
Partial Replacement of Dietary Fat with Krill Oil or Coconut Oil Alleviates Dyslipidemia by Partly Modulating Lipid Metabolism in Lipopolysaccharide-Injected Rats on a High-Fat Diet

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments and Diets
2.2. Analysis of the Fatty Acid Composition of Experimental Diets
2.3. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT)
2.4. Analysis of Fatty Acids Composition of Whole Blood
2.5. Determination of Serum Metabolic Parameters
2.6. Measurement of Serum IL-1β and MCP-1 Levels
2.7. Measurement of Lipid Contents in the Liver
2.8. Histological Assessment of the Liver and Epididymal Adipose Tissue
2.9. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.10. Statistical Analysis
3. Results
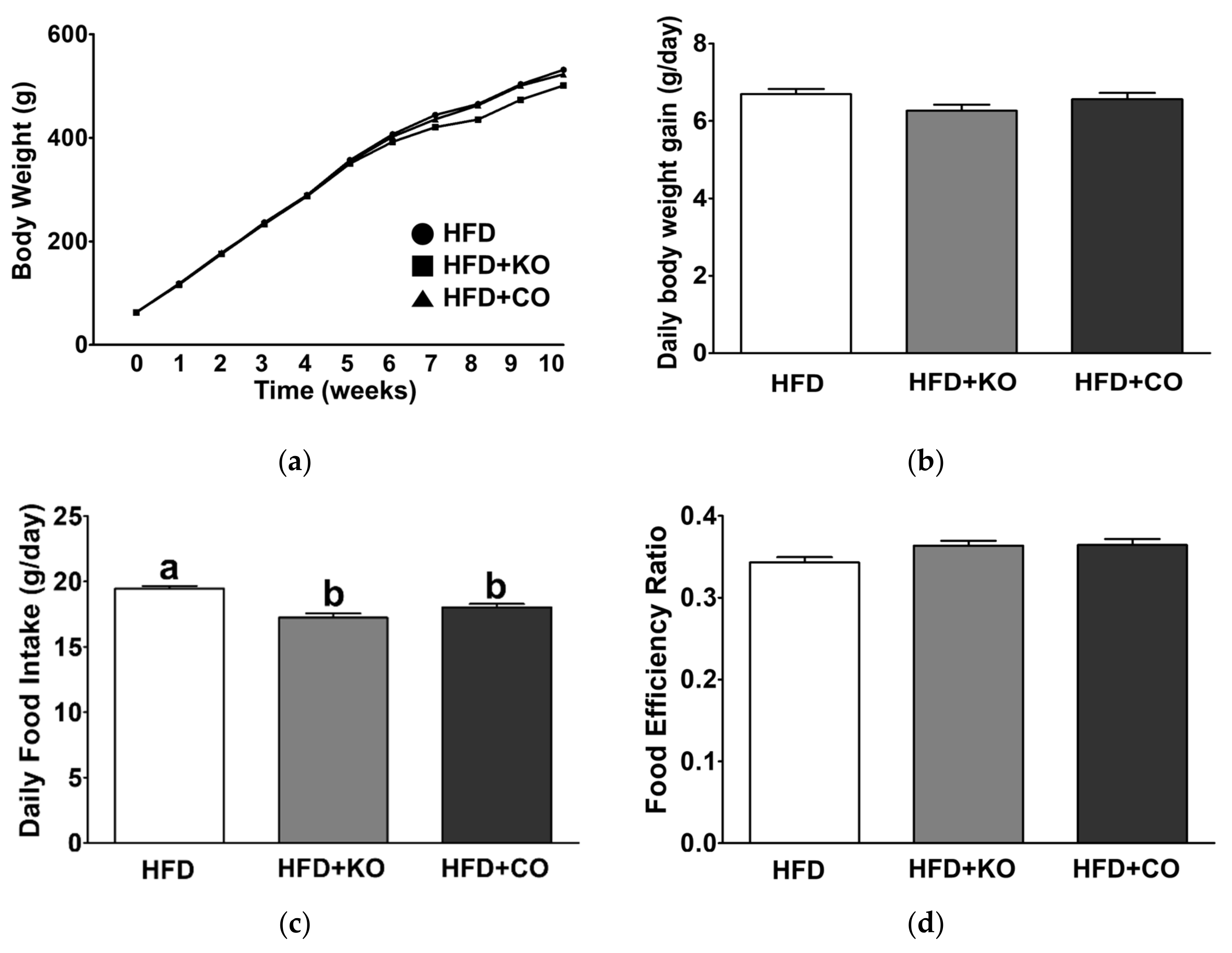
3.1. Effect of the Dietary Fat Replacement with KO or CO on Body Weight Changes, Food Intake, Energy Intake, and Food Efficiency Ratio (FER)
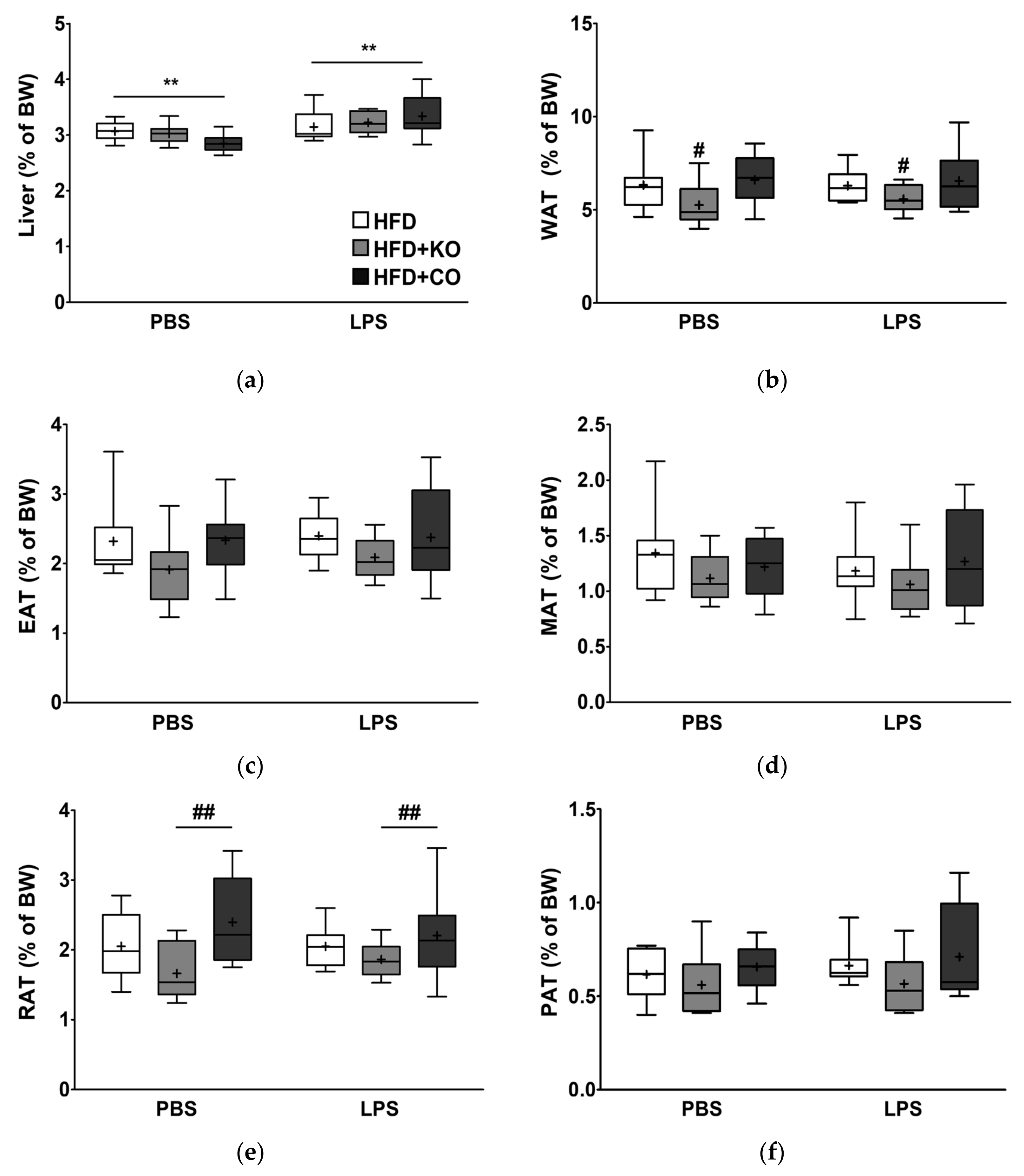
3.2. Effect of the Dietary Fat Replacement with KO or CO on Liver and Adipose Tissue Weights
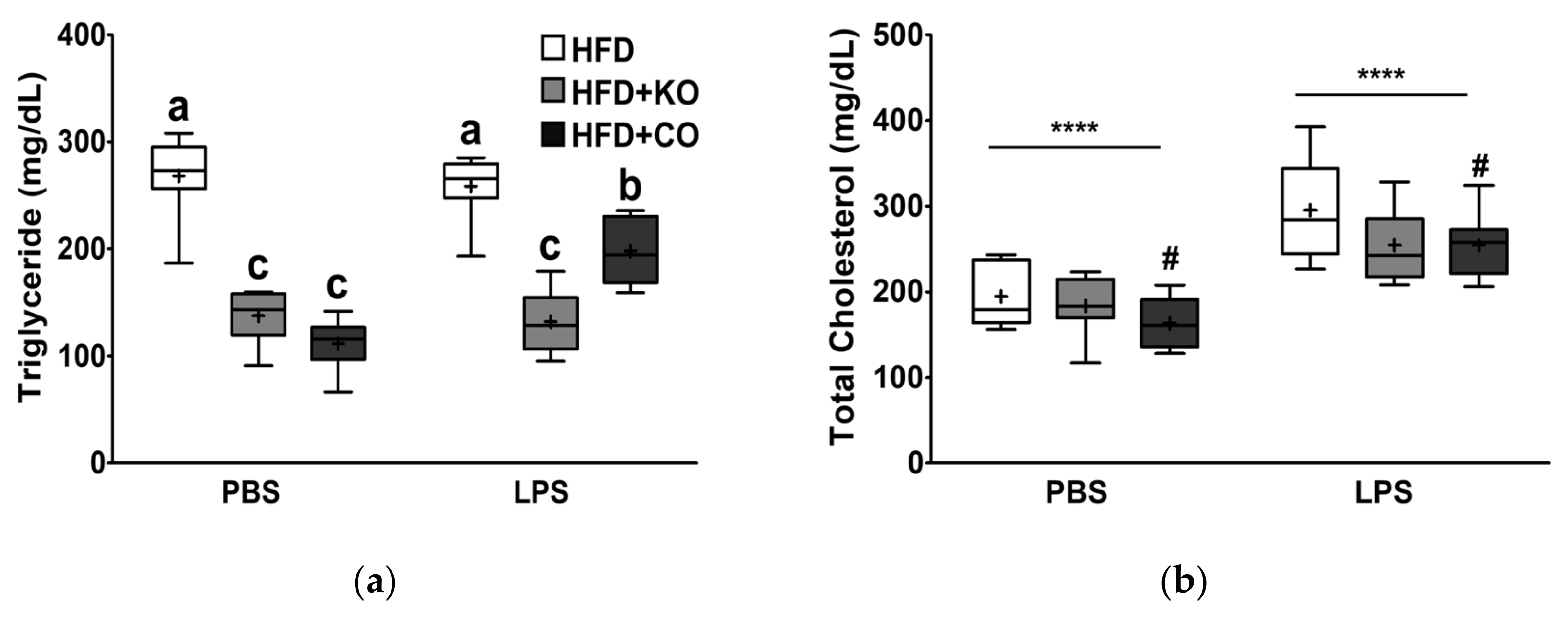
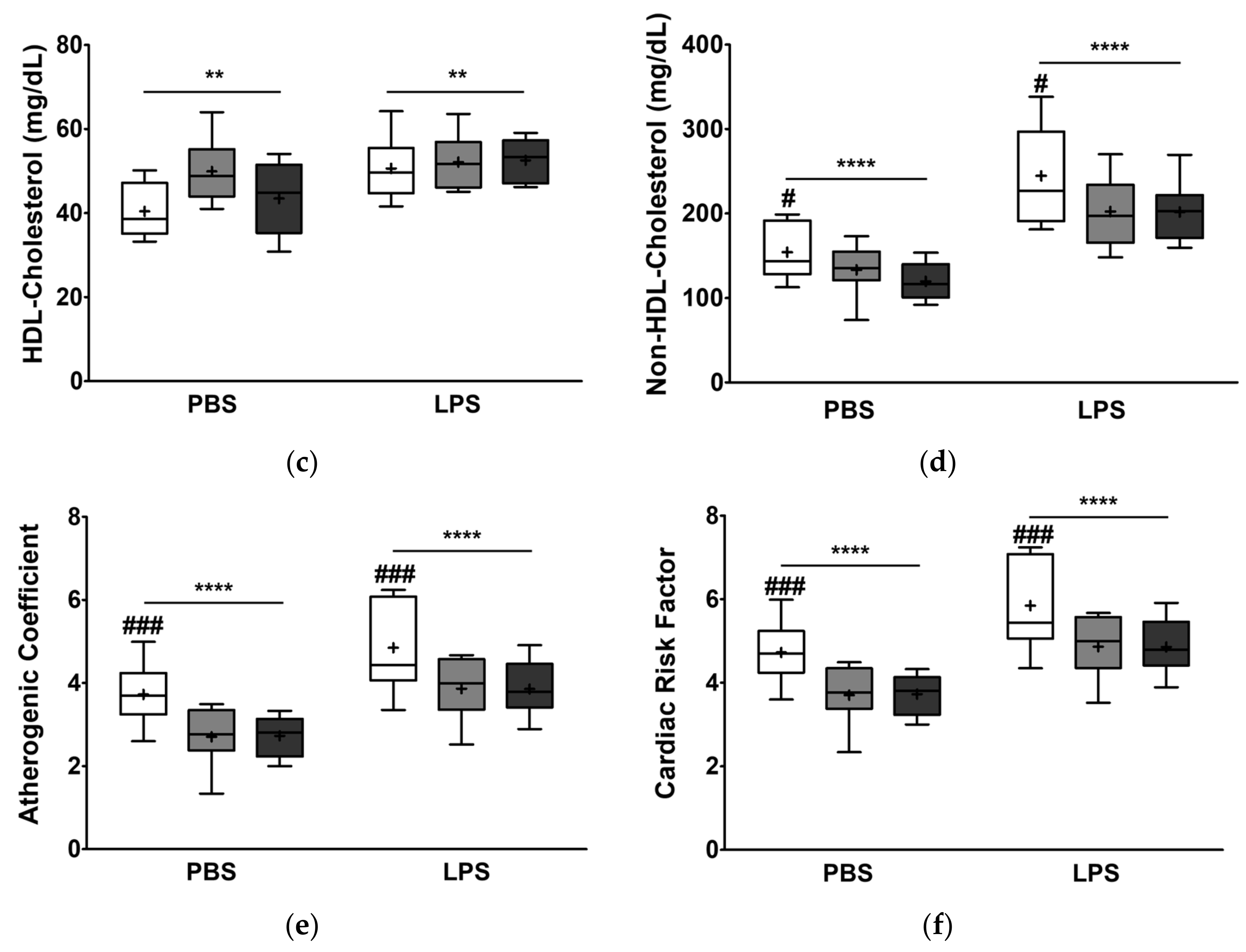
3.3. Effect of the Dietary Fat Replacement with KO or CO on Serum Lipid Profiles and Cardiovascular Parameters
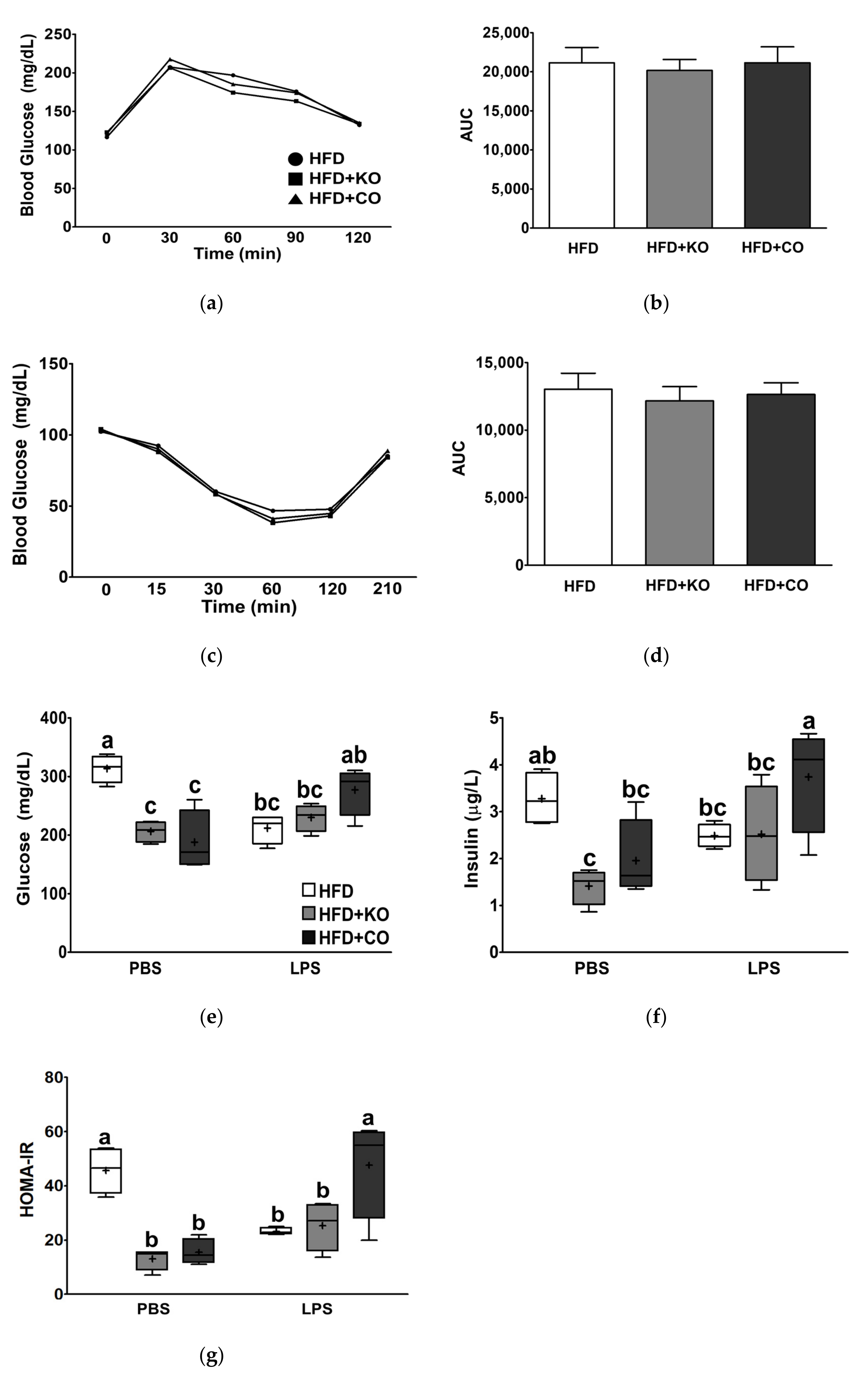
3.4. Effect of the Dietary Fat Replacement with KO or CO on Oral Glucose Tolerance (OGTT) and Insulin Tolerance Test (ITT)


3.5. Effect of the Dietary Fat Replacement with KO or CO on Serum Glucose and Insulin Levels

3.6. Effect of the Dietary Fat Replacement with KO or CO on Blood Fatty Acid Composition
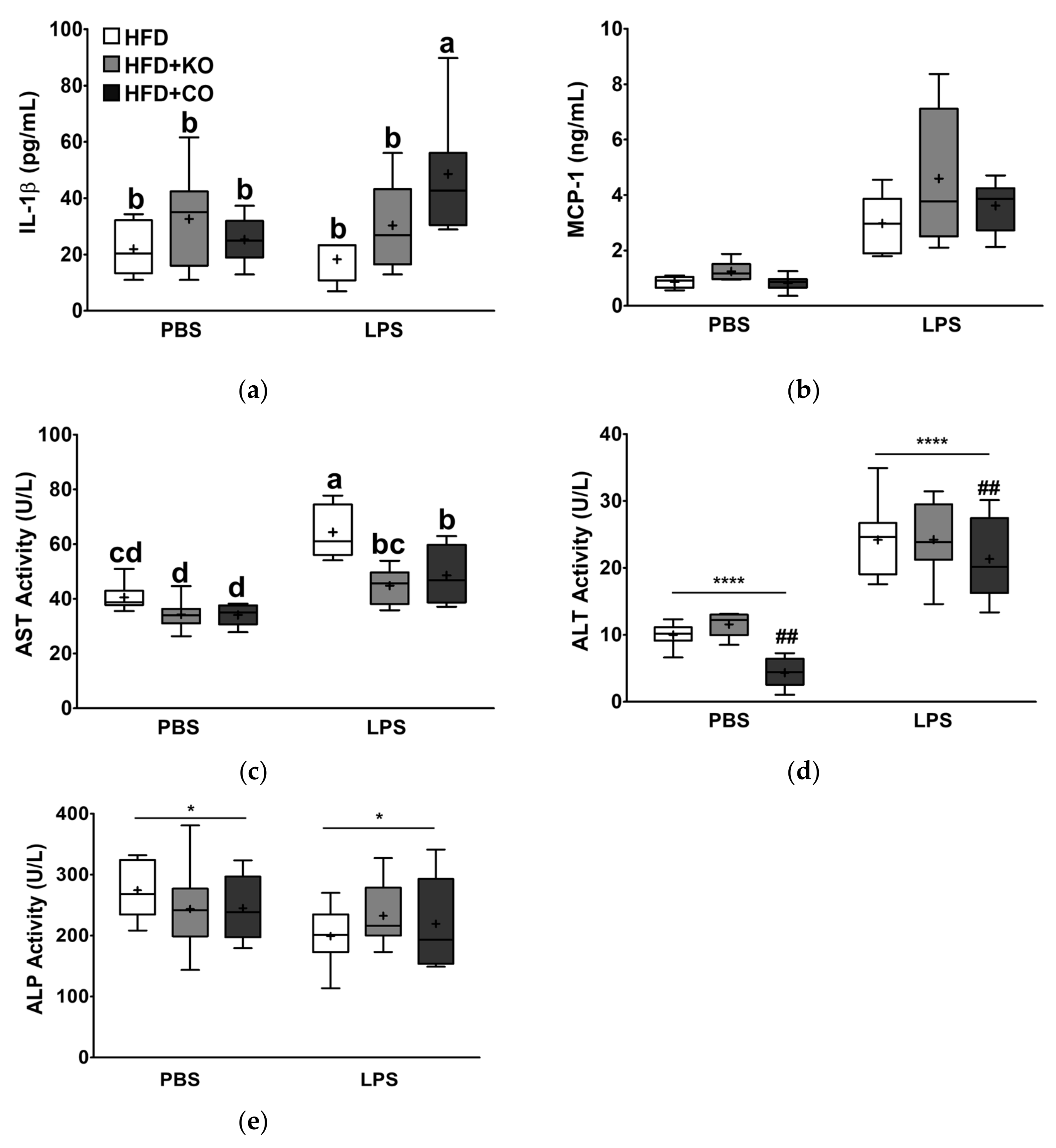
3.7. Effect of the Dietary Fat Replacement with KO or CO on Serum Proinflammatory Cytokines, and Chemokine Levels
3.8. Effect of the Dietary Fat Replacement with KO or CO on Enzyme Profiles Related to Hepatic Function
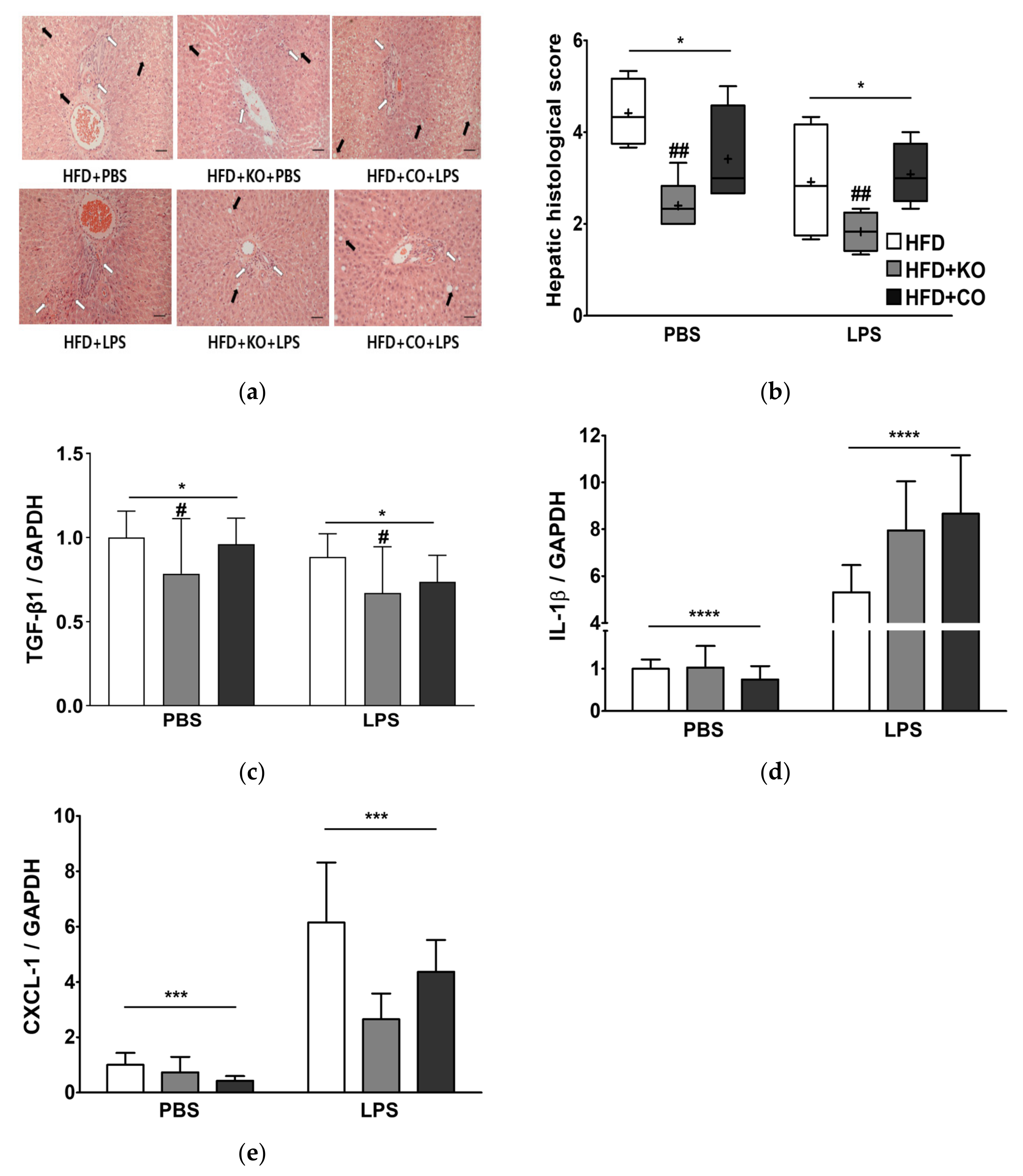
3.9. Effect of the Dietary Fat Replacement with KO or CO on Hepatotoxicity and Hepatic Fibrosis
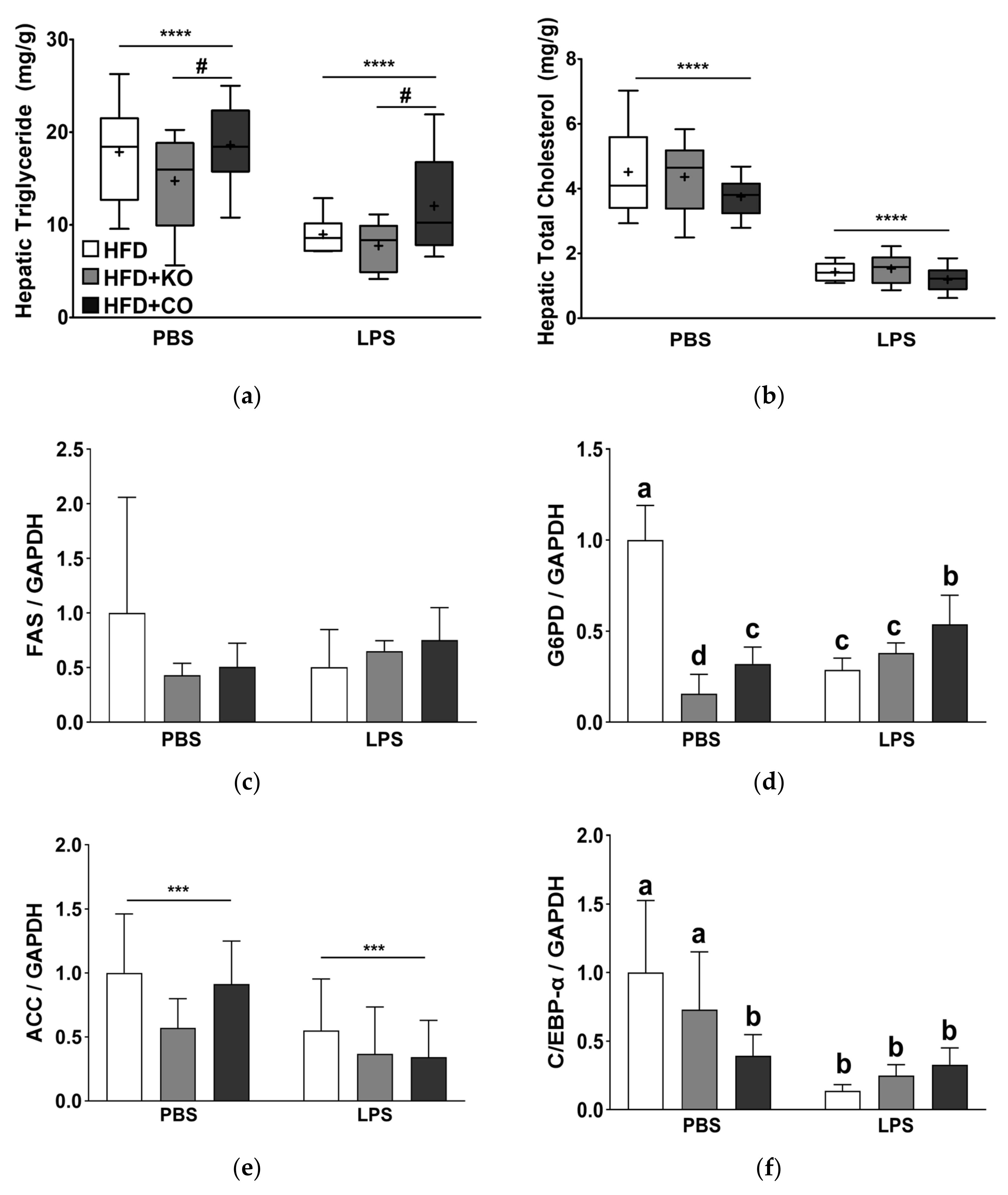
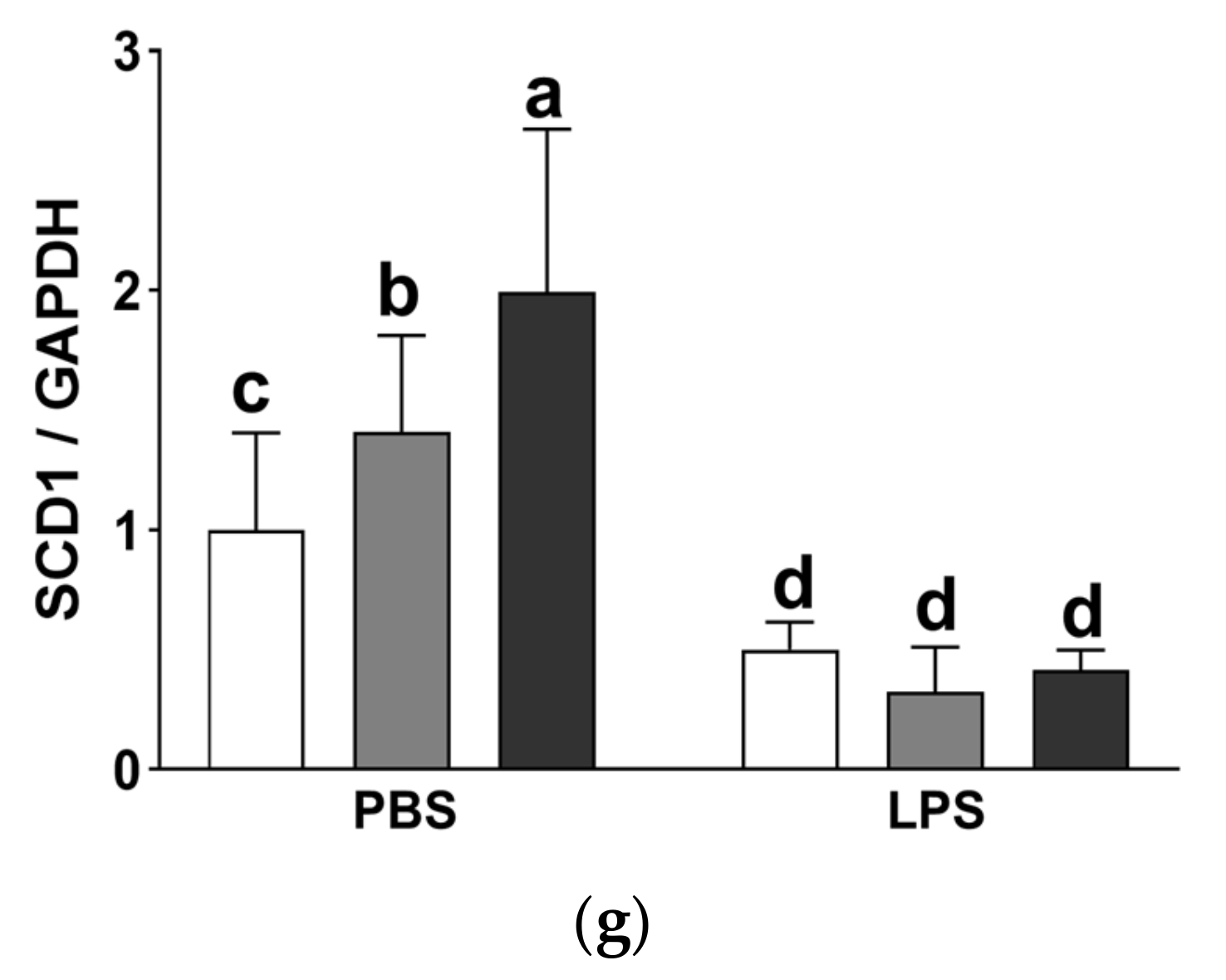
3.10. Effect of the Dietary Fat Replacement with KO or CO on Fat Accumulation and mRNA Expression of Lipogenesis-related Genes in the Liver
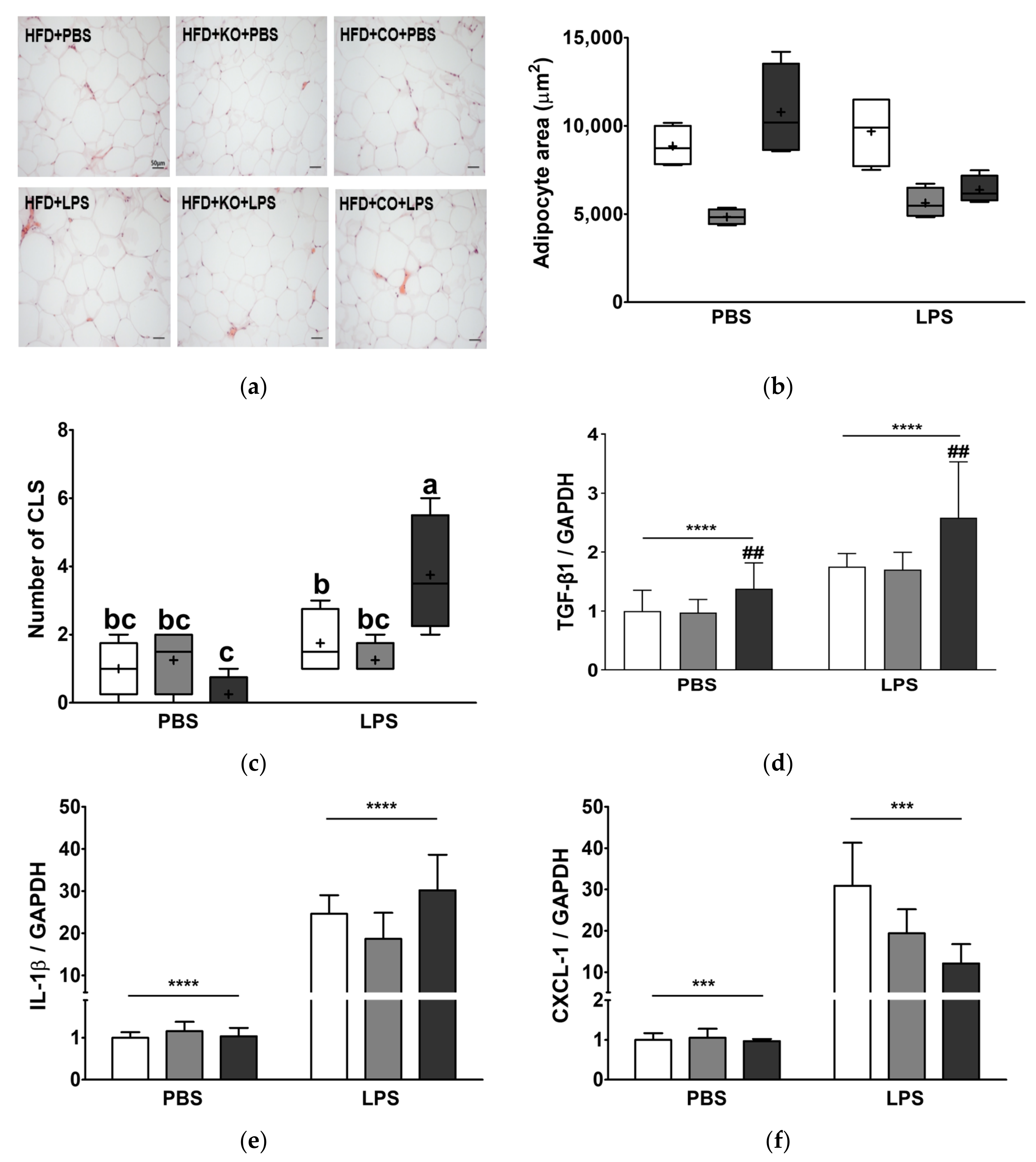
3.11. Effect of the Dietary Fat Replacement with KO or CO on Hypertrophy and Macrophage Infiltration in Epididymal Adipose Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kelly, T.; Yang, W.; Chen, C.-S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. 2008, 32, 1431–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 6 January 2022).
- Center for Disease Control and Prevention. Prevalence of Obesity and Severe Obesity Among Adults: United States, 2017–2018. Available online: https://www.cdc.gov/nchs/products/databriefs/db360.htm (accessed on 6 January 2022).
- Collaboration, N.R.F. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19·2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef] [Green Version]
- Muscogiuri, G.; Barrea, L.; Savastano, S.; Colao, A. Nutritional recommendations for CoVID-19 quarantine. Eur. J. Clin. Nutr. 2020, 74, 850–851. [Google Scholar] [CrossRef] [PubMed]
- Aghili, S.M.M.; Ebrahimpur, M.; Arjmand, B.; Shadman, Z.; Sani, M.P.; Qorbani, M.; Larijani, B.; Payab, M. Obesity in COVID-19 era, implications for mechanisms, comorbidities, and prognosis: A review and meta-analysis. Int. J. Obes. 2021, 45, 998–1016. [Google Scholar] [CrossRef]
- Shin, S.; Ajuwon, K.M. Lipopolysaccharide Alters Thermogenic and Inflammatory Genes in White Adipose Tissue in Mice Fed Diets with Distinct 18-Carbon Fatty-Acid Composition. Lipids 2018, 53, 885–896. [Google Scholar] [CrossRef]
- Camp, H.S.; Ren, D.; Leff, T. Adipogenesis and fat-cell function in obesity and diabetes. Trends Mol. Med. 2002, 8, 442–447. [Google Scholar] [CrossRef]
- Sun, J.; Zhou, W.; Gu, T.; Zhu, D.; Bi, Y. A retrospective study on association between obesity and cardiovascular risk diseases with aging in Chinese adults. Sci. Rep. 2018, 8, 5806. [Google Scholar] [CrossRef] [Green Version]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J. Primary prevention of cardiovascular disease with a Mediterranean diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Masoodi, M.; Kuda, O.; Rossmeisl, M.; Flachs, P.; Kopecky, J. Lipid signaling in adipose tissue: Connecting inflammation & metabolism. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 503–518. [Google Scholar] [CrossRef]
- Evert, A.B.; Boucher, J.L.; Cypress, M.; Dunbar, S.A.; Franz, M.J.; Mayer-Davis, E.J.; Neumiller, J.J.; Nwankwo, R.; Verdi, C.L.; Urbanski, P. Nutrition therapy recommendations for the management of adults with diabetes. Diabetes Care 2014, 37, S120–S143. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.G.; Ford, N.A.; Hu, F.B.; Zelman, K.M.; Mozaffarian, D.; Kris-Etherton, P.M. A healthy approach to dietary fats: Understanding the science and taking action to reduce consumer confusion. Nutr. J. 2017, 16, 53. [Google Scholar] [CrossRef]
- White, B. Dietary fatty acids. Am. Fam. Physician 2009, 80, 345–350. [Google Scholar]
- World Health Organization. Healthy Diet. Available online: https://www.who.int/publications/m/item/healthy-diet-factsheet394 (accessed on 6 January 2022).
- World Health Organization. Diet, Nutrition, and the Prevention of Chronic Diseases: Report of a Joint WHO/FAO Expert Consultation; World Health Organization: Geneva, Switzerland, 2003; Volume 916. [Google Scholar]
- Ulven, S.M.; Holven, K.B. Comparison of bioavailability of krill oil versus fish oil and health effect. Vasc. Health Risk Manag. 2015, 11, 511. [Google Scholar] [CrossRef] [Green Version]
- Kwantes, J.M.; Grundmann, O. A brief review of krill oil history, research, and the commercial market. J. Diet. Suppl. 2015, 12, 23–35. [Google Scholar] [CrossRef]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef] [Green Version]
- Salem, N.; Kuratko, C.N. A reexamination of krill oil bioavailability studies. Lipids Health Dis. 2014, 13, 137. [Google Scholar] [CrossRef] [Green Version]
- Gigliotti, J.C.; Davenport, M.P.; Beamer, S.K.; Tou, J.C.; Jaczynski, J. Extraction and characterisation of lipids from Antarctic krill (Euphausia superba). Food Chem. 2011, 125, 1028–1036. [Google Scholar] [CrossRef]
- Yang, G.; Lee, J.; Lee, S.; Kwak, D.; Choe, W.; Kang, I.; Kim, S.S.; Ha, J. Krill oil supplementation improves dyslipidemia and lowers body weight in mice fed a high-fat diet through activation of AMP-activated protein kinase. J. Med. Food 2016, 19, 1120–1129. [Google Scholar] [CrossRef]
- Kim, H.-D.; Lee, S.-B.; Ko, S.-C.; Jung, W.-K.; Kim, Y.-M.; Kim, S.-B. Anti-inflammatory effect of ozonated krill (Euphausia superba) oil in lipopolysaccharide-stimulated RAW 264.7 macrophages. Fish. Aquat. Sci. 2018, 21, 15. [Google Scholar] [CrossRef]
- Kim, J.H.; Meng, H.W.; He, M.T.; Choi, J.M.; Lee, D.; Cho, E.J. Krill Oil Attenuates Cognitive Impairment by the Regulation of Oxidative Stress and Neuronal Apoptosis in an Amyloid β-Induced Alzheimer’s Disease Mouse Model. Molecules 2020, 25, 3942. [Google Scholar] [CrossRef]
- Ierna, M.; Kerr, A.; Scales, H.; Berge, K.; Griinari, M. Supplementation of diet with krill oil protects against experimental rheumatoid arthritis. BMC Musculoskelet. Disord. 2010, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Cheong, L.-Z.; Sun, T.; Li, Y.; Zhou, J.; Lu, C.; Li, Y.; Huang, Z.; Su, X. Dietary krill oil enhances neurocognitive functions and modulates proteomic changes in brain tissues of d-galactose induced aging mice. Food Funct. 2017, 8, 2038–2045. [Google Scholar] [CrossRef]
- Cui, C.; Li, Y.; Gao, H.; Zhang, H.; Han, J.; Zhang, D.; Li, Y.; Zhou, J.; Lu, C.; Su, X. Modulation of the gut microbiota by the mixture of fish oil and krill oil in high-fat diet-induced obesity mice. PLoS ONE 2017, 12, e0186216. [Google Scholar] [CrossRef]
- Ferramosca, A.; Conte, A.; Burri, L.; Berge, K.; De Nuccio, F.; Giudetti, A.M.; Zara, V. A krill oil supplemented diet suppresses hepatic steatosis in high-fat fed rats. PLoS ONE 2012, 7, e38797. [Google Scholar] [CrossRef] [Green Version]
- Köhler, A.; Sarkkinen, E.; Tapola, N.; Niskanen, T.; Bruheim, I. Bioavailability of fatty acids from krill oil, krill meal and fish oil in healthy subjects–a randomized, single-dose, cross-over trial. Lipids Health Dis. 2015, 14, 19. [Google Scholar] [CrossRef] [Green Version]
- Kroupova, P.; van Schothorst, E.M.; Keijer, J.; Bunschoten, A.; Vodicka, M.; Irodenko, I.; Oseeva, M.; Zacek, P.; Kopecky, J.; Rossmeisl, M. Omega-3 phospholipids from krill oil enhance intestinal fatty acid oxidation more effectively than omega-3 triacylglycerols in high-fat diet-fed obese mice. Nutrients 2020, 12, 2037. [Google Scholar] [CrossRef]
- Rundblad, A.; Holven, K.B.; Bruheim, I.; Myhrstad, M.C.; Ulven, S.M. Effects of krill oil and lean and fatty fish on cardiovascular risk markers: A randomised controlled trial. J. Nutr. Sci. 2018, 7, e3. [Google Scholar] [CrossRef] [Green Version]
- Sistilli, G.; Kalendova, V.; Cajka, T.; Irodenko, I.; Bardova, K.; Oseeva, M.; Zacek, P.; Kroupova, P.; Horakova, O.; Lackner, K. Krill oil supplementation reduces exacerbated hepatic steatosis induced by thermoneutral housing in mice with diet-induced obesity. Nutrients 2021, 13, 437. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, L.; Chen, H.; Feng, R.; Cao, P.; Liu, Y. Effects of Antarctic krill oil on lipid and glucose metabolism in C57BL/6J mice fed with high fat diet. Lipids Health Dis. 2017, 16, 218. [Google Scholar] [CrossRef] [Green Version]
- Tandy, S.; Chung, R.W.; Wat, E.; Kamili, A.; Berge, K.; Griinari, M.; Cohn, J.S. Dietary krill oil supplementation reduces hepatic steatosis, glycemia, and hypercholesterolemia in high-fat-fed mice. J. Agric. Food Chem. 2009, 57, 9339–9345. [Google Scholar] [CrossRef]
- Basson, A.R.; Chen, C.; Sagl, F.; Trotter, A.; Bederman, I.; Gomez-Nguyen, A.; Ilic, S.; Cominelli, F.; Rodriguez-Palacios, A. Regulation of intestinal inflammation by dietary fats. Front. Immunol. 2020, 11, 3639. [Google Scholar] [CrossRef] [PubMed]
- Amarasiri, W.; Dissanayake, A. Coconut fats. Ceylon Med. J. 2006, 51, 47–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, K.C.; Hasse, W.; Dicklin, M.R.; Bell, M.; Buggia, M.A.; Cassens, M.E.; Eren, F. Corn oil lowers plasma cholesterol compared with coconut oil in adults with above-desirable levels of cholesterol in a randomized crossover trial. J. Nutr. 2018, 148, 1556–1563. [Google Scholar] [CrossRef] [PubMed]
- Marina, A.; Man, Y.C.; Amin, I. Virgin coconut oil: Emerging functional food oil. Trends Food Sci. Technol. 2009, 20, 481–487. [Google Scholar] [CrossRef]
- Papamandjaris, A.A.; MacDougall, D.E.; Jones, P.J. Medium chain fatty acid metabolism and energy expenditure: Obesity treatment implications. Life Sci. 1998, 62, 1203–1215. [Google Scholar] [CrossRef]
- Nagao, K.; Yanagita, T. Medium-chain fatty acids: Functional lipids for the prevention and treatment of the metabolic syndrome. Pharmacol. Res. 2010, 61, 208–212. [Google Scholar] [CrossRef]
- Lyte, J.M.; Gabler, N.K.; Hollis, J.H. Postprandial serum endotoxin in healthy humans is modulated by dietary fat in a randomized, controlled, cross-over study. Lipids Health Dis. 2016, 15, 186. [Google Scholar] [CrossRef] [Green Version]
- Remick, D.G.; Newcomb, D.E.; Bolgos, G.L.; Call, D.R. Comparison of the mortality and inflammatory response of two models of sepsis: Lipopolysaccharide vs. cecal ligation and puncture. Shock 2000, 13, 110–116. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, R.; Wang, W.; Xu, F.; Xi, Y.; Brown, R.A.; Zhang, H.; Shi, L.; Zhu, D.; Gong, D.-W. Fc-apelin fusion protein attenuates lipopolysaccharide-induced liver injury in mice. Sci. Rep. 2018, 8, 11428. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Lv, X.-D.; Wang, G.-E.; Li, Y.-F.; Kurihara, H.; He, R.-R. Anti-inflammatory effects of anthocyanins-rich extract from bilberry (Vaccinium myrtillus L.) on croton oil-induced ear edema and Propionibacterium acnes plus LPS-induced liver damage in mice. Int. J. Food Sci. Nutr. 2014, 65, 594–601. [Google Scholar] [CrossRef]
- Nabil-Adam, A.; Shreadah, M.A. Red algae natural products for prevention of lipopolysaccharides (LPS)-induced liver and kidney inflammation and injuries. Biosci. Rep. 2021, 41, BSR20202022. [Google Scholar] [CrossRef]
- Niu, X.; Xing, W.; Li, W.; Fan, T.; Hu, H.; Li, Y. Isofraxidin exhibited anti-inflammatory effects in vivo and inhibited TNF-α production in LPS-induced mouse peritoneal macrophages in vitro via the MAPK pathway. Int. Immunopharmacol. 2012, 14, 164–171. [Google Scholar] [CrossRef]
- Tjahjono, Y.; Karnati, S.; Foe, K.; Anggara, E.; Gunawan, Y.N.; Wijaya, H.; Suyono, H.; Esar, S.Y.; Hadinugroho, W.; Wihadmadyatami, H. Anti-inflammatory activity of 2-((3-(chloromethyl) benzoyl) oxy) benzoic acid in LPS-induced rat model. Prostaglandins Other Lipid Mediat. 2021, 154, 106549. [Google Scholar] [CrossRef]
- Son, H.-K.; Xiang, H.; Park, S.; Lee, J.; Lee, J.-J.; Jung, S.; Ha, J.-H. Partial Replacement of Dietary Fat with Polyunsaturated Fatty Acids Attenuates the Lipopolysaccharide-Induced Hepatic Inflammation in Sprague-Dawley Rats Fed a High-Fat Diet. Int. J. Environ. Res. Public Health 2021, 18, 10986. [Google Scholar] [CrossRef]
- Lange, K.W.; Nakamura, Y.; Gosslau, A.M.; Li, S. Are there serious adverse effects of omega-3 polyunsaturated fatty acid supplements? J. Food Bioact. 2019, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Nestel, P.; Shige, H.; Pomeroy, S.; Cehun, M.; Abbey, M.; Raederstorff, D. The n−3 fatty acids eicosapentaenoic acid and docosahexaenoic acid increase systemic arterial compliance in humans. Am. J. Clin. Nutr. 2002, 76, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Harris, W.; Polreis, J. Measurement of the omega-3 index in dried blood spots. Ann. Clin. Lab. Res. 2016, 4, 137. [Google Scholar] [CrossRef]
- Rosenfeld, L. Lipoprotein analysis. Early methods in the diagnosis of atherosclerosis. Arch. Pathol. Lab. Med. 1989, 113, 1101–1110. [Google Scholar]
- Haffner, S.M.; Miettinen, H.; Stern, M.P. The homeostasis model in the San Antonio heart study. Diabetes Care 1997, 20, 1087–1092. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [Green Version]
- Sollof, L.; Futenberg, H.; Lacko, A. Serum cholesterol esterfication in patients with coronary artery disease. Am. Heart J. 1973, 85, 153–161. [Google Scholar] [CrossRef]
- Bray, G.A.; Krauss, R.M. Overfeeding of polyunsaturated versus saturated fatty acids reduces ectopic fat. Diabetes 2014, 63, 2222–2224. [Google Scholar] [CrossRef] [Green Version]
- Moussavi, N.; Gavino, V.; Receveur, O. Could the quality of dietary fat, and not just its quantity, be related to risk of obesity? Obesity 2008, 16, 7–15. [Google Scholar] [CrossRef]
- Herbert, D.; Franz, S.; Popkova, Y.; Anderegg, U.; Schiller, J.; Schwede, K.; Lorz, A.; Simon, J.C.; Saalbach, A. High-fat diet exacerbates early psoriatic skin inflammation independent of obesity: Saturated fatty acids as key players. J. Investig. Dermatol. 2018, 138, 1999–2009. [Google Scholar] [CrossRef] [Green Version]
- Mensink, R.P. Effects of Saturated Fatty Acids on Serum Lipids and Lipoproteins: A Systematic Review and Regression Analysis; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Li, J.; Li, F.R.; Wei, D.; Jia, W.; Kang, J.X.; Stefanovic-Racic, M.; Dai, Y.; Zhao, A.Z. Endogenous ω-3 Polyunsaturated Fatty Acid Production Confers Resistance to Obesity, Dyslipidemia, and Diabetes in Mice. Mol. Endocrinol. 2014, 28, 1316–1328. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, Y.B.; Hein, G.; Chicco, A. Metabolic syndrome: Effects of n-3 PUFAs on a model of dyslipidemia, insulin resistance and adiposity. Lipids 2007, 42, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-C.; Tsai, T.-H.; Chuang, L.-T.; Li, Y.-Y.; Zouboulis, C.C.; Tsai, P.-J. Anti-bacterial and anti-inflammatory properties of capric acid against Propionibacterium acnes: A comparative study with lauric acid. J. Dermatol. Sci. 2014, 73, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Temme, E.; Mensink, R.P.; Hornstra, G. Comparison of the effects of diets enriched in lauric, palmitic, or oleic acids on serum lipids and lipoproteins in healthy women and men. Am. J. Clin. Nutr. 1996, 63, 897–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, N.F.B.; de Queiroz, T.M.; de Almeida Travassos, R.; Magnani, M.; de Andrade Braga, V. Acute treatment with lauric acid reduces blood pressure and oxidative stress in spontaneously hypertensive rats. Basic Clin. Pharmacol. Toxicol. 2017, 120, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Briggs, M.A.; Petersen, K.S.; Kris-Etherton, P.M. Saturated fatty acids and cardiovascular disease: Replacements for saturated fat to reduce cardiovascular risk. Healthcare 2017, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Brennan, L.; Bloomfield, J.F.; Duff, D.J.; McNulty, B.A.; Flynn, A.; Walton, J.; Gibney, M.J.; Nugent, A.P. Adiposity associated plasma linoleic acid is related to demographic, metabolic health and haplotypes of FADS1/2 genes in Irish adults. Mol. Nutr. Food Res. 2018, 62, 1700785. [Google Scholar] [CrossRef]
- Antonini, J.M.; Kodali, V.; Meighan, T.G.; Roach, K.A.; Roberts, J.R.; Salmen, R.; Boyce, G.R.; Zeidler-Erdely, P.C.; Kashon, M.; Erdely, A. Effect of age, high-fat diet, and rat strain on serum biomarkers and telomere length and global DNA methylation in peripheral blood mononuclear cells. Sci. Rep. 2019, 9, 1996. [Google Scholar] [CrossRef]
- Hintze, K.J.; Benninghoff, A.D.; Cho, C.E.; Ward, R.E. Modeling the western diet for preclinical investigations. Adv. Nutr. 2018, 9, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Dullo, A.; Girardier, L. Influence of dietary composition on energy expenditure during recovery of body weight in the rat: Implications for catch-up growth and obesity relapse. Metabolism 1992, 41, 1336–1342. [Google Scholar] [CrossRef]
- Famurewa, A.C.; Akunna, G.G.; Nwafor, J.; Chukwu, O.C.; Ekeleme-Egedigwe, C.A.; Oluniran, J.N. Nephroprotective activity of virgin coconut oil on diclofenac-induced oxidative nephrotoxicity is associated with antioxidant and anti-inflammatory effects in rats. Avicenna J. Phytomed. 2020, 10, 316. [Google Scholar]
- Hima, L.; Pratap, U.P.; Karrunanithi, S.; Ravichandran, K.A.; Vasantharekha, R.; ThyagaRajan, S. Virgin coconut oil supplementation in diet modulates immunity mediated through survival signaling pathways in rats. J. Complementary Integr. Med. 2020, 17, 1–11. [Google Scholar] [CrossRef]
- Sreevallabhan, S.; Mohanan, R.; Jose, S.P.; Sukumaran, S.; Jagmag, T.; Tilwani, J.; Kulkarni, A. Hepatoprotective effect of essential phospholipids enriched with virgin coconut oil (Phoscoliv) on paracetamol-induced liver toxicity. J. Food Biochem. 2021, 45, e13606. [Google Scholar] [CrossRef]
- Alghamdi, B.S. The Effect of Short-Term Feeding of a High-Coconut Oil or High-Fat Diet on Neuroinflammation and the Performance of an Object–Place Task in Rats. Neurochem. Res. 2021, 46, 287–298. [Google Scholar] [CrossRef]
- Khan, H.U.; Aamir, K.; Jusuf, P.R.; Sethi, G.; Sisinthy, S.P.; Ghildyal, R.; Arya, A. Lauric acid ameliorates lipopolysaccharide (LPS)-induced liver inflammation by mediating TLR4/MyD88 pathway in Sprague Dawley (SD) rats. Life Sci. 2021, 265, 118750. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, W.J.; Olayaki, L.A.; Abdussalam, T.A.; Toriola, A.P.; Olowu, A.B.; Yakub, A.J.; Raji, A.O. Investigation of the effects of dietary modification in experimental obesity: Low dose of virgin coconut oil has a potent therapeutic value. Biomed. Pharmacother. 2020, 126, 110110. [Google Scholar] [CrossRef] [PubMed]
- Famurewa, A.C.; Maduagwuna, E.K.; Folawiyo, A.M.; Besong, E.E.; Eteudo, A.N.; Famurewa, O.A.; Ejezie, F.E. Antioxidant, anti-inflammatory, and antiapoptotic effects of virgin coconut oil against antibiotic drug gentamicin-induced nephrotoxicity via the suppression of oxidative stress and modulation of iNOS/NF-ĸB/caspase-3 signaling pathway in Wistar rats. J. Food Biochem. 2020, 44, e13100. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.; Lee, M.; Bray, T. Weight gain of rats fed medium-chain triglycerides is less than rats fed long-chain triglycerides. Int. J. Obes. 1980, 4, 27–32. [Google Scholar]
- Geliebter, A.; Torbay, N.; Bracco, E.; Hashim, S.A.; Van Itallie, T.B. Overfeeding with medium-chain triglyceride diet results in diminished deposition of fat. Am. J. Clin. Nutr. 1983, 37, 1–4. [Google Scholar] [CrossRef]
- Cavaliere, G.; Trinchese, G.; Bergamo, P.; De Filippo, C.; Mattace Raso, G.; Gifuni, G.; Putti, R.; Moni, B.H.; Canani, R.B.; Meli, R. Polyunsaturated fatty acids attenuate diet induced obesity and insulin resistance, modulating mitochondrial respiratory uncoupling in rat skeletal muscle. PLoS ONE 2016, 11, e0149033. [Google Scholar] [CrossRef] [Green Version]
- Nagy, C.; Einwallner, E. Study of in vivo glucose metabolism in high-fat diet-fed mice using oral glucose tolerance test (OGTT) and insulin tolerance test (ITT). JoVE J. Vis. Exp. 2018, e56672. [Google Scholar] [CrossRef]
- Giknis, M.; Clifford, C. Clinical laboratory parameters for Crl: WI (Han) rats. Accel. Drug Dev. 2008, 1–14. [Google Scholar]
- Bonora, E.; Formentini, G.; Calcaterra, F.; Lombardi, S.; Marini, F.; Zenari, L.; Saggiani, F.; Poli, M.; Perbellini, S.; Raffaelli, A. HOMA-estimated insulin resistance is an independent predictor of cardiovascular disease in type 2 diabetic subjects: Prospective data from the Verona Diabetes Complications Study. Diabetes Care 2002, 25, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Wallace, D.F. The regulation of iron absorption and homeostasis. Clin. Biochem. Rev. 2016, 37, 51. [Google Scholar]
- Ghafoorunissa; Ibrahim, A.; Natarajan, S. Substituting dietary linoleic acid with α-linolenic acid improves insulin sensitivity in sucrose fed rats. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2005, 1733, 67–75. [Google Scholar] [CrossRef]
- Han, J.; Hamilton, J.A.; Kirkland, J.L.; Corkey, B.E.; Guo, W. Medium-chain oil reduces fat mass and down-regulates expression of adipogenic genes in rats. Obes. Res. 2003, 11, 734–744. [Google Scholar] [CrossRef]
- Wein, S.; Wolffram, S.; Schrezenmeir, J.; Gašperiková, D.; Klimeš, I.; Šeböková, E. Medium-chain fatty acids ameliorate insulin resistance caused by high-fat diets in rats. Diabetes/Metab. Res. Rev. 2009, 25, 185–194. [Google Scholar] [CrossRef]
- Liang, H.; Hussey, S.E.; Sanchez-Avila, A.; Tantiwong, P.; Musi, N. Effect of lipopolysaccharide on inflammation and insulin action in human muscle. PLoS ONE 2013, 8, e63983. [Google Scholar] [CrossRef]
- Bosomworth, N.J. Approach to identifying and managing atherogenic dyslipidemia: A metabolic consequence of obesity and diabetes. Can. Fam. Physician 2013, 59, 1169–1180. [Google Scholar]
- Choi, G.-h.; Jung, Y.-s.; Shin, H.-c. The effects of haedoksamul-tang on oxidative stress and hyperlipidemia in LPS-induced ICR mouse. J. Korean Med. 2016, 37, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Lassenius, M.I.; Pietiläinen, K.H.; Kaartinen, K.; Pussinen, P.J.; Syrjänen, J.; Forsblom, C.; Pörsti, I.; Rissanen, A.; Kaprio, J.; Mustonen, J. Bacterial endotoxin activity in human serum is associated with dyslipidemia, insulin resistance, obesity, and chronic inflammation. Diabetes Care 2011, 34, 1809–1815. [Google Scholar] [CrossRef] [Green Version]
- Wahby, M.M.; Yacout, G.; Kandeel, K.; Awad, D. LPS-induced oxidative inflammation and hyperlipidemia in male rats: The protective role of Origanum majorana extract. Beni-Suef Univ. J. Basic Appl. Sci. 2015, 4, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Wen, Y.; Li, J.; Zhang, H.; Liu, Y. Prenatal lipopolysaccharide exposure promotes dyslipidemia in the male offspring rats. Front. Physiol. 2018, 9, 542. [Google Scholar] [CrossRef]
- Famurewa, A.C.; Ekeleme-Egedigwe, C.A.; Nwali, S.C.; Agbo, N.N.; Obi, J.N.; Ezechukwu, G.C. Dietary supplementation with virgin coconut oil improves lipid profile and hepatic antioxidant status and has potential benefits on cardiovascular risk indices in normal rats. J. Diet. Suppl. 2018, 15, 330–342. [Google Scholar] [CrossRef]
- Nevin, K.; Rajamohan, T. Beneficial effects of virgin coconut oil on lipid parameters and in vitro LDL oxidation. Clin. Biochem. 2004, 37, 830–835. [Google Scholar] [CrossRef]
- Speliotes, E.K.; Balakrishnan, M.; Friedman, L.S.; Corey, K.E. Treatment of dyslipidemia in common liver diseases. Clin. Gastroenterol. Hepatol. 2018, 16, 1189–1196. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.-K.; Patra, M.C.; Shin, H.-J.; Gui, X.; Achek, A.; Panneerselvam, S.; Kim, D.-J.; Song, S.-J.; Hong, R.; Kim, K.S. A cell-penetrating peptide blocks Toll-like receptor-mediated downstream signaling and ameliorates autoimmune and inflammatory diseases in mice. Exp. Mol. Med. 2019, 51, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Li, S.; Li, J.; Wang, J.; Zhang, R.; Zhou, Y.; Yin, Q.; Zheng, Y.; Wang, F.; Xia, Y. Effects of omega-3 fatty acid in nonalcoholic fatty liver disease: A meta-analysis. Gastroenterol. Res. Pract. 2016, 2016, 1459790. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, H.; Tehilla, M.; Attal-Singer, J.; Bruck, R.; Luzzatti, R.; Singer, P. The therapeutic potential of long-chain omega-3 fatty acids in nonalcoholic fatty liver disease. Clin. Nutr. 2011, 30, 6–19. [Google Scholar] [CrossRef]
- Spadaro, L.; Magliocco, O.; Spampinato, D.; Piro, S.; Oliveri, C.; Alagona, C.; Papa, G.; Rabuazzo, A.; Purrello, F. Effects of n-3 polyunsaturated fatty acids in subjects with nonalcoholic fatty liver disease. Dig. Liver Dis. 2008, 40, 194–199. [Google Scholar] [CrossRef]
- Liu, L.; Mei, M.; Yang, S.; Li, Q. Roles of chronic low-grade inflammation in the development of ectopic fat deposition. Mediat. Inflamm. 2014, 2014, 418185. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Altintas, M.M.; Rossetti, M.A.; Nayer, B.; Puig, A.; Zagallo, P.; Ortega, L.M.; Johnson, K.B.; McNamara, G.; Reiser, J.; Mendez, A.J. Apoptosis, mastocytosis, and diminished adipocytokine gene expression accompany reduced epididymal fat mass in long-standing diet-induced obese mice. Lipids Health Dis. 2011, 10, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, P.; Morrison, M.; Wielinga, P.; Van Duyvenvoorde, W.; Kooistra, T.; Kleemann, R. Surgical removal of inflamed epididymal white adipose tissue attenuates the development of non-alcoholic steatohepatitis in obesity. Int. J. Obes. 2016, 40, 675–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafontan, M. Adipose tissue and adipocyte dysregulation. Diabetes Metab. 2014, 40, 16–28. [Google Scholar] [CrossRef]
- Liang, W.; Tonini, G.; Mulder, P.; Kelder, T.; van Erk, M.; van den Hoek, A.M.; Mariman, R.; Wielinga, P.Y.; Baccini, M.; Kooistra, T. Coordinated and interactive expression of genes of lipid metabolism and inflammation in adipose tissue and liver during metabolic overload. PLoS ONE 2013, 8, e75290. [Google Scholar] [CrossRef] [Green Version]
- Duval, C.; Thissen, U.; Keshtkar, S.; Accart, B.; Stienstra, R.; Boekschoten, M.V.; Roskams, T.; Kersten, S.; Müller, M. Adipose tissue dysfunction signals progression of hepatic steatosis towards nonalcoholic steatohepatitis in C57BL/6 mice. Diabetes 2010, 59, 3181–3191. [Google Scholar] [CrossRef] [Green Version]
- Gart, E.; Salic, K.; Morrison, M.C.; Caspers, M.; van Duyvenvoorde, W.; Heijnk, M.; Giera, M.; Bobeldijk-Pastorova, I.; Keijer, J.; Storsve, A.B. Krill oil treatment increases distinct pufas and oxylipins in adipose tissue and liver and attenuates obesity-associated inflammation via direct and indirect mechanisms. Nutrients 2021, 13, 2836. [Google Scholar] [CrossRef]
- Awada, M.; Meynier, A.; Soulage, C.O.; Hadji, L.; Géloën, A.; Viau, M.; Ribourg, L.; Benoit, B.; Debard, C.; Guichardant, M. n-3 PUFA added to high-fat diets affect differently adiposity and inflammation when carried by phospholipids or triacylglycerols in mice. Nutr. Metab. 2013, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Saraswathi, V.; Kumar, N.; Gopal, T.; Bhatt, S.; Ai, W.; Ma, C.; Talmon, G.A.; Desouza, C. Lauric acid versus palmitic acid: Effects on adipose tissue inflammation, insulin resistance, and non-alcoholic fatty liver disease in obesity. Biology 2020, 9, 346. [Google Scholar] [CrossRef]
- Ströher, D.J.; de Oliveira, M.F.; Martinez-Oliveira, P.; Pilar, B.C.; Cattelan, M.D.P.; Rodrigues, E.; Bertolin, K.; Gonçalves, P.B.D.; Piccoli, J.d.C.E.; Manfredini, V. Virgin coconut oil associated with high-fat diet induces metabolic dysfunctions, adipose inflammation, and hepatic lipid accumulation. J. Med. Food 2020, 23, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Achek, A.; Yesudhas, D.; Choi, S. Toll-like receptors: Promising therapeutic targets for inflammatory diseases. Arch. Pharm. Res. 2016, 39, 1032–1049. [Google Scholar] [CrossRef]
- O’neill, L.A.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors—redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef]
- Zuany-Amorim, C.; Hastewell, J.; Walker, C. Toll-like receptors as potential therapeutic targets for multiple diseases. Nat. Rev. Drug Discov. 2002, 1, 797–807. [Google Scholar] [CrossRef]
- Laugerette, F.; Vors, C.; Peretti, N.; Michalski, M.-C. Complex links between dietary lipids, endogenous endotoxins and metabolic inflammation. Biochimie 2011, 93, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Raetzsch, C.F.; Brooks, N.L.; Alderman, J.M.; Moore, K.S.; Hosick, P.A.; Klebanov, S.; Akira, S.; Bear, J.E.; Baldwin, A.S.; Mackman, N. Lipopolysaccharide inhibition of glucose production through the Toll-like receptor-4, myeloid differentiation factor 88, and nuclear factor κb pathway. Hepatology 2009, 50, 592–600. [Google Scholar] [CrossRef] [Green Version]
- Imajo, K.; Fujita, K.; Yoneda, M.; Nozaki, Y.; Ogawa, Y.; Shinohara, Y.; Kato, S.; Mawatari, H.; Shibata, W.; Kitani, H. Hyperresponsivity to low-dose endotoxin during progression to nonalcoholic steatohepatitis is regulated by leptin-mediated signaling. Cell Metab. 2012, 16, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Tesfay, M.; Goldkamp, W.J.; Neuschwander-Tetri, B.A. NASH: The emerging most common form of chronic liver disease. Mo. Med. 2018, 115, 225. [Google Scholar]
- Wellen, K.E.; Hotamisligil, G.S. Inflammation, stress, and diabetes. J. Clin. Investig. 2005, 115, 1111–1119. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.-L.; Chen, H.; Wang, C.-L.; Liang, L. Pathogenesis of non-alcoholic fatty liver disease in children and adolescence: From “two hit theory” to “multiple hit model”. World J. Gastroenterol. 2018, 24, 2974. [Google Scholar] [CrossRef]
- Alwayn, I.P.; Gura, K.; Nosé, V.; Zausche, B.; Javid, P.; Garza, J.; Verbesey, J.; Voss, S.; Ollero, M.; Andersson, C. Omega-3 fatty acid supplementation prevents hepatic steatosis in a murine model of nonalcoholic fatty liver disease. Pediatric Res. 2005, 57, 445–452. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Montgomery, M.K.; Fiveash, C.E.; Osborne, B.; Cooney, G.J.; Bell-Anderson, K.; Turner, N. PPARα-independent actions of omega-3 PUFAs contribute to their beneficial effects on adiposity and glucose homeostasis. Sci. Rep. 2015, 4, 5538. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.W.; Kim, K.M.; Choi, Y.J.; An, H.J.; Lee, B.; Kim, D.H.; Lee, E.K.; Im, E.; Lee, J.; Im, D.S. The critical role played by endotoxin-induced liver autophagy in the maintenance of lipid metabolism during sepsis. Autophagy 2017, 13, 1113–1129. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | HFD | HFD + KO | HFD + CO | |
|---|---|---|---|---|
| Ingredient (g/kg) | ||||
| Casein | 220 | 220 | 220 | |
| L-cysteine | 3.4 | 3.4 | 3.4 | |
| Sucrose | 100 | 100 | 100 | |
| Corn starch | 160 | 160 | 160 | |
| Dextrose | 155 | 155 | 155 | |
| Cellulose | 58 | 58 | 58 | |
| Mineral mix 1 | 43 | 43 | 43 | |
| Vitamin mix 2 | 19 | 19 | 19 | |
| Choline bitartrate | 2.8 | 2.8 | 2.8 | |
| Western blend 3 | 184 | 134 | 134 | |
| Lard | 55 | 55 | 55 | |
| tert-Butylhydroquinone | 0.034 | 0.034 | 0.034 | |
| Krill oil | 50 | |||
| Coconut oil | 50 | |||
| Energy (kcal/g) | 4.78 | 4.78 | 4.78 | |
| Fat (kcal %) | 45% | 45% | 45% | |
| Groups | HFD | HFD + KO | HFD + CO | |
|---|---|---|---|---|
| Fatty Acid (% Total Fatty Acids) | ||||
| C4:0, Butyric acid | 0.49 ± 0.03 a | 0.36 ± 0.02 b | 0.35 ± 0.02 b | |
| C6:0, Caproic acid | 2.08 ± 0.11 a | 1.63 ± 0.09 b | 1.69 ± 0.09 b | |
| C8:0, Caprylic acid | 1.98 ± 0.10 b | 1.67 ± 0.08 b | 4.37 ± 0.22 a | |
| C10:0, Capric acid | 4.10 ± 0.21 b | 3.19 ± 0.16 c | 4.88 ± 0.25 a | |
| C12:0, Lauric acid | 6.20 ± 0.31 b | 4.65 ± 0.23 c | 15.91 ± 0.80 a | |
| C13:0, Tridecanoic acid | ND | ND | ND | |
| C14:0, Myristic acid | 11.53 ± 0.58 a | 9.83 ± 0.50 b | 12.04 ± 0.60 a | |
| C14:1, Myristoleic acid | 0.76 ± 0.04 b | 1.00 ± 0.05 a | 0.76 ± 0.04 b | |
| C15:0, Pentadecanoic acid | 1.88 ± 0.10 a | 1.53 ± 0.08 b | 1.26 ± 0.06 c | |
| C16:0, Palmitic acid | 24.79 ± 1.24 a | 21.97 ± 1.10 b | 18.96 ± 0.95 c | |
| C16:1, Palmitoleic acid | 2.5 ± 0.13 b | 3.78 ± 0.19 a | 2.05 ± 0.11 c | |
| C17:0, Heptadecanoic acid | 2.98 ± 0.15 b | 4.31 ± 0.22 a | 3.08 ± 0.16 b | |
| C18:0, Stearic acid | 13.5 ± 0.68 a | 10.99 ± 0.55 b | 11.13 ± 0.56 b | |
| C18:1, Oleic acid | 18.23 ± 0.92 | 16.64 ± 0.83 | 16.32 ± 0.82 | |
| C18:2, Linoleic acid | 6.63 ± 0.33 | 5.95 ± 0.30 | 6.25 ± 0.32 | |
| C18:3, Linolenic acid | 1.27 ± 0.07 b | 1.68 ± 0.08 a | ND | |
| C20:1, Eicosenoic acid | 0.36 ± 0.02 b | 0.48 ± 0.02 a | 0.37 ± 0.02 b | |
| C20:2, Eicosadienoic acid | 0.29 ± 0.02 b | 0.96 ± 0.05 a | 0.27 ± 0.02 b | |
| C20:3, Eicosatienoic acid | ND | 0.50 ± 0.03 a | ND | |
| C20:5, Eicosapentaenoic acid | 0.22 ± 0.01 b | 5.41 ± 0.28 a | 0.13 ± 0.01 b | |
| C22:1, Erucic acid | ND | 0.30 ± 0.02 a | ND | |
| C22:6, Docosahexaenoic acid | ND | 3.17 ± 0.16 a | ND | |
| Others 1 | 0.20 ± 0.01 a | ND | 0.18 ± 0.01 b | |
| SFAs | 69.55 ± 3.48 a | 60.14 ± 3.01 b | 73.67 ± 3.69 a | |
| MUFAs | 21.85 ± 1.10 ab | 22.20 ± 1.11 a | 19.50 ± 0.98 b | |
| PUFAs | 8.41 ± 0.42 b | 17.66 ± 0.89 a | 6.65 ± 0.34 c | |
| n-6 | 6.92 ± 0.35 | 6.91 ± 0.35 | 6.52 ± 0.33 | |
| n-3 | 1.50 ± 0.08 b | 10.75 ± 0.54 a | 0.13 ± 0.01 c | |
| Gene | Primer Sequence | |
|---|---|---|
| Il-1β | Forward | 5′-AAA AAT GCC TCG TGC TGT CT-3′ |
| Reverse | 5′-TCG TTG CTT GTC TCT CCT TG-3′ | |
| CXCL-1 | Forward | 5′-CCA CAC TCA AGA ATG GTC GC-3′ |
| Reverse | 5′-GTT GTC AGA AGC CAG CGT TC-3′ | |
| Fas | Forward | 5′-AAA AGG AAA GTA GAG TGT GC-3′ |
| Reverse | 5′-GAC ACA TTC TGT TCA CTA CAG-3′ | |
| G6pd | Forward | 5′-GTT TGG CAG CGG CAA CTA A-3′ |
| Reverse | 5′-GGC ATC ACC CTG GTA CAA CTC-3′ | |
| C/EBP-α | Forward | 5′-GCC AAG AAG TCG GTG GAT AA-3′ |
| Reverse | 5′-CGG TCA TTG TCA CTG GTC AA-3′ | |
| Scd1 | Forward | 5′-TGT TCG TCA GCA CCT TCT TG-3′ |
| Reverse | 5′-AGT TGA TGT GCC AGC GGT A-3′ | |
| Acc | Forward | 5′-CAA CGC CTT CAC ACC ACC TT-3′ |
| Reverse | 5′-AGC CCA TTA CTT CAT CAA AGA TCC T-3′ | |
| Tgf-β1 | Forward | 5′-ACC GAC CCT TCC TGC TCC TCA T-3′ |
| Reverse | 5′-GAT CCA CTT CCA ACC CAG GTC CT-3′ | |
| Gapdh | Forward | 5′-CTG TGT CTT TCC GCT GTT TTC-3′ |
| Reverse | 5′-TGT GCT GTG CTT ATG GTC TCA-3′ |
| Parameter | Factor, p Values | ||
|---|---|---|---|
| LPS | Diet | LPS X Diet | |
| Relative liver and adipose tissue weights | |||
| Liver | ** p < 0.01 | ns p = 0.931 | ns p = 0.057 |
| WAT | ns p = 0.832 | * p < 0.05 | ns p = 0.884 |
| EAT | ns p = 0.509 | ns p = 0.079 | ns p = 0.926 |
| MAT | ns p = 0.555 | ns p = 0.265 | ns p = 0.673 |
| RAT | ns p = 0.995 | ** p < 0.01 | ns p = 0.508 |
| PAT | ns p = 0.485 | ns p = 0.134 | ns p = 0.925 |
| Serum lipid levels | |||
| Triglyceride | ** p < 0.01 | **** p < 0.0001 | **** p < 0.0001 |
| Total cholesterol | **** p < 0.0001 | * p < 0.05 | ns p = 0.589 |
| HDL-cholesterol | ** p < 0.01 | ns p = 0.095 | ns p = 0.227 |
| Non-HDL-cholesterol | **** p < 0.0001 | * p < 0.05 | ns p = 0.735 |
| Atherogenic coefficient | **** p < 0.0001 | *** p < 0.001 | ns p = 0.997 |
| Cardiac risk factor | **** p < 0.0001 | *** p < 0.001 | ns p = 0.997 |
| Serum glucose and insulin levels | |||
| Glucose | ns p = 0.782 | * p < 0.05 | *** p < 0.001 |
| Insulin | * p < 0.05 | * p < 0.05 | * p < 0.05 |
| HOMA-IR | ns p = 0.078 | * p < 0.05 | *** p < 0.001 |
| Fatty acid levels in whole blood | |||
| C14:0, Myristic acid | ns p = 0.428 | ns p = 0.091 | ns p = 0.658 |
| C16:0, Palmitic acid | ns p = 0.199 | ns p = 0.767 | ns p = 0.28 |
| C16:1, Palmitoleic acid | ns p = 0.311 | ns p = 0.196 | ns p = 0.397 |
| C18:0, Stearic acid | ns p = 0.643 | * p < 0.05 | ns p = 0.35 |
| C18:1, Oleic acid | ns p = 0.26 | ns p = 0.055 | ns p = 0.353 |
| C18:2, Linoleic acid | ns p = 0.908 | * p < 0.05 | ns p = 0.105 |
| C18:3n-3, α-linolenic acid | ns p = 0.499 | ns p = 0.336 | ns p = 0.636 |
| C18:3n-6, γ-linolenic acid | ns p = 0.276 | ns p = 0.287 | ns p = 0.195 |
| C20:1, Eicosenoic acid | ns p = 0.885 | ns p = 0.155 | ns p = 0.700 |
| C20:2, Eicosadienoic acid | ns p = 0.274 | ns p = 0.23 | ns p = 0.402 |
| C20:3, Dihomo-γ-linolenic acid | ns p = 0.28 | ns p = 0.817 | * p < 0.05 |
| C20:4, Aachidonic acid | ns p = 0.941 | *** p < 0.001 | ns p = 0.111 |
| C20:5, Eicosapentaenoic acid | * p < 0.05 | *** p < 0.001 | * p < 0.05 |
| C22:4, Docosatetraenoic acid | ns p = 0.335 | *** p < 0.001 | ns p = 0.405 |
| C22:5, Docosapentaenoic acid | ns p = 0.979 | *** p < 0.001 | ns p = 0.67 |
| C22:6n-3, Docosahexaenoic acid | * p < 0.05 | *** p < 0.001 | ns p = 0.417 |
| C22:6n-6, Docosapentaenoic acid | ns p = 0.716 | *** p < 0.001 | ns p = 0.288 |
| C24:0, Lignoceric acid | ns p = 0.504 | ns p = 0.525 | ns p = 0.899 |
| C24:1, Nervonic acid | ns p = 0.335 | * p < 0.05 | ns p = 0.552 |
| Others | ns p = 0.743 | ** p < 0.01 | ns p = 0.29 |
| SFAs | ns p = 0.169 | ns p = 0.875 | ns p = 0.269 |
| MUFAs | ns p = 0.402 | ns p = 0.057 | ns p = 0.356 |
| PUFAs | ns p = 0.877 | * p < 0.05 | ns p = 0.464 |
| n-6 | ns p = 0.904 | *** p < 0.001 | ns p = 0.404 |
| n-3 | ns p = 0.609 | *** p < 0.001 | ns p = 0.891 |
| n-6/n-3 | ns p = 0.385 | *** p < 0.001 | ns p = 0.780 |
| AA/EPA | ns p = 0.879 | *** p < 0.001 | ns p = 0.993 |
| Serum cytokine levels | |||
| IL-1β | ns p = 0.219 | * p < 0.05 | * p < 0.05 |
| MCP-1 | **** p < 0.0001 | ns p = 0.056 | ns p = 0.338 |
| Serum levels of hepatic function parameter | |||
| AST | **** p < 0.0001 | **** p < 0.0001 | * p < 0.05 |
| ALT | **** p < 0.0001 | ** p < 0.01 | ns p = 0.357 |
| ALP | * p < 0.05 | ns p = 0.958 | ns p = 0.282 |
| Hepatic histology, inflammatory and fibrosis related gene expressions | |||
| Hepatic histological score | * p < 0.05 | ** p < 0.01 | ns p = 0.357 |
| TGF-β1 mRNA expression | * p < 0.05 | * p < 0.05 | ns p = 0.713 |
| IL-1β mRNA expression | **** p < 0.0001 | ns p = 0.532 | ns p = 0.461 |
| CXCL-1 mRNA expression | *** p < 0.001 | ns p = 0.263 | ns p = 0.368 |
| Hepatic lipid level, lipogenesis related gene expressions | |||
| Hepatic triglyceride | **** p < 0.0001 | * p < 0.05 | ns p = 0.733 |
| Hepatic total cholesterol | **** p < 0.0001 | ns p = 0.168 | ns p = 0.666 |
| FAS mRNA expression | ns p = 0.945 | ns p = 0.466 | ns p = 0.058 |
| G6PD mRNA expression | * p < 0.05 | **** p < 0.0001 | **** p < 0.0001 |
| ACC mRNA expression | *** p < 0.001 | ns p = 0.062 | ns p = 0.341 |
| C/EBP-α mRNA expression | **** p < 0.0001 | ns p = 0.132 | ** p < 0.01 |
| SCD1 mRNA expression | **** p < 0.0001 | ** p < 0.01 | ** p < 0.01 |
| EAT inflammatory and fibrosis related gene expressions | |||
| Adipocyte area | ns p = 0.157 | *** p < 0.001 | ** p < 0.01 |
| Number of CLS | ** p < 0.01 | ns p = 0.294 | ** p < 0.01 |
| IL-1β mRNA expression | **** p < 0.0001 | ns p = 0.485 | ns p = 0.471 |
| CXCL-1 mRNA expression | *** p < 0.001 | ns p = 0.221 | ns p = 0.222 |
| TGF-β1 mRNA expression | **** p < 0.0001 | ** p < 0.01 | ns p = 0.302 |
| Groups | PBS | LPS | |||||
|---|---|---|---|---|---|---|---|
| Fatty Acid (% Total Fatty Acids) | HFD | HFD + KO | HFD + CO | HFD | HFD + KO | HFD + CO | |
| C14:0, Myristic acid | 1.11 ± 0.45 | 0.83 ± 0.16 | 1.53 ± 0.59 | 0.88 ± 0.27 | 0.92 ± 0.19 | 1.24 ± 0.42 | |
| C16:0, Palmitic acid | 28.47 ± 0.55 | 30.5 ± 0.52 | 26.9 ± 0.95 | 28.07 ± 0.49 | 22.43 ± 11.12 | 26.57 ± 0.21 | |
| C16:1, Palmitoleic acid | 1.17 ± 0.4 | 0.74 ± 0.27 | 0.80 ± 0.39 | 0.84 ± 0.04 | 0.85 ± 0.25 | 0.61 ± 0.09 | |
| C18:0, Stearic acid | 15.17 ± 1.1 | 16.57 ± 0.87 | 16.97 ± 0.64 | 15.77 ± 0.68 | 15.83 ± 0.23 | 16.57 ± 0.95 | |
| C18:1, Oleic acid | 18.03 ± 3.40 | 14.67 ± 1.45 | 13.93 ± 2.03 | 17.37 ± 1.05 | 15.9 ± 0.20 | 16.4 ± 0.92 | |
| C18:2, Linoleic acid | 7.70 ± 1.08 | 7.92 ± 0.50 | 7.82 ± 0.63 | 6.66 ± 0.64 | 8.39 ± 0.73 | 8.51 ± 0.43 | |
| C18:3n-3, α-linolenic acid | 0.15 ± 0.11 | 0.06 ± 0.05 | 0.10 ± 0.06 | 0.11 ± 0.12 | 0.09 ± 0.03 | 0.05 ± 0.02 | |
| C18:3n-6, γ-linolenic acid | 0.05 ± 0.02 | 0.03 ± 0.02 | 0.08 ± 0.05 | 0.04 ± 0.02 | 0.04 ± 0.01 | 0.04 ± 0.01 | |
| C20:1, Eicosenoic acid | 0.18 ± 0.01 | 0.10 ± 0.06 | 0.12 ± 0.04 | 0.16 ± 0.12 | 0.09 ± 0.06 | 0.16 ± 0.02 | |
| C20:2, Eicosadienoic acid | 0.32 ± 0.12 | 0.18 ± 0.07 | 0.26 ± 0.08 | 0.20 ± 0.07 | 0.18 ± 0.06 | 0.24 ± 0.07 | |
| C20:3, Dihomo-γ-linolenic acid | 0.65 ± 0.07 a | 0.55 ± 0.08 ab | 0.58 ± 0.11 ab | 0.50 ± 0.05 b | 0.63 ± 0.04 a | 0.55 ± 0.04 ab | |
| C20:4, Aachidonic acid | 20.00 ± 2.25 | 11.00 ± 0.92 | 23.7 ± 1.06 | 21.73 ± 1.19 | 11.00 ± 0.20 | 22.10 ± 0.95 | |
| C20:5, Eicosapentaenoic acid | 0.36 ± 0.12 c | 7.58 ± 0.48a | 0.22 ± 0.06 c | 0.39 ± 0.12 c | 6.64 ± 0.48 b | 0.20 ± 0.03 c | |
| C22:4, Docosatetraenoic acid | 0.89 ± 0.24 | 0.09 ± 0.06 | 1.57 ± 0.37 | 0.89 ± 0.07 | 0.08 ± 0.02 | 1.3 ± 0.11 | |
| C22:5, Docosapentaenoic acid | 1.27 ± 0.29 | 3.47 ± 0.64 | 1.22 ± 0.17 | 1.2 ± 0.46 | 3.67 ± 0.1 | 1.07 ± 0.14 | |
| C22:6n-3, Docosahexaenoic acid | 2.57 ± 0.41 | 3.97 ± 0.74 | 2.11 ± 0.25 | 3.1 ± 0.42 | 4.89 ± 0.51 | 2.31 ± 0.21 | |
| C22:6n-6, Docosapentaenoic acid | 0.24 ± 0.07 | 0.02 ± 0.01 | 0.66 ± 0.18 | 0.35 ± 0.06 | 0.06 ± 0.01 | 0.57 ± 0.17 | |
| C24:0, Lignoceric acid | 0.32 ± 0.09 | 0.36 ± 0.23 | 0.37 ± 0.04 | 0.26 ± 0.04 | 0.35 ± 0.05 | 0.33 ± 0.05 | |
| C24:1, Nervonic acid | 0.21 ± 0.05 | 0.29 ± 0.09 | 0.11 ± 0.03 | 0.12 ± 0.11 | 0.23 ± 0.15 | 0.13 ± 0.03 | |
| Others 1 | 0.28 ± 0.02 | 0.20 ± 0.05 | 0.21 ± 0.04 | 0.23 ± 0.05 | 0.16 ± 0.02 | 0.16 ± 0.04 | |
| SFAs | 45.07 ± 1.27 | 48.26 ± 1.57 | 45.76 ± 1.1 | 44.97 ± 0.68 | 39.54 ± 11.38 | 44.7 ± 1.29 | |
| MUFAs | 19.6 ± 3.76 | 15.79 ± 1.67 | 14.96 ± 2.37 | 18.48 ± 1.08 | 17.07 ± 0.53 | 17.3 ± 1.01 | |
| PUFAs | 34.2 ± 2.79 | 34.86 ± 2.29 | 38.31 ± 1.63 | 35.17 ± 1.26 | 35.68 ± 0.84 | 36.92 ± 1.15 | |
| n-6 | 29.85 ± 2.42 | 19.79 ± 0.63 | 34.66 ± 1.69 | 30.37 ± 0.51 | 20.38 ± 0.68 | 33.3 ± 1.23 | |
| n-3 | 4.35 ± 0.45 | 15.07 ± 1.67 | 3.65 ± 0.15 | 4.8 ± 0.75 | 15.29 ± 0.94 | 3.62 ± 0.08 | |
| n-6/n-3 | 6.88 ± 0.48 | 1.32 ± 0.11 | 9.51 ± 0.72 | 6.43 ± 0.98 | 1.34 ± 0.11 | 9.21 ± 0.53 | |
| AA/EPA | 58.24 ± 12.03 | 1.45 ± 0.06 | 112.14 ± 27.47 | 59.25 ± 17.89 | 1.66 ± 0.15 | 114.63 ± 21.89 | |
| Strain | Inducer | Treatment | Biological Markers | Ref. |
|---|---|---|---|---|
| C57BL/6J mouse | HFD | 5 % (w/w) for 12 weeks | Liver and epididymal fat weights ↓ | [33] |
| Serum TC, LDL-C levels ↓ | ||||
| Serum and liver MDA level, ALT activity ↓ | ||||
| Serum and liver SOD levels ↑ | ||||
| C57BL/6N mouse | HFD | 2% (w/w) for 12 weeks | Body weight and abdominal fat (visceral and subcutaneous) contents ↓ | [22] |
| Hepatic weight and hepatic steatosis ↓ | ||||
| Serum TG, LDL-C levels ↓ | ||||
| Fatty acid synthesis related genes expression ↓ | ||||
| Sprague-Dawley rats | HFD | 2.5% (w/w) for 12 weeks | Body weight gain ↓ | [28] |
| Hepatic fatty acid oxidation related gene expression ↑ | ||||
| Plasma TG levels ↓ | ||||
| Hepatic TG, TC levels ↓ | ||||
| Glucose and insulin concentration ↓ | ||||
| C57BL/6 mouse | HFD | 1.25, 2.5, and 5% (w/w) for 8 weeks | Liver weight ↓ | [34] |
| Hepatic TG, TC levels ↓ | ||||
| Serum TC, glucose ↓ | ||||
| Serum adiponectin ↑ | ||||
| Hepatic TNF-α level and mRNA expression ↓ | ||||
| Hepatic FA synthesis associated with gene expression (FAS, ACC, SCD-1) ↓ | ||||
| Hepatic FA catabolism associated with gene expression (MGLL, LIPE) ↓ | ||||
| Hepatic FA oxidation associated with gene expression (CPT-1) ↓ | ||||
| Hepatic cholesterol metabolism associated with gene expression (HMG CoA-R, LDL-R) ↓ | ||||
| Hepatic transcription factors gene expression (SREBP-1c, SREBP-2, PPAR-α) ↓ |
| Strain | Inducer | Treatment | Biological Markers | Ref. |
|---|---|---|---|---|
| Wistar rats | Paracetamol, 750 mg/kg body weight (i.p.) | Virgin coconut oil (phoscoliv) 0.5 mL/100 g body weight | Serum proinflammatory cytokines (IL-6, TNF-α) ↓ | [74] |
| Histological scoring of liver tissue ↓ | ||||
| Wistar rats | HFD (60% of calories from fat) | high-coconut oil diet (60% of calories from coconut oil) for 3 days | Serum glucose, insulin, leptin concentrations ↓ | [75] |
| Protein levels of inflammatory markers in the hippocampi and prefrontal cortices (IL-1β, IL-6, TNF-α and NF-κB) ↓ | ||||
| Sprague-Dawley rats | LPS 10 mg/kg body weight (i.p.) | Lauric acid 50, 100 mg/kg body weight (p.o.) for 14 days | Serum AST, ALT levels ↓ | [76] |
| Serum TC, non-HDL-C, LDL-C and TG ↓ | ||||
| Serum HDL-C ↑ | ||||
| Serum proinflammatory cytokines (TNF-α, IL-6, IL-1β) ↓ | ||||
| Hepatic proinflammatory cytokines (TNF-α, IL-6, IL-1β) ↓ | ||||
| Hepatic protein expression of TLR4, MyD88, NF-κB, TNF-α, IL-1β ↓ | ||||
| C57BL/6 mouse | HFD | 1.25, 2.5, and 5% (w/w) for 8 weeks | Liver weight ↓ | [77] |
| Hepatic TG, TC levels ↓ | ||||
| Serum TC, glucose ↓ | ||||
| Serum adiponectin ↑ | ||||
| Hepatic TNF-α level, mRNA expression ↓ | ||||
| Hepatic FA synthesis associated with gene expression (FAS, ACC, SCD-1) ↓ | ||||
| Wistar rats | Gentamicin, 100 mg/kg body weight (i.p.) | Virgin coconut oil 10% (w/w) for 16 days | Renal oxidative stress index (SOD, CAT, GPx, GSH, MDA) ↓ | [78] |
| Renal inflammatory cytokines (IL-1β, IL-6, TNF-α, NO, iNOS) ↓ | ||||
| Renal NF-κB level ↓ | ||||
| Renal caspase-3 activity ↓ | ||||
| Renal histomorphology ↓ | ||||
| Wistar rats | Virgin coconut oil 4, 8, 16% (w/w) for 30 days | Proinflammatory cytokine (TNF-α) production in the splenocytes ↓ | [73] | |
| p-mTOR, SIRT1, and p-LKB1 expression in the spleen ↑ | ||||
| p-ERK1/2/Total ERK1/2, p-CREB/Total CREB expression in the spleen ↑ | ||||
| Splenic SOD and CAT activities ↑ | ||||
| Serum HDL-C level ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, H.-K.; Kim, B.-H.; Lee, J.; Park, S.; Oh, C.-B.; Jung, S.; Lee, J.K.; Ha, J.-H. Partial Replacement of Dietary Fat with Krill Oil or Coconut Oil Alleviates Dyslipidemia by Partly Modulating Lipid Metabolism in Lipopolysaccharide-Injected Rats on a High-Fat Diet. Int. J. Environ. Res. Public Health 2022, 19, 843. https://doi.org/10.3390/ijerph19020843
Son H-K, Kim B-H, Lee J, Park S, Oh C-B, Jung S, Lee JK, Ha J-H. Partial Replacement of Dietary Fat with Krill Oil or Coconut Oil Alleviates Dyslipidemia by Partly Modulating Lipid Metabolism in Lipopolysaccharide-Injected Rats on a High-Fat Diet. International Journal of Environmental Research and Public Health. 2022; 19(2):843. https://doi.org/10.3390/ijerph19020843
Chicago/Turabian StyleSon, Hee-Kyoung, Bok-Hee Kim, Jisu Lee, Seohyun Park, Chung-Bae Oh, Sunyoon Jung, Jennifer K. Lee, and Jung-Heun Ha. 2022. "Partial Replacement of Dietary Fat with Krill Oil or Coconut Oil Alleviates Dyslipidemia by Partly Modulating Lipid Metabolism in Lipopolysaccharide-Injected Rats on a High-Fat Diet" International Journal of Environmental Research and Public Health 19, no. 2: 843. https://doi.org/10.3390/ijerph19020843