
In Touch with the Heartbeat: Newborns’ Cardiac Sensitivity to Affective and Non-Affective Touch


Abstract
:1. Introduction
2. Materials and Methods
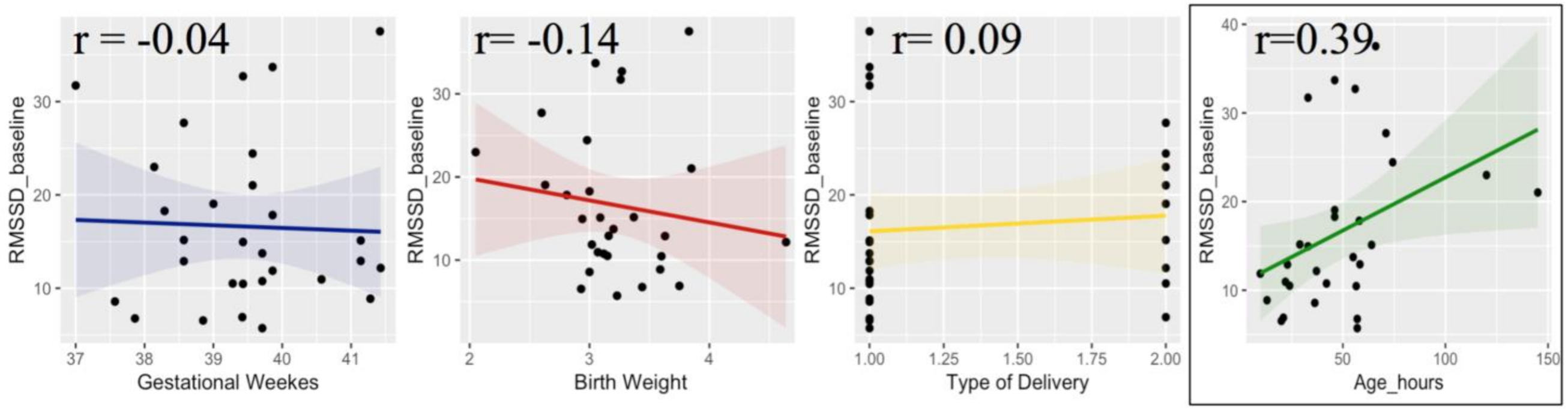
2.1. Participants
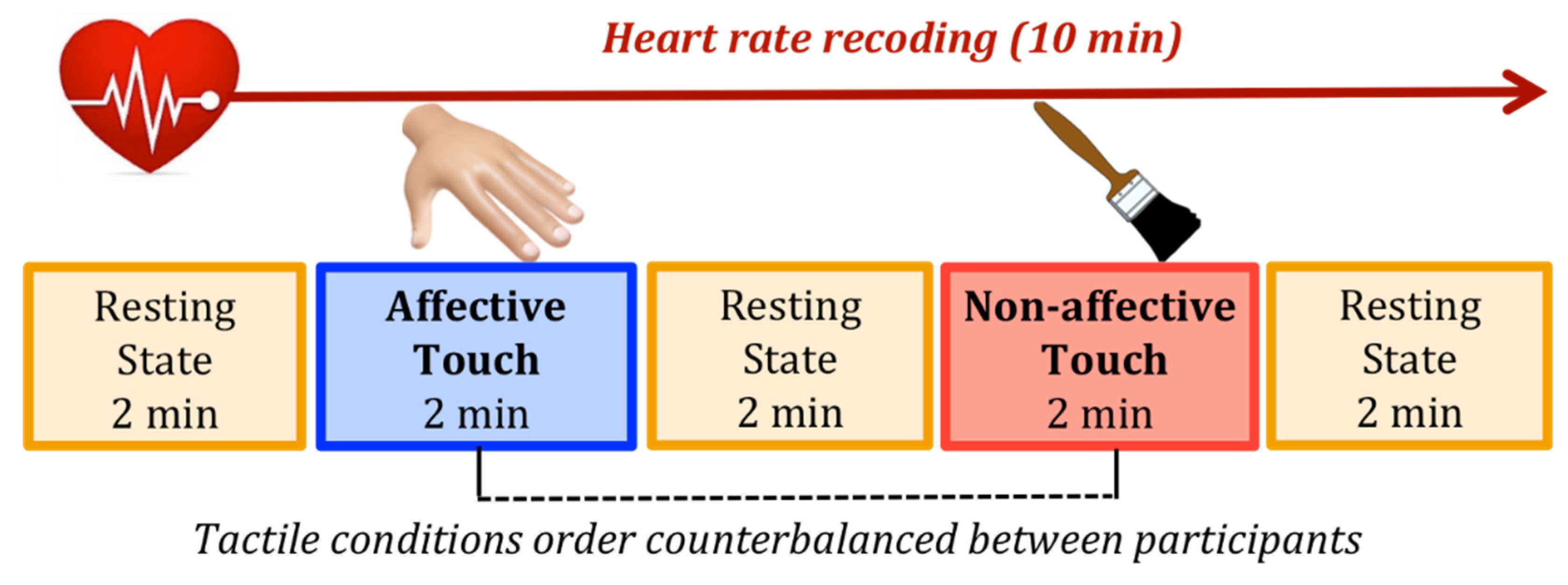
2.2. Stimuli and Procedure
2.3. Electrophysiological Data Recording and Processing
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bornstein, M.H.; Suess, P.E. Physiological Self-Regulation and Information Processing in Infancy: Cardiac Vagal Tone and Habituation. Child Dev. 2000, 71, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Calkins, S.D.; Fox, N.A. Self-regulatory processes in early personality development: A multilevel approach to the study of childhood social withdrawal and aggression. Dev. Psychopathol. 2002, 14, 477–498. [Google Scholar] [CrossRef] [Green Version]
- Feldman, R. The development of regulatory functions from birth to 5 years: Insights from premature infants. Child Dev. 2009, 80, 544–561. [Google Scholar] [CrossRef] [PubMed]
- Dereymaeker, A.; Pillay, K.; Vervisch, J.; De Vos, M.; Van Huffel, S.; Jansen, K.; Naulaers, G. Review of sleep-EEG in preterm and term neonates. Early Hum. Dev. 2017, 113, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.F.; Lucas, R.; White-Traut, R.C. Concept clarification of neonatal neurobehavioural organization. J. Adv. Nurs. 2008, 61, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Fox, N.A.; Calkins, S.D. The Development of Self-Control of Emotion: Intrinsic and Extrinsic Influences. Motiv. Emot. 2003, 27, 7–26. [Google Scholar] [CrossRef]
- Georgoulas, A.; Jones, L.; Laudiano-Dray, M.P.; Meek, J.; Fabrizi, L.; Whitehead, K. Sleep-wake regulation in preterm and term infants. Sleep 2021, 44, zsaa148. [Google Scholar] [CrossRef]
- Feldman, R.; Weller, A.; Sirota, L.; Eidelman, A.I. Skin-to-Skin contact (Kangaroo care) promotes self-regulation in premature infants: Sleep-wake cyclicity, arousal modulation, and sustained exploration. Dev. Psychol. 2002, 38, 194–207. [Google Scholar] [CrossRef]
- Schneider, U.; Bode, F.; Schmidt, A.; Nowack, S.; Rudolph, A.; Doelcker, E.-M.; Schlattmann, P.; Götz, T.; Hoyer, D. Deve-lopmental milestones of the autonomic nervous system revealed via longitudinal monitoring of fetal heart rate variability. PLoS ONE 2018, 13, e0200799. [Google Scholar] [CrossRef]
- Berntson, G.G.; Bigger, J.T.J.; Eckberg, D.L.; Grossman, P.; Kaufmann, P.G.; Malik, M.; Nagaraja, H.N.; Porges, S.W.; Saul, J.P.; Stone, P.H.; et al. Heart rate variability: Origins, methods, and interpretive caveats. Psychophysiology 1997, 34, 623–648. [Google Scholar] [CrossRef]
- McCraty, R.; Shaffer, F. Heart Rate Variability: New Perspectives on Physiological Mechanisms, Assessment of Self-regulatory Capacity, and Health risk. Glob. Adv. Health Med. 2015, 4, 46–61. [Google Scholar] [CrossRef] [Green Version]
- Chiera, M.; Cerritelli, F.; Casini, A.; Barsotti, N.; Boschiero, D.; Cavigioli, F.; Corti, C.G.; Manzotti, A. Heart Rate Variability in the Perinatal Period: A Critical and Conceptual Review. Front. Neurosci. 2020, 14, 561186. [Google Scholar] [CrossRef] [PubMed]
- Berntson, G.G.; Lozano, D.L.; Chen, Y.-J. Filter properties of root mean square successive difference (RMSSD) for heart rate. Psychophysiology 2005, 42, 246–252. [Google Scholar] [CrossRef]
- Shaffer, F.; Ginsberg, J.P. An Overview of Heart Rate Variability Metrics and Norms. Front. Public Health 2017, 5, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, O.M.; Korotchikova, I.; Lightbody, G.; Marnane, W.; Kerins, D.; Boylan, G.B. Heart rate variability during sleep in healthy term newborns in the early postnatal period. Physiol. Meas. 2009, 30, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Javorka, K.; Javorka, M.; Tonhajzerova, I.; Calkovska, A.; Lehotska, Z.; Bukovinska, Z.; Zibolen, M. Determinants of Heart Ra-te in Newborns. Acta Med. Martiniana 2011, 11, 7–16. [Google Scholar] [CrossRef]
- Van Vonderen, J.J.; Roest, A.A.; Siew, M.L.; Walther, F.J.; Hooper, S.B.; te Pas, A.B. Measuring physiological changes during the transi-tion to life after birth. Neonatology 2014, 105, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Kozar, M.; Tonhajzerova, I.; Mestanik, M.; Matasova, K.; Zibolen, M.; Calkovska, A.; Javorka, K. Heart rate variability in heal-thy term newborns is related to delivery mode: A prospective observational study. BMC Pregnancy Childbirth 2018, 18, 264. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.; Silva, M.J.; Guimarães, H. Autonomic nervous system in newborns: A review based on heart rate variability. Child’s Nerv. Syst. ChNS Off. J. Int. Soc. Pediatric Neurosurg. 2017, 33, 1053–1063. [Google Scholar] [CrossRef]
- Porges, S.W. Orienting in a defensive world: Mammalian modifications of our evolutionary heritage. A Polyvagal Theory. Psychophysiology 1995, 32, 301–318. [Google Scholar] [CrossRef]
- Thayer, J.F.; Hansen, A.L.; Saus-Rose, E.; Johnsen, B.H. Heart rate variability, prefrontal neural function, and cognitive per-formance: The neurovisceral integration perspective on self-regulation, adaptation, and health. Ann. Behav. Med. 2009, 37, 141–153. [Google Scholar] [CrossRef]
- Fox, N.A. Heart-rate variability and behavioral reactivity: Individual differences in autonomic patterning and their relation to infant and child temperament. In Perspectives on Behavioral Inhibition; University of Chicago Press: Chicago, IL, USA, 1989; pp. 177–195. [Google Scholar]
- Holzman, J.B.; Bridgett, D.J. Heart rate variability indices as bio-markers of top-down self-regulatory mechanisms: A me-ta-analytic review. Neurosci. Biobehav. Rev. 2017, 74, 233–255. [Google Scholar] [CrossRef]
- Grossman, P. Respiration, Stress, and Cardiovascular Function. Psychophysiology 1983, 20, 284–300. [Google Scholar] [CrossRef]
- Feldman, R.; Eidelman, A.I. Skin-to-skin contact (Kangaroo Care) accelerates autonomic and neurobehavioural maturation in preterm infants. Dev. Med. Child Neurol. 2003, 45, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Hostinar, C.E.; Sullivan, R.M.; Gunnar, M.R. Psychobiological mechanisms underlying the social buffering of the hypothala-mic-pituitary-adrenocortical axis: A review of animal models and human studies across development. Psychol. Bull. 2014, 140, 256–282. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, K.A.; Sheridan, M.A.; Tibu, F.; Fox, N.A.; Zeanah, C.H.; Nelson, C.A. Causal effects of the early caregiving envi-ronment on development of stress response systems in children. Proc. Natl. Acad. Sci. USA 2015, 112, 5637–5642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunbar, R.I.M. The social role of touch in humans and primates: Behavioural function and neurobiological mechanisms. Neurosci. Biobehav. Rev. 2010, 34, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, C.M.; Schanberg, S.M. Responses to maternal separation: Mechanisms and mediators. Int. J. Dev. Neurosci. 1998, 16, 261–270. [Google Scholar] [CrossRef]
- Laudenslager, M.L.; Boccia, M.L.; Berger, C.L.; Gennaro-Ruggles, M.M.; McFerran, B.; Reite, M.L. Total cortisol, free cortisol, and growth hormone associated with brief social separation experiences in young macaques. Dev. Psychobiol. 1995, 28, 199–211. [Google Scholar] [CrossRef]
- Liu, D.; Caldji, C.; Sharma, S.; Plotsky, P.M.; Meaney, M.J. Influence of neonatal rearing conditions on stress-induced adreno-corticotropin responses and norepinepherine release in the hypothalamic paraventricular nucleus. J. Neuroendocrinol. 2000, 12, 5–12. [Google Scholar] [CrossRef]
- Liu, D.; Diorio, J.; Tannenbaum, B.; Caldji, C.; Francis, D.; Freedman, A.; Sharma, S.; Pearson, D.; Plotsky, P.M.; Meaney, M.J. Maternal Care, Hippocampal Glucocorticoid Receptors, and Hypothalamic-Pituitary-Adrenal Responses to Stress. Science 1997, 277, 1659–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferber, S.G.; Makhoul, I.R. The effect of skin-to-skin contact (kangaroo care) shortly after birth on the neurobehavioral re-sponses of the term newborn: A randomized, controlled trial. Pediatrics 2004, 113, 858–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winberg, J. Mother and newborn baby: Mutual regulation of physiology and behavior—A selective review. Dev. Psychobiol. 2005, 47, 217–229. [Google Scholar] [CrossRef]
- Fairhurst, M.T.; Löken, L.; Grossmann, T. Physiological and behavioral responses reveal 9-month-old infants’ sensitivity to pleasant touch. Psychol. Sci. 2014, 25, 1124–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uvnäs-Moberg, K.; Bruzelius, G.; Alster, P.; Lundeberg, T. The antinociceptive effect of non-noxious sensory stimulation is mediated partly through oxytocinergic mechanisms. Acta Physiol. Scand. 1993, 149, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Singer, M.; Zagoory, O. Touch attenuates infants’ physiological reactivity to stress. Dev. Sci. 2010, 13, 271–278. [Google Scholar] [CrossRef]
- Stack, D.M.; Muir, D.W. Adult Tactile Stimulation during Face-to-Face Interactions Modulates Five-Month-Olds’ Affect and Attention. Child Dev. 1992, 63, 1509–1525. [Google Scholar] [CrossRef]
- Hertenstein, M.J.; Campos, J.J. Emotion Regulation Via Maternal Touch. Infancy 2001, 2, 549–566. [Google Scholar] [CrossRef] [Green Version]
- Jahromi, L.B.; Putnam, S.P.; Stifter, C.A. Maternal Regulation of Infant Reactivity From 2 to 6 Months. In Developmental Psy-chology; American Psychological Association: Washington, DC, USA, 2004; Volume 40, pp. 477–487. [Google Scholar] [CrossRef] [Green Version]
- Kaffashi, F.; Scher, M.S.; Ludington-Hoe, S.M.; Loparo, K.A. An analysis of the kangaroo care intervention using neonatal EEG complexity: A preliminary study. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2013, 124, 238–246. [Google Scholar] [CrossRef]
- Scher, M.S.; Ludington-Hoe, S.; Kaffashi, F.; Johnson, M.W.; Holditch-Davis, D.; Loparo, K.A. Neurophysiologic assessment of brain maturation after an 8-week trial of skin-to-skin contact on preterm infants. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2009, 120, 1812–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bystrova, K.; Ivanova, V.; Edhborg, M.; Matthiesen, A.-S.; Ransjö-Arvidson, A.-B.; Mukhamedrakhimov, R.; Uvnäs-Moberg, K.; Widström, A.-M. Early Contact versus Separation: Effects on Mother–Infant Interaction One Year Later. Birth 2009, 36, 97–109. [Google Scholar] [CrossRef]
- Diego, M.A.; Field, T.; Hernandez-Reif, M. Vagal Activity, Gastric Motility, and Weight Gain in Massaged Preterm Neonates. J. Pediatr. 2005, 147, 50–55. [Google Scholar] [CrossRef]
- Manzotti, A.; Cerritelli, F.; Esteves, J.E.; Lista, G.; Lombardi, E.; La Rocca, S.; Gallace, A.; McGlone, F.P.; Walker, S.C. Dynamic touch reduces physiological arousal in preterm infants: A role for c-tactile afferents? Dev. Cogn. Neurosci. 2019, 39, 100703. [Google Scholar] [CrossRef]
- McGlone, F.; Wessberg, J.; Olausson, H. Discriminative and Affective Touch: Sensing and Feeling. Neuron 2014, 82, 737–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björnsdotter, M.; Gordon, I.; Pelphrey, K.A.; Olausson, H.; Kaiser, M.D. Development of brain mechanisms for processing af-fective touch. Front. Behav. Neurosci. 2014, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Gordon, I.; Voos, A.C.; Bennett, R.H.; Bolling, D.Z.; Pelphrey, K.A.; Kaiser, M.D. Brain mechanisms for processing affective touch. Hum. Brain Mapp. 2013, 34, 914–922. [Google Scholar] [CrossRef]
- Olausson, H.; Lamarre, Y.; Backlund, H.; Morin, C.; Wallin, B.G.; Starck, G.; Ekholm, S.; Strigo, I.; Worsley, K.; Vallbo, Å.B.; et al. Unmyelinated tactile afferents signal touch and project to insular cortex. Nat. Neurosci. 2002, 5, 900–904. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, R.; Wasling, H.B.; Liljencrantz, J.; Olausson, H.; Johnson, R.D.; Wessberg, J. Human C-Tactile Afferents Are Tuned to the Temperature of a Skin-Stroking Caress. J. Neurosci. 2014, 34, 2879–2883. [Google Scholar] [CrossRef] [PubMed]
- Vallbo, Å.B.; Olausson, H.; Wessberg, J. Unmyelinated Afferents Constitute a Second System Coding Tactile Stimuli of the Human Hairy Skin. J. Neurophysiol. 1999, 81, 2753–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, I.; Loken, L.; Olausson, H. The Skin as a Social Organ. Exp. Brain Res. 2010, 204, 305–314. [Google Scholar] [CrossRef]
- Löken, L.S.; Wessberg, J.; Morrison, I.; McGlone, F.; Olausson, H. Coding of pleasant touch by unmyelinated afferents in humans. Nat. Neurosci. 2009, 12, 547–548. [Google Scholar] [CrossRef] [PubMed]
- Pawling, R.; Cannon, P.R.; McGlone, F.P.; Walker, S.C. C-tactile afferent stimulating touch carries a positive affective value. PLoS ONE 2017, 12, e0173457. [Google Scholar] [CrossRef] [Green Version]
- Craig, A.D. Interoception: The sense of the physiological condition of the body. Curr. Opin. Neurobiol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Van Puyvelde, M.; Gorissen, A.-S.; Pattyn, N.; McGlone, F. Does touch matter? The impact of stroking versus non-stroking maternal touch on cardio-respiratory processes in mothers and infants. Physiol. Behav. 2019, 207, 55–63. [Google Scholar] [CrossRef]
- Bernhardt, J. Sensory capacities of the fetus. MCN Am. J. Matern. Child Nurs. 1987, 12, 44–47. [Google Scholar] [CrossRef]
- Kisilevsky, B.S.; Muir, D.W.; Low, J.A. Maturation of Human Fetal Responses to Vibroacoustic Stimulation. Child Dev. 1992, 63, 1497–1508. [Google Scholar] [CrossRef]
- Bystrova, K. Novel mechanism of human fetal growth regulation: A potential role of lanugo, vernix caseosa and a second tac-tile system of unmyelinated low-threshold C-afferents. Med. Hypotheses 2009, 72, 143–146. [Google Scholar] [CrossRef]
- Porter, R.H. The biological significance of skin-to-skin contact and maternal odours. Acta Paediatr. 2004, 93, 1560–1562. [Google Scholar] [CrossRef] [PubMed]
- Peláez-Nogueras, M.; Gewirtz, J.L.; Field, T.; Cigales, M.; Malphurs, J.; Clasky, S.; Sanchez, A. Infants’ preference for touch stimulation in face-to-face interactions. J. Appl. Dev. Psychol. 1996, 17, 199–213. [Google Scholar] [CrossRef]
- Della Longa, L.; Gliga, T.; Farroni, T. Tune to touch: Affective touch enhances learning of face identity in 4-month-old infants. Dev. Cogn. Neurosci. 2019, 35. [Google Scholar] [CrossRef] [PubMed]
- Marx, V.; Nagy, E. Fetal Behavioural Responses to Maternal Voice and Touch. PLoS ONE 2015, 10, e0129118. [Google Scholar] [CrossRef] [Green Version]
- Crucianelli, L.; Filippetti, M.L. Developmental Perspectives on Interpersonal Affective Touch. Topoi 2020, 39, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, V.; von Rosenberg, W.; Montaldo, P.; Adjei, T.; Mendoza, J.; Shivamurthappa, V.; Mandic, D.; Thayyil, S. Early Post-natal Heart Rate Variability in Healthy Newborn Infants. Front. Physiol. 2019, 10, 922. [Google Scholar] [CrossRef] [Green Version]
- Apgar, V. A proposal for a new method of evaluation of the newborn infant. Curr. Res. Anesth. Analg. 1953, 32, 260–267. [Google Scholar] [CrossRef]
- Chong, D.S.; Karlberg, J. Refining the Apgar score cut-off point for newborns at risk. Acta Paediatr. 2004, 93, 53–59. [Google Scholar] [CrossRef]
- Baldzer, K.; Dykes, F.D.; Jones, S.A.; Brogan, M.; Carrigan, T.A.; Giddens, D.P. Heart rate variability analysis in full-term in-fants: Spectral indices for study of neonatal cardiorespiratory control. Pediatr. Res. 1989, 26, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longin, E.; Schaible, T.; Lenz, T.; König, S. Short term heart rate variability in healthy neonates: Normative data and physiological observations. Early Hum. Dev. 2005, 81, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.L.; van Roon, A.; Riese, H.; Thio, C.; Oostenbroek, E.; Westrik, I.; de Geus, E.J.C.; Gansevoort, R.; Lefrandt, J.; Nolte, I.M.; et al. Validity of (ultra-)short recordings for heart rate variability measurements. PLoS ONE 2015, 10, e0138921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordin, M. Low-threshold mechanoreceptive and nociceptive units with unmyelinated (C) fibres in the human supraorbital nerve. J. Physiol. 1990, 426, 229–240. [Google Scholar] [CrossRef]
- Kress, I.U.; Minati, L.; Ferraro, S.; Critchley, H.D. Direct skin-to-skin versus indirect touch modulates neural responses to stroking versus tapping. Neuroreport 2011, 22, 646–651. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.0.0. 2009. Available online: http://r-forge.r-project.org/projects/mumin/ (accessed on 24 February 2021).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: New York, NY, USA, 2007. [Google Scholar]
- Baayen, R.H.; Davidson, D.J.; Bates, D.M. Mixed-effects modeling with crossed random effects for subjects and items. J. Mem. Lang. 2008, 59, 390–412. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Proceedings of the Second International Symposium On Information Theory; Petrov, B.N., Caski, F., Eds.; Akademiai Kiado: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- McElreath, R. Statistical Rethinking: A Bayesian Course with Examples in R and Stan; CRC Press: Boca Raton, FL, USA, 2016; Volume 122. [Google Scholar]
- Hooper, D.; Coughlan, J.; Mullen, M. Structural equation modelling: Guidelines for determining model fit. Electron. J. Bus. Res. Methods 2008, 6, 53–60. [Google Scholar]
- Bazhenova, O.V.; Plonskaia, O.; Porges, S.W. Vagal reactivity and affective adjustment in infants during interaction challen-ges. Child Dev. 2001, 72, 1314–1326. [Google Scholar] [CrossRef]
- Field, T.; Hernandez-Reif, M.; Diego, M.; Feijo, L.; Vera, Y.; Gil, K. Massage therapy by parents improves early growth and development. Infant Behav. Dev. 2004, 27, 435–442. [Google Scholar] [CrossRef]
- Takatani, T.; Takahashi, Y.; Yoshida, R.; Imai, R.; Uchiike, T.; Yamazaki, M.; Shima, M.; Nishikubo, T.; Ikada, Y.; Fujimoto, S. Relationship between frequency spectrum of heart rate variability and autonomic nervous activities during sleep in newborns. Brain Dev. 2018, 40, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.J. Measurement of Heart Rate Variability. West. J. Nurs. Res. 1995, 17, 32–48. [Google Scholar] [CrossRef]
- Birns, B.; Blank, M.; Bridger, W.H. The effectiveness of various soothing techniques on human neonates. Psychosom. Med. 1966, 28, 316–322. [Google Scholar] [CrossRef]
- Bloch-Salisbury, E.; Zuzarte, I.; Indic, P.; Bednarek, F.; Paydarfar, D. Kangaroo care: Cardio-respiratory relationships between the infant and caregiver. Early Hum. Dev. 2014, 90, 843–850. [Google Scholar] [CrossRef]
- Van Puyvelde, M.; Loots, G.; Meys, J.; Neyt, X.; Mairesse, O.; Simcock, D.; Pattyn, N. Whose clock makes yours tick? How maternal cardiorespiratory physiology influences newborns’ heart rate variability. Biol. Psychol. 2015, 108, 132–141. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, J.A.; Costigan, K.A.; Pressman, E.K.; Doussard-Roosevelt, J.A. Antenatal origins of individual differences in heart rate. Dev. Psychobiol. 2000, 37, 221–228. [Google Scholar] [CrossRef]
- Nair, M.N.G.; Gupta, G.; Jatana, S.K. NICU Environment: Can we be Ignorant? Med. J. Armed Forces India 2003, 59, 93–95. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, D.N.; Miller, L.J.; Shyu, V.; Hagerman, R.J. Sensory-modulation disruption, electrodermal responses, and functional behaviors. Dev. Med. Child Neurol. 1999, 41, 608–615. [Google Scholar] [CrossRef]
- Schaaf, R.; Benevides, T.; Blanche, E.; Brett-Green, B.; Burke, J.; Cohn, E.; Koomar, J.; Lane, S.; Miller, L.; May-Benson, T.; et al. Parasympathetic functions in children with sensory processing disorder. Front. Integr. Neurosci. 2010, 4, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
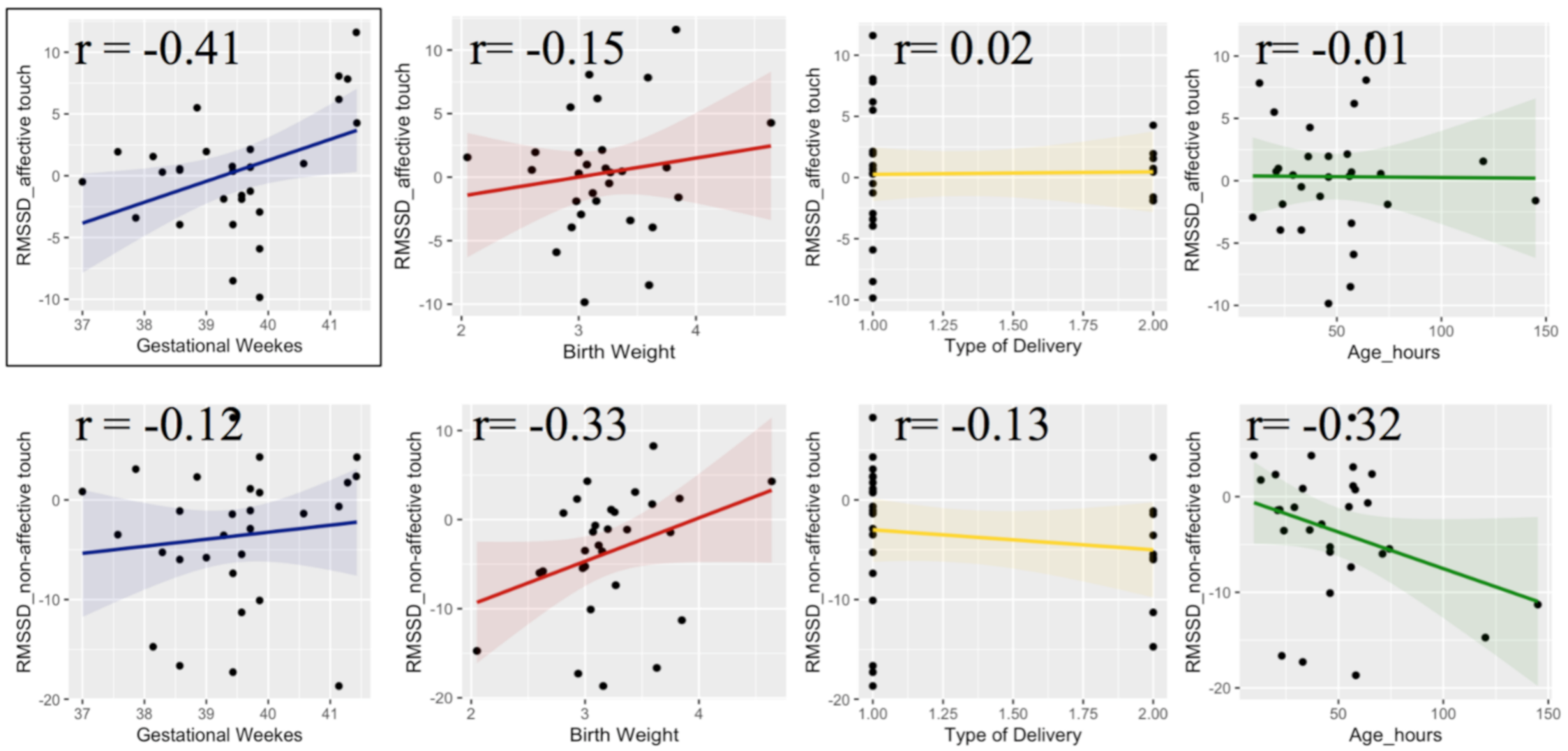{kind=link}
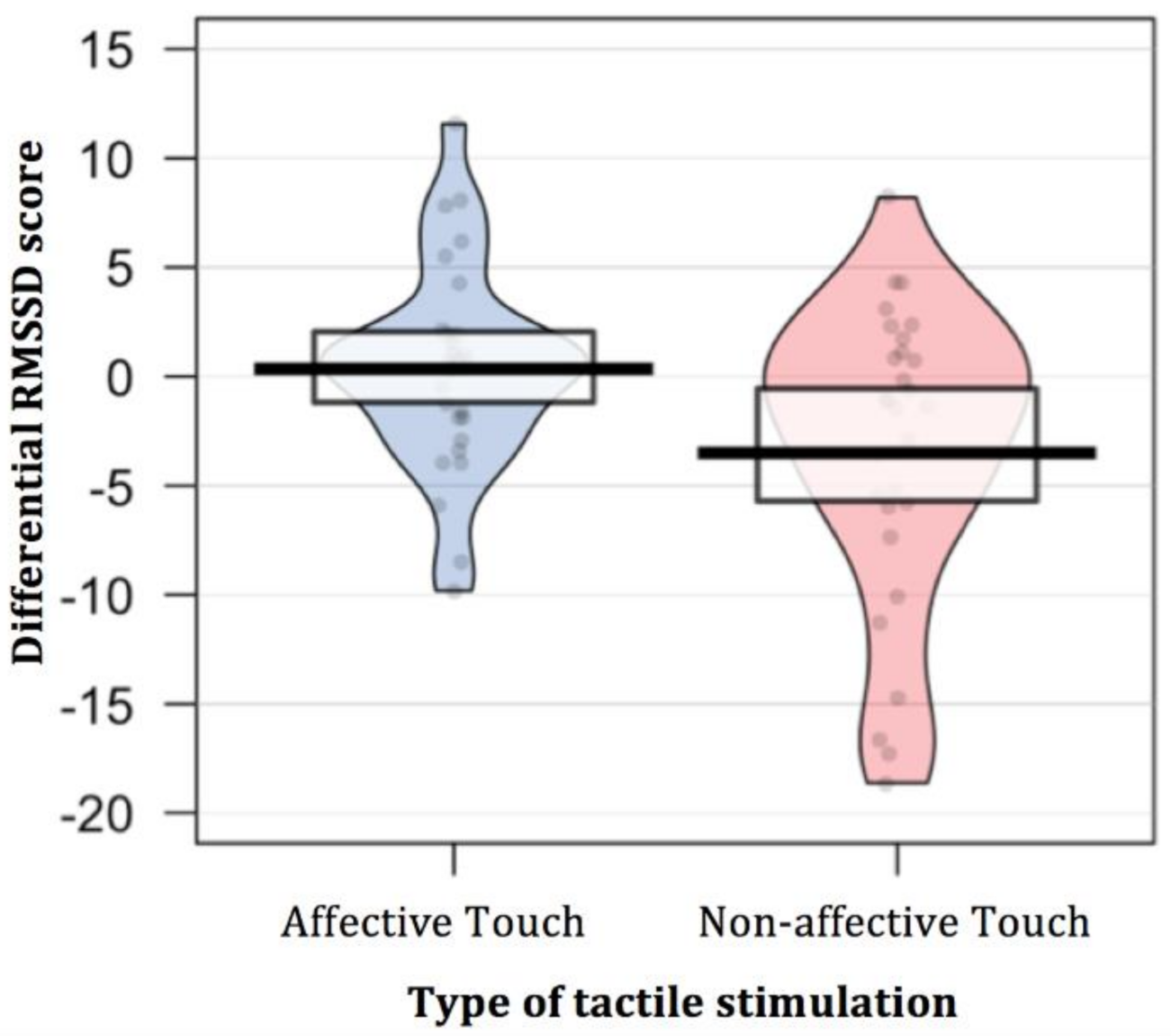
| Type of Touch | Differential RMSSD Score | Simple t-Test (Null Level) | Paired t-Test |
|---|---|---|---|
| Affective Touch | 0.35 (4.66) | t (29) = 0.41; p = 0.682 Cohen’s d = 0.07 | t (29) = 2.71; p = 0.011 Cohen’s d = 0.66 |
| Non-affective Touch | −3.51 (6.83) | t (29) = −2.81; p = 0.009 Cohen’s d = −0.51 |
| Type of Touch | Differential RMSSD Score | Simple t-Test (Null Level) | Paired t-Test |
|---|---|---|---|
| Affective Touch | 1.90 (9.19) | t (29) = 1.13; p = 0.267 Cohen’s d = 0.21 | t (29) = 2.04; p = 0.051 Cohen’s d = 0.62 |
| Non-affective Touch | −3.47 (8.03) | t (29) = −2.37; p = 0.025 Cohen’s d = −0.43 |
| Tested Models | Variables | AIC | Delta AIC | Marginal R2 | χ2 | p |
|---|---|---|---|---|---|---|
| Model 0 | Random effect of Participants | 780.84 | ||||
| Model 1 | + Presence of touch | 779.49 | 2.88 | 0.011 | 3.349 | 0.067 |
| Model 2 | + Type of touch | 779.15 | 1.83 | 0.017 | 2.337 | 0.126 |
| Model 3 | + Interaction Presence * Type of touch | 775.76 | 6.13 | 0.033 | 5.393 | 0.020 |
| Model 4 | + Order of tactile stimulation | 777.06 | 0.15 | 0.035 | 0.704 | 0.401 |
| Model 5 | + Interaction with Order | 780.41 | 8.58 | 0.041 | 2.651 | 0.449 |
| Variables | B (SE) | T | p |
|---|---|---|---|
| Presence of touch | 0.352 (1.178) | 0.299 | 0.766 |
| Type of touch | 3.229 (1.178) | 2.741 | 0.007 |
| Presence of touch × Type of touch | −3.861 (1.666) | −2.318 | 0.023 |
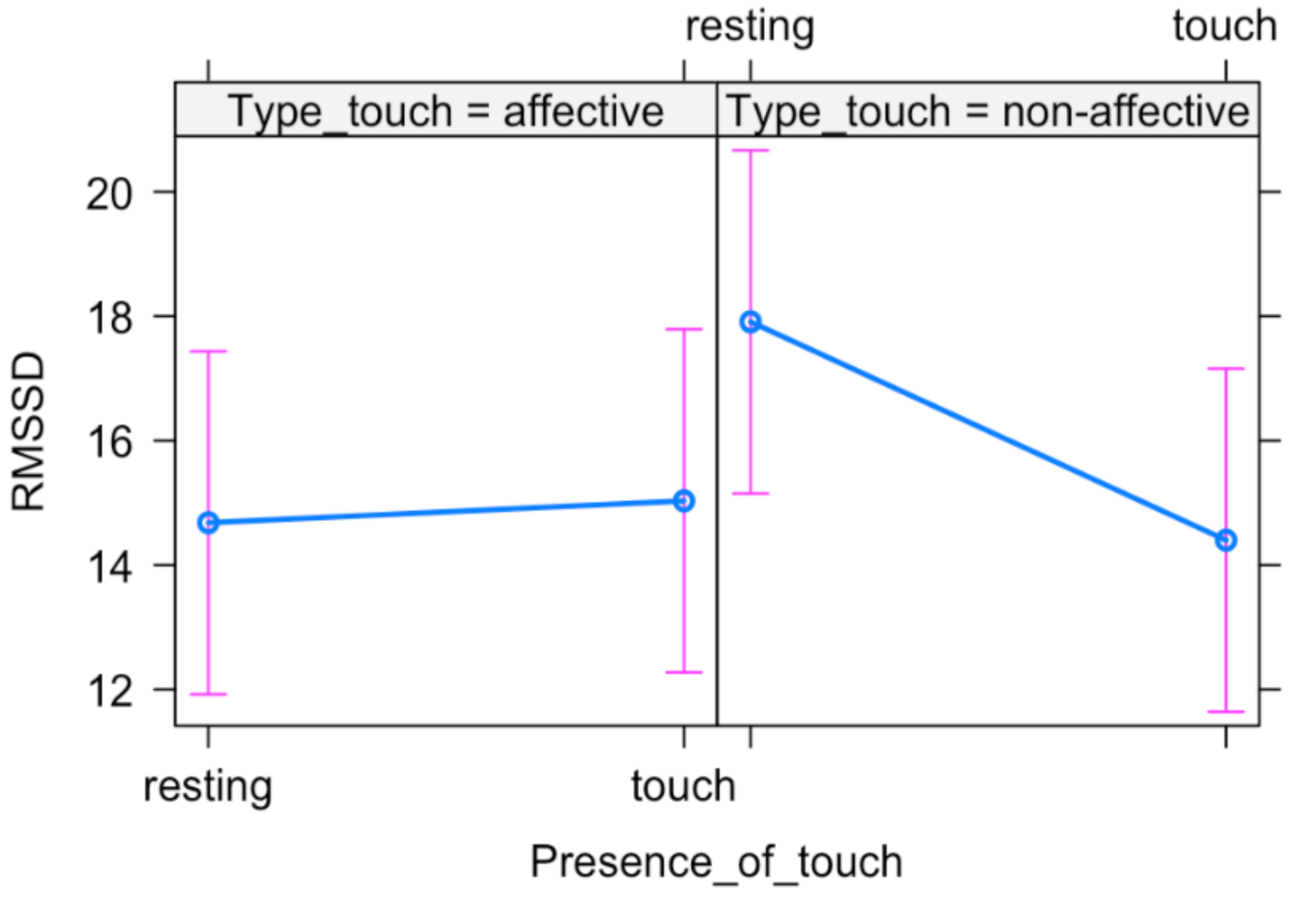
| Tested Models | Variables | AIC | Delta AIC | Marginal R2 | χ2 | p |
|---|---|---|---|---|---|---|
| Model 0 | Random effect of Participants | 1173 | ||||
| Model 1 | + Time | 1173 | 1.99 | 0.006 | 2.935 | 0.231 |
| Model 2 | + Type of touch | 1175 | −0.68 | 0.006 | 0.042 | 0.838 |
| Model 3 | + Interaction Time × Type of touch | 1170 | 10.95 | 0.024 | 9.400 | 0.009 |
| Model 4 | + Order of tactile stimulation | 1171 | 0.89 | 0.028 | 1.722 | 0.190 |
| Model 5 | + Interaction with Order | 1176 | 13.86 | 0.036 | 4.759 | 0.446 |
| Variables | B (SE) | T | p |
|---|---|---|---|
| Time (PreTouch–Touch) | 0.29 (1.24) | 0.23 | 0.814 |
| Time (PreTouch–PostTouch) | 1.89 (1.24) | 1.528 | 0.128 |
| Type of touch Time (PreTouch–Touch) × (NonAffective) Time (PreTouch–PostTouch) × (NonAffective) | 3.28 (1.24) −3.75 (1.75) −5.40 (1.75) | 2.649 −2.142 −3.084 | 0.009 0.034 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Della Longa, L.; Dragovic, D.; Farroni, T. In Touch with the Heartbeat: Newborns’ Cardiac Sensitivity to Affective and Non-Affective Touch. Int. J. Environ. Res. Public Health 2021, 18, 2212. https://doi.org/10.3390/ijerph18052212
Della Longa L, Dragovic D, Farroni T. In Touch with the Heartbeat: Newborns’ Cardiac Sensitivity to Affective and Non-Affective Touch. International Journal of Environmental Research and Public Health. 2021; 18(5):2212. https://doi.org/10.3390/ijerph18052212
Chicago/Turabian StyleDella Longa, Letizia, Danica Dragovic, and Teresa Farroni. 2021. "In Touch with the Heartbeat: Newborns’ Cardiac Sensitivity to Affective and Non-Affective Touch" International Journal of Environmental Research and Public Health 18, no. 5: 2212. https://doi.org/10.3390/ijerph18052212