
Diversity and Phosphate Solubilizing Characteristics of Cultivable Organophosphorus-Mineralizing Bacteria in the Sediments of Sancha Lake

Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of Sancha Lake and Collection of Sediments
2.2. Isolation and Screening of OPB
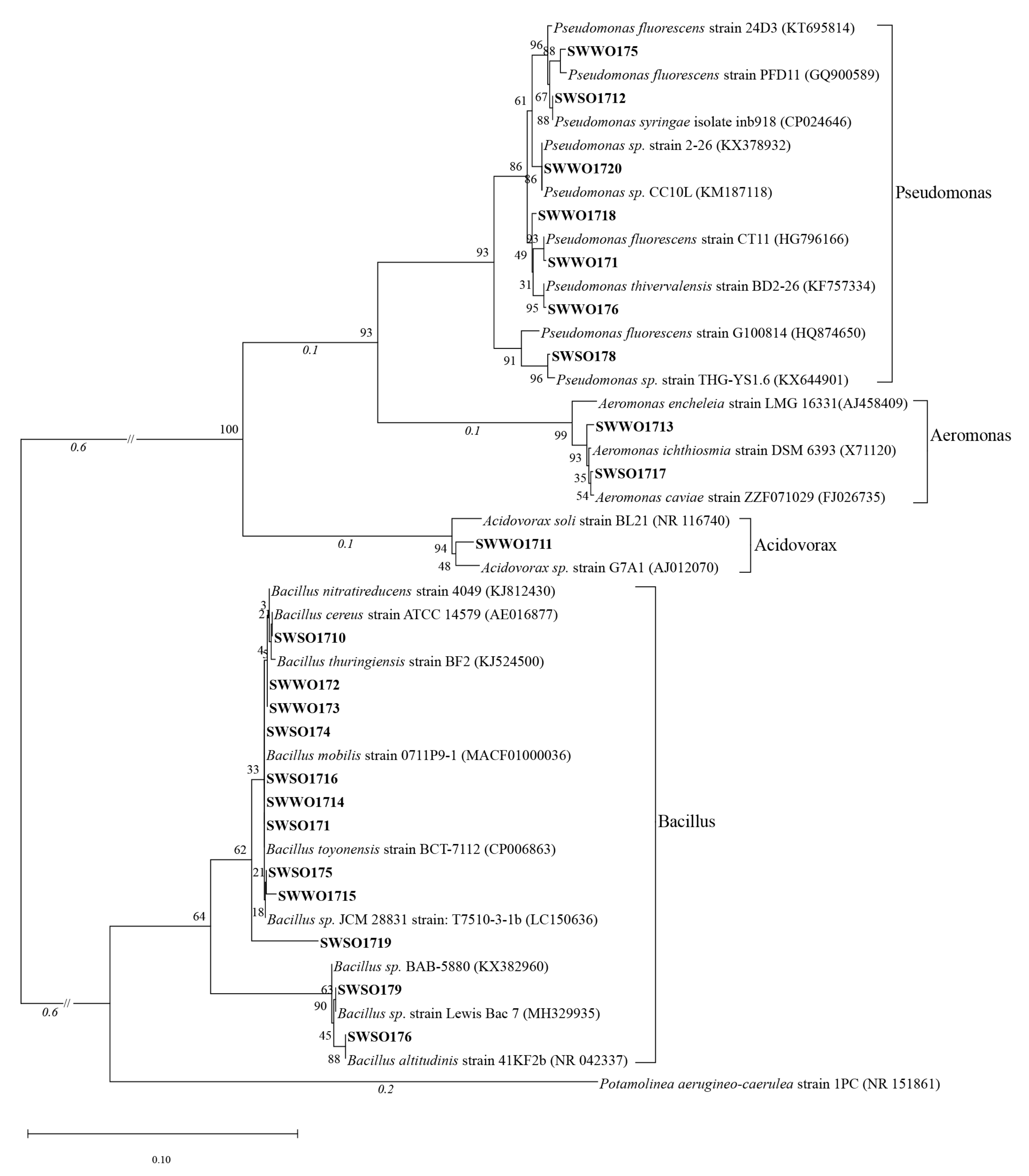
2.3. 16S rDNA Amplification, Sequencing, and Phylogenetic Analysis
2.4. Evaluation of P-Solubilizing Ability of the OPB Strains
2.4.1. Determination of P-Solubilizing Ability by Solid Lecithin Plate
2.4.2. Determination of P-Solubilizing Ability by Liquid Lecithin Medium
2.4.3. Microcosm Setup and P-Release by Bacterial Strains
2.4.4. Statistical Analysis
3. Results and Discussion
3.1. Results of OPB Screening and Identification
3.2. P-Solubilizing Ability
3.2.1. P-Solubilizing Activity of Screened OPB Strains
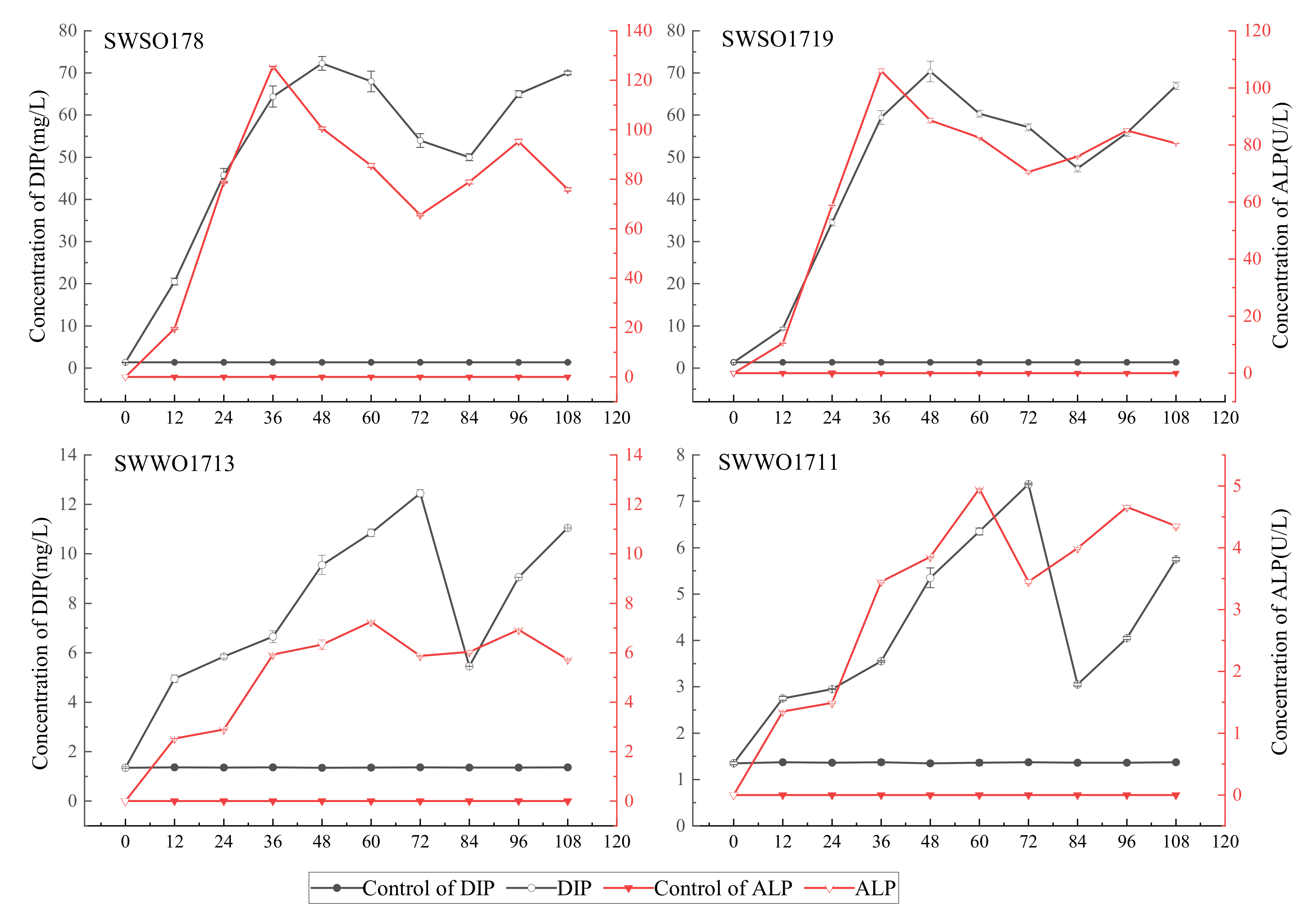
3.2.2. Changes in Alkaline Phosphatase and pH of the Organophosphate Culture of the Four Representative Organophosphate-Mineralizing Bacteria Strains and Their Organophosphate Solubilizing Ability
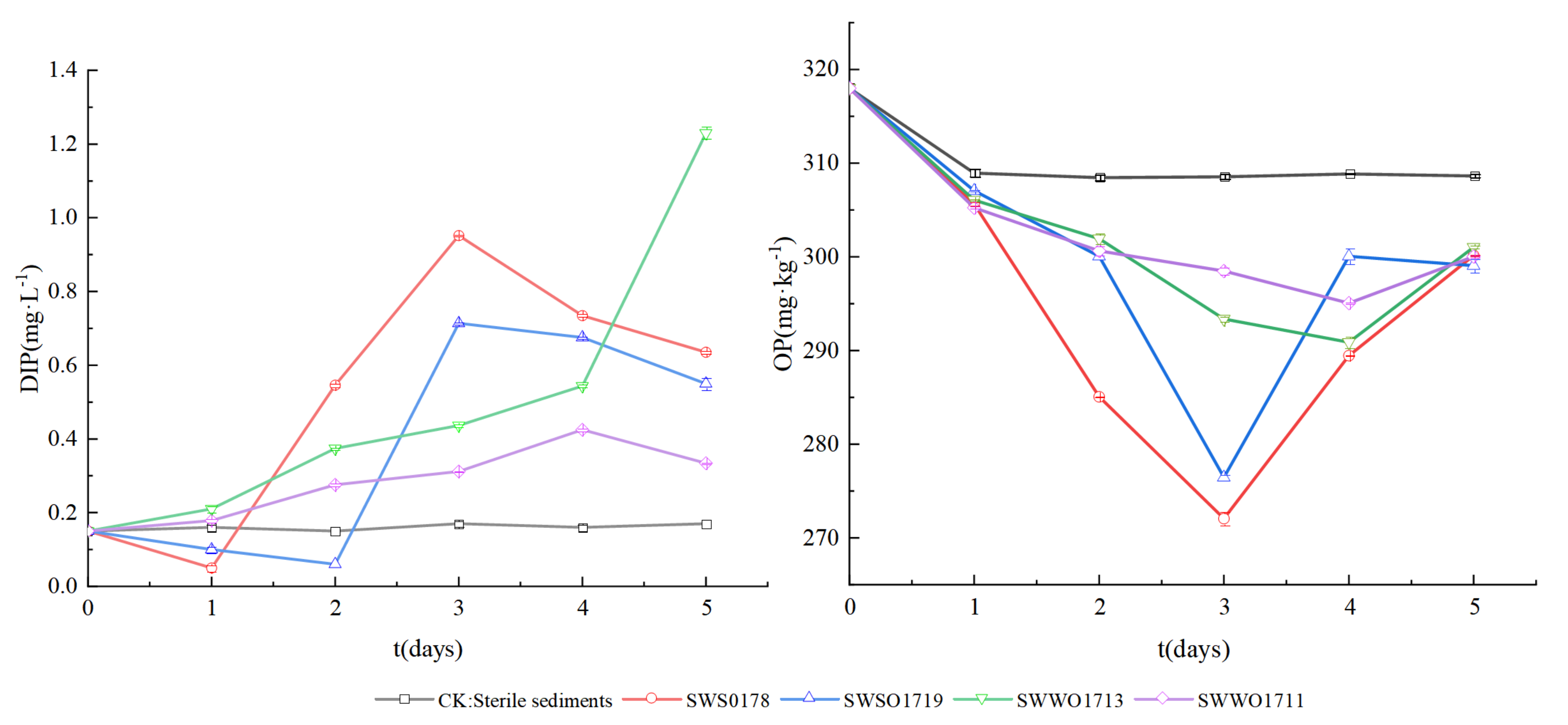
3.2.3. P-Release by OPB Strains in Sediments
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Xie, L.Q.; Xie, P.; Tang, H.J. Enhancement of dissolved phosphorus release from sediment to lake water by Microcystis blooms—An enclosure experiment in a hyper-eutrophic, subtropical Chinese lake. Environ. Pollut. 2003, 122, 391–399. [Google Scholar] [CrossRef]
- Peng, L.; Zhao, J.; Zhang, Y.; Hua, Y. Seasonal variation of bacterial community in sediments of urban eutrophic lakes. Chin. J. Appl. Environ. Biol. 2015, 21, 1012–1018. [Google Scholar]
- Wu, Q.H.; Zhang, R.D.; Huang, S.; Zhang, H.J. Effects of bacteria on nitrogen and phosphorus release from river sediment. J. Environ. Sci. 2008, 20, 404–412. [Google Scholar] [CrossRef]
- Van-Mourik, A.; Bleumink-Pluym, N.M.C.; Van-Dijk, L.; Van-Putten, J.P.M.; Wosten, M.M.S.M. Functional analysis of a Campylobacter jejuni alkaline phosphatase secreted via the Tat export machinery. Microbiology 2008, 154, 584–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilanjan, M.; Sanjib, K.M.; Srikanta, S.; Keka, S.; Debashri, D.; Chinmay, B.; Sanjeev, K.S.; Anil, P.S. Ecological significance and phosphorus release potential of phosphate solubilizing bacteria in freshwater ecosystems. Hydrobiologia 2015, 745, 69–83. [Google Scholar]
- Qian, Y.C.; Shi, J.Y.; Chen, Y.X.; Lou, L.P.; Cui, X.Y.; Cao, R.K.; Li, P.F.; Tang, J. Characterization of phosphate solubilizing bacteria from the sediments of shallow eutrophic lake and wetland: Isolation, molecular identification and phosphorus release ability determination. Molecules 2010, 15, 8518–8533. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tian, X.X.; Qu, L.Y. The phosphate-dissolving characteristics of organic phosphorus bacterium isolated from Jiulong River. Mar. Environ. Sci. 2013, 32, 736–740. [Google Scholar]
- De-Souza, M.; Nair, S.; Chandramohan, D. Phosphate solubilizing bacteria around Indian peninsula. Indian J. Mar. Sci. 2000, 29, 48–51. [Google Scholar]
- Seshadri, S.; Ignacimuthu, S.; Lakshminarsimhan, C. Variations in heterotrophic and phosphate solubilizing bacteria from Chennai, southeast coast of India. Indian J. Mar. Sci. 2002, 31, 69–72. [Google Scholar]
- Mudryk, Z. Decomposition of organic and solubilisation of inorganic phosphorus compounds by bacteria isolated from a marine sandy beach. Mar. Biol. 2004, 145, 1227–1234. [Google Scholar] [CrossRef]
- Zhou, C.; Song, C.L.; Huang, D.Z.; Liu, Y.B.; Cao, X.Y.; Zhou, Y.Y. Isolation and characterization of organic phosphorus-mineralizing bacteria in sediment of a Chinese large shallow eutrophic lake (Lake Taihu). Geomicrobiol. J. 2011, 28, 660–666. [Google Scholar] [CrossRef]
- Jia, B.Y.; Tang, Y.; Fu, W.L.; Yu, J.; Zhang, C. Relationship among sediment characteristics, eutrophication process and human activities in the Sancha lake. China Environ. Sci. 2013, 33, 1638–1644. [Google Scholar]
- Li, Y.; Zhang, J.Q.; Gong, Z.L.; Fu, W.L.; Wu, D.M. Fractions and temporal and spatial distribution of phosphorus in the sediments of Sancha lake. Appl. Ecol. Environ. Res. 2019, 17, 11731–11743. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.Q.; Gong, Z.L.; Xu, W.L.; Mou, Z.S. Gcd gene diversity of quinoprotein glucose dehydrogenase in the sediment of Sancha lake and its response to the environment. Int. J. Environ. Res. Public Health 2019, 16, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, J.J.; Zhang, J.Q.; Xu, W.L.; Mou, Z.S. Characteristics of Inorganic Phosphate-Solubilizing Bacteria from the Sediments of a Eutrophic Lake. Int. J. Environ. Res. Public Health 2019, 16, 2141. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Niewolak, S. Occurrence of microorganisms in fertilized lakes. II. Lecithin-mineralizing microorganisms. Polskie Arch. Hydrobiol. 1980, 27, 53–71. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic studies. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Studier, J.A.; Keppler, K.J. A note on the neighbor-joining algorithm of Saitou and Nei. Mol. Biol. Evol. 1988, 6, 729–731. [Google Scholar]
- Qiao, C.C.; Wang, T.T.; Wang, R.F.; Liu, C.; Gao, Q.; Li, R.; Shen, Q.R. Screening phosphate solubilizing bacterial strains from maize rhizosphere and research on their plant growth promotion effect. J. Nanjing Agric. Univ. 2017, 40, 664–670. [Google Scholar]
- Huang, Q.H.; Wang, D.H.; Wang, C.X.; Ma, M.; Wang, Z.J. Relationship between phosphorus morphology in sediment and lake eutrophication. China Environ. Sci. 2003, 23, 583–586. [Google Scholar]
- You, X.J.; Su, Y.J.; Zhan, X.C.; Zhuang, Y.T.; Chen, Y.Z.; Lin, W.Z. Research on effect of microbe on phosphorus releasing from sediment in dutang reservoir. J. Fujian Norm. Univ. (Nat. Sci. Ed.) 2021, 28, 88–93. [Google Scholar]
- Ruban, V.; Brigault, S.; Demare, D.; Philippe, A.M. An investigation of the origin and mobility of phosphorus in freshwater sediments from Bort-Les-Orgues Reservoir, France. J. Environ. Monit. 1999, 1, 403–407. [Google Scholar] [CrossRef]
- Ruban, V.; López-Sánchez, J.F.; Pardo, P.; Rauret, G.; Muntau, H.; Quevauviller, P. Harmonized Protocol and certified reference material for the determination of extractable contents of phosphorus in freshwater sediments—A synthesis of recent works. Fresenius J. Anal. Chem. 2001, 370, 224–228. [Google Scholar] [CrossRef]
- Wu, G.F.; Zhou, X.P. Characterization of phosphorus-releasing bacteria in a small eutrophic shallow lake, Eastern China. Water Res. 2005, 39, 4623–4632. [Google Scholar]
- Dong, Y.M.X.; Fan, Z.Q.; Zhang, Z.; Xia, X.H.; Tian, S.P.; Zhuo, J.M. Experimental researches on phosphorus decomposing and concentrating of microorganism in Dianchi Lake: An example of modern phosphorus deposition and microorganism mineralization. J. Chang. Univ. Sci. Technol. 2003, 33, 282–289. [Google Scholar]
- Li, Y.; Ai, M.J.; Sun, Y.; Zhang, Y.Q.; Zhang, J.Q. Spirosoma lacussanchae sp. nov., a phosphate-solubilizing bacterium isolated from a freshwater reservoir. Int. J. Syst. Evol. Microbiol. 2017, 67, 3144–3149. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Shi, J.Y. Isolation, purification, and phosphate-solubilizing capability of phosphorous bacteria in West Lake sediment. Chin. J. Appl. Ecol. 2006, 17, 2112–2116. [Google Scholar]
- Labry, C.; Delmas, D.; Herbland, A. Phytoplankton and bacterial alkaline phosphatase activities in relation to phosphate and DOP availability within the Gironde plume waters (Bay of Biscay). J. Exp. Mar. Biol. Ecol. 2005, 318, 213–225. [Google Scholar] [CrossRef] [Green Version]
- Rossolinia, G.M.; Schippab, S.; Riccioa, M.L.; Berlutti, F.; Macaskie, L.E.; Thaller, M.C. Bacterial nonspecific acid phosphatases: Physiology, evolution, and use as tools in microbial biotechnology. Cell. Mol. Life Sci. 1998, 54, 833–850. [Google Scholar] [CrossRef]
- Behera, B.C.; Singevsachan, S.K.; Mishra, R.R.; Dutta, S.K.; Thatoil, H.N. Diversity, mechanism and biotechnology of phosphate solubilising microorganism in mangrove—A review. Biocatal. Agric. Biotechnol. 2014, 3, 97–110. [Google Scholar] [CrossRef]
- Sebastián, M.; Arístegui, J.; Montero, M.F.; Escanez, J.; Niell, F.X. Alkaline phosphatase activity and its relationship to inorganic phosphorus in the transition zone of the North-western African upwelling system. Prog. Oceanogr. 2004, 62, 31–150. [Google Scholar] [CrossRef]
- Chappell, K.R.; Goulder, R. Enzymes as river pollutants and the response of native epilithic extracellular-enzyme activity. Environ. Pollut. 1994, 86, 161–169. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, X.L.; Cui, L.B.; Shang, K.; Qu, L.Y. Diversity and characteristics of culturable phosphate-solubilizing bacteria of Indian Ocean. Microbiol. China 2015, 42, 1847–1857. [Google Scholar]
- Oh, W.S.; Im, Y.S.; Yeon, K.Y.; Yoon, Y.J.; Kim, J.W. Phosphate and carbon source regulation of alkaline phosphatase and phospholipase in Vibrio vulnificus. J. Microbiol. 2007, 45, 311–317. [Google Scholar] [PubMed]
- Cao, X.Y.; Song, C.L.; Zhou, Y.Y. Limitations of using extracellular alkaline phosphatase activities as a general indicator for describing P deficiency of phytoplankton in Chinese shallow lakes. J. Appl. Phycol. 2010, 22, 33–41. [Google Scholar] [CrossRef]
- Zhao, X.R.; Lin, Q.M.; Sun, Y.X.; Yao, J.; Zhang, Y.S. The methods for quantifying capacity of bacteria in dissolving P compounds. Microbiol. China 2001, 28, 1–4. [Google Scholar]
- Søndergaard, M.; Jensen, J.; Jeppesen, E. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 2003, 506, 135–145. [Google Scholar] [CrossRef]
- Hu, X.J.; Xu, Y.N.; Hu, B.W.; Xu, Y.; Yang, K.; Wen, G.L.; Li, Z.J.; Cao, Y.C. Phosphate solubilizing effects of phosphate solubilizing bacteria PSBHY-3 on pond sediment. J. South. Agric. 2018, 49, 2096–2102. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria Strain | Most Closely Related Strains (Accession Number) a | Gene Identity (%) b | Taxonomical Assignment | Query Length | Accession Number c |
|---|---|---|---|---|---|
| SWSO171 | Bacillus thuringiensis strain ATCC 10792(ACNF01000156) | 100 | Bacillus thuringiensis | 1460 | MK828267 |
| SWSO172 | Bacillus altitudinis strain 41KF2b (ASJC01000029) | 99.93 | Bacillus altitudinis | 1456 | MK828268 |
| SWSO174 | Bacillus mobilis strain 0711P9-1(MACF01000036) | 100 | Bacillus mobilis | 1492 | MK828270 |
| SWSO175 | Bacillus proteolyticus strain TD42(MACH01000033) | 99.66 | Bacillus proteolyticus | 1475 | MK828271 |
| SWSO176 | Bacillus altitudinis 41KF2b (ASJC01000029) | 100 | Bacillus altitudinis | 1456 | MK828272 |
| SWSO177 | Bacillus stratosphericus strain 41KF2a (ASJC01000029) | 100 | Bacillus stratosphericus | 1472 | MK828273 |
| SWSO179 | Bacillus pumilus strain ATCC 7061(ABRX01000007) | 99.79 | Bacillus pumilus | 1457 | MK828275 |
| SWSO1710 | Bacillus pacificus strain EB422 (KJ812450) | 99.86 | Bacillus pacificus | 1473 | MK828276 |
| SWSO1711 | Bacillus altitudinis 41KF2b (ASJC01000029) | 100 | Bacillus altitudinis | 1469 | MK828277 |
| SWSO1713 | Bacillus thuringiensis strain ATCC 10792(ACNF01000156) | 100 | Bacillus thuringiensis | 1474 | MK828278 |
| SWSO1714 | Bacillus luti strain TD41(MACI01000041) | 99.80 | Bacillus luti | 1472 | MK828279 |
| SWSO1715 | Bacillus albus strain N35-10-2(MAOE01000087) | 99.86 | Bacillus albus | 1476 | MK828280 |
| SWSO1716 | Bacillus luti strain TD41(MACI01000041) | 100 | Bacillus luti | 1459 | MK828281 |
| SWSO1719 | Bacillus albus strain N35-10-2(MAOE01000087) | 96.52 | Bacillus jysancha sp.nov | 1464 | MK863518 |
| SWWO172 | Bacillus paramycoides strain MCCC 1A04098(NR157734.1) | 100 | Bacillus paramycoides | 1458 | MK828283 |
| SWWO173 | Bacillus nitratireducens strain 4049(KJ812430) | 99.93 | Bacillus nitratireducens | 1460 | MK828284 |
| SWWO178 | Bacillus altitudinis 41KF2b(ASJC01000029) | 100 | Bacillus altitudinis | 1469 | MK828289 |
| SWWO179 | Bacillus altitudinis 41KF2b(ASJC01000029) | 100 | Bacillus altitudinis | 1460 | MK828290 |
| SWWO1712 | Bacillus thuringiensis strain ATCC 10792(ACNF01000156) | 100 | Bacillus thuringiensis | 1469 | MK828292 |
| SWWO1714 | Bacillus toyonensis strain BCT-7112 (CP006863) | 100 | Bacillus toyonensis | 1460 | MK828294 |
| SWWO1715 | Bacillus sp. strain AFS096926(NVLJ01000028) | 99.58 | Bacillus sp. | 1418 | MK828295 |
| SWWO1716 | Bacillus mobilis strain 0711P9-1(MACF01000036) | 99.93 | Bacillus mobilis | 1472 | MK828296 |
| SWSO178 | Pseudomonas sp. strain LY1(LSSW01000001) | 97.93 | Pseudomonas sp. | 1413 | MK828274 |
| SWSO1712 | Pseudomonas fluorescens strain PgKB31(MH553942.1) | 99.51 | Pseudomonas fluorescens | 1443 | MK834812 |
| SWWO171 | Pseudomonas silesiensis strain A3 (KX276592) | 99.86 | Pseudomonas silesiensis | 1446 | MK828282 |
| SWWO174 | Pseudomonas sp. strain R28 (CM002330) | 100 | Pseudomonas sp. | 1443 | MK828285 |
| SWWO175 | Pseudomonas sp. strain R28 (CM002330) | 99.79 | Pseudomonas sp. | 1453 | MK828286 |
| SWWO176 | Pseudomonas kilonensis strain ATCC 49054(EU391388) | 99.38 | Pseudomonas kilonensis | 1450 | MK828287 |
| SWWO177 | Pseudomonas silesiensis strain A3(KX276592) | 100 | Pseudomonas silesiensis | 1459 | MK828288 |
| SWWO1710 | Pseudomonas baetica strain a390(FM201274) | 99.05 | Pseudomonas baetica | 1465 | MK834811 |
| SWWO1718 | Pseudomonas sp. strain 11K1(CP035088) | 100 | Pseudomonas sp. | 1469 | MK863513 |
| SWWO1720 | Pseudomonas yamanorum strain 8H1(EU557337) | 99.86 | Pseudomonas yamanorum | 1457 | MK863515 |
| SWWO1713 | Aeromonas veronii strain CECT 4257(CDDK01000015) | 99.44 | Aeromonas veronii | 1437 | MK828293 |
| SWSO1717 | Aeromonas encheleia strain LMG 16331(AJ458409) | 98.56 | Aeromonas encheleia | 1443 | MK863516 |
| SWSO1718 | Aeromonas encheleia strain LMG 16331(AJ458409) | 98.52 | Aeromonas encheleia | 1438 | MK863517 |
| SWWO1719 | Aeromonas veronii strain CECT 4257(CDDK01000015) | 99.83 | Aeromonas veronii | 1468 | MK863514 |
| SWWO1717 | Aeromonas veronii strain CECT 4257(CDDK01000015) | 98.63 | Aeromonas veronii | 1426 | MK863512 |
| SWWO1721 | Spirosoma soli MIMBbqt12T(KT347096) | 96.32 | Spirosoma lacussanchae sp.nov | 1459 | KX580025 |
| SWWO1711 | Acidovorax sp. strain Root219(LMIJ01000041) | 99.51 | Acidovorax sp. | 1434 | MK828291 |
| Bacteria Strain | Phosphate Solubilizing Halo (HD/CD) ⅰ | DIP (mg·L−1) ⅱ | ALP (U·L−1) ⅲ | Cell Density (log CFU mL−1) ⅳ |
|---|---|---|---|---|
| Control | — | 1.35 ± 0.16 k | 0 | 0 |
| SWSO1712 | 4.1 ± 0.3 ab | 21.92 ± 0.35 f | 8.8 ± 0.9 cd | 5.10 f |
| SWSO1718 | 1.2 ± 0.1 h | 2.26 ± 0.20 j | 5.6 ± 0.4 fg | 5.98 cd |
| SWSO1717 | 1.3 ± 0.1 g | 10.36 ± 0.20 gh | 12.3 ± 0.16 b | 5.06 f |
| SWSO171 | 3.0 ± 0.3 d | 51.67 ± 1.12 c | 6.2 ± 0.5 f | 6.75 b |
| SWSO172 | 4.3 ± 0.4 a | 62.65 ± 1.20 b | 5.0 ± 0.5 g | 6.85 ab |
| SWSO173 | 4.1 ± 0.3 ab | 51.53 ± 1.32 c | 5.4 ± 0.4 g | 6.19 c |
| SWSO174 | 4.2 ± 0.2 ab | 62.12 ± 0.82 b | 5.1 ± 0.3 g | 7.39 a |
| SWSO175 | 4.1 ± 0.3 ab | 32.02 ± 1.42 e | 8.1 ± 0.7 d | 6.27 c |
| SWSO176 | 4.0 ± 0.2 b | 32.25 ± 1.20 e | 8.2 ± 0.9 d | 6.65 b |
| SWSO177 | 3.0 ± 0.2 d | 31.36 ± 0.97 e | 7.9 ± 1.0 d | 6.93 b |
| SWSO178 | 2.3 ± 0.2 ef | 72.26 ± 1.27 a | 3.5 ± 0.2 h | 7.14 a |
| SWSO179 | 4.2 ± 0.2 ab | 37.23 ± 1.30 d | 7.5 ± 0.6 cd | 5.79 cd |
| SWSO1710 | 3.5 ± 0.2 c | 31.75 ± 1.10 e | 8.0 ± 0.6 d | 6.71 b |
| SWSO1711 | 3.7 ± 0.3 bc | 22.09 ± 0.35 f | 8.9 ± 1.1 cd | 6.16 c |
| SWSO1713 | 3.6 ± 0.2 c | 21.82 ± 0.35 f | 8.9 ± 0.6 cd | 6.75 b |
| SWSO1714 | 1.4 ± 0.1 g | 9.82 ± 0.35 gh | 12.7 ± 1. 6 ab | 5.99 cd |
| SWSO1715 | 1.8 ± 0.3 fg | 3.78 ± 0.15 i | 6.9 ± 0.6 e | 6.09 cd |
| SWSO1716 | 2.8 ± 0.3 e | 13.82 ± 0.15 g | 10.2 ± 0.9 c | 5.27 e |
| SWSO1719 | 4.5 ± 0.1a | 70.36 ± 1.20 a | 3.8 ± 0.2 h | 7.16 a |
| SWWO171 | 3.2 ± 0.2 cd | 21.78 ± 0.12 f | 8.5 ± 0.6 cd | 6.62 b |
| SWWO172 | 1.3 ± 0.1 g | 2.74 ± 0.11 j | 6.4 ± 0. 6 f | 5.67 d |
| SWWO173 | 2.1 ± 0.1 f | 3.67 ± 0.10 i | 6.8 ± 0.9 e | 6.02 cd |
| SWWO174 | 1.3 ± 0.3 g | 12.27 ± 0.22 g | 11.5 ± 1.6 bc | 5.90 cd |
| SWWO175 | ND | 1.95 ± 0.09 jk | 6.2 ± 0.2 f | 5.06 f |
| SWWO176 | 3.1 ± 0.2 d | 22.07 ± 0.38 f | 8.5 ± 0.2 cd | 6.16 c |
| SWWO177 | 2.1 ± 0.2 f | 19.49 ± 0.05 f | 9.3 ± 0.6 c | 6.05 cd |
| SWWO178 | 3.1 ± 0.1d | 30.5 ± 0.05 e | 8.3 ± 0.7 d | 5.67 d |
| SWWO1718 | ND | 50.56 ± 1.31 c | 7.2 ± 0.6 e | 6.99 b |
| SWWO1719 | ND | 2.41 ± 0.35 j | 6.7 ± 0.7 e | 5.82 cd |
| SWWO1720 | ND | 2.37 ± 0.35 j | 6.2 ± 0.6 f | 6.99 b |
| SWWO179 | 3.0 ± 0.2 d | 27.52 ± 0.51 ef | 8.1 ± 0.9 d | 6.16 c |
| SWWO1710 | 1.3 ± 0.1 g | 2.52 ± 0.08 j | 6.0 ± 0.8 f | 5.08 f |
| SWWO1711 | 1.1 ± 0.1 h | 7.37 ± 0.2 h | 13.7 ± 1.6 a | 5.16 f |
| SWWO1712 | 2.0 ± 0.2 f | 18.82 ± 0.18 fg | 9.4 ± 1.1 c | 5.29 d |
| SWWO1713 | 1.2 ± 0.1 h | 12.45 ± 0.08 g | 11.3 ± 1.6 bc | 5.25 e |
| SWWO1714 | 1.2 ± 0.2 h | 3.32 ± 0.35 i | 6.5 ± 0.6 ef | 4.81 g |
| SWWO1715 | 1.6 ± 0.4 fg | 2.09 ± 0.35 j | 5.3 ± 0.4 g | 5.83 cd |
| SWWO1716 | 1.3 ± 0.2 g | 1.92 ± 0.35 jk | 4.9 ± 0.3 g | 4.80 g |
| SWWO1717 | 1.5 ± 0.1 g | 1.82 ± 0.35 jk | 4.5 ± 0.5 g | 4.87 fg |
| SWWO1721 | 1.2 ± 0.2 h | 3.25 ± 0.12 i | 10.4 ± 0.9 c | 5.26 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yu, X.; Zheng, J.; Gong, Z.; Xu, W. Diversity and Phosphate Solubilizing Characteristics of Cultivable Organophosphorus-Mineralizing Bacteria in the Sediments of Sancha Lake. Int. J. Environ. Res. Public Health 2022, 19, 2320. https://doi.org/10.3390/ijerph19042320
Li Y, Yu X, Zheng J, Gong Z, Xu W. Diversity and Phosphate Solubilizing Characteristics of Cultivable Organophosphorus-Mineralizing Bacteria in the Sediments of Sancha Lake. International Journal of Environmental Research and Public Health. 2022; 19(4):2320. https://doi.org/10.3390/ijerph19042320
Chicago/Turabian StyleLi, Yong, Xintao Yu, Jiarui Zheng, Zhilian Gong, and Wenlai Xu. 2022. "Diversity and Phosphate Solubilizing Characteristics of Cultivable Organophosphorus-Mineralizing Bacteria in the Sediments of Sancha Lake" International Journal of Environmental Research and Public Health 19, no. 4: 2320. https://doi.org/10.3390/ijerph19042320