
Effects of Gadolinium Deposits in the Cerebellum: Reviewing the Literature from In Vitro Laboratory Studies to In Vivo Human Investigations


Abstract
:1. Introduction
2. Gadolinium-Based Contrast Agent (GBCA) Deposition in the Cerebellum
3. Effects of GBCAs on Cerebellar Structure and Function
3.1. Human Studies
3.2. Animal Studies
4. Possible Mechanisms of GBCA Toxicity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MRI | Magnetic resonance imaging |
| GBCA | Gadolinium-based contrast agent |
| T1WI | T1-weighted images |
| FDA | Food and drug administration |
| DN | Dentate nucleus |
| DCN | Deep cerebellar nucleus |
| GP | Globus pallidus |
| ICP-MS | Inductively-coupled plasma mass spectrometry |
| Gd | Gadolinium |
| Fe | Ferrum |
| Zn | Zinc |
| Cu | Cuprum |
| Gd-DTPA | Gadolinium-diethylenetriaminepentacetate |
| Gd-DTPA-BMA | Gadolinium-diethylenetriaminepentacetate-bis-(methylamide) |
| Gd-DOTA | Gadolinium-tetra-azacyclo-dodecane-tetra-acetic acid |
| ORT | Object recognition test |
| OLT | Object-in-location test |
| Ca | Calcium |
| DIV | Days in vitro |
| TH | Thyroid hormone |
| TR | Thyroid hormone receptor |
| TRE | Thyroid hormone response element |
| CaMKIV | Ca2+/calmodulin-dependent protein kinase type IV |
References
- Caravan, P.; Ellison, J.J.; McMurry, T.J.; Lauffer, R.B. Gadolinium(III) chelates as MRI contrast agents: Structure, dynamics, and applications. Chem. Rev. 1999, 99, 2293–2352. [Google Scholar] [CrossRef] [PubMed]
- Akakuru, O.U.; Iqbal, M.Z.; Saeed, M.; Liu, C.; Paunesku, T.; Woloschak, G.; Hosmane, N.S.; Wu, A. The Transition from Metal-Based to Metal-Free Contrast Agents for T1 Magnetic Resonance Imaging Enhancement. Bioconjug. Chem. 2019, 30, 2264–2286. [Google Scholar] [CrossRef]
- Idée, J.M.; Port, M.; Raynal, I.; Schaefer, M.; Le Greneur, S.; Corot, C. Clinical and biological consequences of transmetallation induced by contrast agents for magnetic resonance imaging: A review. Fundam. Clin. Pharmacol. 2006, 20, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Rogosnitzky, M.; Branch, S. Gadolinium-based contrast agent toxicity: A review of known and proposed mechanisms. BioMetals 2016, 29, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baykara, M.; Ozcan, M.; Bilgen, M.; Kelestimur, H. Effects of gadolinium and gadolinium chelates on intracellular calcium signaling in sensory neurons. Neurosci. Lett. 2019, 707, 134295. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, L.; Napolitano, A.; Visconti, E.; Longo, D.; Romano, A.; Tomà, P.; Espagnet, M.C.R. Gadolinium-Based Contrast Agent-Related Toxicities. CNS Drugs 2018, 32, 229–240. [Google Scholar] [CrossRef]
- Frenzel, T.; Apte, C.; Jost, G.; Schöckel, L.; Lohrke, J.; Pietsch, H. Quantification and Assessment of the Chemical Form of Residual Gadolinium in the Brain After Repeated Administration of Gadolinium-Based Contrast Agents. Investig. Radiol. 2017, 52, 396–404. [Google Scholar] [CrossRef] [Green Version]
- Kanda, T.; Osawa, M.; Oba, H.; Toyoda, K.; Kotoku, J.; Haruyama, T.; Takeshita, K.; Furui, S. High signal intensity in dentate nucleus on unenhanced T1-weighted MR images: Association with linear versus macrocyclic gadolinium chelate administration. Radiology 2015, 275, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Kanda, T.; Fukusato, T.; Matsuda, M.; Toyoda, K.; Oba, H.; Haruyama, T.; Kitajima, K.; Furui, S. Gadolinium-based contrast agent accumulates in the brain even in subjects without severe renal dysfunction: Evaluation of autopsy brain specimens with inductively coupled plasma mass spectroscopy. Neuroradiology 2015, 276, 228–232. [Google Scholar] [CrossRef]
- Robert, P.; Violas, X.; Grand, S.; Lehericy, S.; Idée, J.-M.; Ballet, S.; Corot, C. Linear Gadolinium-Based Contrast Agents Are Associated With Brain Gadolinium Retention in Healthy Rats. Investig. Radiol. 2016, 51, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Robert, P.; Fingerhut, S.; Factor, C.; Vives, V.; Letien, J.; Sperling, M.; Rasschaert, M.; Santus, R.; Ballet, S.; Idée, J.M.; et al. One-year retention of gadolinium in the brain: Comparison of gadodiamide and gadoterate meglumine in a rodent model. Radiology 2018, 288, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Jost, G.; Frenzel, T.; Boyken, J.; Lohrke, J.; Nischwitz, V.; Pietsch, H. Long-term Excretion of Gadolinium-based Contrast Agents: Linear versus Macrocyclic Agents in an Experimental Rat Model. Radiology 2019, 290, 340–348. [Google Scholar] [CrossRef] [PubMed]
- McDonald, R.J.; McDonald, J.S.; Dai, D.; Schroeder, D.; Jentoft, M.E.; Murray, D.L.; Kadirvel, R.; Eckel, L.J.; Kallmes, D.F. Comparison of Gadolinium Concentrations within Multiple Rat Organs after Intravenous Administration of Linear versus Macrocyclic Gadolinium Chelates. Radiology 2017, 285, 536–545. [Google Scholar] [CrossRef] [Green Version]
- Mcdonald, R.J.; Mcdonald, J.S.; Kallmes, D.F.; Jentoft, M.E.; Paolini, M.A.; Murray, D.L.; Williamson, E.E.; Eckel, L.J. Gadolinium Deposition in Human Brain Tissues after Contrast-enhanced MR Imaging in Adult Patients without Intracranial Abnormalities. Radiology 2017, 285, 546–554. [Google Scholar] [CrossRef]
- Rasschaert, M.; Idée, J.; Robert, P.; Fretellier, N.; Vives, V.; Violas, X.; Ballet, S.; Corot, C. Moderate Renal Failure Accentuates T1 Signal Enhancement in the Deep Cerebellar Nuclei of Gadodiamide-Treated Rats. Investig. Radiol. 2017, 52, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasschaert, M.; Schroeder, J.A.; Wu, T.D.; Marco, S.; Emerit, A.; Siegmund, H.; Fischer, C.; Fretellier, N.; Idée, J.M.; Corot, C.; et al. Multimodal Imaging Study of Gadolinium Presence in Rat Cerebellum: Differences between Gd Chelates, Presence in the Virchow-Robin Space, Association with Lipofuscin, and Hypotheses about Distribution Pathway. Investig. Radiol. 2018, 53, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Lohrke, J.; Frisk, A.-L.; Frenzel, T.; Schöckel, L.; Rosenbruch, M.; Jost, G.; Lenhard, D.C.; Sieber, M.A.; Nischwitz, V.; Küppers, A.; et al. Histology and Gadolinium Distribution in the Rodent Brain After the Administration of Cumulative High Doses of Linear and Macrocyclic Gadolinium-Based Contrast Agents. Investig. Radiol. 2017, 52, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Hamrani, D.E.; Vives, V.; Buchholz, R.; Même, W.; Factor, C.; Fingerhut, S.; Sperling, M.; Karst, U.; Robert, P.; Même, S. Effect of Long-Term Retention of Gadolinium on Metabolism of Deep Cerebellar Nuclei after Repeated Injections of Gadodiamide in Rats. Investig. Radiol. 2020, 55, 120–128. [Google Scholar] [CrossRef]
- Gianolio, E.; Bardini, P.; Arena, F.; Stefania, R.; Di Gregorio, E.; Iani, R.; Aime, S. Gadolinium Retention in the Rat Brain: Assessment of the Amounts of Insoluble Gadolinium-containing Species and Intact Gadolinium Complexes after Repeated Administration of Gadolinium-based Contrast Agents. Radiology 2017, 285, 839–849. [Google Scholar] [CrossRef]
- Khairinisa, M.A.; Takatsuru, Y.; Amano, I.; Erdene, K.; Nakajima, T.; Kameo, S.; Koyama, H.; Tsushima, Y.; Koibuchi, N. The Effect of Perinatal Gadolinium-Based Contrast Agents on Adult Mice Behavior. Investig. Radiol. 2018, 53, 110–118. [Google Scholar] [CrossRef]
- D’Angelo, E. Physiology of the Cerebellum, 1st ed.; Elsevier, B.V.: Amsterdam, The Netherlands, 2018; Volume 154. [Google Scholar]
- Bohne, P.; Schwarz, M.K.; Herlitze, S.; Mark, M.D. A New Projection from the Deep Cerebellar Nuclei to the Hippocampus via the Ventrolateral and Laterodorsal Thalamus in Mice. Front. Neural Circuits 2019, 13, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habas, C.; Van Dun, K.; Manto, M.; Mariën, P. Deep Cerebellar Nuclei (DCN) and Language; Elsevier Inc.: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Kanda, T.; Oba, H.; Toyoda, K.; Kitajima, K. Brain gadolinium deposition after administration of gadolinium-based contrast agents. Jpn. J. Radiol 2016, 34, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strzeminska, I.; Factor, C.; Robert, P.; Grindel, A.L.; Comby, P.O.; Szpunar, J.; Corot, C.; Lobinski, R. Long-Term Evaluation of Gadolinium Retention in Rat Brain after Single Injection of a Clinically Relevant Dose of Gadolinium-Based Contrast Agents. Investig. Radiol. 2020, 55, 138–143. [Google Scholar] [CrossRef]
- Roberts, D.R.; Holden, K.R. Progressive increase of T1 signal intensity in the dentate nucleus and globus pallidus on unenhanced T1-weighted MR images in the pediatric brain exposed to multiple doses of gadolinium contrast. Brain Dev. 2016, 38, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.R.; Chatterjee, A.R.; Yazdani, M.; Marebwa, B.; Brown, T.; Collins, H.; Bolles, G.; Jenrette, J.M.; Nietert, P.J.; Zhu, X. Pediatric patients demonstrate progressive t1-weighted hyperintensity in the dentate nucleus following multiple doses of gadolinium-based contrast agent. AJNR Am. J. Neuroradiol. 2016, 37, 2340–2347. [Google Scholar] [CrossRef] [Green Version]
- Clough, T.J.; Jiang, L.; Wong, K.L.; Long, N.J. Ligand design strategies to increase stability of gadolinium-based magnetic resonance imaging contrast agents. Nat. Commun. 2019, 10, 1420. [Google Scholar] [CrossRef] [Green Version]
- Sherry, A.D.; Caravan, P.; Lenkinski, R.E. Primer on gadolinium chemistry. J. Magn. Reson. Imaging 2009, 30, 1240–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, Z.; Thurmond, A.S.; Ross, P.L.; Jones, M.K.; Thornburg, K.L.; Katzberg, R.W. Gadolinium-DTPA transplacental transfer and distribution in fetal tissue in rabbits. Investig. Radiol. 1993, 28, 828–830. [Google Scholar] [CrossRef]
- Vahter, M.; Akesson, A.; Lidén, C.; Ceccatelli, S.; Berglund, M. Gender differences in the disposition and toxicity of metals. Environ. Res. 2007, 104, 85–95. [Google Scholar] [CrossRef]
- Semelka, R.C.; Ramalho, J.; Vakharia, A.; AlObaidy, M.; Burke, L.M.; Jay, M.; Ramalho, M. Gadolinium deposition disease: Initial description of a disease that has been around for a while. Magn. Reson. Imaging 2016, 34, 1383–1390. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.; Grimm, H. Gadolinium Toxicity A Survey of the Chronic Effects of Retained Gadolinium from Contrast MRIs. 2014. Available online: www.gadoliniumToxicity.com (accessed on 10 May 2021).
- Welk, B.; McArthur, E.; Morrow, S.A.; MacDonald, P.; Hayward, J.; Leung, A.; Lum, A. Association between gadolinium contrast exposure and the risk of parkinsonism. JAMA J. Am. Med. Assoc. 2016, 316, 96–98. [Google Scholar] [CrossRef] [Green Version]
- Perrotta, G.; Metens, T.; Absil, J.; Lemort, M.; Manto, M. Absence of clinical cerebellar syndrome after serial injections of more than 20 doses of gadoterate, a macrocyclic GBCA: A monocenter retrospective study. J. Neurol. 2017, 264, 2277–2283. [Google Scholar] [CrossRef]
- Bird, S.T.; Gelperin, K.; Sahin, L.; Bleich, K.B.; Fazio-Eynullayeva, E.; Woods, C.; Radden, E.; Greene, P.; McCloskey, C.; Johnson, T.; et al. First-trimester exposure to gadolinium-based contrast agents: A utilization study of 4.6 Million, U.S. Pregnancies. Radiology 2019, 293, 193–200. [Google Scholar] [CrossRef]
- Ray, D.E.; Holton, J.L.; Nolan, C.C.; Cavanagh, J.B.; Harpur, E.S. Neurotoxic potential of gadodiamide after injection into the lateral cerebral ventricle of rats. AJNR Am. J. Neuroradiol. 1998, 19, 1455–1462. [Google Scholar]
- Ray, D.E.; Cavanagh, J.B.; Nolan, C.C.; Williams, S.C.R. Neurotoxic effects of gadopentetate dimeglumine: Behavioral disturbance and morphology after intracerebroventricular injection in rats. AJNR Am. J. Neuroradiol. 1996, 17, 365–373. [Google Scholar]
- Toney, G.M.; Chavez, H.A.; Ibarra, R.; Jinkins, J.R. Acute and subacute physiological and histological studies of the central nervous system after intrathecal gadolinium injection in the anesthetized rat. Investig. Radiol. 2001, 36, 33–40. [Google Scholar] [CrossRef]
- Adams, M.; Barco, S.; Gale, K.; Hynes, M.; Ralston, W. Motor Coordination Testing of the Neurotoxicity of Paramagnetic Chelates. In New Developments in Contrast Agents: Proceedings of the 3rd Special Topic Seminar of the European Magnetic Resonance Forum; European Magnetic Resonance Forum Foundation: Hamburg, Germany, 1993; pp. 51–58. [Google Scholar]
- Koibuchi, N. Animal models to study thyroid hormone action in cerebellum. Cerebellum 2009, 8, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.A.; Miall, R.C.; Ivry, R.B. The Cerebellum: Adaptive Prediction for Movement and Cognition. Trends Cogn. Sci. 2017, 21, 313–332. [Google Scholar] [CrossRef] [Green Version]
- Robert, P.; Lehericy, S.; Grand, S.; Violas, X.; Fretellier, N.; Ideé, J.M.; Ballet, S.; Corot, C. T1-Weighted Hypersignal in the Deep Cerebellar Nuclei after Repeated Administrations of Gadolinium-Based Contrast Agents in Healthy Rats: Difference between Linear and Macrocyclic Agents. Investig. Radiol. 2015, 50, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Bussi, S.; Coppo, A.; Botteron, C.; Fraimbault, V.; Fanizzi, A.; De Laurentiis, E.; Colombo Serra, S.; Kirchin, M.A.; Tedoldi, F.; Maisano, F. Differences in gadolinium retention after repeated injections of macrocyclic MR contrast agents to rats. J. Magn. Reson. Imaging 2018, 47, 746–752. [Google Scholar] [CrossRef]
- Aggleton, J.P.; Albasser, M.M.; Aggleton, D.J.; Poirier, G.L.; Pearce, J.M. Lesions of the rat perirhinal cortex spare the acquisition of a complex configural visual discrimination yet impair object recognition. Behav. Neurosci. 2010, 124, 55–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckmaster, C.; Eichenbaum, H.; Amaral, D.; Suzuki, W.A.; Rapp, P.R. Entorhinal cortex lesions disrupt the relational organization of memory in monkeys. J. Neurosci. 2004, 24, 9811–9825. [Google Scholar] [CrossRef] [Green Version]
- Ariyani, W.; Iwasaki, T.; Miyazaki, W.; Khongorzul, E.; Nakajima, T.; Kameo, S.; Koyama, H.; Tsushima, Y.; Koibuchi, N. Effects of gadolinium-based contrast agents on thyroid hormone receptor action and thyroid hormone-induced cerebellar purkinje cell morphogenesis. Front. Endocrinol. 2016, 7, 115. [Google Scholar] [CrossRef] [Green Version]
- Toczylowska, B.; Zieminska, E.; Goch, G.; Milej, D.; Gerega, A.; Liebert, A. Neurotoxic effects of indocyanine green -cerebellar granule cell culture viability study. Biomed. Opt. Express 2014, 5, 800. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.D.; Xia, Q.; Yuan, L.; Huang, H.F.; Yang, X.D.; Wang, K. Gadolinium triggers unfolded protein responses (UPRs) in primary cultured rat cortical astrocytes via promotion of an influx of extracellular Ca2+. Cell Biol. Toxicol. 2011, 27, 1–12. [Google Scholar] [CrossRef]
- Bellin, M.F.; Van Der Molen, A.J. Extracellular gadolinium-based contrast media: An overview. Eur. J. Radiol. 2008, 66, 160–167. [Google Scholar] [CrossRef]
- Baykara, M.; Ozcan, M.; Bilgen, M.; Kelestimur, H. Interference of gadolinium dechelated from MR contrast agents by calcium signaling in neuronal cells of GnRH. J. Cell Physiol. 2021, 236, 2139–2143. [Google Scholar] [CrossRef]
- Bower, D.V.; Richter, J.K.; Von Tengg-Kobligk, H.; Heverhagen, J.T.; Runge, V.M. Gadolinium-Based MRI Contrast Agents Induce Mitochondrial Toxicity and Cell Death in Human Neurons, and Toxicity Increases With Reduced Kinetic Stability of the Agent. Investig. Radiol. 2019, 54, 453–463. [Google Scholar] [CrossRef]
- Koibuchi, N. The role of thyroid hormone on functional organization in the cerebellum. Cerebellum 2013, 12, 304–306. [Google Scholar] [CrossRef]
- Kuno-Murata, M.; Koibuchi, N.; Fukuda, H.; Murata, M.; Chin, W.W. Augmentation of thyroid hormone receptor-mediated transcription by Ca2+/calmodulin-dependent protein kinase IV. Endocrinology 2001, 142, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Goglia, F.; Leonard, J.L. Nongenomic actions of thyroid hormone. Nat. Rev. Endocrinol. 2016, 12, 111–121. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Investigators | Model of Study | Type of GBCAs | Doses | Durations | Findings |
|---|---|---|---|---|---|
| Toczylowska et al., 2014 [48] | In Vitro: Primary culture of cerebellar granule cells | Gadobutrol | 0.1, 1 and 10 mM | 30–60 min exposure at DIV7 | Reduction of the toxic effects of indocyanine green by increasing the cell viability of cerebellar granule cells culture 24 h after the exposure. |
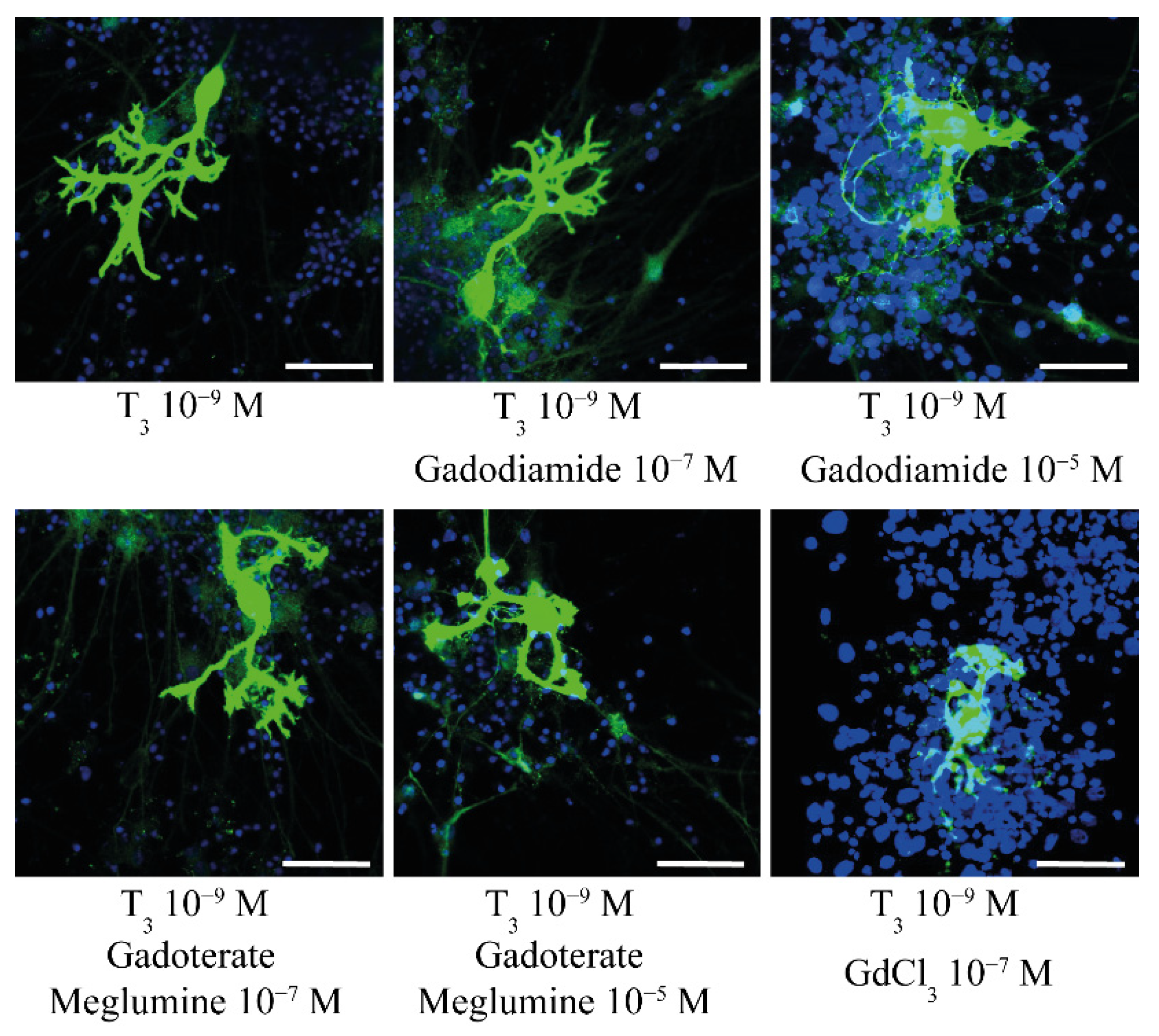
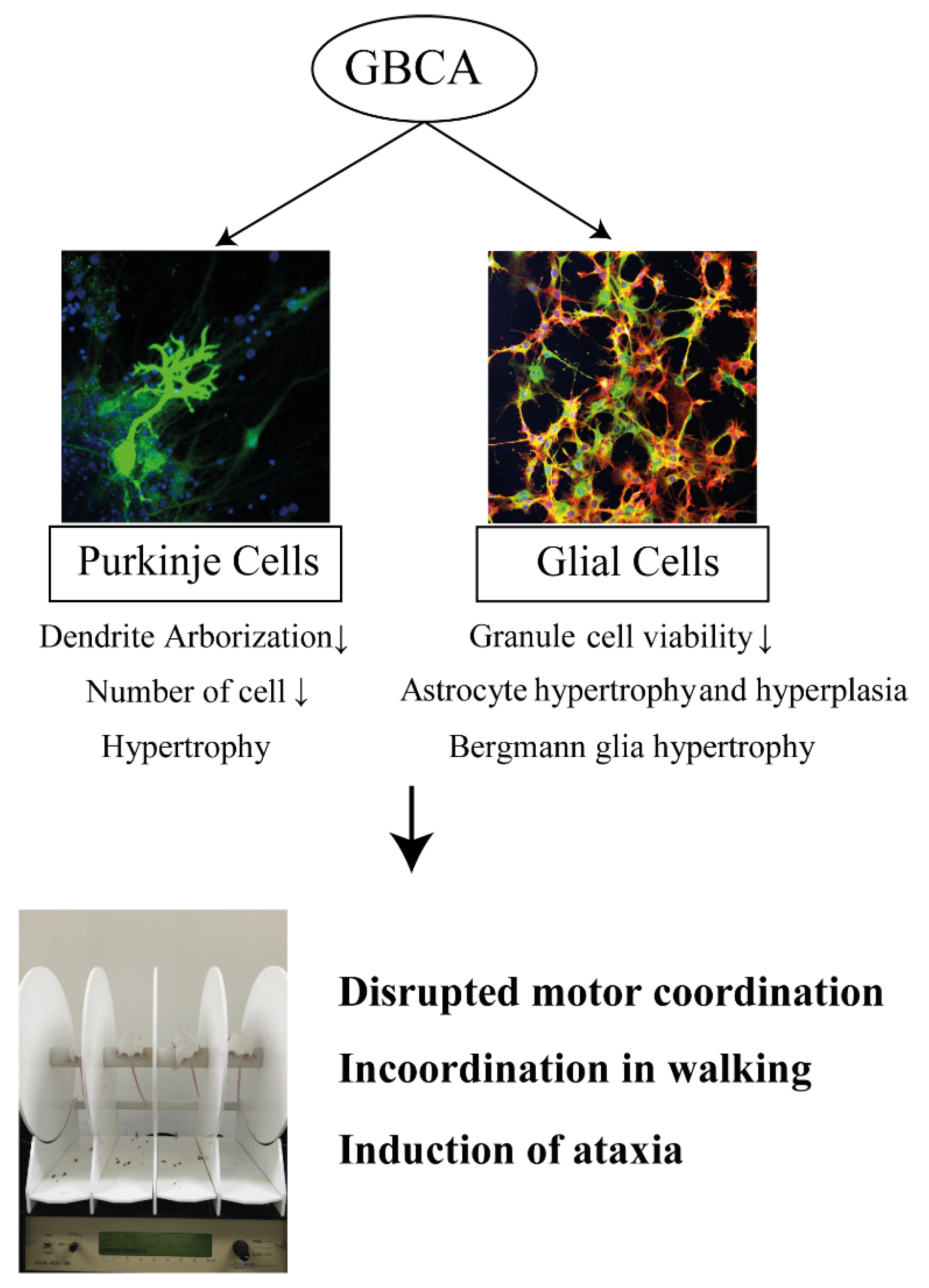
| Ariyani et al., 2016 [47] | In Vitro: Primary culture of cerebellar Purkinje cells | GdCl3; gadodiamide; gadoterate meglumine | 0.1 and 10 µM | DIV1–DIV17 | Alteration of dendrite arborization and cell number of Purkinje cells by gadodiamide and GdCl3, but not gadoterate meglumine. |
| Ray et al., 1996 [38] | In Vivo: Rat | Gadopentetate dimeglumine | 10 µmol/g brain (brain weight estimates at 2 g) | Single intracerebroventricular injection | Morphology: Focal lesions within the thalamus, brain stem, and spinal cord, with necrosis of glia, loss of myelin, and sparing of neurons and nerve fibers, but no description of cerebellar morphology. Behavior: Induction of ataxia with various grades starting at 28–220 min after injection, and lasting for up to 8 days. |
| Ray et al., 1998 [37] | In Vivo: Rat | gadodiamide; gadopentetate dimeglumine | 10 µmol/g brain (brain weight estimates at 2 g) | Single intracerebroventricular injection | Morphology: Astrocyte hypertrophy and hyperplasia by gadodiamide, starting from 3 days after the injection. Hypertrophy of Bergmann glia and Purkinje cell Behavior: Incoordination in walking for several hours after gadodiamide injection. Development of ataxia and a high-stepping gait by a higher dose of gadodiamide later than 4 days after injection. Development of ataxia by gadopentetate dimeglumine at 24 h after injection; The symptom being more severe with transient anesthesia. |
| Khairinisa et al., 2017 [20] | In Vivo: Mouse | gadodiamide; gadoterate meglumine | 2 mmol/kg body weight per day | day 15 to 19 of pregnancy | Induced of the disrupted motor coordination and impaired memory function by gadodiamide and gadoterate meglumine in adulthood. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khairinisa, M.A.; Ariyani, W.; Tsushima, Y.; Koibuchi, N. Effects of Gadolinium Deposits in the Cerebellum: Reviewing the Literature from In Vitro Laboratory Studies to In Vivo Human Investigations. Int. J. Environ. Res. Public Health 2021, 18, 7214. https://doi.org/10.3390/ijerph18147214
Khairinisa MA, Ariyani W, Tsushima Y, Koibuchi N. Effects of Gadolinium Deposits in the Cerebellum: Reviewing the Literature from In Vitro Laboratory Studies to In Vivo Human Investigations. International Journal of Environmental Research and Public Health. 2021; 18(14):7214. https://doi.org/10.3390/ijerph18147214
Chicago/Turabian StyleKhairinisa, Miski Aghnia, Winda Ariyani, Yoshito Tsushima, and Noriyuki Koibuchi. 2021. "Effects of Gadolinium Deposits in the Cerebellum: Reviewing the Literature from In Vitro Laboratory Studies to In Vivo Human Investigations" International Journal of Environmental Research and Public Health 18, no. 14: 7214. https://doi.org/10.3390/ijerph18147214