
The Effect of the Distance from a Path on Abiotic Conditions and Vascular Plant Species in the Undergrowth of Urban Forests and Parks


Abstract
:1. Introduction
2. Materials and Methods
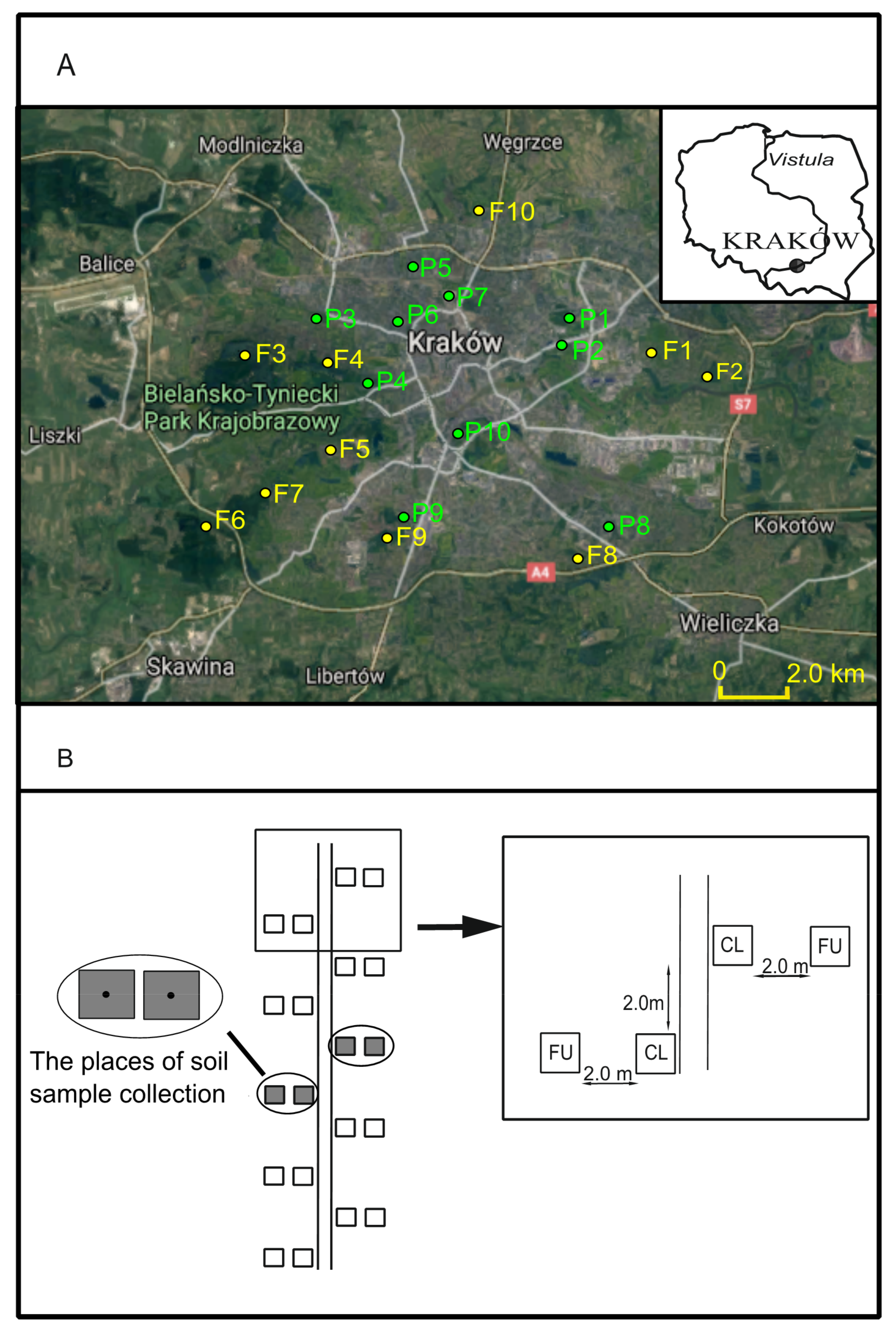
2.1. Study Area
2.2. Plot Sampling Design
2.3. Measurement of Abiotic Traits within the Plots
2.4. Measurement of Vegetation Cover within the Plots
- “+”—species covers less than 1% of the plot area,
- “1”—species covers 1–5% of the plot area,
- “2”—species covers 6–25% of the plot area,
- “3”—species covers 26–50% of the plot area,
- “4”—species covers 51–75% of the plot area,
- “5”—species covers 76–100% of the plot area,
2.5. Selection of Ecological Traits of the Species
2.6. Statistical Analyses
3. Results
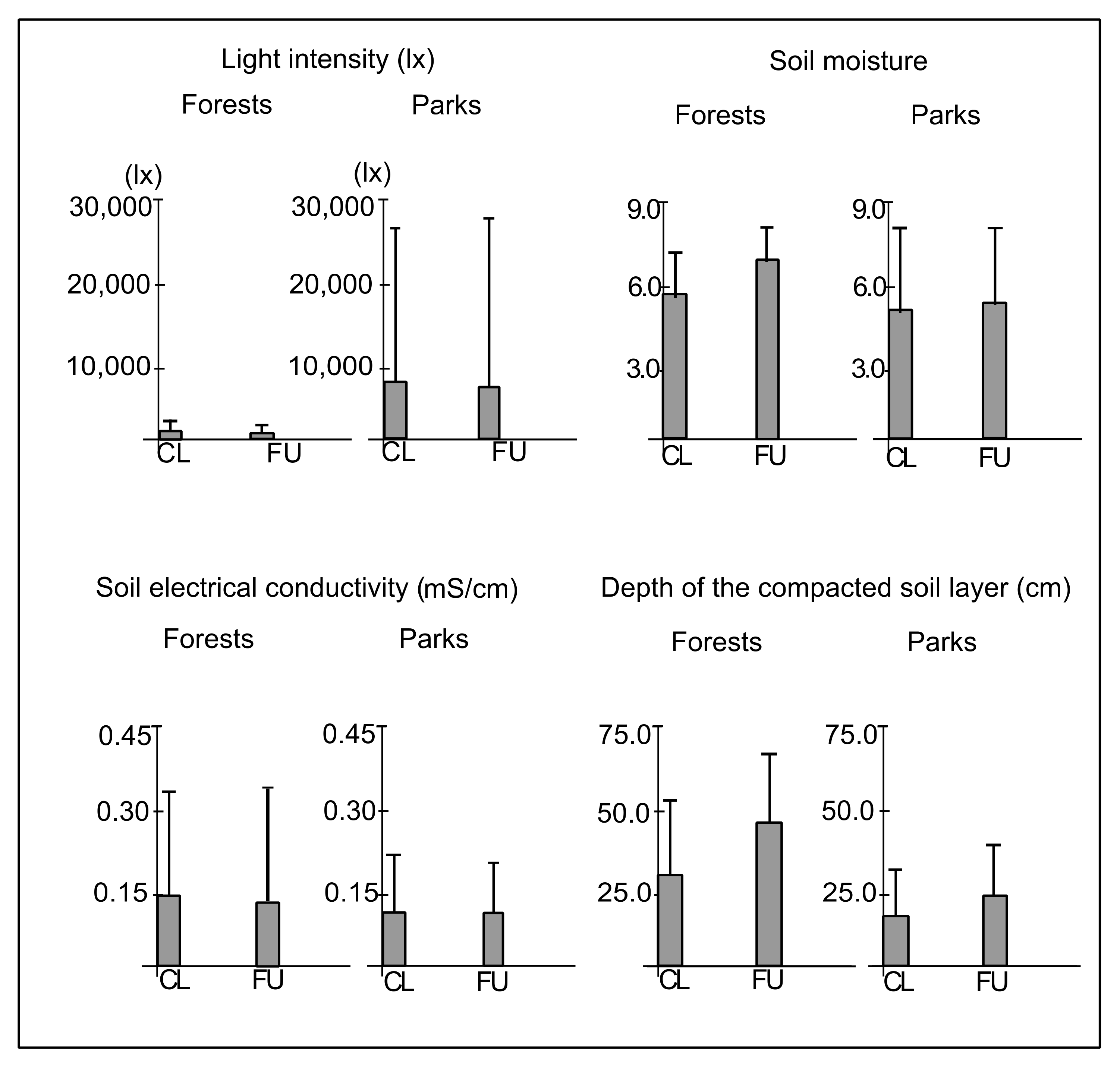
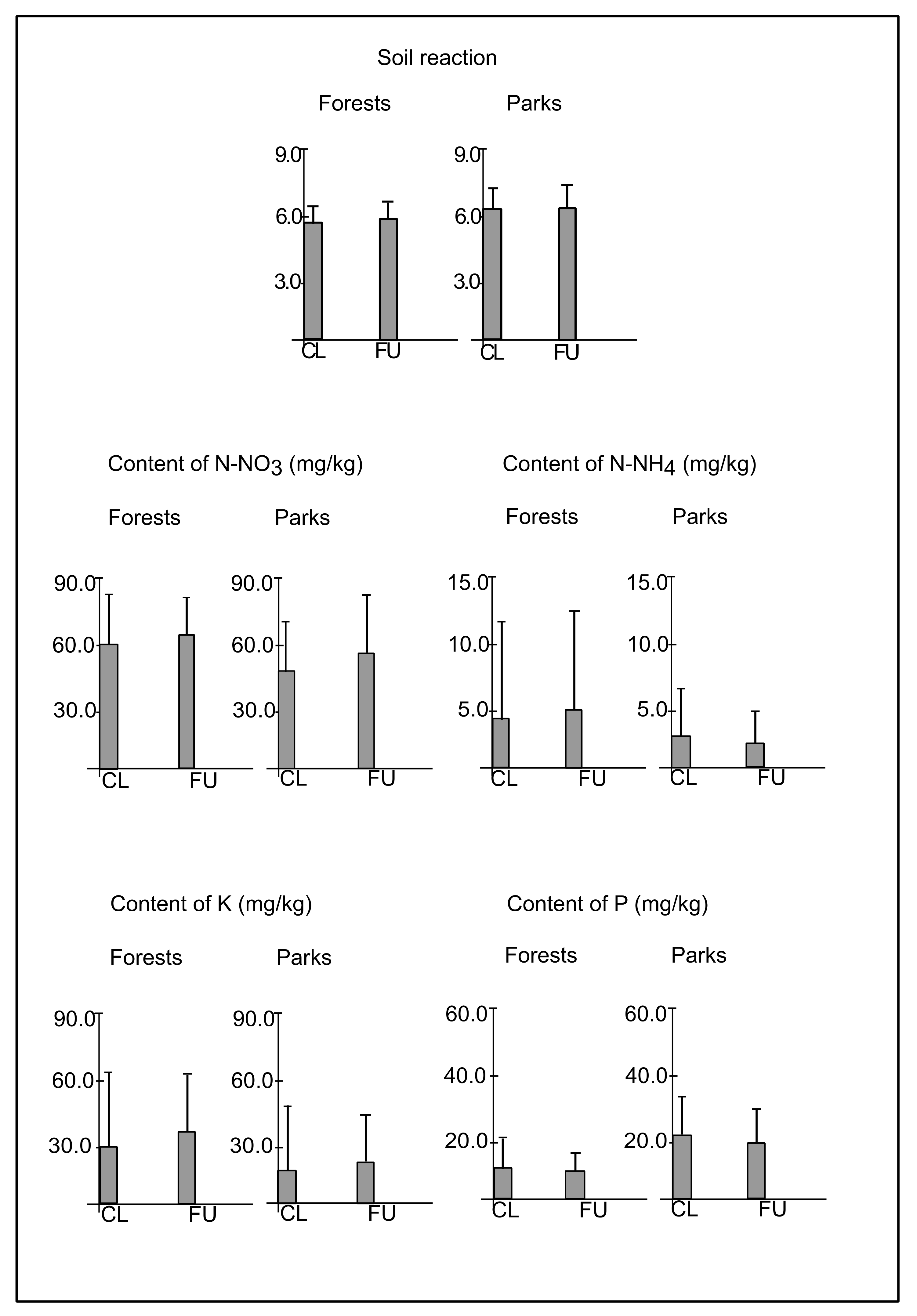
3.1. Light Intensity and Soil Conditions
3.2. Vegetation Cover Traits and Number of Species
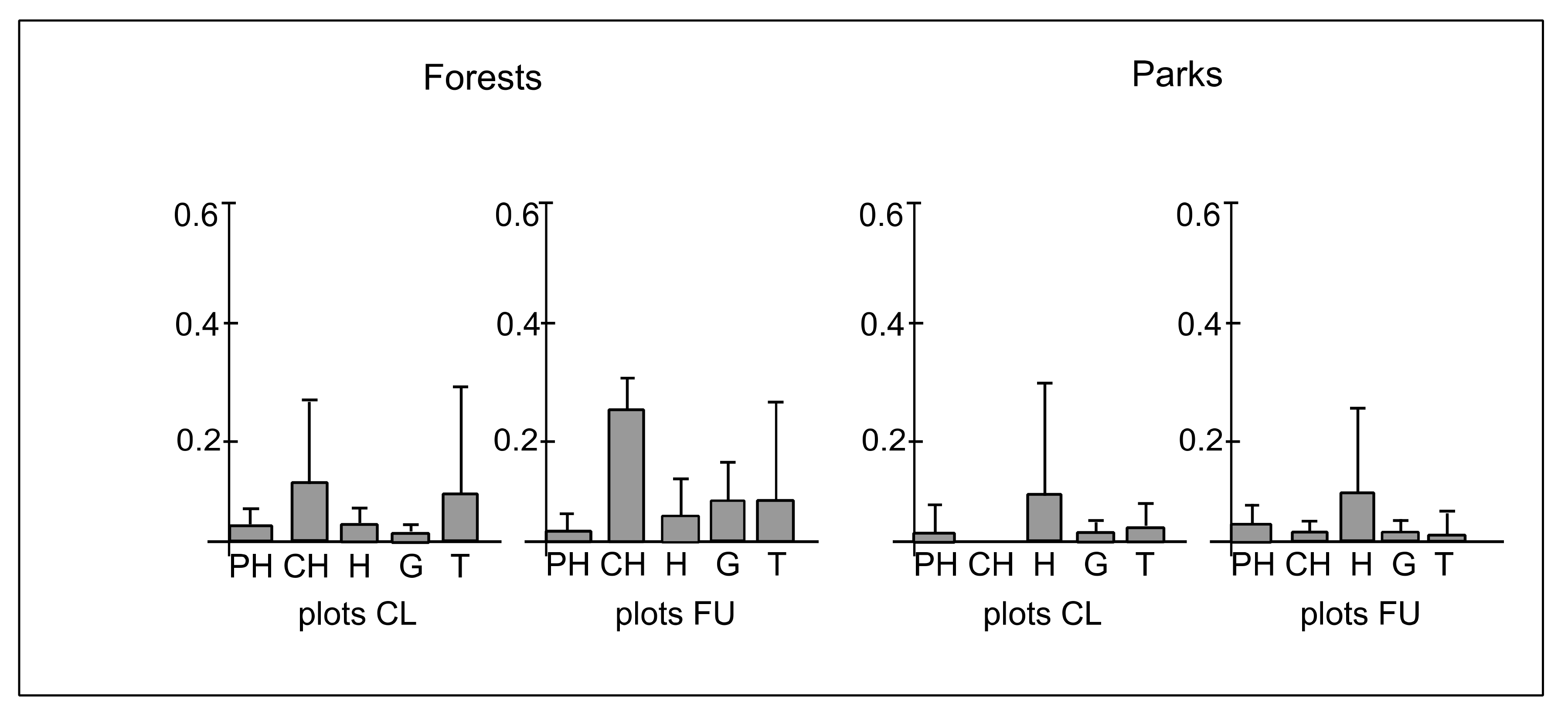
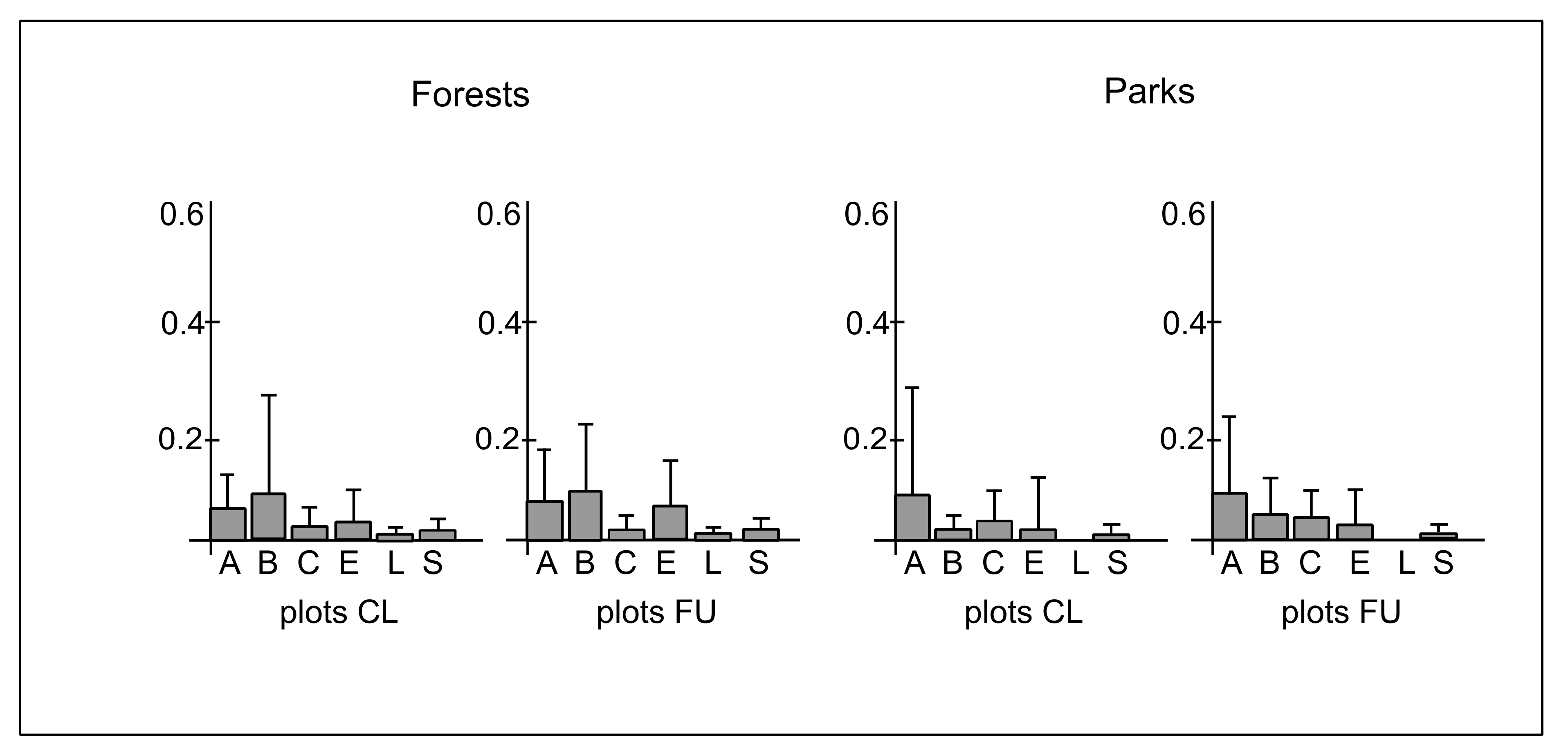
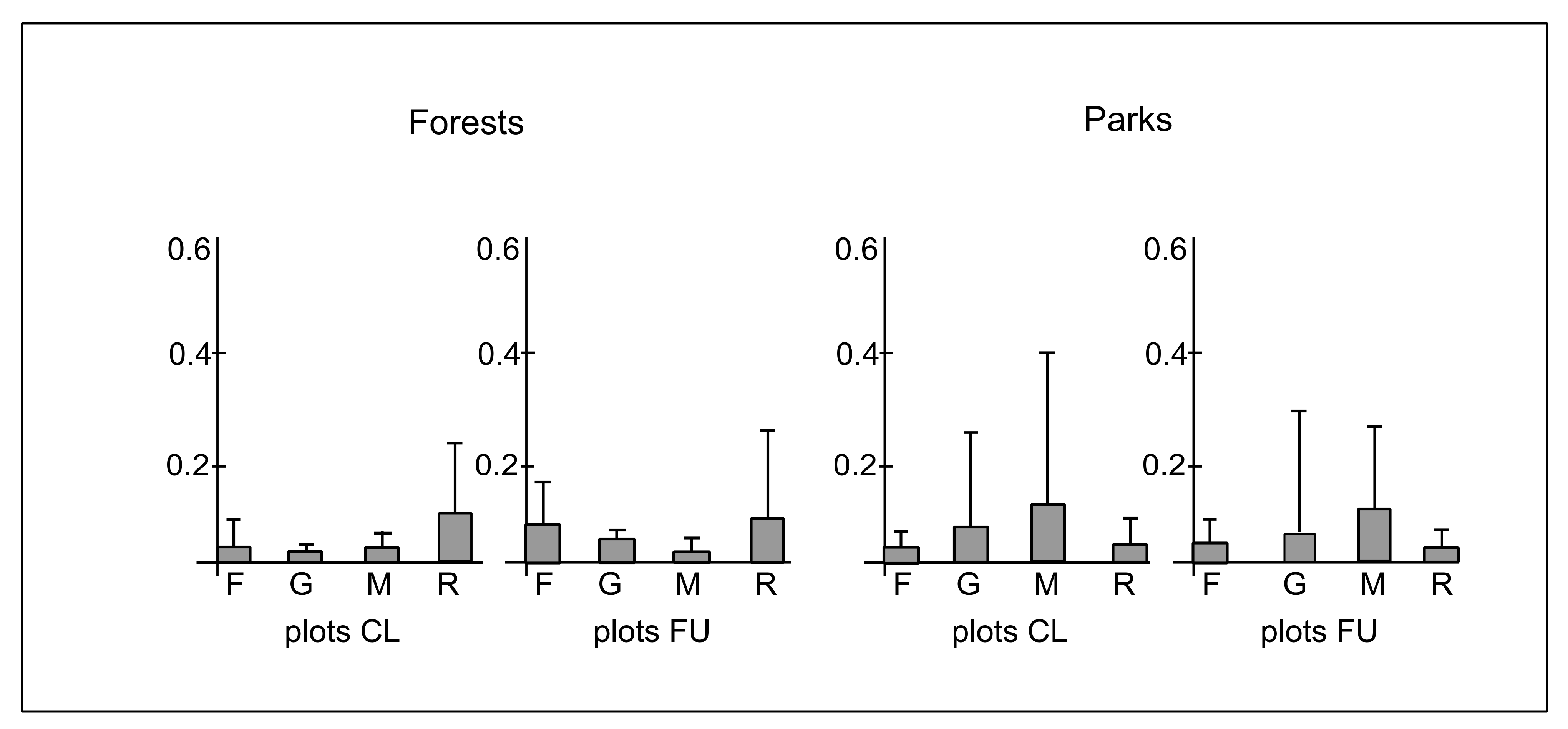
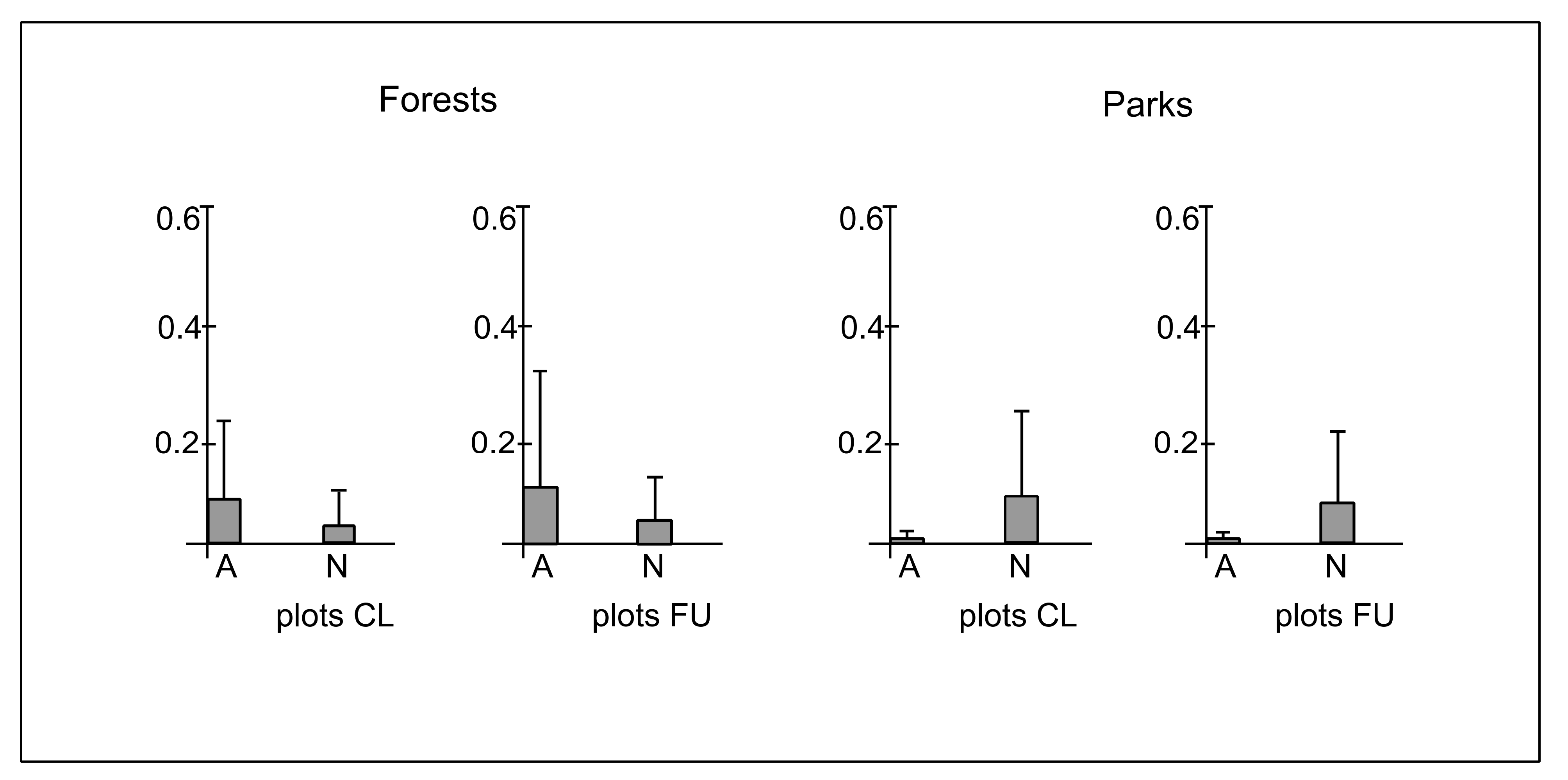
3.3. Ecological Characteristics of Species
4. Discussion
4.1. The Effect of Distance from the Path on Abiotic Conditions
4.2. The Effect of Distance from the Path on Vegetation Cover and Number of Species
4.3. The Effect of Distance from the Path on Ecological Diversity and Abundance of Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon (Speciesorgenus) | Presence or Absencein Plots | Life Form | Dispersalmode | Habitat Affiliation | Origin | |||
|---|---|---|---|---|---|---|---|---|
| Forests | Parks | |||||||
| CL | FU | CL | FU | |||||
| Acer platanoides L. | 1 | 1 | 1 | 1 | PH | Epilobium | Forest | Native |
| Acer pseudoplatanus L. | 1 | 1 | 1 | 1 | PH | Epilobium | Forest | Native |
| Acer saccharinum L. | 0 | 0 | 0 | 1 | PH | Epilobium | Ruderal | Alien |
| Achillea millefolium L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Actaea spicata L. | 1 | 0 | 0 | 0 | H | Cornus | Forest | Native |
| Aegopodium podagraria L. | 1 | 1 | 1 | 1 | H | Allium | Forest | Native |
| Agrimonia eupatoria L. | 1 | 0 | 1 | 1 | H | Bidens | Grassland | Native |
| Agrostis capillaris L. | 1 | 1 | 1 | 1 | H | Allium | Grassland | Native |
| Ajuga reptans L. | 1 | 0 | 0 | 0 | H | Allium | Forest | Native |
| Alchemilla micans Buser | 1 | 0 | 0 | 1 | H | Allium | Meadow | Native |
| Alliaria petiolata (M. Bieb.) Cavara& Grande | 1 | 1 | 1 | 1 | H | Allium | Ruderal | Native |
| Alnus glutinosa (L.) Gaertn. | 0 | 1 | 0 | 0 | PH | Epilobium | Forest | Native |
| Anemone nemorosa L. | 1 | 1 | 0 | 1 | G | Allium | Forest | Native |
| Arctium tomentosum Mill. | 1 | 0 | 0 | 0 | H | Bidens | Ruderal | Native |
| Arenaria serpyllifolia L. | 0 | 0 | 1 | 1 | T | Allium | Grassland | Native |
| Arrhenatherum elatius (L.) P. Beauv. ex J. Presl& C. Presl | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Artemisia vulgaris L. | 0 | 0 | 0 | 1 | H | Allium | Ruderal | Native |
| Asarum europaeum L. | 1 | 1 | 0 | 0 | H | Allium | Forest | Native |
| Asperula odorata L. | 1 | 1 | 0 | 0 | H | Bidens | Forest | Native |
| Atriplex patula L. | 0 | 1 | 1 | 1 | T | Allium | Ruderal | Native |
| Avenula pubescens (Huds.) Dumort. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Bellis perennis L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Berteroa incana (L.) DC. | 0 | 0 | 1 | 0 | T | Allium | Ruderal | ? |
| Bidens frondosa L. | 1 | 0 | 0 | 0 | T | Bidens | Ruderal | Alien * |
| Bromus hordeaceus L. | 0 | 0 | 1 | 0 | T | Allium | Meadow | Native |
| Bromus sterilis L. | 0 | 0 | 0 | 1 | T | Allium | Ruderal | Alien |
| Calystegia sepium (L.) R. Br. | 0 | 0 | 1 | 0 | G | Allium | Ruderal | Native |
| Capsella bursa-pastoris (L.) Med. | 0 | 0 | 1 | 1 | T | Allium | Ruderal | Alien |
| Carex brizoides L. | 1 | 1 | 0 | 0 | G | Allium | Forest | Native |
| Carex hirta L. | 0 | 0 | 1 | 1 | G | Allium | Meadow | Native |
| Carex ovalis Good. | 1 | 1 | 0 | 1 | H | Allium | Meadow | Native |
| Carex remota L. | 1 | 1 | 0 | 0 | H | Sparganium | Forest | Native |
| Carex sp. | 1 | 1 | 1 | 1 | nd | nd | nd | nd |
| Carex spicata Huds. | 0 | 0 | 1 | 1 | H | Allium | Grassland | Native |
| Carex sylvatica Huds. | 1 | 0 | 0 | 0 | H | Allium | Forest | Native |
| Carpinus betulus L. | 1 | 1 | 1 | 1 | PH | Epilobium | Forest | Native |
| Cerastium arvense L. | 0 | 0 | 1 | 1 | CH | Allium | Ruderal | Native |
| Cerastium holosteoides Fr. | 0 | 0 | 1 | 1 | CH | Allium | Meadow | Native |
| Cerasus avium (L.) Moench | 1 | 1 | 0 | 0 | PH | Cornus | Forest | Native |
| Chaerophyllum aromaticum L. | 1 | 0 | 1 | 1 | H | Allium | Ruderal | Native |
| Chaerophyllum temulum L. | 0 | 0 | 1 | 0 | H | Allium | Ruderal | Native |
| Chelidonium majus L. | 1 | 1 | 0 | 1 | H | Allium | Ruderal | Native |
| Circaea lutetiana L. | 1 | 1 | 0 | 0 | G | Bidens | Forest | Native |
| Cirsium vulgare (Savi) Ten. | 0 | 0 | 1 | 1 | H | Epilobium | Ruderal | ? |
| Convallaria majalis L. | 1 | 1 | 0 | 0 | G | Cornus | Forest | Native |
| Convolvulus arvensis L. | 0 | 0 | 1 | 1 | G | Allium | Ruderal | Native |
| Conyza canadensis(L.) Cronquist | 0 | 0 | 1 | 0 | T | Epilobium | Ruderal | Alien * |
| Cornus sanguinea L. | 1 | 1 | 1 | 1 | PH | Cornus | Grassland | Native |
| Coronilla varia L. | 0 | 0 | 1 | 1 | H | Allium | Grassland | Native |
| Corylus avellana L. | 1 | 0 | 0 | 0 | PH | Cornus | Forest | Native |
| Crataegus sp. | 1 | 1 | 1 | 1 | nd | nd | nd | nd |
| Crepis biennis L. | 0 | 0 | 1 | 1 | H | Epilobium | Meadow | Native |
| Dactylis glomerata L. | 1 | 0 | 1 | 1 | nd | Allium | Meadow | Native |
| Deschampsia cespitosa (L.) P. Beauv. | 1 | 1 | 0 | 1 | H | Allium | Meadow | Native |
| Dryopteris sp. | 0 | 1 | 0 | 0 | nd | nd | nd | nd |
| Dryopteris carthusiana (Vill.) H. P. Fuchs | 0 | 1 | 0 | 0 | H | Lycopodium | Forest | Native |
| Duchesnea indica (Andrews) Focke | 0 | 0 | 1 | 1 | H | Cornus | Ruderal | Alien |
| Echium vulgare L. | 0 | 0 | 1 | 0 | H | Allium | Ruderal | Native |
| Elymus repens (L.) Gould | 1 | 0 | 1 | 1 | G | Allium | Ruderal | Native |
| Epilobium montanum L. | 0 | 0 | 1 | 0 | H | Epilobium | Ruderal | Native |
| Equisetum arvense L. | 1 | 0 | 0 | 0 | G | Lycopodium | Ruderal | Native |
| Erigeron annuus (L.) Pers. | 0 | 0 | 0 | 1 | H | Epilobium | Ruderal | Alien * |
| Euonymus europaeus L. | 1 | 1 | 0 | 1 | PH | Cornus | Forest | Native |
| Euonymus verrucosus Scop. | 1 | 1 | 0 | 0 | PH | Cornus | Forest | Native |
| Euphorbia cyparissias L. | 0 | 0 | 1 | 0 | G | Allium | Grassland | Native |
| Euphorbia peplus L. | 0 | 0 | 0 | 1 | T | Allium | Ruderal | Alien |
| Fagus sylvatica L. | 1 | 1 | 0 | 0 | PH | Cornus | Forest | Native |
| Fallopia convolvulus (L.) Å. Löve | 0 | 0 | 0 | 1 | T | Allium | Ruderal | Alien |
| Festuca arundinacea Schreb. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Festuca gigantea (L.) Vill. | 1 | 0 | 0 | 1 | H | Allium | Forest | Native |
| Festuca rubra L. | 1 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Filipendula ulmaria (L.) Maxim. | 1 | 1 | 0 | 0 | H | Allium | Meadow | Native |
| Fragaria viridis (Duchesne) Weston | 1 | 0 | 1 | 1 | H | Cornus | Grassland | Native |
| Fraxinus excelsior L. | 1 | 1 | 1 | 1 | PH | Epilobium | Forest | Native |
| Galeobdolon luteum Huds. | 1 | 1 | 0 | 0 | CH | Allium | Forest | Native |
| Galeopsis tetrahit L. | 1 | 1 | 0 | 0 | T | Allium | Ruderal | Native |
| Galinsoga ciliata (Raf.) S. F. Blake | 0 | 0 | 0 | 1 | T | Epilobium | Ruderal | Alien * |
| Galium aparine L. | 0 | 1 | 0 | 1 | T | Bidens | Ruderal | Native |
| Galium mollugo L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Galium odoratum (L.) Scop. | 1 | 1 | 0 | 0 | H | Bidens | Forest | Native |
| Geranium phaeum L. | 1 | 1 | 0 | 0 | H | Allium | Forest | Native |
| Geranium pratense L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Geranium pusillum Burm. F. ex L | 0 | 0 | 1 | 0 | T | Allium | Ruderal | Alien |
| Geranium robertianum L. | 1 | 1 | 0 | 1 | T | Allium | Ruderal | Native |
| Geum urbanum L. | 1 | 1 | 1 | 1 | H | Bidens | Ruderal | Native |
| Glechoma hederacea L. | 1 | 1 | 1 | 1 | H | Allium | Ruderal | Native |
| Hedera helix L. | 1 | 1 | 1 | 1 | PH | Cornus | Forest | Native |
| Heracleum sphondylium L. | 0 | 0 | 0 | 1 | H | Allium | Meadow | Native |
| Hieracium pilosella L. | 0 | 0 | 0 | 1 | H | Epilobium | Grassland | Native |
| Holcus lanatus L. | 0 | 0 | 0 | 1 | H | Allium | Meadow | Native |
| Humulus lupulus L. | 0 | 1 | 0 | 1 | H | Allium | Forest | Native |
| Impatiens glandulifera Royle | 0 | 1 | 0 | 0 | T | Allium | Ruderal | Alien * |
| Impatiens parviflora DC. | 1 | 1 | 0 | 1 | T | Allium | Ruderal | Alien * |
| Juglans regia L. | 1 | 0 | 0 | 0 | PH | Cornus | Ruderal | Alien * |
| Juncus bufonius L. | 0 | 0 | 1 | 0 | T | Sparganium | Ruderal | Native |
| Juncus tenuis Willd. | 0 | 0 | 1 | 0 | H | Allium | Meadow | Alien * |
| Lactuca serriola L. | 0 | 0 | 1 | 1 | H | Epilobium | Ruderal | Alien |
| Lamium album L. | 0 | 0 | 1 | 1 | H | Allium | Ruderal | Alien |
| Lamium purpureum L. | 0 | 0 | 1 | 0 | T | Allium | Ruderal | Alien |
| Lapsana communis L. | 0 | 0 | 1 | 0 | T | Allium | Ruderal | Native |
| Lathyrus vernus (L.) Bernh. | 1 | 0 | 0 | 0 | G | Allium | Forest | Native |
| Leontodon autumnalis L. | 0 | 0 | 1 | 0 | H | Epilobium | Meadow | Native |
| Leontodon hispidus L. | 0 | 0 | 0 | 1 | H | Epilobium | Meadow | Native |
| Ligustrum vulgare L. | 0 | 1 | 0 | 0 | PH | Cornus | Grassland | ? |
| Lolium perenne L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Lonicera xylosteum L. | 1 | 0 | 0 | 0 | PH | Cornus | Forest | Native |
| Lysimachia nummularia L. | 1 | 1 | 1 | 1 | H | Allium | Meadow | Native |
| Lysimachia vulgaris L. | 1 | 0 | 0 | 1 | G | Allium | Meadow | Native |
| Maianthemum bifolium (L.) F. W. Schmidt | 1 | 1 | 0 | 0 | G | Cornus | Forest | Native |
| Medicago lupulina L. | 0 | 0 | 1 | 1 | T | Allium | Ruderal | Native |
| Medicago falcata L | 0 | 0 | 1 | 1 | H | Allium | Grassland | Native |
| Melandrium album (Mill.) Garcke | 0 | 0 | 1 | 0 | H | Allium | Ruderal | Alien |
| Melica nutans L. | 1 | 0 | 0 | 0 | H | Allium | Forest | Native |
| Mentha sp. | 0 | 0 | 1 | 0 | nd | nd | nd | nd |
| Mercurialis perennis L. | 1 | 1 | 0 | 0 | G | Allium | Forest | Native |
| Milium effusum L. | 1 | 1 | 0 | 0 | H | Allium | Forest | Native |
| Moehringia trinervia (L.) Clairv. | 0 | 1 | 0 | 0 | H | Allium | Ruderal | Native |
| Myosotis arvensis (L.) Hill | 0 | 0 | 0 | 1 | T | Allium | Ruderal | Alien |
| Oxalis acetosella L. | 1 | 1 | 0 | 0 | G | Allium | Forest | Native |
| Oxalis fontana Bunge | 0 | 0 | 1 | 1 | G | Allium | Ruderal | Alien * |
| Padus avium Mill. | 1 | 1 | 0 | 0 | PH | Cornus | Forest | Native |
| Padus serotina (Ehrh.) Borkh. | 1 | 1 | 0 | 1 | PH | Cornus | Forest | Alien * |
| Paris quadrifolia L. | 1 | 0 | 0 | 0 | G | Cornus | Forest | Native |
| Plantago lanceolata L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Plantago major L. | 1 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Poa annua L. | 1 | 1 | 1 | 1 | H | Allium | Meadow | Native |
| Poa nemoralis L. | 1 | 0 | 0 | 0 | H | Allium | Forest | Native |
| Poa pratensis L. | 0 | 1 | 1 | 1 | H | Allium | Meadow | Native |
| Poa trivialis L. | 1 | 1 | 1 | 1 | H | Allium | Meadow | Native |
| Polygonatum multiflorum (L.) All. | 1 | 1 | 0 | 0 | G | Cornus | Meadow | Native |
| Polygonatum odoratum (Mill.) Druce | 1 | 0 | 0 | 0 | G | Cornus | Grassland | Native |
| Polygonum aviculare L. | 0 | 0 | 1 | 1 | T | Allium | Ruderal | Native |
| Polygonum lapathifolium L. | 0 | 0 | 1 | 1 | T | Sparganium | Ruderal | Native |
| Potentilla anserina L. | 0 | 0 | 1 | 0 | H | Allium | Meadow | Native |
| Potentilla arenaria P. Gaertn., B. Mey. &Scherb | 0 | 0 | 1 | 1 | H | Allium | Grassland | Native |
| Potentilla argentea L. | 0 | 0 | 1 | 0 | H | Allium | Grassland | Native |
| Potentilla reptans L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Prunella vulgaris L. | 1 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Prunus domestica L. | 0 | 1 | 0 | 1 | PH | Cornus | Ruderal | Alien |
| Pteridium aquilinum (L.) Kuhn | 0 | 1 | 0 | 0 | G | Lycopodium | Forest | Native |
| Pulmonaria obscura Dumort. | 1 | 1 | 0 | 0 | H | Allium | Forest | Native |
| Quercus robur L. | 1 | 1 | 1 | 0 | PH | Cornus | Forest | Native |
| Quercus rubra L. | 1 | 1 | 1 | 1 | PH | Cornus | Forest | Alien * |
| Quercus petraea (Matt.) Liebl. | 0 | 1 | 0 | 0 | PH | Cornus | Forest | Native |
| Ranunculus acris L. | 1 | 0 | 0 | 1 | H | Allium | Meadow | Native |
| Ranunculus lanuginosus L. | 0 | 1 | 0 | 1 | H | Allium | Forest | Native |
| Ranunculus repens L. | 0 | 1 | 1 | 1 | H | Allium | Meadow | Native |
| Ribes uva-crispa L. | 1 | 1 | 0 | 0 | PH | Cornus | Forest | Native |
| Robinia pseudoacacia L. | 0 | 0 | 1 | 1 | PH | Allium | Forest | Alien * |
| Rubus caesius L. | 1 | 1 | 1 | 1 | PH | Cornus | Ruderal | Native |
| Rubus idaeus L. | 1 | 1 | 0 | 0 | PH | Cornus | Ruderal | Native |
| Rubus sp. | 1 | 1 | 0 | 1 | nd | nd | nd | nd |
| Rumex obtusifolius L. | 0 | 0 | 1 | 1 | H | Allium | Ruderal | Native |
| Rumex thyrsiflorus Fingerh. | 0 | 0 | 1 | 0 | H | Allium | Meadow | ? |
| Sagina procumbens L. | 0 | 0 | 1 | 0 | CH | Allium | Meadow | Native |
| Sambucus nigra L. | 1 | 1 | 0 | 1 | PH | Cornus | Ruderal | Native |
| Setaria viridis (L.) P. Beauv. | 0 | 0 | 1 | 0 | T | Bidens | Ruderal | Alien * |
| Silene nutans L. | 0 | 0 | 0 | 1 | H | Allium | Grassland | Native |
| Solidago canadensis L. | 0 | 0 | 1 | 0 | H | Epilobium | Ruderal | Alien * |
| Solidago gigantea Aiton | 1 | 0 | 0 | 1 | H | Epilobium | Ruderal | Alien * |
| Sorbus aucuparia L. | 1 | 0 | 0 | 0 | PH | Cornus | Forest | Native |
| Stachys sylvatica L. | 0 | 1 | 0 | 0 | H | Allium | Forest | Native |
| Stellaria graminea L. | 0 | 0 | 1 | 0 | H | Allium | Meadow | Native |
| Stellaria holostea L. | 1 | 0 | 0 | 1 | CH | Allium | Forest | Native |
| Stellaria media (L.) Vill. | 1 | 1 | 1 | 1 | T | Allium | Ruderal | Native |
| Symphytum tuberosum L. | 1 | 1 | 0 | 0 | H | Allium | Forest | Native |
| Taraxacum officinale F.H. Wiggers | 1 | 0 | 1 | 1 | H | Epilobium | Meadow | Native |
| Thymus serpyllum L. | 0 | 0 | 1 | 1 | CH | Allium | Grassland | Native |
| Tilia cordata Mill. | 0 | 1 | 0 | 1 | PH | Epilobium | Forest | Native |
| Tilia sp. | 1 | 1 | 0 | 0 | PH | nd | nd | nd |
| Trifolium pratense L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Trifolium repens L. | 0 | 0 | 1 | 1 | H | Allium | Meadow | Native |
| Tussilago farfara L. | 1 | 0 | 0 | 0 | H | Epilobium | Ruderal | Native |
| Ulmus laevis Pall. | 0 | 0 | 1 | 1 | PH | Epilobium | Forest | Native |
| Ulmus minor Mill. | 1 | 0 | 0 | 0 | PH | Epilobium | Forest | Native |
| Urtica dioica L. | 1 | 1 | 1 | 1 | H | Allium | Ruderal | Native |
| Veronica arvensis L. | 0 | 0 | 1 | 1 | T | Allium | Ruderal | Alien |
| Veronica chamaedrys L | 1 | 0 | 1 | 1 | H | Allium | Grassland | Native |
| Veronica serpyllifolia L. | 0 | 0 | 1 | 1 | H | Allium | Ruderal | Native |
| Viciatetrasperma (L.) Schreb. | 0 | 0 | 0 | 1 | T | Allium | Ruderal | Alien |
| Viola odorata L. | 1 | 0 | 1 | 1 | H | Allium | Ruderal | ? |
| Viola reichenbachiana Jord. ex Boreau | 1 | 1 | 0 | 1 | H | Allium | Forest | Native |
| Viola sp. | 0 | 0 | 1 | 0 | nd | nd | nd | nd |
References
- Dwyer, J.F.; McPherson, E.G.; Schroeder, H.W.; Rowntree, R.A. Assessing the benefits and costs of the urban forest. J. Arboric. 1992, 18, 227–234. [Google Scholar]
- Alvey, A.A. Promoting and preserving biodiversity in the urban forest. Urban For. Urban Green. 2006, 5, 195–201. [Google Scholar] [CrossRef]
- Nowak, D.J.; Crane, D.E.; Stevens, J.C. Air pollution removal by urban trees and shrubs in the United States. Urban For. Urban Green. 2006, 4, 115–123. [Google Scholar] [CrossRef]
- Loures, L.; Costa, L. The role of urban parks to enhance metropolitan sustainability: The case of Oporto. Int. J. Energy Environ. 2012, 6, 453–461. [Google Scholar]
- Yao, L.; Chen, L.; Wei, W.; Sun, R. Potential reduction in urban runoff by green spaces in Beijing: A scenario analysis. Urban For. Urban Green. 2015, 14, 300–308. [Google Scholar] [CrossRef]
- Livesley, S.J.; McPherson, E.G.; Calfapietra, C. The urban forest and ecosystem services: Impacts on urban water, heat, and pollution cycles at the tree, street, and city scale. J. Environ. Qual. 2016, 45, 119–124. [Google Scholar] [CrossRef]
- Wang, W.; Wang, H.; Xiao, L.; He, X.; Zhou, W.; Wang, Q.; Wei, C. Microclimate regulating functions of urban forests in Changchun City (north-east China) and their association with different factors. iForest 2018, 11, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Wilken, C. The Effect of Urban Forests on Air Quality and Human Health. Bachelor’s Thesis, University of Nebraska, Lincoln, NE, USA, 2021. [Google Scholar]
- Schroeder, H.W. Preferred features of urban forests and parks. J. Arboric. 1982, 8, 317–322. [Google Scholar]
- Chiesura, A. The role of urban parks for the sustainable city. Lands. Urban Plan. 2004, 68, 129–138. [Google Scholar] [CrossRef]
- Hajzeri, A. The management of urban parks and its contribution to social interactions. Arboric. J. 2020, 43, 187–195. [Google Scholar] [CrossRef]
- Nowak, D.J.; Stein, S.M.; Randler, P.B.; Greenfield, E.J.; Comas, S.J.; Carr, M.A.; Alig, R.J. Sustaining America’s Urban Trees and Forests: A Forests on the Edge Report; Gen. Tech. Rep. NRS-62; Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2010; p. 27. [Google Scholar]
- Referowska-Chodak, E. Pressures and threats to nature related to human activities in European urban and suburban forests. Forests 2019, 10, 765. [Google Scholar] [CrossRef] [Green Version]
- Sikorski, P.; Jackowiak, K.; Szumacher, I. Interdisciplinary environmental studies in urban parks as a basis for their sustainable management. Misc. Geogr. 2008, 13, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Ballantyne, M.; Pickering, C.M. The impacts of trail infrastructure on vegetation and soils: Current literature and future directions. J. Environ. Manag. 2015, 164, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McWilliam, W.; Eagles, P.; Seasons, M.; Brown, R. Assessing the degradation effects of local residents on urban forests in Ontario, Canada. Arboric. Urban For. 2010, 36, 253–260. [Google Scholar] [CrossRef]
- Tonnesen, A.S.; Ebersole, J.J. Human trampling effects on regeneration and age structures of Pinus edulis and Juniperus monosperma. Great Basin Nat. 1997, 57, 50–56. [Google Scholar]
- Sarah, P.; Zhevelev, H.M.; Oz, A. Urban park soil and vegetation: Effects of natural and anthropogenic factors. Pedosphere 2015, 25, 392–404. [Google Scholar] [CrossRef]
- Simpson, G.D.; Parker, J.; Gibbens, E.; Ladd, P.G. A hybrid method for citizen science monitoring of recreational trampling in urban remnants: A case study from Perth, Western Australia. Urban Sci. 2020, 4, 72. [Google Scholar] [CrossRef]
- Kostrakiewicz-Gierałt, K.; Pliszko, A.; Gmyrek, K. The effect of informal tourist trails on the abiotic conditions and floristic composition of deciduous forest undergrowth in an urban area. Forests 2021, 12, 423. [Google Scholar] [CrossRef]
- Recycling Council for British Columbia. Illegal Dumping in BC: An Overview. October 2017. Available online: https://www.rcbc.ca (accessed on 27 October 2021).
- Lee, J.M.; Tan, J.; Gill, A.S.; McGuire, K.L. Evaluating the effects of canine urine on urban soil microbial communities. Urban Ecosyst. 2019, 22, 721–732. [Google Scholar] [CrossRef]
- Allen, J.A.; Setälä, H.; Kotze, D.J. Dog urine has acute impacts on soil chemistry in urban green spaces. Front. Ecol. Evol. 2020, 8, 615979. [Google Scholar] [CrossRef]
- Badach, J.; Dymnicka, M.; Baranowski, A. Urban vegetation in air quality management: A review and policy framework. Sustainability 2020, 12, 1258. [Google Scholar] [CrossRef] [Green Version]
- Lõhmus, K.; Liira, J. Old rural parks support higher biodiversity than forest remnants. Basic. Appl. Ecol. 2013, 14, 165–173. [Google Scholar] [CrossRef]
- Bell, S.; Blom, D.; Rautamäki, M.; Castel-Branco, C.; Simson, A.; Olsen, I.A. Design of urban forests. In Urban Forests and Trees, a Reference Book; Konijnendijk, C.C., Nilsson, K., Randrup, T.B., Schipperijn, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 149–186. [Google Scholar]
- Kowarik, I.; Körner, S. (Eds.) Wild Urban Woodlands. New Perspectives for Urban Forestry; Springer: Berlin/Heidelberg, Germany, 2005; p. 287. [Google Scholar]
- LaPaix, R.; Freedman, B. Vegetation structure and composition within urban parks of Halifax Regional Municipality, Nova Scotia, Canada. Landsc. Urban Plan. 2010, 98, 124–135. [Google Scholar] [CrossRef]
- Afrianto, W.F.; Wati, S.I.; Hidayatullah, T. The suitability assessment of the tree species in the urban parks and urban forest in Kediri City, East Java, Indonesia. Nus. Biosci. 2021, 13, 131–139. [Google Scholar] [CrossRef]
- Almas, A.D.; Conway, T.M. The role of native species in urban forest planning and practice: A case study of Carolinian Canada. Urban For. Urban Green. 2016, 17, 54–62. [Google Scholar] [CrossRef]
- Rahmonov, O.; Pukowiec-Kurda, K.; Banaszek, J.; Brom, K. Floristic diversity in selected city parks in southern Poland. Environ. Nat. Resour. J. 2019, 30, 8–17. [Google Scholar] [CrossRef]
- Zhao, J.J.; Ouyang, Z.Y.; Zheng, H.; Xu, W.H.; Wang, X.K. Species composition and spatial structure of plants in urban parks of Beijing. Ying Yong Sheng Tai Xue Bao 2009, 20, 298–306. [Google Scholar]
- Wang, J.; Liu, H.; Wu, Z.; Li, Y. Landscape pattern analysis of the urban forest and green network structure in the Pudong district of Shanghai, China. Rev. Chapingo Ser. Cienc. For. Ambient. 2017, 23, 457–473. [Google Scholar] [CrossRef]
- Sudnik-Wójcikowska, B.; Jędrzejewska-Szmek, K.; Sikorski, P. Flora parku Mokotowskie Pole w Warszawie. Pr. Stud. Geogr. 2020, 65, 33–42. [Google Scholar]
- Kowarik, I.; von der Lippe, M.; Cierjacks, A. Prevalence of alien versus native species of woody plants in Berlin differs between habitats and at different scales. Preslia 2013, 85, 113–132. [Google Scholar]
- Čeplová, N.; Lososová, Z.; Kalusová, V. Urban ornamental trees: A source of current invaders; a case study from a European city. Urban Ecosyst. 2017, 20, 1135–1140. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.R.U.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D. Richardson DM Non-native species in urban environments: Patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef] [Green Version]
- Vojík, M.; Sádlo, J.; Petřík, P.; Pyšek, P.; Man, M.; Pergl, J. Two faces of parks: Sources of invasion and habitat for threatened native plants. Preslia 2020, 92, 353–373. [Google Scholar] [CrossRef]
- Celesti-Grapow, L.; Ricotta, C. Plant invasion as an emerging challenge for the conservation of heritage sites: The spread of ornamental trees on ancient monuments in Rome, Italy. Biol. Invasions 2021, 23, 1191–1206. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Barber, P.; Harper, R.; Linh, T.V.K.; Dell, B. Vegetation trends associated with urban development: The role of golf courses. PLoS ONE 2020, 15, e0228090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornal-Pieniak, B.; Ollik, M.; Schwerk, A. Impact of different levels of anthropogenic pressure on the plant species composition in woodland sites. Urban For. Urban Green. 2019, 38, 295–304. [Google Scholar] [CrossRef]
- Kowarik, I.; Fischer, L.K.; Kendal, D. Biodiversity conservation and sustainable urban development. Sustainability 2020, 12, 4964. [Google Scholar] [CrossRef]
- Santos, T.; Nogueira Mendes, R.; Vasco, A. Recreational activities in urban parks: Spatial interactions among users. J. Outdoor Recreat. Tour. 2016, 15, 1–9. [Google Scholar] [CrossRef]
- Zhai, Y.; Baran, P.K. Urban park pathway design characteristics and senior walking behavior. Urban For. Urban Green. 2017, 21, 60–73. [Google Scholar] [CrossRef]
- Nemec, K.T.; Allen, C.R.; Alai, A.; Clements, G.; Kessler, A.C.; Kinsell, T.; Major, A.; Bruce, J.; Stephen, B.J. Woody invasions of urban trails and the changing face of urban forests in the Great Plains, USA. Am. Midl. Nat. 2011, 165, 241–256. [Google Scholar] [CrossRef] [Green Version]
- GUS. Area and Population in the Territorial Profile in 2021; Statistics Poland: Warsaw, Poland, 2021; p. 23. [Google Scholar]
- Matuszko, D.; Piotrowicz, K. Cechy klimatu miasta a klimat Krakowa. In Miasto w Badaniach Geografów; Trzepacz, P., Więcław-Michniewska, J., Brzosko-Sermak, A., Kołoś, A., Eds.; Instytut Geografii i Gospodarki Przestrzennej Uniwersytetu Jagiellońskiego: Kraków, Poland, 2015; Volume 1, pp. 221–241. [Google Scholar]
- Załącznik do Zarządzenia nr 2282Prezydenta Miasta Krakowa z Dnia 29 Września 2019 r. Kierunki Rozwoju i ZarządzaniaTerenami Zieleni w Krakowie na Lata 2019–2030. Rozdział I-V. Urząd Miasta Krakowa, Wydział Kształtowania Środowiska, Os. Zgody 2, Kraków. Available online: https://zzm.krakow.pl/images/pliki/KRiZTZ/KRiZTZ_ROZDZIAL_I_V.pdf (accessed on 27 January 2022).
- Grzegorczyk, M.; Perzanowska, J. (Eds.) Skarby Przyrody i Kultury Krakowai i Okolic. Ekologiczne Ścieżki Edukacyjne; WAM Publisher: Krakow, Poland, 2005; p. 495. [Google Scholar]
- Sikora, Z.; Podwika, M. Szlak Lasów Miejskich Krakowa; Fundacja Miejski Park i Ogród Zoologiczny w Krakowie: Kraków, Poland, 2011; p. 58. [Google Scholar]
- The Management of Urban Green Areas in Krakow. Available online: https://zzm.krakow.pl (accessed on 25 March 2022).
- Csapodý, V. Keimlingsbestimmungsbuch der Dikotyledonen; Akademiai Kiado: Budapeszt, Hungary, 1968; p. 286. [Google Scholar]
- Muller, F.M. Seedlings of the North-Western European Lowland. A Flora of Seedlings, 1st ed.; Springer: Wageningen, The Netherlands, 1978; p. 653. [Google Scholar]
- Rutkowski, L. Klucz do Oznaczania Roślin Naczyniowych Polski Niżowej; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2004; p. 814. [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. VascularPlants of Poland. An Annotated Checklist; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2020; p. 526. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer: Berlin, Germany, 1964; p. 631. [Google Scholar]
- Klotz, S.; Kühn, I.; Durka, W. Eine Datenbankzubiologisch-okologischen Merkmalen der Gefabpflanzen in Deutschland. Schriftenreihe fur Vegetationkunde, Bundesamt fur Naturschutz. 2002. Available online: https://www.ufz.de/biolflor/index.jsp (accessed on 7 October 2021).
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimes, L.; Klimesová, J.; et al. The LEDA Traitbase: A database of life-history traits of Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Pladias. Database of the Czech Flora and Vegetation. 2014. Available online: http://www.pladias.org (accessed on 7 October 2021).
- Sádlo, J.; Chytrý, M.; Pergl, J.; Pyšek, P. Plant dispersal strategies: A new classification based on the multiple dispersal modes of individual species. Preslia 2018, 90, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Matuszkiewicz, W.A. Guide for Identification of Polish Plant Communities; Polish Scientific Publishers PWN: Warsaw, Poland, 2017; p. 540. [Google Scholar]
- Zając, M.; Zając, A. The Geographic Elements of Native Flora of Poland; Edited by Laboratory of Computer Chorology; Institute of Botany, Jagiellonian University: Kraków, Poland, 2009; p. 94. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; GeneralDirection of Environmental Protection: Warsaw, Poland, 2012; p. 197. [Google Scholar]
- Tokarska-Guzik, B.; Bzdęga, K.; Dajdok, Z.; Mazurska, K.; Solarz, W. Invasive alien plants in Poland—The state of research and the use of the results in practice. Environ. Socio-Econ. Stud. 2021, 9, 71–95. [Google Scholar] [CrossRef]
- Statistics Libretexts. 2019. Available online: https://stats.libretexts.org/@go/page/34 (accessed on 7 October 2021).
- Roovers, P.; Bossuyt, B.; Gulinck, H.; Hermy, M. Vegetation recovery on closed paths in temperate deciduous forests. J. Environ. Manag. 2005, 74, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Fekete, I.; Varga, C.; Biró, B.; Tóth, J.A.; Várbíró, G.; Lajtha, K.; Szabó, G.; Kotroczó, Z. The effects of litter production and litter depth on soil microclimate in a central european deciduous forest. Plant Soil 2016, 398, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Othaman, N.N.C.; Md Isa, N.; Ismail, R.C.; Ahmad, M.I.; Hui, C. K Factors that affect soil electrical conductivity (EC) based system for smart farming application. AIP Conf. Proc. 2020, 2203, 020055. [Google Scholar] [CrossRef]
- Pająk, M.; Krzaklewski, W.; Duda, K.; Gruba, P. Spatial and temporal variation in soil salinity as a result of chemical de-icing of road in Kraków, Poland. Fresenius Environ. Bull. 2015, 24, 3363–3370. [Google Scholar]
- Shannon, T.P.; Ahler, S.J.; Mathers, A.; Ziter, C.D.; Dugan, H.A. Road salt impact on soil electrical conductivity across an urban landscape. J. Urban Ecol. 2020, 6, juaa006. [Google Scholar] [CrossRef]
- Bartkowiak, A.; Lemanowicz, J.; Siwik-Ziomek, A. Assessment of selected heavy metals and enzymes in soil within the range of impact of illegal dumping sites. Int. J. Environ. Res. 2016, 10, 245–254. [Google Scholar]
- Gamar, A.; Khiya, Z.; Zair, T.; El Kabriti, M.; Bouhlal, A.; El Hilali, F. Assessment of physicochemical quality of the polluting load of leachates from the wild dump of El Hajeb city (Morocco). Int. J. Res. Granthaalayah 2017, 5, 63–71. [Google Scholar] [CrossRef]
- Gamar, A.; Zair, T.; El Kabriti, M.; El Hilali, F. Study of the impact of the wild dump leachates of the region of El Hajeb (Morocco) on the physicochemical quality of the adjacent water table. Karbala. Int. J. Mod. Sci. 2018, 4, 382–392. [Google Scholar] [CrossRef]
- Kida, K.; Kawahigashi, M. Influence of asphalt pavement construction processes on urban soil formation in Tokyo. Soil Sci. Plant Nutr. 2015, 61, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Huang, G.; Buyantuev, A.; Wu, J.; Luo, S.; Ma, K. Spatial heterogeneity of urban soils: The case of the Beijing metropolitan region, China. Ecol. Process 2014, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Muras, P. Standardy Zakładania i Pielęgnacji Podstawowych Rodzajów Terenów Zieleni w Krakowie na lata 2019–2030. 2016. Available online: https://zzm.krakow.pl/images/pliki/KRiZTZ/12_E_251069_0_zal5_aneks_III_standardy_zakladania_pielegnacji_zieleni.pdf (accessed on 24 March 2022).
- Qin, G.; Wu, J.; Zheng, X.; Zhou, R.; Wei, Z. Phosphorus forms and associated properties along an urban–rural gradient in Southern China. Water 2019, 11, 2504. [Google Scholar] [CrossRef] [Green Version]
- Paradeis, B.; Lovas, S.; Aipperspach, A.; Kazmierczak, A.; Boche, M.; He, Y.; Corrigan, P.; Chambers, K.; Gao, Y.; Norland, J.; et al. Dog-park soils: Concentration and distribution of urine-borne constituents. Urban Ecosyst. 2013, 16, 351–365. [Google Scholar] [CrossRef]
- Bonner, C.; Agnew, A.D.Q. Soil phosphorus as an indicator of canine faecal pollution in urban recreation areas. Environ. Pollut. Ser. B Chem. Phys. 1983, 6, 145–156. [Google Scholar] [CrossRef]
- Nawaz, M.F.; Bourrié, G.; Trolard, F. Soil compaction impact and modelling. A review. Agron. Sustain. Dev. 2013, 33, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Randrup, T.B.; Dralle, K. Influence of planning and design on soil compaction inconstruction sites. Landsc. Urban Plan. 1997, 38, 87–92. [Google Scholar] [CrossRef]
- Kostrakiewicz-Gierałt, K.; Pliszko, A.; Gmyrek-Gołąb, K. The effect of visitors on the properties of vegetation of calcareous grasslands in the context of width and distances from tourist trails. Sustainability 2020, 12, 454. [Google Scholar] [CrossRef] [Green Version]
- Pfitzenmeyer, C.D.C. Biological Flora of British Isles. Arrhenatherum elatius (L.) J. &. C. Presl (A. Avenaceum Beauv.). J. Ecol. 1962, 50, 235–245. [Google Scholar] [CrossRef]
- Beddows, A.R. Biological Flora of British Isles. Holcus lanatus L. J. Ecol. 1961, 49, 421–430. [Google Scholar] [CrossRef]
- Beddows, A.R. Biological Flora of British Isles. Lolium perenne L. J. Ecol. 1967, 55, 567–587. [Google Scholar] [CrossRef]
- Pigot, C.D. Biological Flora of British Isles. Thymus L. J. Ecol. 1955, 43, 365–387. [Google Scholar] [CrossRef]
- Kobayashi, T.; Hori, Y.; Nomoto, N. Effects of trampling and vegetation removal on species diversity and micro-environment under different shade conditions. J. Veg. Sci. 1997, 8, 873–880. [Google Scholar] [CrossRef]
- Moszkowicz, Ł.; Krzeptowska-Moszkowicz, I.; Porada, K. Relationship between parameters of public parks and their surroundings and the richness, diversity and species composition of vascular herbaceous plants on the example of Krakow in Central Europe. Landsc. Online 2021, 94, 1–16. [Google Scholar] [CrossRef]
- Sehrt, M.; Bossdorf, O.; Freitag, M.; Bucharova, A. Less is more! Rapid increase in plant species richness after reduced mowing in urban grasslands. Basic Appl. Ecol. 2020, 42, 47–53. [Google Scholar] [CrossRef]
- Jim, C.Y. Soil characteristics and management in an urban park in Hong Kong. Environ. Manag. 1998, 22, 683–695. [Google Scholar] [CrossRef]
- Sun, D. Trampling resistance, recovery and growth rate of eight plant species. Agric. Ecosyst. Environ. 1992, 38, 265–273. [Google Scholar] [CrossRef]
- Banaszek, J.; Leksy, M.; Rahmonov, O. The ecological diversity of vegetation within Urban Parks in the Dabrowski Basin (southern Poland). In Proceedings of the 10th International Conference “Environmental Engineering”, Vilnius Gediminas Technical University Lithuania, Vilnius, Lithuania, 27–28 April 2017. [Google Scholar] [CrossRef] [Green Version]
- Ignatieva, M.; Haase, D.; Dushkova, D.; Haase, A. Lawns in cities: From aglobalised urban green space phenomenon to sustainable nature-based solutions. Land 2020, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Bedla, D.; Halecki, W. The use of web application in monitoring the effects of introducing flower meadows in Kraków’s city parks. Geomat. Landmanag. Landsc. 2020, 4, 7–15. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press London: London, UK, 1977. [Google Scholar]
- Wódkiewicz, M.; Gruszczyńska, B. Genetic Diversity and spatial genetic structure of Stellaria holostea populations from urban forest islands. Acta Biol. Crac. Ser. Bot. 2014, 56, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Vakhlamova, T.; Rusterholz, H.-P.; Kamkin, V.; Baur, B. Recreational use of urban and suburban forests affects plant diversity in a Western Siberian city. Urban For. Urban Green. 2016, 17, 92–103. [Google Scholar] [CrossRef]
- Cao, Y.; Natuhara, Y. Effect of urbanization on vegetation in riparian area: Plant communities in artificial and semi-natural habitats. Sustainability 2020, 12, 204. [Google Scholar] [CrossRef] [Green Version]
- Duchesneau, K.; Derickx, L.; Antunes, P.M. Assessing the relative importance of human and spatial pressures on non-native plant establishment in urban forests using citizen science. NeoBiota 2021, 65, 1–21. [Google Scholar] [CrossRef]
- Szumańska, I.; Lubińska-Mielińska, S.; Kamiński, D.; Rutkowski, L.; Nienartowicz, A.; Piernik, A. Invasive Plant Species Distribution Is Structured by Soil and Habitat Type in the City Landscape. Plants 2021, 10, 773. [Google Scholar] [CrossRef]
- Bomanowska, A.; Rewicz, A.; Wolski, G.; Krasoń, K. Invasive alien plants in protected areas within city borders, Łódź, (Poland). Pak. J. Bot. 2017, 49, 311–316. [Google Scholar]
- Dauer, J.T.; Mortensen, D.A.; Vangessel, M.J. Temporal and spatial dynamics of long-distance Conyza canadensis seed dispersal. J. Appl. Ecol. 2007, 44, 105–114. [Google Scholar] [CrossRef]
- Huang, H.; Guo, S.; Chen, G. Reproductive biology in an invasive plant Solidago canadensis. Front. Biol. China 2007, 2, 196–204. [Google Scholar] [CrossRef]
- Von der Lippe, K.; Kowarik, I. Do cities export biodiversity? Traffic as dispersal vector across urban–rural gradients. Divers. Distrib. 2008, 14, 18–25. [Google Scholar] [CrossRef]
- Pickering, C.; Mount, A. Do tourists disperse weed seed? A global review of unintentional human-mediated terrestrial seed dispersal on clothing, vehicles and horses. J. Sustain. Tour. 2010, 18, 239–256. [Google Scholar] [CrossRef]
- Gelmi-Candusso, T.A.; Hämäläinen, A.M. Seeds and the city: The interdependence of zoochory and ecosystem dynamics in urban environments. Front. Ecol. Evol. 2019, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Dzwonko, Z.; Loster, S. Wskaźnikowe gatunki starych lasów i ich znaczenie dla ochrony przyrody i kartografii roślinności. Prace Georg. 2001, 178, 119–132. [Google Scholar]








| Name of Study Site | Code | GPS Coordinates | Altitude (m a.s.l.) | Width of the Path (cm) | Surface of the Path |
|---|---|---|---|---|---|
| Łęgowski Forest | F1 | N50°03.390′ E20°01.814′ | 203 | 220 | Natural |
| Mogilski Forest | F2 | N50°03.233′ E20°03.341′ | 210 | 260 | Anthropogenic (asphalt) |
| Wolski Forest | F3 | N50°03.327′ E19°51.468′ | 331 | 190 | Natural |
| Forest in Sikornik Hill | F4 | N50°03.509′ E19°53.236′ | 258 | 130 | Natural |
| Forest in Górka Pychowicka | F5 | N50°01.903′ E19°52.977′ | 240 | 230 | Natural |
| Tyniec Forest | F6 | N50°00.633′ E19°49.712′ | 277 | 250 | Natural |
| Forest in Skotniki | F7 | N50°01.251′ E19°51.120′ | 209 | 220 | Anthropogenic (gravel) |
| RżąckiForest | F8 | N50°00.342′ E19°59.797′ | 266 | 100 | Natural |
| Borkowski Forest | F9 | N50°00.608′ E19°54.795′ | 260 | 300 | Anthropogenic (gravel) |
| Witkowice Forest | F10 | N50°06.471′ E19°57.001′ | 249 | 100 | Natural |
| PolishAviator’s Park | P1 | N50°04.377′ E19°59.441′ | 223 | 160 | Anthropogenic (asphalt) |
| Dąbie Park | P2 | N50°03.608′ E19°59.055′ | 206 | 220 | Natural |
| Decius Park | P3 | N50°03.855′ E19°52.384′ | 219 | 260 | Anthropogenic (asphalt) |
| Twardowski Rocks Park | P4 | N50°02.366′ E19°54.154′ | 220 | 300 | Anthropogenic (asphalt) |
| StanisławWyspiański’sPark | P5 | N50°05.144′ E19°55.245′ | 235 | 310 | Anthropogenic (asphalt) |
| Henryk Jordan’s Park | P6 | N50°03.864′ E19°55.087′ | 206 | 230 | Anthropogenic (asphalt) |
| Kleparski Park | P7 | N50°04.572′ E19°56.310′ | 226 | 310 | Natural |
| Aleksandra’s Park | P8 | N50°00.827′ E20°00.828′ | 243 | 160 | Anthropogenic (gravel) |
| Solvay Park | P9 | N50°00.905′ E19°55.591′ | 273 | 180 | Natural |
| Wojciech Bednarski’s Park | P10 | N50°02.548′ E19°57.000′ | 218 | 370 | Anthropogenic (asphalt) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostrakiewicz-Gierałt, K.; Gmyrek, K.; Pliszko, A. The Effect of the Distance from a Path on Abiotic Conditions and Vascular Plant Species in the Undergrowth of Urban Forests and Parks. Int. J. Environ. Res. Public Health 2022, 19, 5621. https://doi.org/10.3390/ijerph19095621
Kostrakiewicz-Gierałt K, Gmyrek K, Pliszko A. The Effect of the Distance from a Path on Abiotic Conditions and Vascular Plant Species in the Undergrowth of Urban Forests and Parks. International Journal of Environmental Research and Public Health. 2022; 19(9):5621. https://doi.org/10.3390/ijerph19095621
Chicago/Turabian StyleKostrakiewicz-Gierałt, Kinga, Katarzyna Gmyrek, and Artur Pliszko. 2022. "The Effect of the Distance from a Path on Abiotic Conditions and Vascular Plant Species in the Undergrowth of Urban Forests and Parks" International Journal of Environmental Research and Public Health 19, no. 9: 5621. https://doi.org/10.3390/ijerph19095621