Zhang, J.; You, Z.; Hu, T.; Shao, G.; Zhang, Z.; Xu, Z.; Yu, X. TWIST2 inhibits EMT and induces oxidative stress in lung cancer cells by regulating the FGF21-mediated AMPK/mTOR pathway. Exp. Cell Res. 2021, 405, 112661. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Patients | 34 |
|---|---|
| Median age (years) | 60 |
| Age range (years) | 35–80 |
| Pathology Diagnosis | |
| Endometrioid adenocarcinoma | 30 |
| Endometrial clear cell carcinoma | 1 |
| Endometrioid adenocarcinoma with squamous metaplasia | 3 |
| Grade | |
| Grade 1 | 4 |
| Grade 2 | 17 |
| Grade 3 | 13 |
| FIGO Stage | |
| IA | 30 |
| IB | 2 |
| II | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, S.; Liu, W.; Xue, Y.; Pan, Z.; Zhao, L.; Wang, F.; Qi, X. Follicle-Stimulating Hormone Promotes the Development of Endometrial Cancer In Vitro and In Vivo. Int. J. Environ. Res. Public Health 2022, 19, 15344. https://doi.org/10.3390/ijerph192215344
Sheng S, Liu W, Xue Y, Pan Z, Zhao L, Wang F, Qi X. Follicle-Stimulating Hormone Promotes the Development of Endometrial Cancer In Vitro and In Vivo. International Journal of Environmental Research and Public Health. 2022; 19(22):15344. https://doi.org/10.3390/ijerph192215344
Chicago/Turabian StyleSheng, Shuman, Wei Liu, Yafei Xue, Zhengwu Pan, Lanlan Zhao, Fei Wang, and Xiaoyi Qi. 2022. "Follicle-Stimulating Hormone Promotes the Development of Endometrial Cancer In Vitro and In Vivo" International Journal of Environmental Research and Public Health 19, no. 22: 15344. https://doi.org/10.3390/ijerph192215344