The Effects of Pro-Inflammatory and Anti-Inflammatory Agents for the Suppression of Intimal Hyperplasia: An Evidence-Based Review

 ,
,
Abstract
:1. Introduction
1.1. Vascular Intimal Hyperplasia, Risk Factors and Current Therapy
1.2. Chemokines and/or Cytokines Effects in Inhibiting Intimal Hyperplasia Development
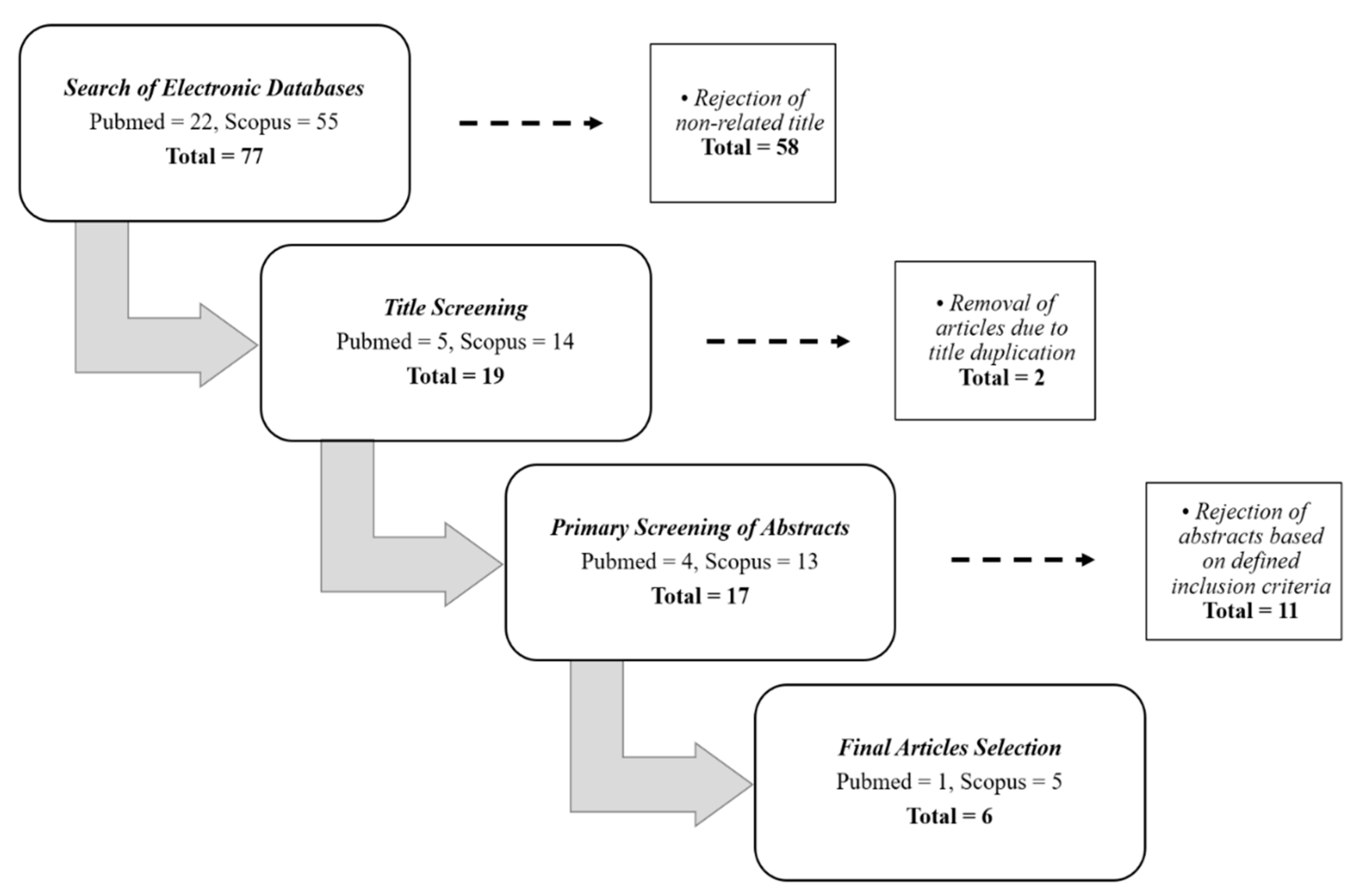
2. Methods
2.1. Search Strategy in Selected Databases
2.2. Criteria of Selection
2.3. Management of Data Extraction Table
3. Results
3.1. Search Results
3.2. Study Characteristics
4. Discussion
4.1. Utilisation of Natural Product to Combat IH
4.2. Genetic Manipulation and Protein Inhibition to Attenuate the Progression of IH
4.3. Targeted Drugs in Clinical Setting to Suppress IH
4.4. Emerging Role of Chemokines and Cytokines Induction to Combat IH/Cardiovascular Disease
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Clark, H. NCDs: A challenge to sustainable human development. Lancet 2013, 381, 510–511. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.; Han, S.; Rane, P.P.; Fox, K.M.; Qian, Y.; Suh, H.S. Prevalence and incidence of atherosclerotic cardiovascular disease and its risk factors in Korea: A nationwide population-based study. BMC Public Health 2019, 19, 1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhang, S.; Wu, J.; Wu, S.; Xu, G.; Wei, D. Essential role of nonessential amino acid glutamine in atherosclerotic cardiovascular disease. DNA Cell Biol. 2020, 39, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shioi, A.; Ikari, Y. Plaque calcification during atherosclerosis progression and regression. J. Atheroscler. Thromb. 2018, 25, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Chen, M.; Li, Y.; Wong, F.H.; Thiam, C.W.; Hossain, M.Z.; Poh, K.K.; Hirohata, S.; Ogawa, H.; Angeli, V.; et al. Loss of ADAMTS4 reduces high fat diet-induced atherosclerosis and enhances plaque stability in ApoE-/- mice. Sci. Rep. 2016, 6, 31130. [Google Scholar] [CrossRef] [Green Version]
- Rajamani, K.; Fisher, M. An overview of atherosclerosis. Prim. Cerebrovasc. Dis. Second Ed. 2017, 105–108. [Google Scholar] [CrossRef]
- Warboys, C.M.; Amini, N.; De Luca, A.; Evans, P.C. The role of blood flow in determining the sites of atherosclerotic plaques. F1000 Med. Rep. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Muscogiuri, G.; Annweiler, C.; Duval, G.; Karras, S.; Tirabassi, G.; Salvio, G.; Balercia, G.; Kimball, S.; Kotsa, K.; Mascitelli, L.; et al. Vitamin D and cardiovascular disease: From atherosclerosis to myocardial infarction and stroke. Int. J. Cardiol. 2017, 230, 577–584. [Google Scholar] [CrossRef]
- Best, P.J.M.; Lerman, A. Endothelin in cardiovascular disease: From atherosclerosis to heart failure. J. Cardiovasc. Pharmacol. 2000, 35, S61–S63. [Google Scholar] [CrossRef]
- Ross, R.; Agius, L. The process of atherogenesis—Cellular and molecular interaction: From experimental animal models to humans. Diabetologia 1992, 35, S34–S40. [Google Scholar] [CrossRef]
- Stary, H.C.; Chandler, A.B.; Glagov, S.; Guyton, J.R.; Insull, W., Jr.; Rosenfeld, M.E.; Schaffer, S.A.; Schwartz, C.J.; Wagner, W.D.; Wissler, R.W. A definition of initial, fatty streak, and intermediate lesions of atherosclerosis. Circulation 1994, 89, 2462–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, P.-K.; Tran-Lundmark, K.; Soininen, R.; Tryggvason, K.; Thyberg, J.; Hedin, U. Increased intimal hyperplasia and smooth muscle cell proliferation in transgenic mice with heparan sulfate–deficient perlecan. Circ. Res. 2004, 94, 550–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigel, B.; Swami, V.; Can, A.; Parsons, R.E.; Golub, R.M.; Kolecki, R.; Kitamura, H. Intimal hyperplasia producing thrombus organization in an experimental venous thrombosis model. J. Vasc. Surg. 1994, 19, 350–360. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.K.; Agrawal, T.; Rai, V.; Del Core, M.G.; Hunter, W.J.; Agrawal, D.K. Vitamin D supplementation reduces intimal hyperplasia and restenosis following coronary intervention in atherosclerotic swine. PLoS ONE 2016, 11, e0156857. [Google Scholar] [CrossRef] [PubMed]
- Casscells, W. Migration of smooth muscle and endothelial cells: Critical events in restenosis. Circulation 1992, 86, 723–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, S.F.; Zahradka, P. Vascular smooth muscle cell motility: From migration to invasion. Exp. Clin. Cardiol. 2010, 15, e75–e85. [Google Scholar] [PubMed]
- Jennette, J.C.; Stone, J.R. Diseases of medium-sized and small vessels. In Cellular and Molecular Pathobiology of Cardiovascular Disease; Elsevier: Amsterdam, The Netherlands, 2014; pp. 197–219. [Google Scholar] [CrossRef]
- Ducasse, E.; Cosset, J.M.; Eschwège, F.; Chevalier, J.; De Ravignan, D.; Puppinck, P.; Lartigau, E. Hyperplasia of the arterial intima due to smooth muscle cell proliferation. Current data, experimental treatments and perspectives. J. Mal. Vasc. 2003, 28, 130–144. [Google Scholar] [PubMed]
- Zalewski, A.; Shi, Y.; Johnson, A.G. Diverse origin of intimal cells. Circ. Res. 2002, 91, 652–655. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Waltham, M.; Wadoodi, A.; Burnand, K.; Smith, A. The role of endothelial cells and their progenitors in intimal hyperplasia. Ther. Adv. Cardiovasc. Dis. 2010, 4, 129–141. [Google Scholar] [CrossRef]
- Tsai, S.; Butler, J.; Rafii, S.; Liu, B.; Kent, K.C. The role of progenitor cells in the development of intimal hyperplasia. J. Vasc. Surg. 2009, 49, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Al-Ani, M.K.; Pan, X.; Chi, Q.; Dong, N.; Qiu, X. Plant-derived products for treatment of vascular intima hyperplasia selectively inhibit vascular smooth muscle cell functions. Evidence-Based Complement. Altern. Med. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lemson, M.; Tordoir, J.; Daemen, M.; Kitslaar, P. Intimal hyperplasia in vascular grafts. Eur. J. Vasc. Endovasc. Surg. 2000, 19, 336–350. [Google Scholar] [CrossRef] [Green Version]
- Zubilewicz, T.; Wronski, J.; Bourriez, A.; Terlecki, P.; Guinault, A.M.; Muscatelli-Groux, B.; Michalak, J.; Méllière, D.; Becquemin, J.; Allaire, E. Injury in vascular surgery-the intimal hyperplastic response. Med. Sci. Monit. 2001, 7, 316–324. [Google Scholar]
- Gellman, J.; Ezekowitz, M.D.; Sarembock, I.J.; Azrin, M.A.; Nochomowitz, L.E.; Lerner, E.; Haudenschild, C.C. Effect of lovastatin on intimal hyperplasia after balloon angioplasty: A study in an atherosclerotic hypercholesterolemic rabbit. J. Am. Coll. Cardiol. 1991, 17, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Carrozza, J.P.; Kuntz, R.E.; Fishman, R.F.; Baim, D.S. Restenosis after arterial injury caused by coronary stenting in patients with diabetes mellitus. Ann. Intern. Med. 1993, 118, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Salam, T.A.; Taylor, B.; Suggs, W.D.; Hanson, S.R.; Lumsden, A.B. Reaction to injury following balloon angioplasty and intravascular stent placement in the canine femoral artery. Am. Surg. 1994, 60, 353–357. [Google Scholar] [PubMed]
- Kornowski, R.; Hong, M.K.; Tio, F.O.; Bramwell, O.; Wu, H.; Leon, M.B. In-stent restenosis: Contributions of inflammatory responses and arterial injury to neointimal hyperplasia. J. Am. Coll. Cardiol. 1998, 31, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Lacolley, P.; Regnault, V.; Avolio, A.P. Smooth muscle cell and arterial aging: Basic and clinical aspects. Cardiovasc. Res. 2018, 114, 513–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakoshi, A.; Hatano, T.; Tsukahara, T.; Murakami, M.; Arai, D.; Yamaguchi, S. Percutaneous transluminal angioplasty for atherosclerotic stenosis of the subclavian or innominate artery: Angiographic and clinical outcomes in 36 patients. Neurosurg. Rev. 2011, 35, 121–126. [Google Scholar] [CrossRef]
- Cooper, C.J.; Murphy, T.P.; Cutlip, D.E.; Jamerson, K.; Henrich, W.; Reid, D.M.; Cohen, D.J.; Matsumoto, A.H.; Steffes, M.; Jaff, M.R.; et al. Stenting and medical therapy for atherosclerotic renal-artery stenosis. N. Engl. J. Med. 2014, 370, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Higo, T.; Ueda, Y.; Oyabu, J.; Okada, K.; Nishio, M.; Hirata, A.; Kashiwase, K.; Ogasawara, N.; Hirotani, S.; Kodama, K. Atherosclerotic and thrombogenic neointima formed over sirolimus drug-eluting stent. JACC Cardiovasc. Imaging 2009, 2, 616–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackensen, G.; Swaminathan, M.; Ti, L.K.; Grocott, H.P.; Phillips-Bute, B.G.; Mathew, J.P.; Newman, M.F.; Milano, C.A.; Stafford-Smith, M. Preliminary report on the interaction of apolipoprotein E polymorphism with aortic atherosclerosis and acute nephropathy after CABG. Ann. Thorac. Surg. 2004, 78, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Osgood, M.J.; Hocking, K.M.; Voskresensky, I.V.; Li, F.D.; Komalavilas, P.; Cheung-Flynn, J.; Brophy, C.M. Surgical vein graft preparation promotes cellular dysfunction, oxidative stress, and intimal hyperplasia in human saphenous vein. J. Vasc. Surg. 2013, 60, 202–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellings, W.E.; Moll, F.L.; De Vries, J.P.P.; Ackerstaff, R.G.; Seldenrijk, K.A.; Met, R.; Velema, E.; Derksen, W.J.; De Kleijn, D.P.; Pasterkamp, G. Atherosclerotic plaque composition and occurrence of restenosis after carotid endarterectomy. JAMA J. Am. Med. Assoc. 2008, 299, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.W.; Roubin, G.S.; King, S.B. Restenosis after coronary angioplasty. Potential biologic determinants and role of intimal hyperplasia. Circulation 1989, 79, 1374–1387. [Google Scholar] [CrossRef] [Green Version]
- Rosenfield, K.; Schainfeld, R.; Pieczek, A.; Haley, L.; Isner, J.M. Restenosis of endovascular stents from stent compression. J. Am. Coll. Cardiol. 1997, 29, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Charo, I.F.; Taubman, M.B. Chemokines in the pathogenesis of vascular disease. Circ. Res. 2004, 95, 858–866. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Tseng, S.C.; Rollins, B.J. Monocyte chemoattractant protein-1. Chem. Immunol. 1999, 72, 7–29. [Google Scholar] [CrossRef]
- Roque, M.; Kim, W.J.; Gazdoin, M.; Malik, A.; Reis, E.D.; Fallon, J.T.; Badimon, J.J.; Charo, I.F.; Taubman, M.B. CCR2 deficiency decreases intimal hyperplasia after arterial injury. Arter. Thromb. Vasc. Biol. 2002, 22, 554–559. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, Y.; Matsumori, A.; Ohashi, N.; Shioi, T.; Ono, K.; Harada, A.; Matsushima, K.; Sasayama, S. Anti-monocyte chemoattractant protein-1/monocyte chemotactic and activating factor antibody inhibits neointimal hyperplasia in injured rat carotid arteries. Circ. Res. 1999, 84, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Idriss, H.T.; Naismith, J.H. TNFα and the TNF receptor superfamily: Structure-function relationship(s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- Shaw, J.; Tang, Z.; Schneider, H.; Saljé, K.; Hansson, S.R.; Guller, S. Inflammatory processes are specifically enhanced in endothelial cells by placental-derived TNF-α: Implications in preeclampsia (PE). Placenta 2016, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Brand, K.; Page, S.; Rogler, G.; Barstch, A.; Brandl, R.; Knuechel, R.; Page, M.; Kaltschmidt, C.; Baeuerle, P.A.; Neumeier, D. Activated transcription factor nuclear factor-kappa B is present in the atherosclerotic lesion. J. Clin. Investig. 1996, 97. [Google Scholar] [CrossRef] [PubMed]
- Maleknia, M.; Ansari, N.; Haybar, H.; Maniati, M.; Saki, N. Inflammatory growth factors and in-stent restenosis: Effect of cytokines and growth factors. SN Compr. Clin. Med. 2020, 2, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interf. Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Li, B.; Pozzi, A.; Young, P.P. TNFα accelerates monocyte to endothelial transdifferentiation in tumors by the induction of integrin α5 expression and adhesion to fibronectin. Mol. Cancer Res. 2011, 9, 702–711. [Google Scholar] [CrossRef] [Green Version]
- Ko, I.K.; Lee, S.J.; Atala, A.; Yoo, J.J. In situ tissue regeneration through host stem cell recruitment. Exp. Mol. Med. 2013, 45, e57. [Google Scholar] [CrossRef]
- Choi, S.; Park, M.; Kim, J.; Park, W.; Kim, S.; Lee, D.-K.; Hwang, J.Y.; Choe, J.; Won, M.-H.; Ryoo, S.; et al. TNF-α elicits phenotypic and functional alterations of vascular smooth muscle cells by miR-155-5p–dependent down-regulation of cGMP-dependent kinase 1. J. Biol. Chem. 2018, 293, 14812–14822. [Google Scholar] [CrossRef] [Green Version]
- Fardoun, M.; Iratni, R.; Dehaini, H.; Eeid, A.; Ghaddar, T.; El-Elimat, T.; Alali, F.; Badran, A.; Eid, A.H.; Baydoun, E.; et al. 7-O-methylpunctatin, a novel homoisoflavonoid, inhibits phenotypic switch of human arteriolar smooth muscle cells. Biomolecules 2019, 9, 716. [Google Scholar] [CrossRef] [Green Version]
- Kenagy, R.D.; Civelek, M.; Kikuchi, S.; Chen, L.; Grieff, A.; Sobel, M.; Lusis, A.J.; Clowes, A.W. Scavenger receptor class A member 5 (SCARA5) and suprabasin (SBSN) are hub genes of coexpression network modules associated with peripheral vein graft patency. J. Vasc. Surg. 2016, 64, 202–209.e6. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Zhu, X.; Cai, W.; Qiu, L.Y. Hypaphorine attenuates lipopolysaccharide-induced endothelial inflammation via regulation of TLR4 and PPAR-γ dependent on PI3K/Akt/mTOR signal pathway. Int. J. Mol. Sci. 2017, 18, 844. [Google Scholar] [CrossRef] [Green Version]
- De Jong, R.C.M.; Ewing, M.M.; De Vries, M.R.; Karper, J.C.; Bastiaansen, A.J.N.M.; Peters, H.A.B.; Baghana, F.; Elsen, P.J.V.D.; Gongora, C.; Jukema, J.W.; et al. The epigenetic factor PCAF regulates vascular inflammation and is essential for intimal hyperplasia development. PLoS ONE 2017, 12, e0185820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Chai, S.; Dai, L.; Gu, C. Collagen external scaffolds mitigate intimal hyperplasia and improve remodeling of vein grafts in a rabbit arteriovenous graft model. Biomed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.L.; Lin, Y.C.; Ni, H.; Mo, F.E. Ganoderma triterpenoids exert antiatherogenic effects in mice by alleviating disturbed flow-induced oxidative stress and inflammation. Oxidative Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Pang, D.; Yuan, T.; Li, Z.; Li, Z.; Zhang, M.; Ren, W.; Ouyang, H.; Tang, X. N-3 polyunsaturated fatty acids attenuates triglyceride and inflammatory factors level in hfat-1 transgenic pigs. Lipids Health Dis. 2016, 15, 89. [Google Scholar] [CrossRef] [Green Version]
- Sanders, W.G.; Li, H.; Zhuplatov, I.; He, Y.; Kim, S.E.; Cheung, A.K.; Agarwal, J.; Terry, C.M. Autologous fat transplants to deliver glitazone and adiponectin for vasculoprotection. J. Control. Release 2017, 264, 237–246. [Google Scholar] [CrossRef]
- Schlosser, A.; Pilecki, B.; Hemstra, L.E.; Kejling, K.; Kristmannsdottir, G.B.; Wulf-Johansson, H.; Moeller, J.B.; Füchtbauer, E.M.; Nielsen, O.; Kirketerp-Møller, K.; et al. MFAP4 promotes vascular smooth muscle migration, proliferation and accelerates neointima formation. Arterioscler. Thromb. Vasc. Biol. 2016, 36. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Deng, P.; Sun, Y.; Ye, P.; Zhang, A.; Wu, C.; Yue, Z.; Chen, Z.; Xia, J. MicroRNA-155 promotes neointimal hyperplasia through smooth muscle-like cell-derived RANTES in arteriovenous fistulas. J. Vasc. Surg. 2018, 67. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Brahmbhatt, A.; Torres, E.N.; Thielen, B.; McCall, D.L.; Engel, S.; Bansal, A.; Pandey, M.K.; Dietz, A.B.; Leof, E.B.; et al. Tracking and therapeutic value of human adipose tissue-derived mesenchymal stem cell transplantation in reducing venous neointimal hyperplasia associated with arteriovenous fistula. Radiology 2016, 279. [Google Scholar] [CrossRef] [Green Version]
- Moreira, D.M.; da Silva, R.L.; Vieira, J.L.; Fattah, T.; Lueneberg, M.E.; Gottschall, C.A.M. Role of vascular inflammation in coronary artery disease: Potential of anti-inflammatory drugs in the prevention of atherothrombosis: Inflammation and anti-inflammatory drugs in coronary artery disease. Am. J. Cardiovasc. Drugs 2015, 15, 1–11. [Google Scholar] [CrossRef]
- Christodoulidis, G.; Vittorio, T.J.; Fudim, M.; Lerakis, S.; Kosmas, C.E. Inflammation in coronary artery disease. Cardiol. Rev. 2014, 22, 279–288. [Google Scholar] [CrossRef]
- Kosmas, C.E.; Silverio, D.; Tsomidou, C.; Salcedo, M.D.; Montan, P.D.; Guzman, E. The impact of insulin resistance and chronic kidney disease on inflammation and cardiovascular disease. Clin. Med. Insights Endocrinol. Diabetes 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.L.; Yeh, J.L.; Tsai, P.C.; Chang, T.H.; Huang, W.C.; Lee, S.T.; Wassler, M.; Geng, Y.J.; Sulistyowati, E. Inhibition of neointima hyperplasia, inflammation, and reactive oxygen species in balloon-injured arteries by HVJ envelope vector-mediated delivery of superoxide dismutase gene. Transl. Stroke Res. 2018, 10, 413–427. [Google Scholar] [CrossRef] [Green Version]
- Sotokawauchi, A.; Ishibashi, Y.; Matsui, T.; Yamagishi, S.I. Aqueous extract of glucoraphanin-rich broccoli sprouts inhibits formation of advanced glycation end products and attenuates inflammatory reactions in endothelial cells. Evidence-Based Complement. Altern. Med. 2018, 2018, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galkina, E.V.; Ley, K. Immune and inflammatory mechanisms of atherosclerosis. Ann. Rev. Immunol. 2009, 27, 165–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedgui, A.; Mallat, Z. Cytokines in atherosclerosis: Pathogenic and regulatory pathways. Physiol. Rev. 2006, 86, 515–581. [Google Scholar] [CrossRef] [Green Version]
- Bucova, M.; Bernadic, M.; Buckingham, T. C-reactive protein, cytokines and inflammation in cardiovascular diseases. Bratisl Lek List. 2008, 109, 333–340. [Google Scholar]
- Liu, M.; Chen, J.; Huang, D.; Ke, J.; Wu, W. A meta-analysis of proinflammatory cytokines in chronic heart failure. Hear. Asia 2014, 6, 130–136. [Google Scholar] [CrossRef]
- Kitagaki, M.; Isoda, K.; Kamada, H.; Kobayashi, T.; Tsunoda, S.; Tsutsumi, Y.; Niida, T.; Kujiraoka, T. Inflammatory reactions in the pathogenesis of atherosclerosis inflammation and intimal hyperplasia in Mice. J. Atheroscler. Thromb. 2012, 19, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Wajant, H.; Pfizenmaier, K.; Scheurich, P. Tumor necrosis factor signaling. Cell Death Differ. 2003, 10, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Watanabe, T. Inflammatory reactions in the pathogenesis of atherosclerosis. J. Atheroscler. Thromb. 2003, 10, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| No. | Authors | Type of Cells Used | Type of Cytokines/Chemokines/Extract/Substances/Scaffolds Used | Type of Disease | Methodology | Results | Conclusion |
|---|---|---|---|---|---|---|---|
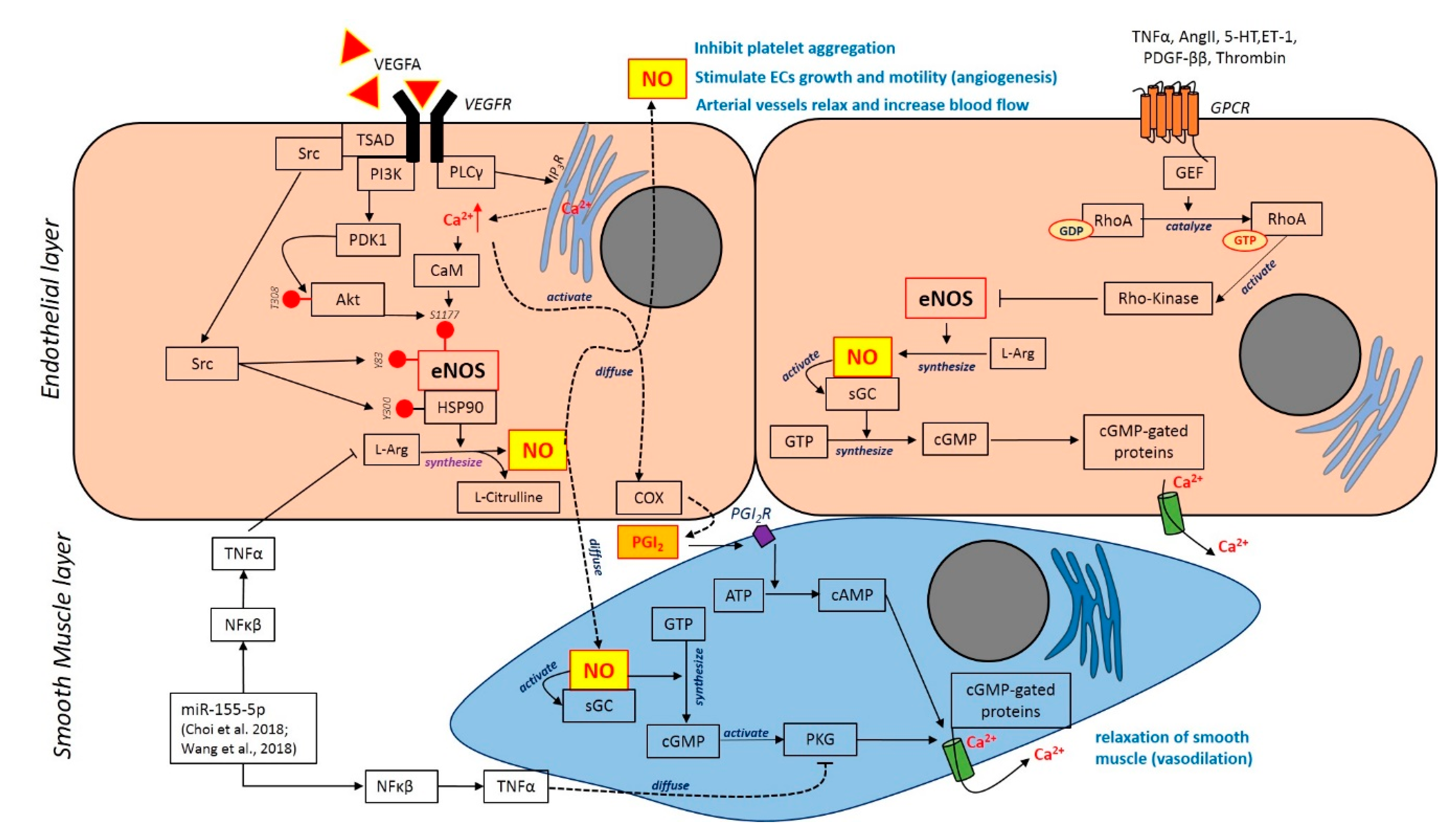
| 1 | Choi et al., 2018 [49] | Vascular smooth muscle cell | TNF-α and IL-1β | Atherosclerosis, hypertension, stroke | VSMC was culture, treated and subjected to reporter gene assay, immunofluorescence, wound healing, proliferation, RTPCR, vascular tension assay and FACs analysis | TNF-alpha down-regulates PKG1 expression PKG1 suppression interceded by TNF-a was controlled by NF-kB NF-kB–responsive miR-155-5p has a direct aim on PKG1 VSMC induced by TNF-a moderated the expression of PKG1 The inhibition of PKG1 imitated the effect of TNF-a in interceding the alteration of VSMC phenotype PKG1 suppression reduced cGMP that moderate the vascular relaxation The stimulation of NF-kB plays a vital role in preventing the vasodilation disorder | The discovery elucidated the explicit role of inflammatory agents such as TNF-a in a pathological process of hypertension and atherosclerosis, which possibly caused by VSMC disorder through certain pathways. Thus, the signalling pathways involved is essential in determining the pathogenesis of other vascular diseases |
| 2 | Fardoun et.al, 2019 [50] | Humanvascular smooth muscle cells | 7-O-Methyl Punctatin (MP) | Cardio-vasculo-pathologies (hypertension) | VSMC were isolated from redundant tissue of a newborn baby. The cells are then treated with MP MP was extracted, characterized, and purified prior to use Cells were subjected to BrdU assay, cell cycle analysis, RTPCR, SEM, wound healing, invasion and cell adhesion assay, monocyte adhesion assay, luciferase reporter assay and western blot | The cell proliferation of VSMC was suppressed by MP VSMC cell cycle was reduced by MP Cyclin D1 and CDK4 expression were attenuated, while CDK inhibitors, p21 and p27 were upregulated in VSMC, induced by MP. VSMC apoptosis were mediated by MP MP decreased cell migration and adhesion in VSMC MP reduced the expression of MMP-2 and MMP-9 in VSMC MP upregulated differentiation markers and downregulated the de-differentiation marker MP downregulated the expression of transcription factor, NF-κB and suppressed the initiation of its inhibitor | These findings showed that MP could be treated as a regulator in cardio-vasculo pathologies, by attenuating the alteration in VSMC phenotype |
| 3 | Kenagy et al., 2016 [51] | Human saphenous vein cell lines | 1. Platelet-derived growth factor (PDGF)-BB 2. Collagen gel | Intimal hyperplasia | Human saphenous vein cell lines used was subjected to gene expression analysis Cells were stimulated with PDGF-BB or serum within 4 h The regulation of gene expression was evaluated by Patterns of Gene Expression software Cell migration was performed using a microchemotaxis chamber to investigate the effects of PDGF-BB Collagen gel contraction, which was used as a model of vascular remodelling, was determined using bovine collagen Bovine collagen gel has been used for vaso-remodelling | Migration in response to PDGF-BB showed no correlation with graft outcome The graft outcome has not correlated with cell migration treated with PDGF-BB Cell migration induced by PDGF-BB was not affected by heparin Cell treated with FBS showed the expression of 1188 unique genes, whereas cell treated with PDGF expressed 1340 genes Sky-blue modules—No significant enrichment in GO terms after treatment with FBS. Yellow modules—No significant enrichment in Go terms after treatment with PDGF Bovine gel contraction—No significant effects was noticed upon SCARA5 knockdown, whereas SBSN knockdown improved the contraction Treatment with PDGF-BB showed that the expression of SCARA5 was escalated, but no significant effect can be seen for SBSN Proliferation assay—No significant effect on SBSN knockdown. SCARA5 knockdown showed slight increment in cell proliferation | Elucidation of SCARA5 and SBSN illustrated that these genes has a great effect for the future therapy of vein graft impairment |
| 4 | Sun et al., 2017 [52] | Human micro-vascular endothelial cells (HMEC-1) | Hypaphorine (Hy) derived marine | Vascular endothelial dysfunction and atherosclerosis | HMEC-1 was cultivated in MCDB 131 medium with 10% FBS Human EA. hy926 endothelial cells were cultivated in DMEM with 10% FBS until confluent The concentration of Hy inhibitors were selected based on optimization that has been reported previously Parameters: RTPCR, western blot, ICC, siRNA transfections, ELISA | TNF-α, IL-1β, VCAM-1 and MCP-1 were upregulated at 500 ng/mL of Hy, and at higher dose (100 µM) attenuates the expression of these genes. TLR4 inhibition affected Hy inflammatory mechanism PPAR-γ interceded in vascular protection of Hy in HMEC-1 PI3K/Akt/mTOR inhibitor plays a significant effect in HMEC-1 upon inflammation, as showed by negative correlation between PPAR-γ and TLR4 | Hy plays a major role in the anti-inflammatory effect of HMEC-1 via the activation of PPAR-γ and TLR4. Thus, it could potentially act as an alternative anti-inflammatory factor for the treatment of inflammatory-related diseases |
| No. | Authors | Type of Cells/Animals Used | Type of Cytokines/Chemokines/Extract/Substances/Scaffolds Used | Type of Disease | Methodology | Results | Conclusion |
|---|---|---|---|---|---|---|---|
| 1 | de Jong et al., 2017 [53] | In vitro: 1. Vascular smooth muscle cells (VSMC) from PCAF KO aorta 2. Bone-marrow derived macrophages from PCAF KO bone marrow 3. Whole blood-derived leukocytes from PCAF KO blood In vivo: PCAF KO transgenic male mice | Garcinol on PCAF KO mice (Genetic P300/CBP-associated factor | Intimal hyperlasia | Chow diet were fed to PCAF KO and WT, whereas ApoE3-Leiden mice were given a Western diet consisted a mixture of cholesterol and cholate to promote hypercholesterolemia in animal model WT and PCAF KO were subjected to vascular inflammation via femoral artery cuff 10 μL pluronic gel F127 ± 25 mg/mL garcinol treatment was given to ApoE3-Leiden mice. Garcinol was applied for few days at the inflammatory site Parameters: Cell viability, IHC, morphometric analysis and ELISA | The lack of PCAF attenuates the production of inflammatory cytokine PCAF KO mice showed the reduction of intimal hyperplasia VSMC, leukocytes and macrophages showed the downregulation in CCL2, IL-6 and TNF-a Macrophage influx and CCL2 regulation were not affected by PCAF deficiency Inflammatory cytokines were inhibited by garcinol treatment PCAF inhibitory effect decreased injury in vivo | The finding defined an essential contribution of lysine acetyltransferase PCAF in regulating inflammatory effect associated to intimal hyperplasia |
| 2 | Haiming et al., 2017 [54] | In vivo: New Zealand white rabbits | Collagen external scaffold (CES) | Intimal hyperplasia | New Zealand white rabbits (n=36) were divided into several groups:
The AVFs was encircled by CES. The parameters of AVFs were evaluated during surgery and ultrasonic evaluation was performed after 4 weeks of surgery, followed by histological analysis of vein grafts. mRNA level and protein expression were quantified using qPCR and Western blot analysis. The markers used are as follows:
| CES successfully prevented the growth of vein grafts by the alteration of diameter Decrease of blood flow in vein graft was impeded effectively by CES Intimal thickening was decreased by CES and significantly ameliorated the restoration of vein graft The expressions of PCNA and ClvCasp-3 were downregulated, and the expression of SM22-alpha was upregulated by CES | CES provided a significant role in reducing the formation of intimal hyperplasia. It also had a useful impact in ameliorating the vein graft remodelling |
| 3 | Hsu et al., 2018 [55] | In vitro: Human umbilical vein endothelial cells In vivo: Carotid artery-ligation mouse model (Male BALB/c) | Ganoderma triterpenoids (GT) extracted from Ganoderma lucidum (mushroom) | Neointimal hyperplasia, atherosclerosis | Mouse model was subjected to GL and GT treatment that promote atherogenesis GL and GT treatment on neointima formation induced mouse (by a ligature at the end of carotid artery (LCA)) HUVECs were seeded on gelatine-coated plates in medium supplemented with 10% FBS, heparin and 30 μg/mL endothelial cell growth supplement HUVECs were cultivated on plates coated with gelatine. The cells were cultured in a medium supplemented 10% FBS, heparin with additional 30 μg/mL endothelial cell growth factor Pre-treatment of GTs (500 μg/mL) or DMSO vehicle control were given for 1 h prior to treatment with H2O2 (400 μM) that was applied. It was incubated for another 24 h Parameters: Cell characterization by flowcytometry, reactive oxygen species, immunofluorescence and in vitro perfusion system | GL treatment prevented carotid artery from atherogenesis derived flow system GT treatment protected carotid artery from neointima formation GT reduced the oxidative stress and atherogenic events in HUVECs GTs inhibit the fluctuation of flow system-induced inflammation in HUVECs GTs provide a protection in HUVECs against oxidative stress GT treatment successfully halting atherogenesis | Anti-atherogenic effects provided by GL and GT are essential in terminating inflammation of HUVECs, which was promoted by disturbed flow system |
| 4 | Liu et al., 2016 [56] | In vitro: Primary foetal fibroblasts from black pig (as donor cells) In vivo: Pigs (undergone somatic cell nuclear transfer (SCNT) and embryo transfer) to induced hfat-1 transgenic pigs | Not applicable | Cardiovascular disease | The hfat-1 plasmid was transfected into primary foetal fibroblasts. The positive colonies were served as donor cells to conduct SCNT and embryo transfer into pig model. The pigs were housed in 18–20 °C environment. The diets were given three times daily, with a mixture of 15% crude protein and 4 % crude fat Genomic DNA derived piglets were isolated from tails, followed by gene expression analysis (PCR) and subsequently by gel electrophoresis and gene sequencing GC–MS was performed to evaluate the constituents of fatty acid in the ears based on previous preliminary studies 5ml of blood were collected from pig vein following 16 h of food depreciation. Beckman coulter UniCel DxC 800 Synchron was used to determine the level of TAG, total cholesterol (TC), HDL-C and LDL-C | PCR analysis showed positive expression of hfat-1 transgene in all piglets, which proved by the accumulation of n-3 PUFAs The expression of n-3 PUFAs is 2-fold higher in transgenic pigs compared to wild-type pigs There were no significant changes of n-6 PUFAs in transgenic pigs, and the level composition of fatty acids was alleviated in transgenic pigs compared to wild type pigs No significant different of total cholesterol, HDL-C and LDL-C levels in both type of pigs in deprivation condition, whereas TAG level was notably attenuated in transgenic pigs compared to wild-type pigs The transgenic pigs showed downregulation of MCP-1, IL-6 and TNF-α compared to wild-type pigs, which demonstrated that the accumulation of n-3 PUFAs plays a positive role in reducing the inflammatory effects | These findings revealed that the aggregation of n-3 PUFAs provides a promising impact on vasculoprotective studies. The use of transgenic pigs might be beneficial in determining the inflammatory mechanisms in the body |
| 5 | Sanders et al., 2017 [57] | In vitro: Human venous smooth muscle cells In vivo: 1. Porcine model 2. Adipocytes 3. Peripheral blood mononuclear cells | 1. Rosiglitazone drugs (ROS) 2. Pioglitazone drugs (PGZ) | Stenosis caused by intimal hyperplasia (Develops following the installation of arterial synthetic grafts during haemodialysis) | ROS or PGZ in powder form (6–6000 μM) was integrated thoroughly with adipose tissue explants and cultivated in the medium HPLC/MS/MS was performed to quantify the drug that release from adipose tissue. Secretion of adiponectin and monocyte chemotactic protein-1 (MCP-1) in the conditioned media were determined by ELISA. Platelet-derived growth factor-BB (PDGG-BB) was evaluated through DNA binding assay, whereas lipopolysaccharide (LPS) was measured by ELISA HPLC/MS/MS were also performed to determine the kinetics of PGZ. MRI were used to monitor the depository of adipose in a porcine model The effects of PGZ/ fat depository were assessed using a porcine model with the installation of synthetic graft at the region between carotid artery and jugular vein | Powdered PGZ and ROS induced the secretion of adiponectin in conditioned media The secretion of MCP-1 in conditioned media were increased. However, the treatment using PGZ and ROS alleviated MCP-1 level Drugs was not detected in the pig treated with fat/PGZ depot but slight detection (0.06 ± 0.06 μM) can be seen in the pig with fat/ROS depots treatment. MRI monitoring demonstrated that the PGZ/fat depots resided near the perivascular region of jugular vein The expression of adiponectin protein in jugular vein was increased in the fat/PGZ transplant Treatment with fat/PGZ depots showed identical graft lumen compared to control group | Treatment with fat/PGZ depots effectively regulates the expression of adiponectin and reduced inflammatory activities. Hence, the findings could be considered as a promising therapy in halting stenosis due to intimal hyperplasia |
| 6 | Schlosser et al., 2015 [58] | In vitro: Vascular smooth muscle cells and monocyte In vivo: Mfap4-deficient (Mfap4−/−) mice Ex vivo: Neointima of Mfap4-deficient (Mfap4−/−) mice | Microfibrillar-associated protein 4 (MFAP4) | Neointimal hyperplasia | Materials and Methods were described in data supplement that are available online | Mfap4−/− mice impeded neointima development following artery ligation. This is related to the proliferation of VSMC and the invasion of leukocyte The presence of MFAP4 was detected both in healthy and diseased vessel walls, thus stimulating the development of neointima VSMC synthesized MFAP4 and mediated binding to integrin αVβ3 Stimulation MFAP4 mediated cell proliferation and induced the migratory effect on VSMC MFAP4 is effective in protecting VSMC from apoptosis and monocytic cell migration | MFAP4 has a great effect in regulating integrin αVβ which promoted VSMC migration and proliferation, thus increased the formation of neointimal hyperplasia |
| 7 | Wang et al., 2018 [59] | In vitro: Smooth muscle-like cells (SMLCs) from the neointima of the outflow vein In vivo: 1. C57BL/6 mice 2. miR-155-/- mice (C57BL/6) | Not applicable | Neointimal hyperplasia | C57BL/6 and miR-155 mice were subjected to AVFs at the region of jugular vein and carotid artery The mice were euthanized. At days 7, 14, 21 and 28, the segments were harvested. The AVFs were subjected to histological staining, and gene expressions were evaluated using RTPCR Native saphenous vein was obtained from patients who underwent CABG surgery and suffered from chronic kidney failure. The samples were assessed by histological staining | Knockout of miR-155 decreases the level of proinflammatory cytokines in the outflow vein and attenuates the formation of neointimal hyperplasia Knockout of miR-155 halted SMLCs proliferation and alleviates the regulation of ECM RANTES was upregulated in SMLCs with the increment of miR-155 SMLCs proliferation and ECM expression were enhanced by miR-155 due to high level of RANTES Upregulation of RANTES expression was promoted by miR-155 Patients with neointimal hyperplasia showed the high expression of inflammatory factors in the outflow vein | miR-155 promotes the formation of intimal hyperplasia through the upregulation of RANTES that provide inflammatory effects in SMLCs |
| 8 | Yang et al., 2016 [60] | In vitro: Adipose tissue–derived mesenchymal stem cells (MSC) In vivo: B6.Cg-Foxn1nu/J mice | Not applicable | Neointimal hyperplasia | B6. Cg-Foxn1nu/J mice were subjected to arteriovenous fistula (AVF) and utilized for the experiment MSCs was labelled with GFP and directly injected into outflow vein upon AVF installation At day 7, mice were euthanized following AVF installation. The tissues were analysed by RTPCR and histomorphometric analyses. At day 21, the tissues were evaluated only by histomorphometric analysis. MSCs labelled with 89Zr were implanted in a different group. The animals were examined using positron electron tomography for 3 weeks. The data were analysed using two-way analysis of variance and subsequently by Student t test | Transplanted MSCs showed GFP-positive cells that were detected at day 7, whereas at day 21, GFP signal cannot be seen MCP-1 was downregulated at the outflow vein resulted from MSCs transplantation Transplanted MSCs attenuates the neointimal, medial and adventitial regions, and cell concentration was also decreased, whereas the lumen area was increased at day 7 and day 21 TUNEL staining demonstrated positive staining at the outflow vein resulted from MSCs transplantation MSCs transplantation alleviates cell proliferation and decreases the expression of FSP-1, a-SMA, HIF-1a and CD68 | Adventitial transplantation of MSCs decreases MCP-1 gene expression, accompanied by a reduction in venous neointimal hyperplasia MSCs transplantation reduced the expression of MCP-1, and thus reduced the formation of neointimal hyperplasia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Che Man, R.; Sulaiman, N.; Ishak, M.F.; Bt Hj Idrus, R.; Abdul Rahman, M.R.; Yazid, M.D. The Effects of Pro-Inflammatory and Anti-Inflammatory Agents for the Suppression of Intimal Hyperplasia: An Evidence-Based Review. Int. J. Environ. Res. Public Health 2020, 17, 7825. https://doi.org/10.3390/ijerph17217825
Che Man R, Sulaiman N, Ishak MF, Bt Hj Idrus R, Abdul Rahman MR, Yazid MD. The Effects of Pro-Inflammatory and Anti-Inflammatory Agents for the Suppression of Intimal Hyperplasia: An Evidence-Based Review. International Journal of Environmental Research and Public Health. 2020; 17(21):7825. https://doi.org/10.3390/ijerph17217825
Chicago/Turabian StyleChe Man, Rohaina, Nadiah Sulaiman, Mohamad Fikeri Ishak, Ruszymah Bt Hj Idrus, Mohd Ramzisham Abdul Rahman, and Muhammad Dain Yazid. 2020. "The Effects of Pro-Inflammatory and Anti-Inflammatory Agents for the Suppression of Intimal Hyperplasia: An Evidence-Based Review" International Journal of Environmental Research and Public Health 17, no. 21: 7825. https://doi.org/10.3390/ijerph17217825