
Effect of Soybean and Soybean Koji on Obesity and Dyslipidemia in Rats Fed a High-Fat Diet: A Comparative Study



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Steamed Soybean (SS) and Soybean Koji (SK)
2.2. Proximate Composition of SS and SK
2.3. Free Amino Acid Content of SS and SK
2.4. Fatty Acid Content of SS and SK
2.5. Animal Experiments and Diets
2.6. Biochemical and Enzymatic Analysis of Serum Samples
2.7. Tissue and Fecal Lipid Contents
2.8. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.9. Histological Analysis
2.10. Statistical Analysis
3. Results
3.1. Proximate, Free Amino Acid, and Fatty Acid Compositions in Steamed Soybean and Soybean Koji
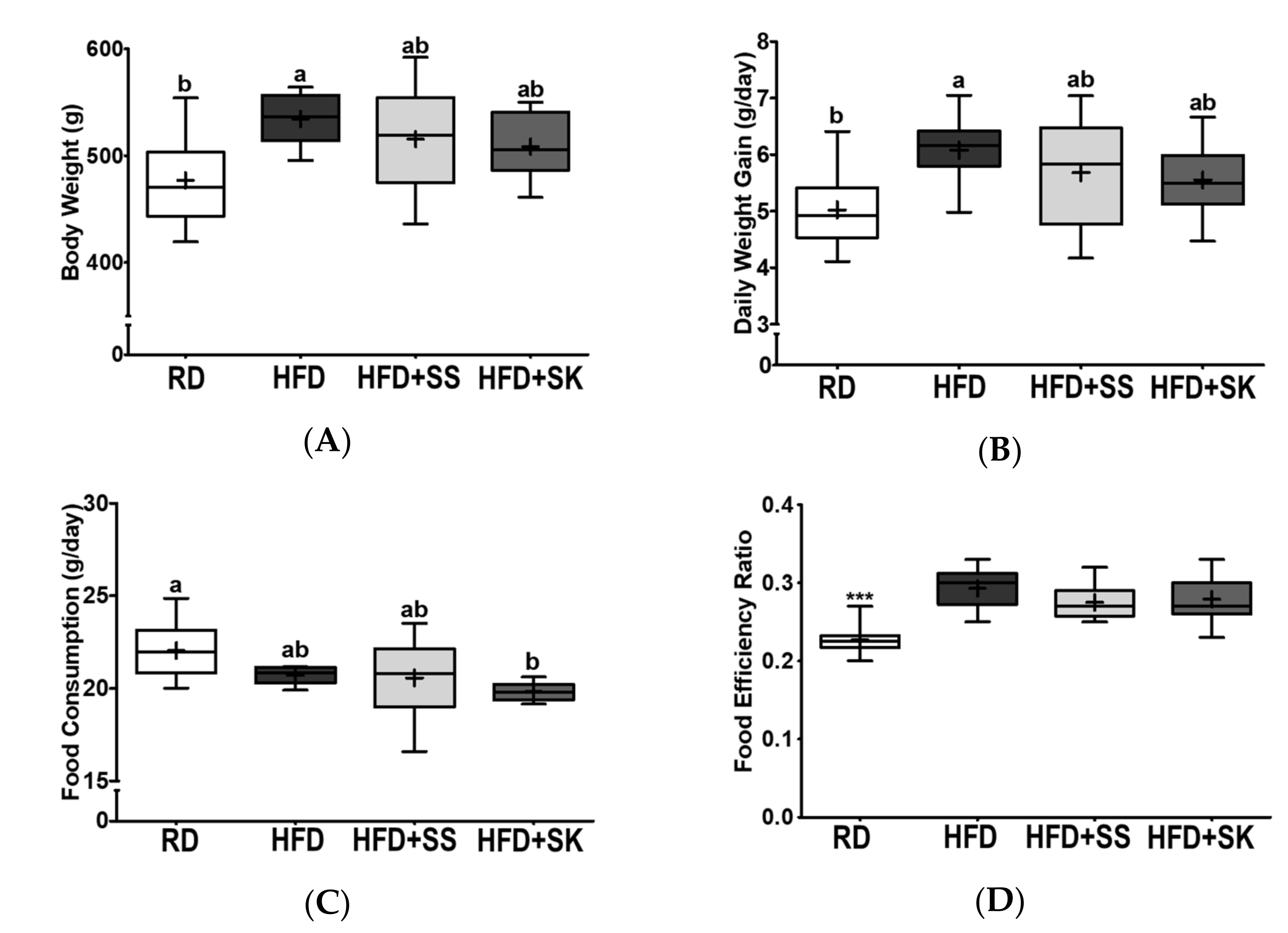
3.2. SS and SK Consumption Tended to Ameliorate HFD-Induced Body Weight Gain
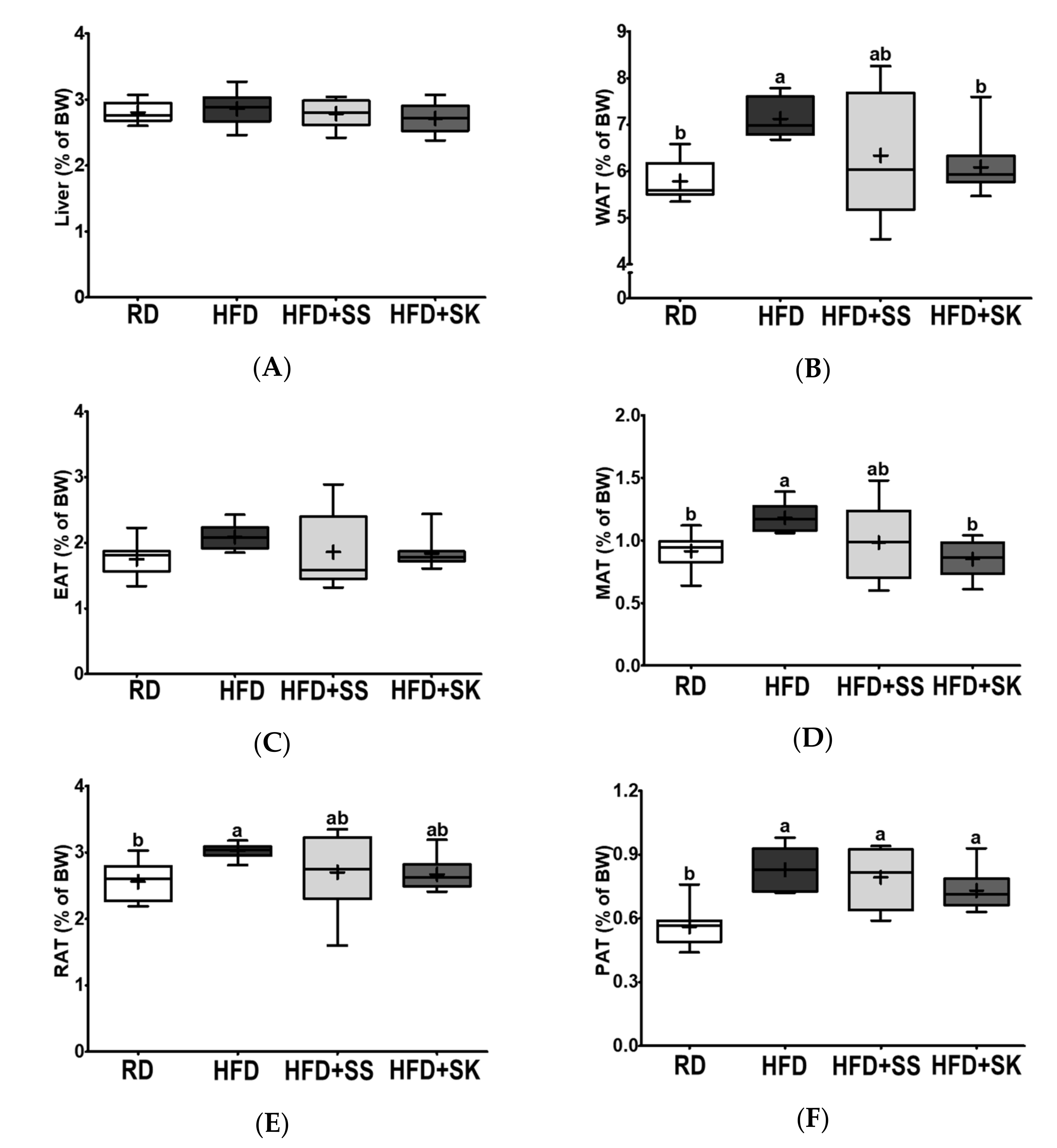
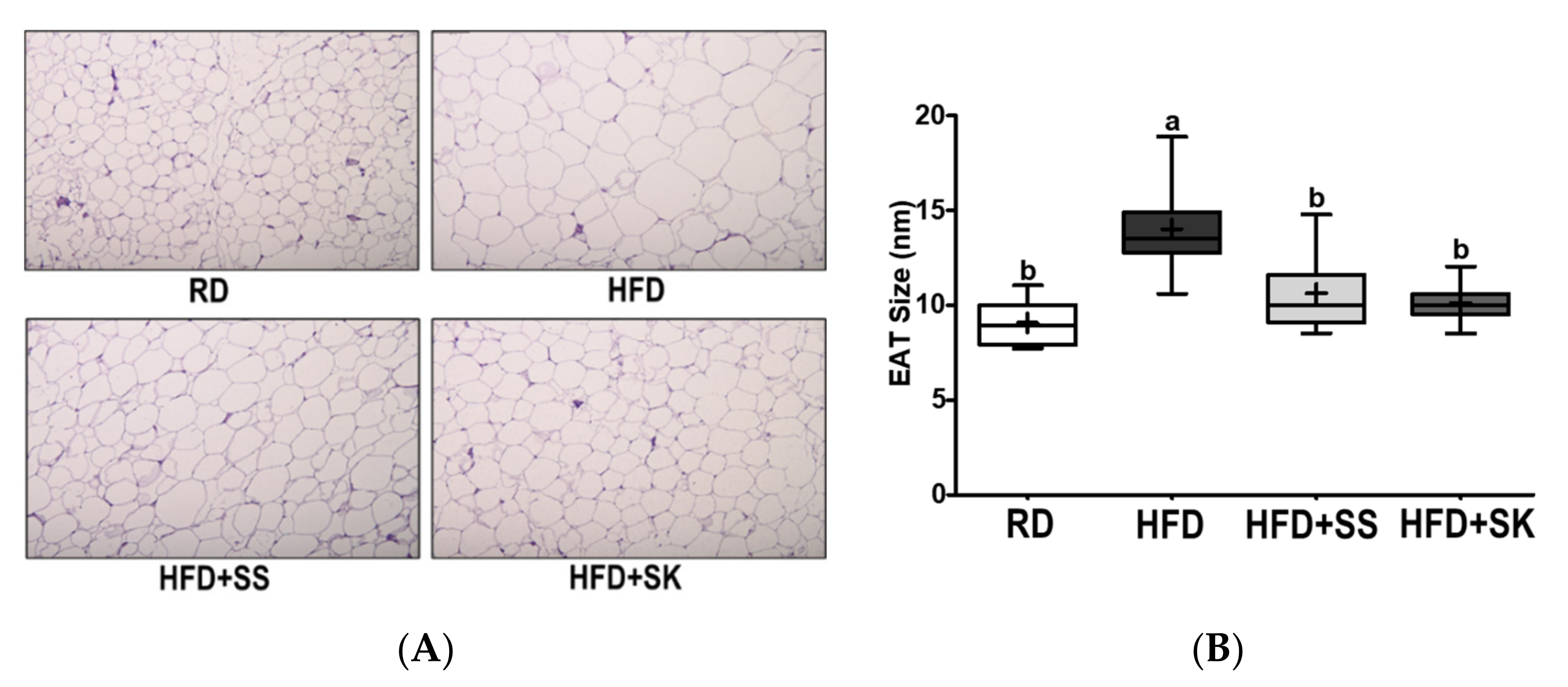
3.3. SK Consumption Ameliorated HFD-Induced Fat Accumulation in White Adipose Tissue
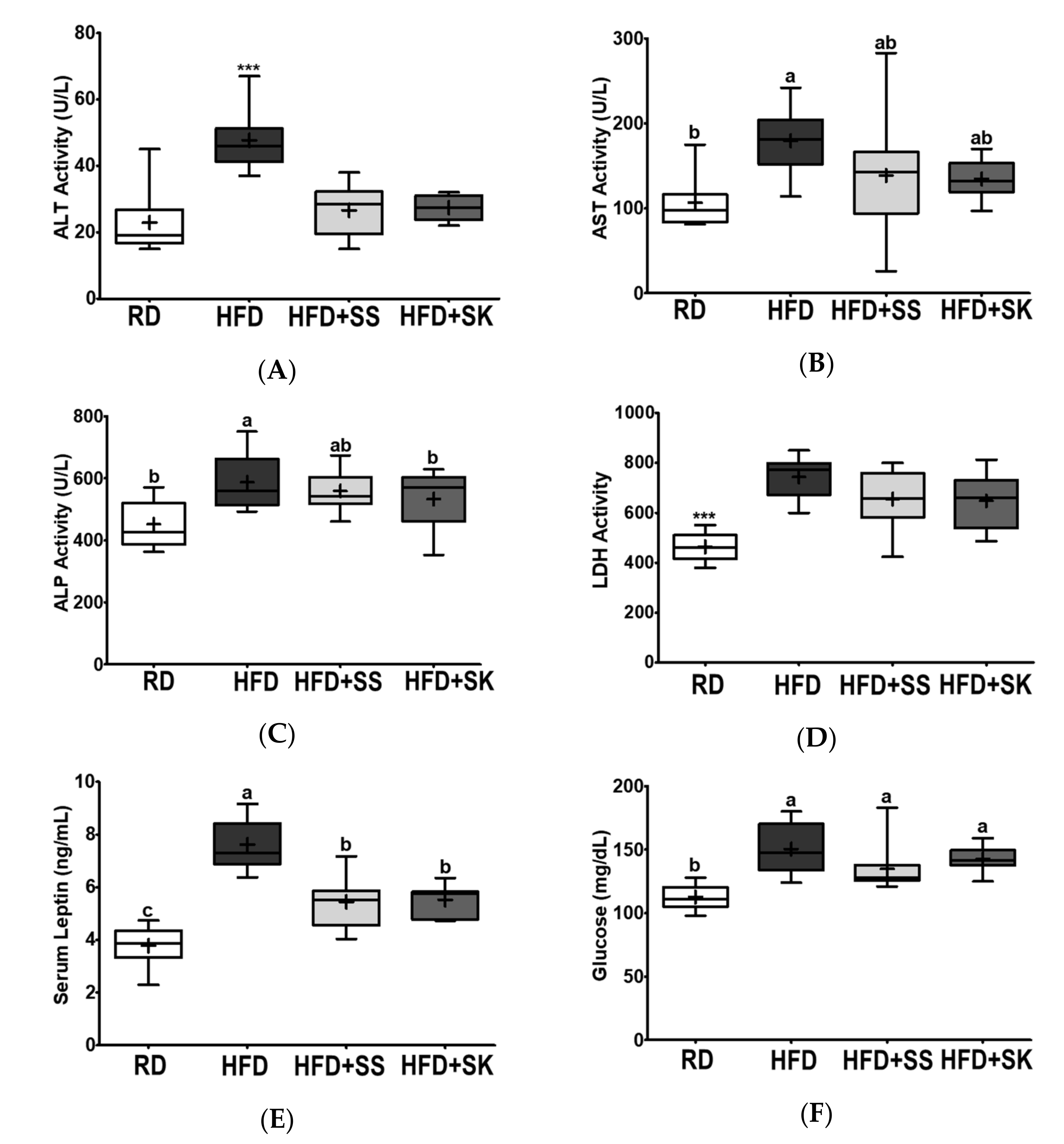
3.4. SS and SK Consumption Improved the Levels of Serum Metabolites Related to Liver Function and Lowered Leptin Level
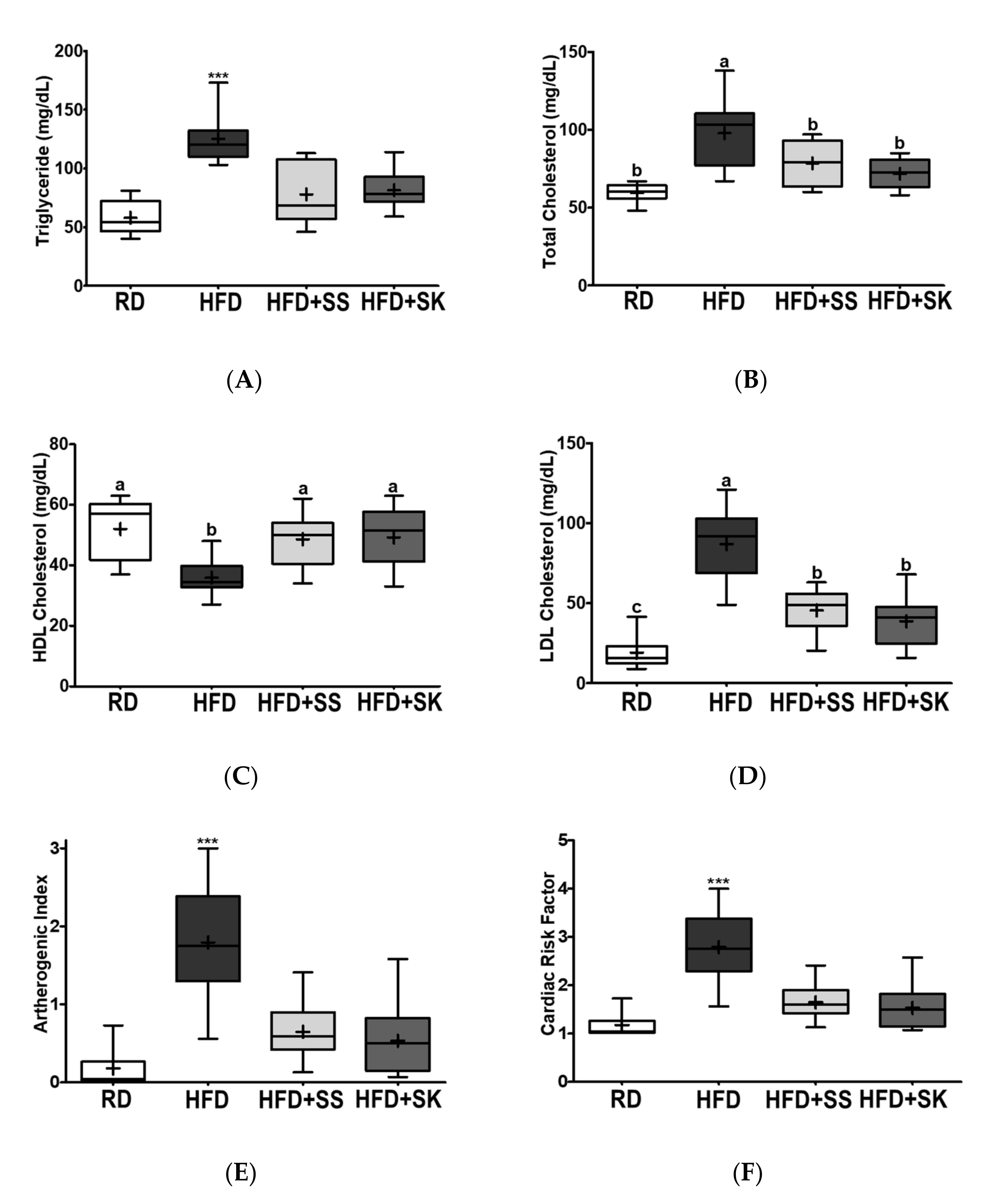
3.5. SS and SK Consumption Improved the Serum Lipid Profile
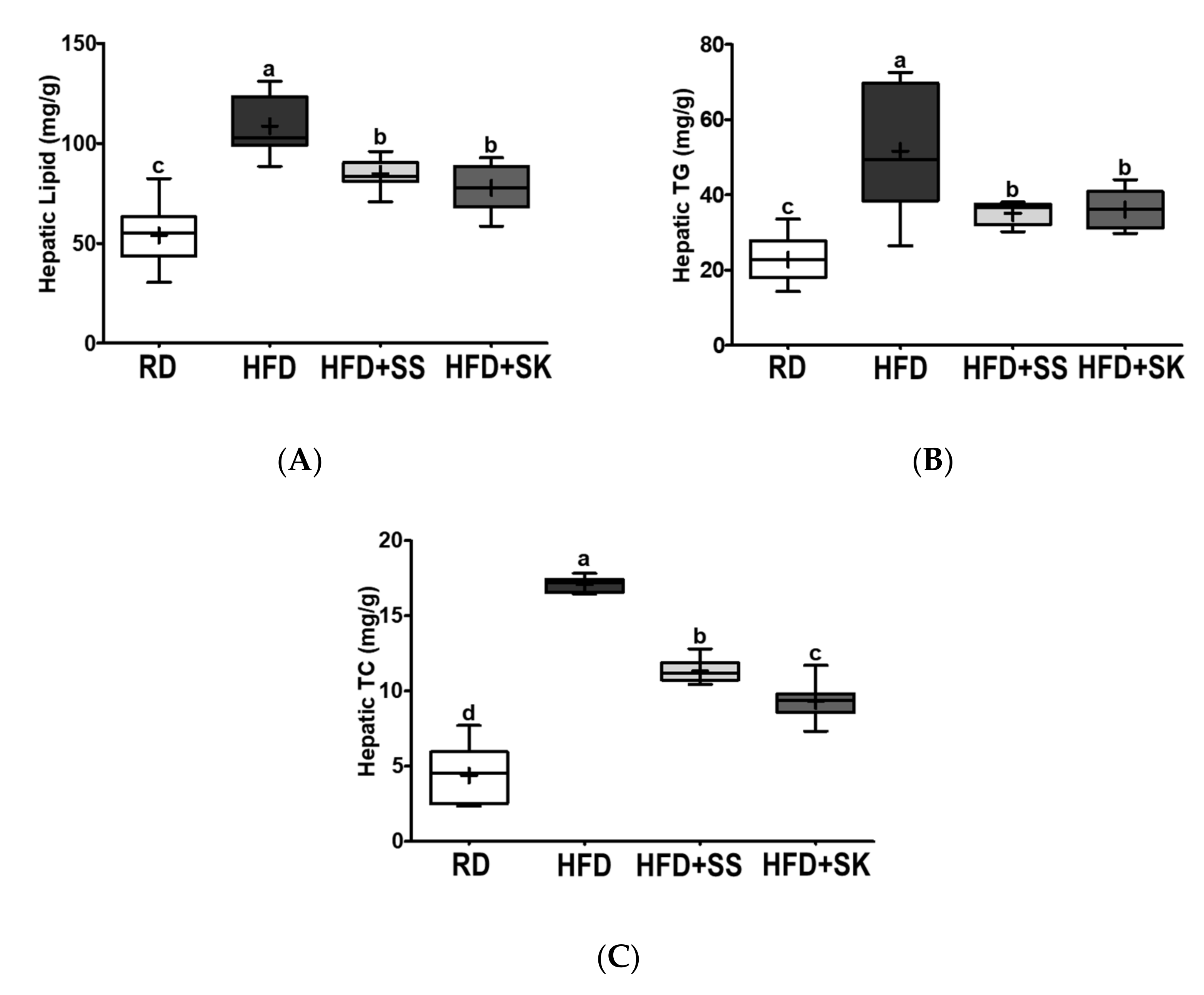
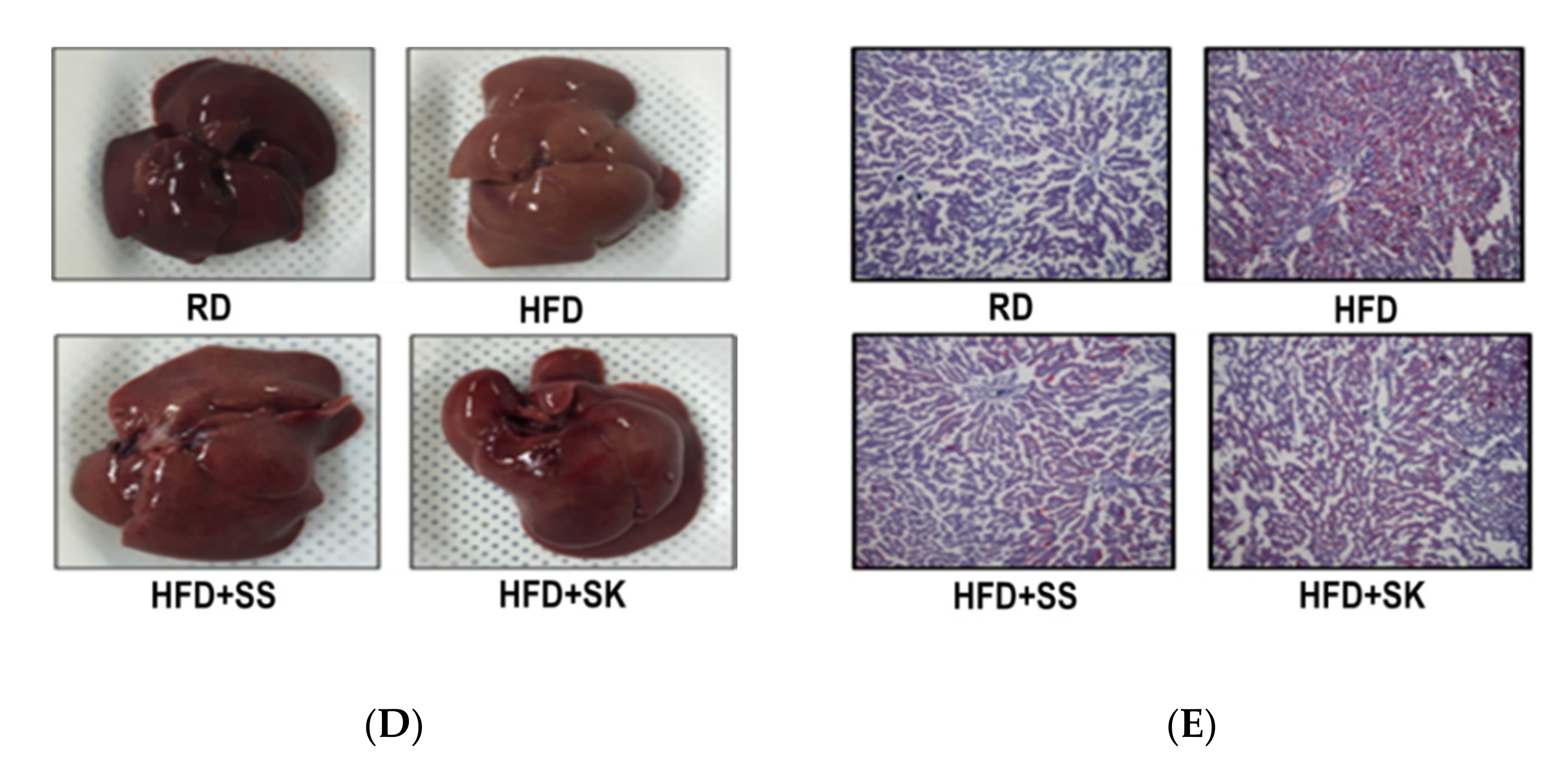
3.6. SS and SK Consumption Inhibited HFD-Induced Hepatic Lipid Accumulation
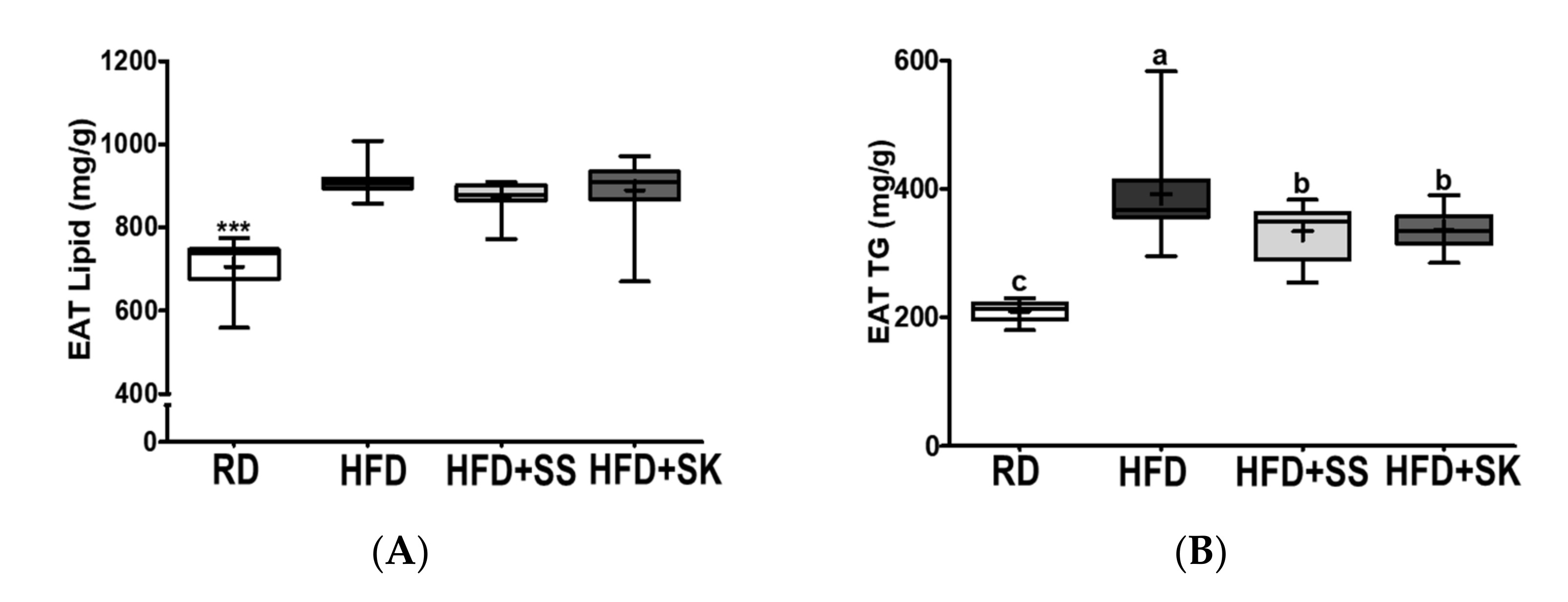
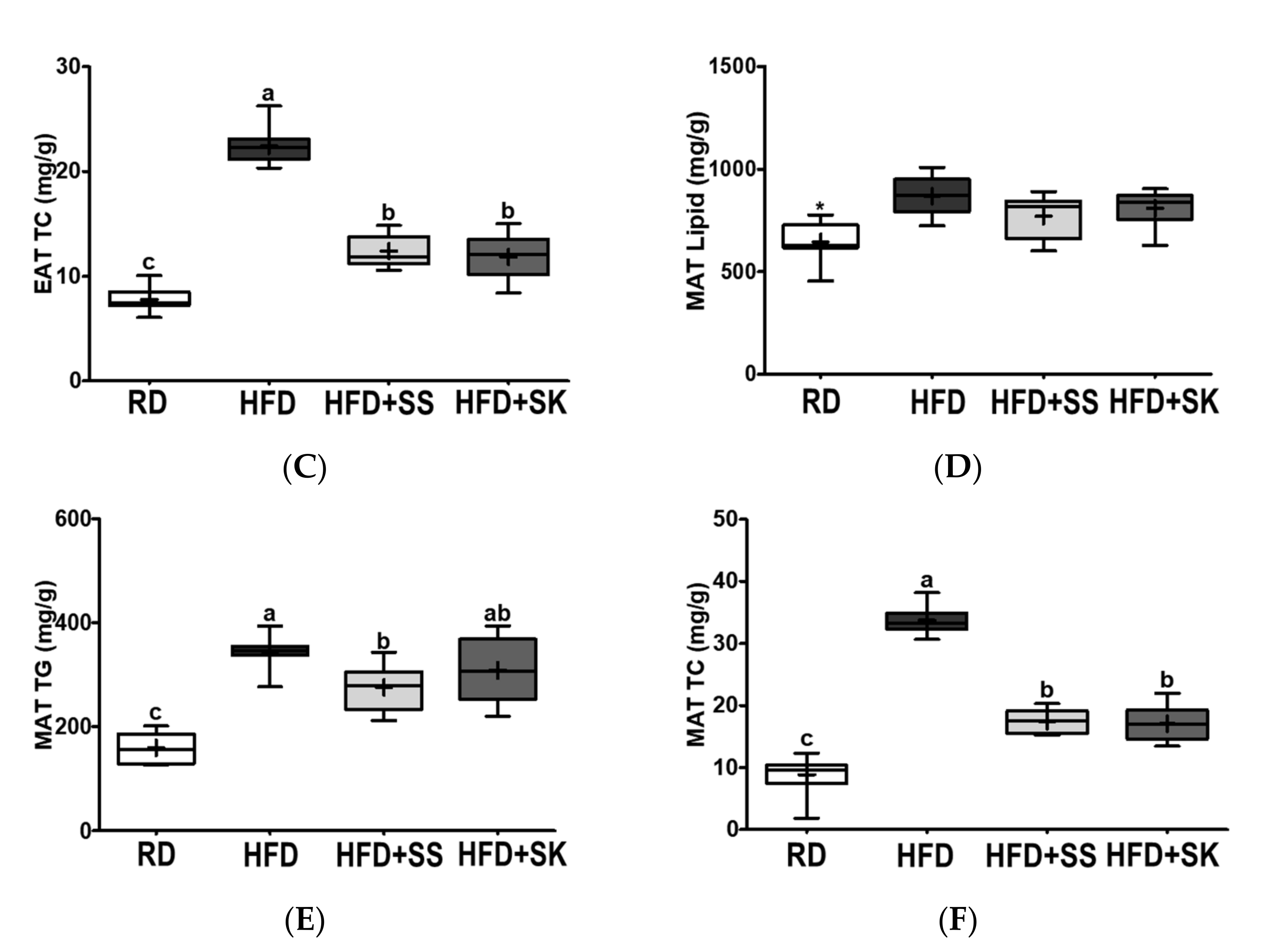
3.7. SS and SK Consumption Inhibited HFD-Induced Lipid Accumulation in White Adipose Tissue
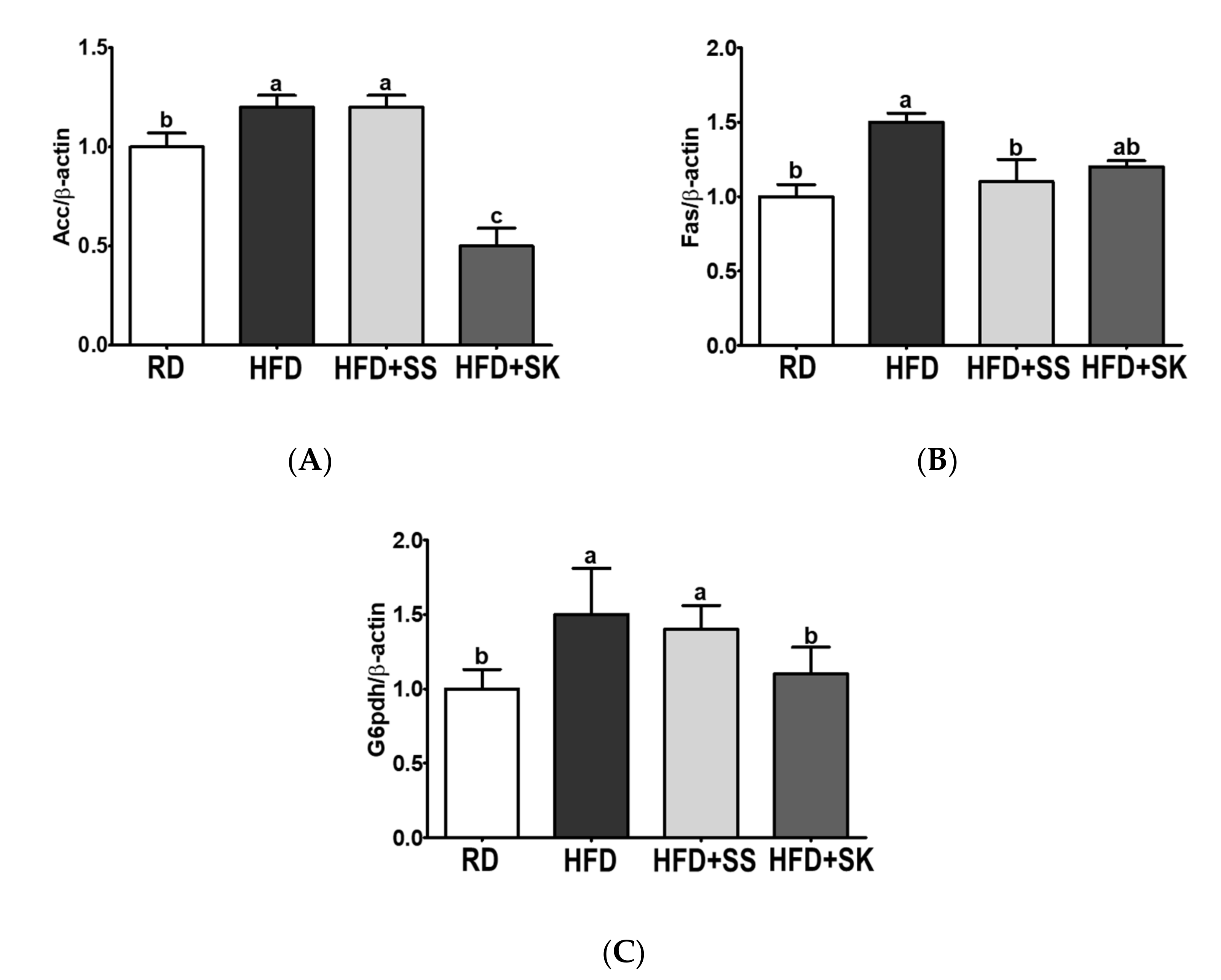
3.8. SS and SK Consumption Suppressed mRNA Expression Involved in Lipogenesis in EAT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbate, J.M.; Macrì, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of protein hydrolysates from anchovy (Engraulis Encrasicolus) waste for twelve weeks decreases metabolic dysfunction-associated fatty liver disease severity in ApoE-/-mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Li, D.; Guruvaiah, P.; Xu, N.; Xie, Z. Dietary supplement of large yellow tea ameliorates metabolic syndrome and attenuates hepatic steatosis in db/db mice. Nutrients 2018, 10, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, W.W.; Smith, E.O.; Stuff, J.E.; Hachey, D.L.; Heird, W.C.; Pownell, H.J. Cholesterol-lowering effect of soy protein in normocholesterolemic and hypercholesterolemic men. Am. J. Clin. Nutr. 1998, 68, 1385S–1389S. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.L.; Wang, X.Y.; Liu, F.C.; Chen, J.C.; Cao, J.; Li, J.X.; Hu, D.S.; Shen, C.; Lu, F.H.; Zhao, Y.X.; et al. Associations of soybean products intake with blood pressure changes and hypertension incidence: The China-PAR project. J. Geriatr. Cardiol. 2020, 17, 384–392. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Rajaratnam, V.; Mukherjee, S.; Das, S.K. Control of peripheral benzodiazepine receptor-mediated breast cancer in rats by soy protein. Mol. Carcinog. 2008, 47, 310–319. [Google Scholar] [CrossRef]
- Rai, A.K.; Jeyaram, K. Health benefits of functional proteins in fermented foods. In Health Benefits of Fermented Foods and Beverages, 1st ed.; Tamang, J.P., Ed.; CRC Press: Boca Raton, FL, USA, 2015; Chapter 14; pp. 455–474. [Google Scholar]
- Luh, B.S. Industrial production of soy sauce. J. Ind. Microbiol. 1995, 14, 467–471. [Google Scholar] [CrossRef]
- Lin, C.H.; Wei, Y.T.; Chou, C.C. Enhanced antioxidative activity of soybean koji prepared with various filamentous fungi. Food Microbiol. 2006, 23, 628–633. [Google Scholar] [CrossRef]
- Qureshi, K.; Abrams, G.A. Metabolic liver disease of obesity and role of adipose tissue in the pathogenesis of nonalcoholic fatty liver disease. World J. Gastroenterol. 2007, 13, 3540–3553. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [Green Version]
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 14 May 2021).
- Prevalence of Obesity and Severe Obesity among Adults: United States, 2017–2018. Available online: https://www.cdc.gov/nchs/products/databriefs/db360.htm (accessed on 14 May 2021).
- Jia, P.; Zhang, L.; Yu, W.; Yu, B.; Liu, M.; Zhang, D.; Yang, S. Impact of COVID-19 lockdown on activity patterns and weight status among youths in China: The COVID-19 impact on lifestyle change survey (COINLICS). Int. J. Obes. 2021, 45, 695–699. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, M.; Ji, Y.; Shi, Y.; Zhou, J.; Li, Q.; Qin, R.; Zhuang, X. "Stay-at-home" lifestyle effect on weight gain during the COVID-19 outbreak confinement in China. Int. J. Environ. Res. Public Health 2021, 18, 1813. [Google Scholar] [CrossRef]
- Williams, D.M.; Nawaz, A.; Evans, M. Drug therapy in obesity: A review of current and emerging treatments. Diabetes Ther. 2020, 11, 1199–1216. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; You, M.K.; Wang, Z.; Lee, Y.H.; Kim, H.A. Red pepper seed water extract suppresses high-fat diet-induced obesity in C57BL/6 mice. Food Sci. Biotechnol. 2019, 29, 275–281. [Google Scholar] [CrossRef]
- Mahmoud, R.H.; Elnour, W.A. Comparative evaluation of the efficacy of ginger and orlistat on obesity management, pancreatic lipase and liver peroxisomal catalase enzyme in male albino rats. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 75–83. [Google Scholar]
- Neyrinck, A.M.; Bindels, L.B.; Geurts, L.; Van Hul, M.; Cani, P.D.; Delzenne, N.M. A polyphenolic extract from green tea leaves activates fat browning in high-fat-diet-induced obese mice. J. Nutr. Biochem. 2017, 49, 15–21. [Google Scholar] [CrossRef]
- Sun, N.N.; Wu, T.Y.; Chau, C.F. Natural dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef]
- Velasquez, M.T.; Bhathena, S.J. Role of dietary soy protein in obesity. Int. J. Med. Sci. 2007, 4, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Zhao, A.; Wu, Y.; Zhao, Y.; Yang, X. Soybean soluble polysaccharides enhance bioavailability of genistein and its prevention against obesity and metabolic syndrome of mice with chronic high fat consumption. Food Funct. 2019, 10, 4153–4165. [Google Scholar] [CrossRef]
- Novel Bacillus Amyloliquefaciens CJ 14-6 Isolated from The Korean Traditional Meju and Manufacturing Method of Soybean Koji Using This Fungi and Soybean Koji Manufactured Therefrom. Available online: https://patents.google.com/patent/KR20170028044A/en (accessed on 14 May 2021).
- Shin, H.W.; Jang, E.S.; Moon, B.S.; Lee, J.J.; Lee, D.E.; Lee, C.H.; Shin, C.S. Anti-obesity effects of gochujang products prepared using rice koji and soybean meju in rats. J. Food Sci. Technol. 2016, 53, 1004–1013. [Google Scholar] [CrossRef] [Green Version]
- Waters Associates. Analysis of Amino Acid in Waters. PICO. TAG System; Young-in Scientific Co. Ltd.: Seoul, Korea, 1990; pp. 41–46. [Google Scholar]
- Van Wijngaarden, D. Modified rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem. 1967, 39, 848–849. [Google Scholar] [CrossRef]
- Park, Y.H.; Lee, J.J.; Son, H.K.; Kim, B.H.; Byun, J.; Ha, J.H. Antiobesity effects of extract from Spergularia marina griseb in adipocytes and high-fat diet-induced obese rats. Nutrients 2020, 12, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Rosenfeld, L. Lipoprotein analysis. Early methods in the diagnosis of atherosclerosis. Arch. Pathol. Lab. Med. 1989, 113, 1101–1110. [Google Scholar] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Biggs, H.G.; Erikson, J.M.; Moorehead, W.R. A manual colormetric assay of triglycerides in serum. Clin. Chem. 1975, 21, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Zlatkis, A.; Zak, B. Study of a new cholesterol reagent. Anal. Biochem. 1969, 29, 143–148. [Google Scholar] [CrossRef]
- Son, H.K.; Shin, H.W.; Jang, E.S.; Moon, B.S.; Lee, C.H.; Lee, J.J. Comparison of antiobesity effects between gochujangs produced using different koji products and tabasco hot sauce in rats fed a high-fat diet. J. Med. Food 2018, 21, 233–243. [Google Scholar] [CrossRef]
- Giannini, E.G.; Testa, R.; Savarino, V. Liver enzyme alteration: A guide for clinicians. Can. Med. Assoc. J. 2005, 172, 367–379. [Google Scholar] [CrossRef] [Green Version]
- D’souza, A.M.; Neumann, U.H.; Glavas, M.M.; Kieffer, T.J. The glucoregulatory actions of leptin. Mol. Metab. 2017, 6, 1052–1065. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Technol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Watanabe, K.; Igarashi, M.; Li, X.; Nakatani, A.; Miyamoto, J.; Inaba, Y.; Sutou, A.; Saito, T.; Sato, T.; Tachibana, N.; et al. Dietary soybean protein ameliorates high-fat diet-induced obesity by modifying the gut microbiota-dependent biotransformation of bile acids. PLoS ONE 2018, 13, e0202083. [Google Scholar] [CrossRef]
- Seo, H.S.; Lee, S.; Singh, D.; Shin, H.W.; Cho, S.A.; Lee, C.H. Untargeted metabolite profiling for koji-fermentative bioprocess unravels the effects of varying substrate types and microbial inocula. Food Chem. 2018, 266, 161–169. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, K.; LeBlanc, R.E.; Loh, D.; Schwartz, G.J.; Yu, Y.H. Increasing dietary leucine intake reduces diet-induced obesity and improves glucose and cholesterol metabolism in mice via multimechanisms. Diabetes 2007, 56, 1647–1654. [Google Scholar] [CrossRef] [Green Version]
- Pohle-Krauza, R.J.; Navia, J.L.; Madore, E.Y.; Nyrop, J.E.; Pelkman, C.L. Effects of L-phenylalanine on energy intake in overweight and obese women: Interactions with dietary restraint status. Appetite 2008, 51, 111–119. [Google Scholar] [CrossRef]
- Deglaire, A.; Moughan, P.J. Animal models for determining amino acid digestibility in humans—A review. Br. J. Nutr. 2012, 108, S273–S281. [Google Scholar] [CrossRef] [Green Version]
- Buettner, R.; Parhofer, K.G.; Woenckhaus, M.; Wrede, C.E.; Kunz-Schughart, L.A.; Schölmerich, J.; Bollheimer, L.C. Defining high-fat-diet rat models: Metabolic and molecular effects of different fat types. J. Mol. Endocrinol. 2006, 36, 485–501. [Google Scholar] [CrossRef] [Green Version]
- Frigolet, M.E.; Torres, N.; Uribe-Figueroa, L.; Rangel, C.; Jimenez-Sanchez, G.; Tovar, A.R. White adipose tissue genome wide-expression profiling and adipocyte metabolic functions after soy protein consumption in rats. J. Nutr. Biochem. 2011, 22, 118–129. [Google Scholar] [CrossRef]
- Choi, J.W. Association between elevated serum hepatic enzyme activity and total body fat in obese humans. Ann. Clin. Lab. Sci. 2003, 33, 257–264. [Google Scholar]
- Kim, S.E. Optimal evaluation of the results of liver function tests. Korean J. Fam. Med. 2019, 94, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Lee, Y.S.; Seol, D.J.; Cho, I.J.; Ku, S.K.; Choi, J.S.; Lee, H.J. Anti-obesity and fatty liver-preventing activities of Lonicera caerulea in high-fat diet-fed mice. Int. J. Mol. Med. 2018, 42, 3047–3064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamohara, S.; Burcelin, R.; Halaas, J.L.; Friedman, J.M.; Charron, M.J. Acute stimulation of glucose metabolism in mice by leptin treatment. Nature 1997, 389, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Wåhlen, K.; Sjölin, E.; Löfgren, P. Role of fat cell size for plasma leptin in a large population based sample. Exp. Clin. Endocrinol. Diabetes 2011, 119, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klop, B.; Elte, J.W.; Cabezas, M.C. Dyslipidemia in obesity: Mechanisms and potential targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [Green Version]
- Tushuizen, M.E.; Diamant, M.; Heine, R.J. Postprandial dysmetabolism and cardiovascular disease in type 2 diabetes. Postgrad. Med. J. 2005, 81, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Cheik, N.C.; Rossi, E.A.; Guerra, R.L.; Tenório, N.M.; Oller do Nascimento, C.M.; Viana, F.P.; Manzoni, M.S.; Carlos, I.Z.; Leão da Silva, P.; Vendramini, R.C.; et al. Effects of a ferment soy product on the adipocyte area reduction and dyslipidemia control in hypercholesterolemic adult male rats. Lipids Health Dis. 2008, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Anthony, M.S.; Clarkson, T.B.; Hughes, C.L.; Morgan, T.M.; Burke, G.L. Soybean isoflavones improve cardiovascular risk factors without affecting the reproductive system of peripubertal rhesus monkeys. J. Nutr. 1996, 126, 43–50. [Google Scholar] [CrossRef]
- Park, J.; Rho, H.K.; Kim, K.H.; Choe, S.S.; Lee, Y.S.; Kim, J.B. Overexpression of glucose-6-phosphate dehydrogenase is associated with lipid dysregulation and insulin resistance in obesity. Mol. Cell. Biol. 2005, 25, 5146–5157. [Google Scholar] [CrossRef] [Green Version]
- Berndt, J.; Kovacs, P.; Ruschke, K.; Klöting, N.; Fasshauer, M.; Schön, M.R.; Körner, A.; Stumvoll, M.; Blüher, M. Fatty acid synthase gene expression in human adipose tissue: Association with obesity and type 2 diabetes. Diabetologia 2007, 50, 1472–1480. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | RD (1) | HFD (2) | HFD+SS | HFD+SK | |
|---|---|---|---|---|---|
| Components | |||||
| Ingredient (g/kg) | |||||
| Casein | 200.000 | 200.000 | 188.84 | 188.00 | |
| L-cystine | 3.000 | 3.000 | 3.000 | 3.000 | |
| Corn starch | 397.486 | 262.46 | 256.88 | 258.47 | |
| Dextrose | 132.000 | 132.000 | 132.000 | 132.000 | |
| Sucrose | 100.000 | 100.000 | 100.000 | 100.000 | |
| Cellulose | 50.000 | 50.000 | 50.000 | 50.000 | |
| Lard | 135.000 | 135.000 | 135.000 | ||
| Soybean oil | 70.000 | 70.000 | 67.930 | 66.280 | |
| Mineral mix (3) | 35.000 | 35.000 | 35.000 | 35.000 | |
| Vitamin mix (4) | 10.000 | 10.000 | 10.000 | 10.000 | |
| Choline chloride | 2.500 | 2.500 | 2.500 | 2.500 | |
| tert-Butylhydroquinone | 0.014 | 0.040 | 0.040 | 0.040 | |
| Steamed soybean | 30.000 | ||||
| Soybean koji | 30.000 | ||||
| Total | 1000.0 | 1000.0 | 1000.0 | 1000.0 | |
| Total energy (kcal) | 4001.9 | 4674.8 | 4667.5 | 4665.5 | |
| Fat (kcal %) | 15.8 | 39.5 | 39.2 | 39.2 | |
| Transcript | Forward Primer | Reverse Primer |
|---|---|---|
| Acc | CAACGCCTTCACACCACCTT | AGCCCATTACTTCATCAAAGATCCT |
| Fas | GGAACTGAACGGCATTACTCG | CATGCCGTTATCAACTTGTCC |
| G6pdh | GTTTGGCAGCGGCAACTAA | GGCATCACCCTGGTACAACTC |
| β-actin | GTGGGGCGCCCCAGGCACCAGGGC | CTCCTTAATGTCACGCACGATTTC |
| Parameter (%) | Steamed Soybean | Soybean Koji |
|---|---|---|
| Moisture | 1.7 ± 0.17 *** | 3.5 ± 0.17 |
| Carbohydrate | 48.4 ± 2.96 * | 39.9 ± 1.96 |
| Crude protein | 38.5 ± 0.9 | 39.9 ± 2.1 |
| Crude fat | 6.9 ± 0.44 *** | 12.4 ± 0.61 |
| Ash | 4.5 ± 0.18 | 4.3 ± 0.11 |
| Dietary fiber | 37.3 ± 2.64 | 34.3 ± 1.29 |
| Amino Acid (mg%) | Steamed Soybean | Soybean Koji |
|---|---|---|
| Urea | 0.10 ± 0.01 *** | 0.38 ± 0.01 |
| Aspartic acid | 0.09 ± 0.00 *** | <LLOQ (1) |
| Threonine | 0.26 ± 0.00 *** | 0.49 ± 0.01 |
| Serine | 0.20 ± 0.01 ** | 0.1 ± 0.00 |
| Asparagine | 0.75 ± 0.02 ** | 0.6 ± 0.02 |
| Glutamic acid | 0.88 ± 0.03 *** | 5.93 ± 0.03 |
| α-aminoadipic acid | 0.53 ± 0.03 *** | <LLOQ |
| Proline | 0.72 ± 0.01 *** | <LLOQ |
| Glycine | 0.18 ± 0.01 ** | 0.26 ± 0.01 |
| Alanine | 2.16 ± 0.06 ** | 1.29 ± 0.06 |
| Citrulline | <LLOQ | 0.18 ± 0.01 |
| Valine | 0.96 ± 0.04 *** | 7.12 ± 0.27 |
| Methionine | <LLOQ | 0.74 ± 0.02 |
| Isoleucine | 0.54 ± 0.03 *** | 3.24 ± 0.10 |
| Leucine | 0.69 ± 0.02 *** | 10.6 ± 0.36 |
| Tyrosine | 1.15 ± 0.04 *** | 5.07 ± 0.14 |
| Phenylalanine | 2.08 ± 0.07 *** | 26.17 ± 0.73 |
| β-alanine | 0.29 ± 0.02 *** | <LLOQ |
| γ-amino-n-butyric acid | 1.95 ± 0.04 *** | 0.52 ± 0.01 |
| Histidine | 0.36 ± 0.01 *** | 1.26 ± 0.02 |
| Tryptophan | 0.85 ± 0.03 *** | 1.84 ± 0.09 |
| Arginine | 0.68 ± 0.07 ** | <LLOQ |
| Total | 15.42 ± 0.41 *** | 65.8 ± 2.21 |
| Fatty Acid (% Total Fatty Acids) | Steamed Soybean | Soybean Koji |
|---|---|---|
| Myristic acid (C14:0) | 0.15 ± 0.01 | 0.17 ± 0.02 |
| Pentadecanoic acid (C15:0) | <LLOQ (1) | 0.49 ± 0.06 |
| Palmitic acid (C16:0) | 13.80 ± 0.76 | 16.03 ± 0.61 |
| Heptadecanoic acid (C17:0) | 0.76 ± 0.02 ** | 0.52 ± 0.04 |
| Stearic acid (C18:0) | 5.53 ± 0.05 ** | 6.40 ± 0.07 |
| Arachidic acid (C20:0) | 0.59 ± 0.02 | 0.55 ± 0.20 |
| Heneicosanoic acid (C21:0) | 22.05 ± 0.18 *** | 10.72 ± 0.39 |
| Behenic acid (C22:0) | 0.61 ± 0.05 | 0.69 ± 0.03 |
| Lignoceric acid (C24:0) | 0.19 ± 0.01 | 0.22 ± 0.02 |
| Saturated | 43.67 ± 0.80 ** | 35.80 ± 0.27 |
| Myristoleic acid (C14:1) | <LLOQ | 0.16 ± 0.02 |
| Palmitoleic acid (C16:1) | 0.13 ± 0.01 | 0.16 ± 0.01 |
| Oleic acid (C18:1n9c) | 18.20 ± 0.07 ** | 20.39 ± 0.45 |
| cis-11-Eicosenoic acid (C20:1) | 0.36 ± 0.03 | 0.30 ± 0.01 |
| Monounsaturated | 18.69 ± 0.20 * | 21.01 ± 0.77 |
| Linolelaidic acid (C18:2n6t) | 0.28 ± 0.20 | <LLOQ |
| Linoleic acid (C18:2n6c) | 29.01 ± 0.56 * | 33.41 ± 0.88 |
| cis-11,14-Eicosadienoic acid (C20:2) | 0.09 ± 0.01 *** | <LLOQ |
| Linolenic acid (C18:3n3) | 7.66 ± 0.18 ** | 9.67 ± 0.35 |
| cis-5,8,11,14,17-Eicosapentaenoic acid (C20:5n3) | 0.61 ± 0.05 *** | 0.10 ± 0.02 |
| Polyunsaturated | 37.64 ± 0.67 ** | 43.19 ± 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Lee, J.-J.; Shin, H.-W.; Jung, S.; Ha, J.-H. Effect of Soybean and Soybean Koji on Obesity and Dyslipidemia in Rats Fed a High-Fat Diet: A Comparative Study. Int. J. Environ. Res. Public Health 2021, 18, 6032. https://doi.org/10.3390/ijerph18116032
Park S, Lee J-J, Shin H-W, Jung S, Ha J-H. Effect of Soybean and Soybean Koji on Obesity and Dyslipidemia in Rats Fed a High-Fat Diet: A Comparative Study. International Journal of Environmental Research and Public Health. 2021; 18(11):6032. https://doi.org/10.3390/ijerph18116032
Chicago/Turabian StylePark, Sihoon, Jae-Joon Lee, Hye-Won Shin, Sunyoon Jung, and Jung-Heun Ha. 2021. "Effect of Soybean and Soybean Koji on Obesity and Dyslipidemia in Rats Fed a High-Fat Diet: A Comparative Study" International Journal of Environmental Research and Public Health 18, no. 11: 6032. https://doi.org/10.3390/ijerph18116032