
Partial-Body Cryostimulation Increases Resting Energy Expenditure in Lean and Obese Women

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Design
2.2. Partial-Body Cryostimulation Procedure
2.3. Statistical Analysis
3. Results
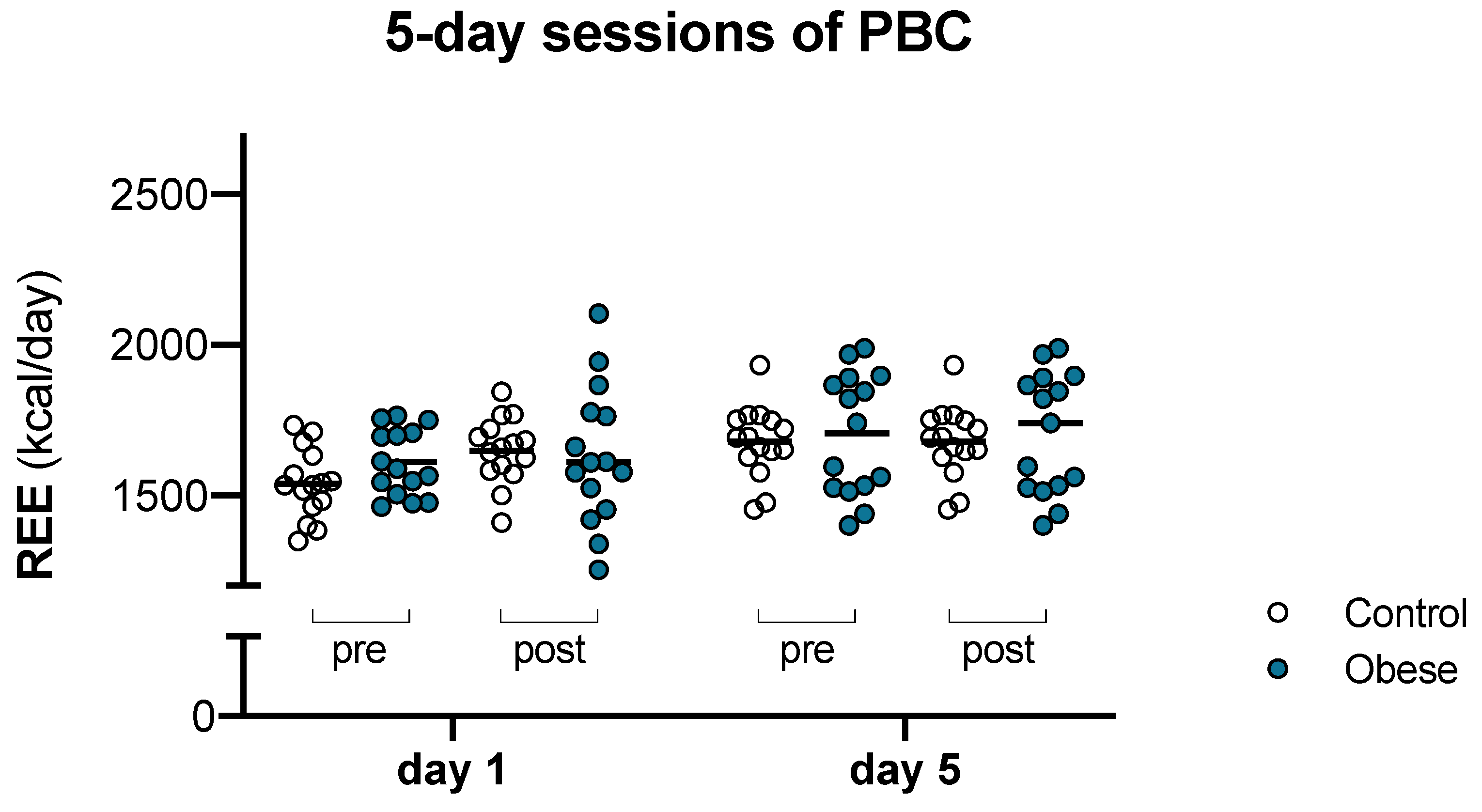
3.1. Multiple PBC Induced Incremental Response in REE in Lean and Obese Women
3.2. PBC Induced a Shift in Substrate Oxidation
4. Discussion
- Brown and Brown-Like Adipocytes Are Less Represented in Obese Compared to Lean Individuals
- Fat-Free Mass (FFM) and Chiefly Skeletal Muscles Are Likely to Provide a Great Contribution to Cold-Induced Thermogenesis
- Dysfunctional WAT Impairs Muscle Metabolism Likely Altering Obese Subjects’ Thermogenetic Capacity
- WAT Response to Cooling Is Probably Poorly Involved in REE Increase in Short-Term Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/en/newsroom/fact-sheets/detail/obesity-and-overweight (accessed on 20 December 2020).
- Maurizi, G.; Della Guardia, L.; Maurizi, A.; Poloni, A. Adipocytes properties and crosstalk with immune system in obesity-related inflammation. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef]
- Maurizi, G.; Petäistö, T.; Maurizi, A.; Della Guardia, L. Key-genes regulating the liposecretion process of mature adipocytes. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Giordano, A.; Frontini, A.; Cinti, S. Convertible visceral fat as a therapeutic target to curb obesity. Nat. Rev. Drug Discov. 2016. [Google Scholar] [CrossRef]
- Ouellet, V.; Labbé, S.M.; Blondin, D.P.; Phoenix, S.; Guérin, B.; Haman, F.; Turcotte, E.E.; Richard, D.; Carpentier, A.C. Brown adipose tissue oxidative metabolism contributes to energy expenditure during acute cold exposure in humans. J. Clin. Investig. 2012, 122, 545–552. [Google Scholar] [CrossRef]
- Rolfe, D.F.; Brown, G.C. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol. Rev. 1997, 77, 731–758. [Google Scholar] [CrossRef] [Green Version]
- Ruige, J.B.; Ballaux, D.P.; Funahashi, T.; Mertens, I.L.; Matsuzawa, Y.; Van Gaal, L.F. Resting metabolic rate is an important predictor of serum adiponectin concentrations: Potential implications for obesity-related disorders. Am. J. Clin. Nutr. 2005, 82, 21–25. [Google Scholar] [CrossRef]
- Astrup, A.; Buemann, B.; Toubro, S.; Ranneries, C.; Raben, A. Low resting metabolic rate in subjects predisposed to obesity: A role for thyroid status. Am. J. Clin. Nutr. 1996, 63, 879–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravussin, E.; Lillioja, S.; Knowler, W.C.; Christin, L.; Freymond, D.; Abbott, W.G.; Boyce, V.; Howard, B.V.; Bogardus, C. Reduced Rate of Energy Expenditure as a Risk Factor for Body-Weight Gain. New Engl. J. Med. 1988, 318, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.F.; Clegg, D.J. Non-shivering thermogenesis as a mechanism to facilitate sustainable weight loss. Obes. Rev. 2017, 18, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Browning, M.G.; Evans, R.K. The contribution of fat-free mass to resting energy expenditure: Implications for weight loss strategies in the treatment of adolescent obesity. Int. J. Adolesc. Med. Health 2015, 27, 241–246. [Google Scholar] [CrossRef]
- Vallerand, A.L.; Jacobs, I. Rates of energy substrates utilization during human cold exposure. Graefe’s Arch. Clin. Exp. Ophthalmol. 1989, 58, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Fonda, B.; De Nardi, M.; Sarabon, N. Effects of whole-body cryotherapy duration on thermal and cardio-vascular response. J. Therm. Biol. 2014, 42, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Lubkowska, A.; Dudzińska, W.; Bryczkowska, I.; Dołęgowska, B. Body Composition, Lipid Profile, Adipokine Concentration, and Antioxidant Capacity Changes during Interventions to Treat Overweight with Exercise Programme and Whole-Body Cryostimulation. Oxidative Med. Cell. Longev. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wiecek, M.; Szymura, J.; Sproull, J.; Szygula, Z. Whole-Body Cryotherapy Is an Effective Method of Reducing Abdominal Obesity in Menopausal Women with Metabolic Syndrome. J. Clin. Med. 2020, 9, 2797. [Google Scholar] [CrossRef] [PubMed]
- Bouzigon, R.; Grappe, F.; Ravier, G.D.B. Whole- and partial-body cryostimulation/cryotherapy: Current technologies and practical applications. J. Therm Biol. 2016, 61, 61–67. [Google Scholar] [CrossRef]
- Mannocci, A.; Di Thiene, D.; Del Cimmuto, A. International Physical Activity Questionnaire: Validation and assessment in an Italian sample. Ital. J. Public Heal. 2010, 7, 369–376. [Google Scholar]
- Ekelund, U.; Ward, A.H.; Norat, T.; Luan, J.; May, A.M.; Weiderpass, E.; Sharp, S.J.; Overvad, K.; Østergaard, J.N.; Tjønneland, A.; et al. Physical activity and all-cause mortality across levels of overall and abdominal adiposity in European men and women: The European Prospective Investigation into Cancer and Nutrition Study (EPIC). Am. J. Clin. Nutr. 2015, 101, 613–621. [Google Scholar] [CrossRef]
- Welch, A.A.; Luben, R.; Khaw, K.T.; Bingham, S.A. The CAFE computer program for nutritional analysis of the EPIC-Norfolk food frequency questionnaire and identification of extreme nutrient values. J. Hum. Nutr. Diet. 2005, 18, 99–116. [Google Scholar] [CrossRef]
- Dupertuis, Y.M.; Oshima, T.; Delsoglio, M.; Achamrah, N.; Heidegger, C.; Pichard, C. In Vitro validation of the new indirect calorimeter for very low O2 consumption and CO2 production. Clin. Nutr. 2018, 37, s175. [Google Scholar] [CrossRef]
- Groeneveld, R.A.; Meeden, G. Measuring Skewness and Kurtosis. J. R. Stat. Soc. Ser. D (Stat.) 1984, 33, 391. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Academic Press: New York, NY, USA, 1988. [Google Scholar]
- Beyer, H.; Tukey, J.W. Exploratory Data Analysis. Addison-Wesley Publishing Company Reading, Mass. Menlo Park, Cal., London, Amsterdam, Don Mills, Ontario, Sydney 1977, XVI, 688. 1981. S. Biom. J. 1981, 23, 413–414. [Google Scholar] [CrossRef]
- Loap, S.L.R. Mechanism Underlying Tissue Cryotherapy to Combat Obesity/Overweight: Triggering Thermogenesis. J. Obes. 2018, 2018, 5789647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, I.P.; Elliott, S.A.; Siervo, M.; Padwal, R.; Bertoli, S.; Battezzati, A.; Prado, C.M. Is Obesity Associated with Altered Energy Expenditure? Adv. Nutr. 2016, 7, 476–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehling, H.; Lusher, J. People with a body mass index ≥30 under-report their dietary intake: A systematic review. J. Health Psychol. 2019, 24, 2042–2059. [Google Scholar] [CrossRef] [PubMed]
- Nowack, J.; Giroud, S.; Arnold, W.; Ruf, T. Muscle Non-shivering Thermogenesis and Its Role in the Evolution of Endothermy. Front. Physiol. 2017, 8, 889. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, S.K.; Shaikh, S.A.; Sopariwala, D.H.; Bal, N.C.; Periasamy, M. Sarcolipin Protein Interaction with Sarco(endo)plasmic Reticulum Ca2+ATPase (SERCA) Is Distinct from Phospholamban Protein, and Only Sarcolipin Can Promote Uncoupling of the SERCA Pump. J. Biol. Chem. 2013, 288, 6881–6889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, M.A.; Pallubinsky, H.B.D. Functional characterization of human brown adipose tissue metabolism. Biochem, J. 2020, 477, 261–1286. [Google Scholar] [CrossRef] [Green Version]
- Blondin, D.P.; Daoud, A.; Taylor, T.; Tingelstad, H.C.; Bézaire, V.; Richard, D.; Carpentier, A.C.; Taylor, A.W.; Harper, M.-E.; Aguer, C.; et al. Four-week cold acclimation in adult humans shifts uncoupling thermogenesis from skeletal muscles to brown adipose tissue. J. Physiol. 2017, 595, 2099–2113. [Google Scholar] [CrossRef]
- Dulian, K.; Laskowski, R.; Grzywacz, T.; Kujach, S.; Flis, D.J.; Smaruj, M.; Ziemann, E. The whole body cryostimulation modifies irisin concentration and reduces inflammation in middle aged, obese men. Cryobiology 2015, 71, 398–404. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, R.; Meng, Y.; Li, S.; Donelan, W.; Zhao, Y.; Qi, L.; Zhang, M.; Wang, X.; Cui, T.; et al. Irisin Stimulates Browning of White Adipocytes through Mitogen-Activated Protein Kinase p38 MAP Kinase and ERK MAP Kinase Signaling. Diabetes 2014, 63, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfannenberg, C.; Werner, M.K.; Ripkens, S.; Stef, I.; Deckert, A.; Schmadl, M.; Reimold, M.; Häring, H.-U.; Claussen, C.D.; Stefan, N. Impact of Age on the Relationships of Brown Adipose Tissue with Sex and Adiposity in Humans. Diabetes 2010, 59, 1789–1793. [Google Scholar] [CrossRef] [Green Version]
- Yoneshiro, T.; Aita, S.; Matsushita, M.; Kayahara, T.; Kameya, T.; Kawai, Y.; Iwanaga, T.; Saito, M. Recruited brown adipose tissue as an antiobesity agent in humans. J. Clin. Investig. 2013, 123, 3404–3408. [Google Scholar] [CrossRef] [Green Version]
- Jung, R.T.; Shetty, P.S.; James, W.P.T.; Barrand, M.A.; Callingham, B.A. Reduced thermogenesis in obesity. Nat. Cell Biol. 1979, 279, 322–323. [Google Scholar] [CrossRef]
- Periasamy, M.; Maurya, S.K.; Sahoo, S.K.; Singh, S.; Reis, F.C.G.; Bal, N.C. Role of SERCA Pump in Muscle Thermogenesis and Metabolism. Compr. Physiol. 2017, 7, 879–890. [Google Scholar] [CrossRef]
- Thurlby, P.L.; Trayhurn, P. The role of thermoregulatory thermogenesis in the development of obesity in genetically-obese (ob/ob) mice pair-fed with lean siblings. B. Br. J. Nutr. 1979, 42, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Ballantyne, C.M. Skeletal muscle inflammation and insulin resistance in obesity. J. Clin. Investig. 2017, 127, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Berggren, J.R.; Hulver, M.W.; Dohm, G.L.; Houmard, J.A. Weight Loss and Exercise: Implications for Muscle Lipid Metabolism and Insulin Action. Med. Sci. Sports Exerc. 2004, 36, 1191–1195. [Google Scholar] [CrossRef]
- de Mello, A.H.; Costa, A.B.; Engel, J.D.G.; Rezin, G.T. Mitochondrial dysfunction in obesity. Life Sci. 2018, 192, 26–32. [Google Scholar] [CrossRef]
- Wold, L.E.; Dutta, K.; Mason, M.M.; Ren, J.; Cala, S.E.; Schwanke, M.L.; Davidoff, A.J. Impaired SERCA function contributes to cardiomyocyte dysfunction in insulin resistant rats. J. Mol. Cell. Cardiol. 2005, 39, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Chakraborty, S.; Muthuchamy, M. Roles of sarcoplasmic reticulum Ca2+ ATPase pump in the impairments of lymphatic contractile activity in a metabolic syndrome rat model. Sci. Rep. 2020, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Corpeleijn, E.; Mensink, M.; Kooi, M.E.; Roekaerts, P.M.; Saris, W.H.; Blaak, E.E. Impaired Skeletal Muscle Substrate Oxidation in Glucose-intolerant Men Improves After Weight Loss. Obesity 2008, 16, 1025–1032. [Google Scholar] [CrossRef]
- Flachs, P.; Rossmeisl, M.; Kuda, O.; Kopecky, J. Stimulation of mitochondrial oxidative capacity in white fat independent of UCP1: A key to lean phenotype. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2013, 1831, 986–1003. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; You, W.; Zhou, Y.; Chen, W.; Wang, Y.; Shan, T. Cold-induced lipid dynamics and transcriptional programs in white adipose tissue. BMC Biol. 2019, 17, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, A.W.; Csikasz, R.I.; von Essen, G.; Cannon, B.; Nedergaard, J. No insulating effect of obesity. Am. J. Physiol Endocrinol Metab. 2016, 311, E202–E213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olichon-Berthe, C.; Van Obberghen, E.; Le Marchand-Brustel, Y. Effect of cold acclimation on the expression of glucose transporter Glut 4. Mol. Cell. Endocrinol. 1992, 89, 11–18. [Google Scholar] [CrossRef]
- Shin, H.; Ma, Y.; Chanturiya, T.; Cao, Q.; Wang, Y.; Kadegowda, A.K.; Jackson, R.; Rumore, D.; Xue, B.; Shi, H.; et al. Lipolysis in Brown Adipocytes Is Not Essential for Cold-Induced Thermogenesis in Mice. Cell Metab. 2017, 26, 764–777.e5. [Google Scholar] [CrossRef] [Green Version]
- Chitraju, C.; Fischer, A.W.; Farese, R.V.; Walther, T.C. Lipid Droplets in Brown Adipose Tissue Are Dispensable for Cold-Induced Thermogenesis. Cell Rep. 2020, 33, 108348. [Google Scholar] [CrossRef]
- Geer, E.B.; Shen, W. Gender differences in insulin resistance, body composition, and energy balance. Gend. Med. 2009, 6, 60–75. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Lean (n = 16) | Obese (n = 15) | p Value | |
|---|---|---|---|
| Age (years) | 40.4 ± 6 | 42.7 ± 10.4 | 0.46 |
| Height (cm) | 162.6 ± 5.5 | 163.7 ± 6.8 | 0.64 |
| Weight (kg) | 58.3 ± 4.5 | 83.2 ± 7.3 | <0.0001 |
| BMI (kg/m2) | 22 ± 1.7 | 31 ± 1.6 | <0.0001 |
| Basal REE * (kcal/day) | 1538 ± 111 | 1610 ± 110 | <0.0001 |
| Physical Activity (MET **-min/week) | 1760 ± 1660 | 2149 ± 934 | 0.7 |
| Dietary intake (kcal/day) | 1254 ± 191 | 1432 ± 820 | 0.67 |
| Lean | Obese | |||||||
|---|---|---|---|---|---|---|---|---|
| Day 1 | Day 5 | Day 1 | Day 5 | |||||
| Pre | Post | Pre | Post | Pre | Post | Pre | Post | |
| O2 (mL/min) | 224 ± 15 | 235 ± 17 | 244 ± 15 | 242 ± 17 | 234 ± 17 | 234 ± 37 | 248 ± 21 | 244 ± 31 |
| CO2 (mL/min) | 176 ± 20 | 203 ± 16 | 186 ± 20 | 197 ± 20 | 186 ± 17 | 201 ± 23 | 193 ± 23 | 212 ± 24 |
| RQ (CO2/O2) | 0.79 ± 0.06 | 0.86 ± 0.08 | 0.76 ± 0.07 | 0.82 ± 0.07 | 0.80 ± 0.08 | 0.87 ± 0.11 | 0.78 ± 0.09 | 0.87 ± 0.07 |
| REE (kcal/day) | 1538 ± 111 | 1645 ± 107 | 1665 ± 106 | 1676 ± 115 | 1610 ± 110 | 1632 ± 229 | 1698 ± 142 | 1706 ± 203 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Nardi, M.; Bisio, A.; Della Guardia, L.; Facheris, C.; Faelli, E.; La Torre, A.; Luzi, L.; Ruggeri, P.; Codella, R. Partial-Body Cryostimulation Increases Resting Energy Expenditure in Lean and Obese Women. Int. J. Environ. Res. Public Health 2021, 18, 4127. https://doi.org/10.3390/ijerph18084127
De Nardi M, Bisio A, Della Guardia L, Facheris C, Faelli E, La Torre A, Luzi L, Ruggeri P, Codella R. Partial-Body Cryostimulation Increases Resting Energy Expenditure in Lean and Obese Women. International Journal of Environmental Research and Public Health. 2021; 18(8):4127. https://doi.org/10.3390/ijerph18084127
Chicago/Turabian StyleDe Nardi, Massimo, Ambra Bisio, Lucio Della Guardia, Carlo Facheris, Emanuela Faelli, Antonio La Torre, Livio Luzi, Piero Ruggeri, and Roberto Codella. 2021. "Partial-Body Cryostimulation Increases Resting Energy Expenditure in Lean and Obese Women" International Journal of Environmental Research and Public Health 18, no. 8: 4127. https://doi.org/10.3390/ijerph18084127