Resistance Exercise Training Attenuates the Loss of Endogenous GLP-1 Receptor in the Hypothalamus of Type 2 Diabetic Rats



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
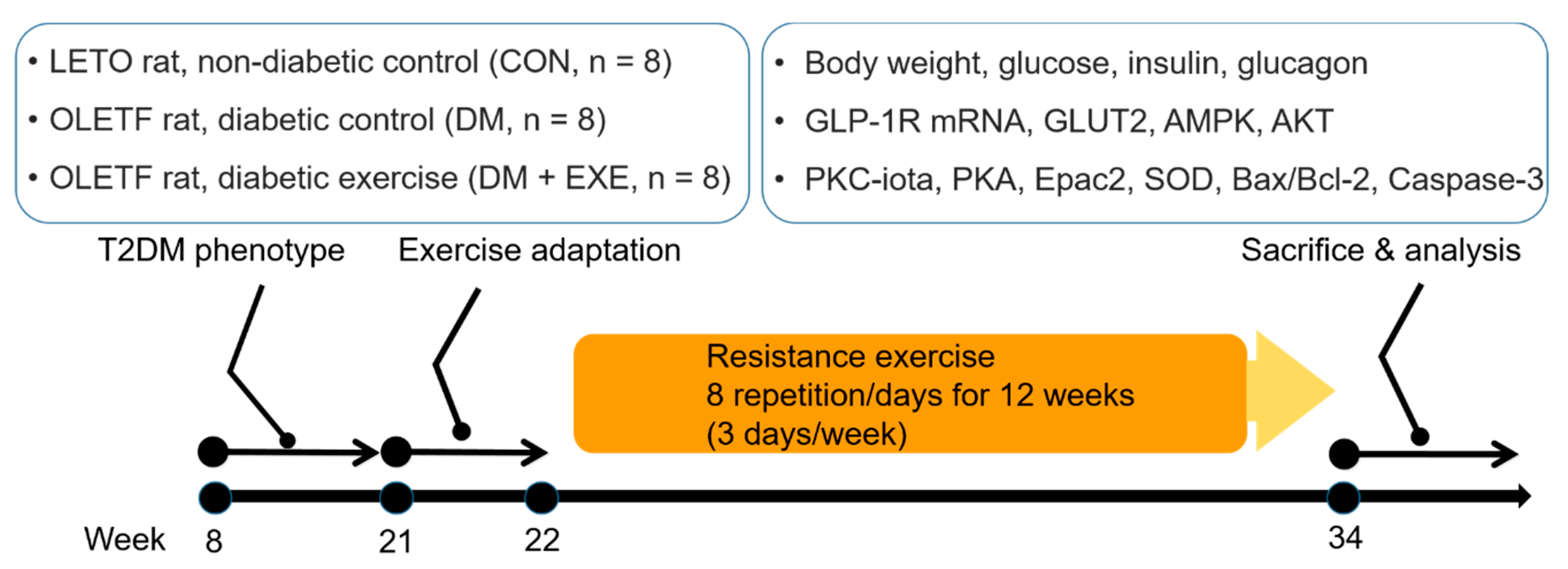
2. Materials and Methods
2.1. Experimental Animals
2.2. Exercise Protocol
2.3. Tissue Preparation and Blood Parameters
2.4. Western Blotting
2.5. Real-Time PCR
2.6. Statistics
3. Results
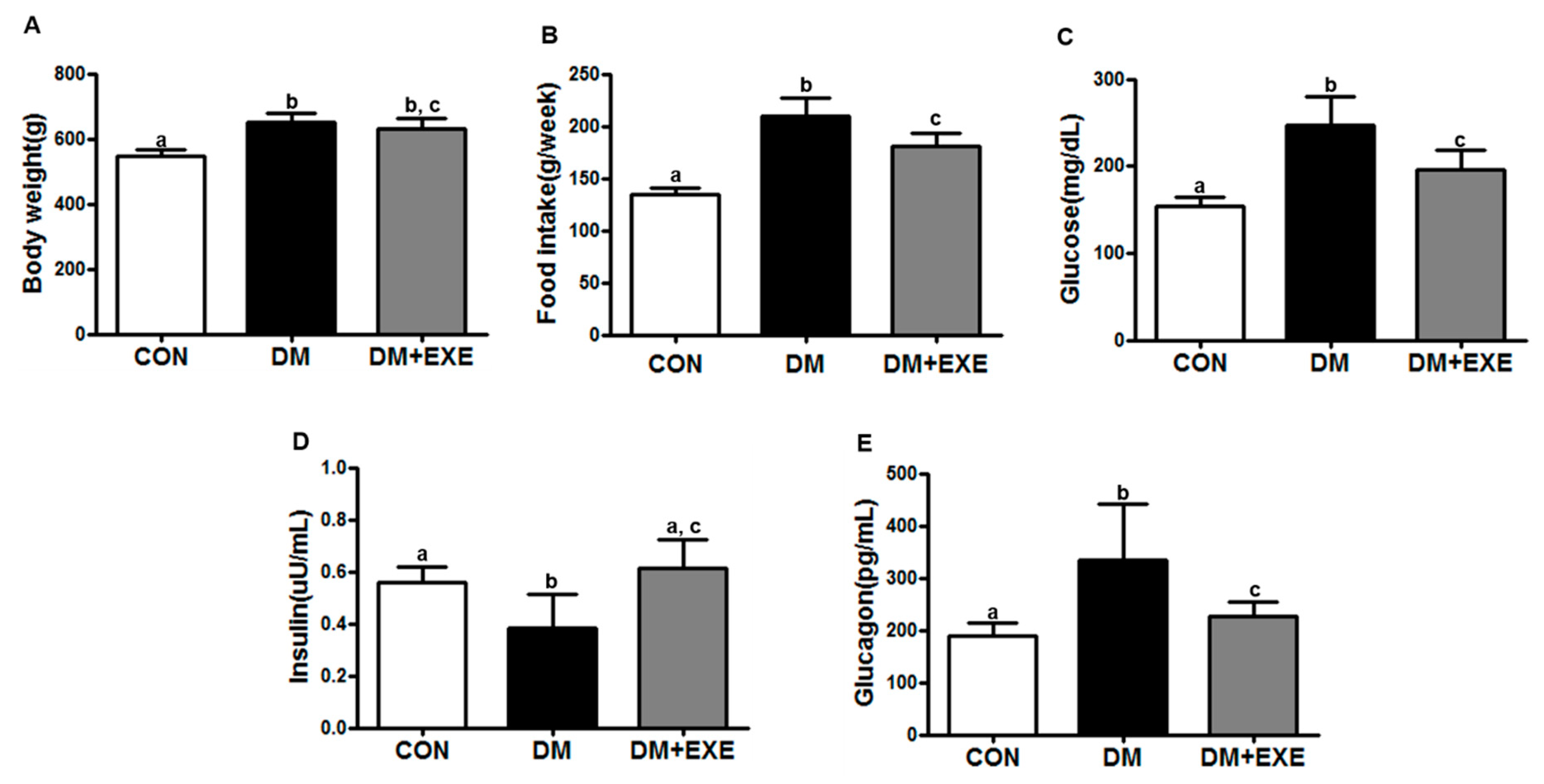
3.1. Resistance Exercise Improved Food Intake, Gluocse, Insulin, and Glucagon Levels in T2DM Rats Regardless of Body Weight
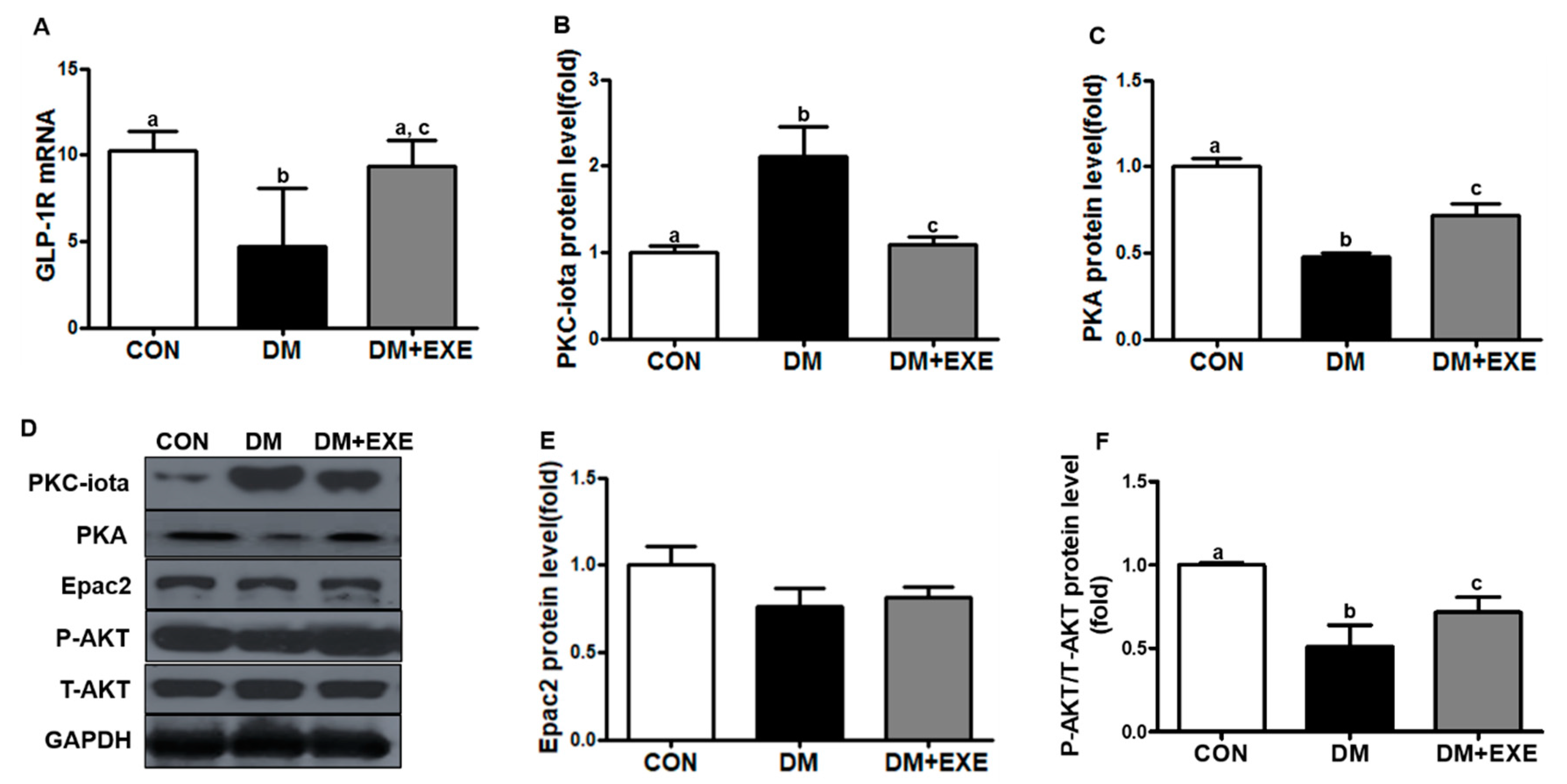
3.2. Resistance Exercise Increased GLP-1R Levels by Decreasing PKC-iota and Enhancing the Related Signaling Pathway in the Hypothalamus
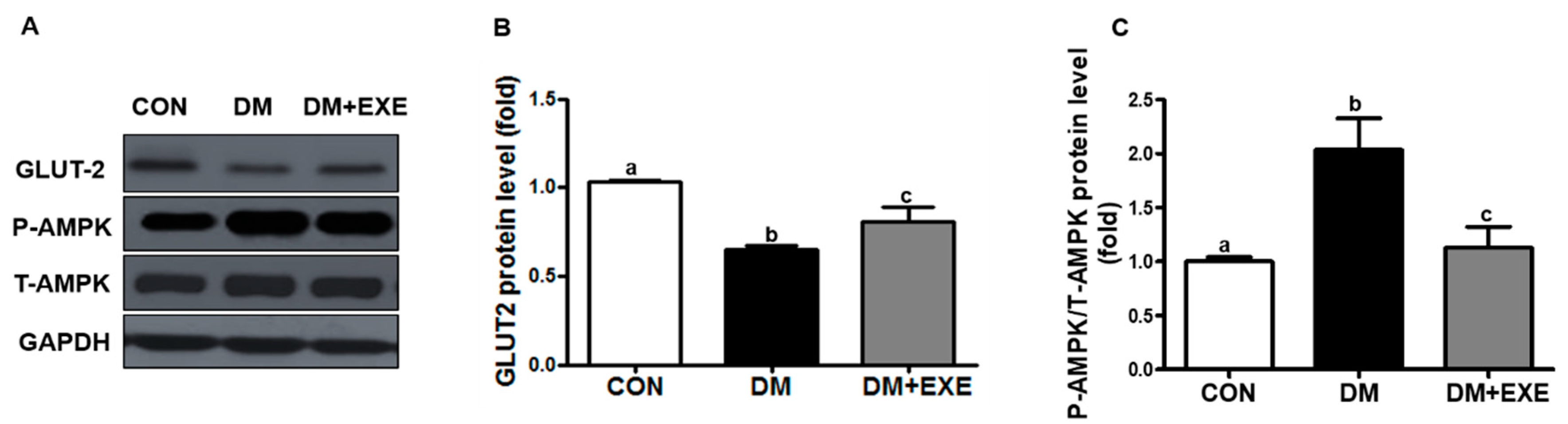
3.3. Resistance Exercise Increased GLUT-2 and Attenuated Increases in AMPK in the Hypothalamus
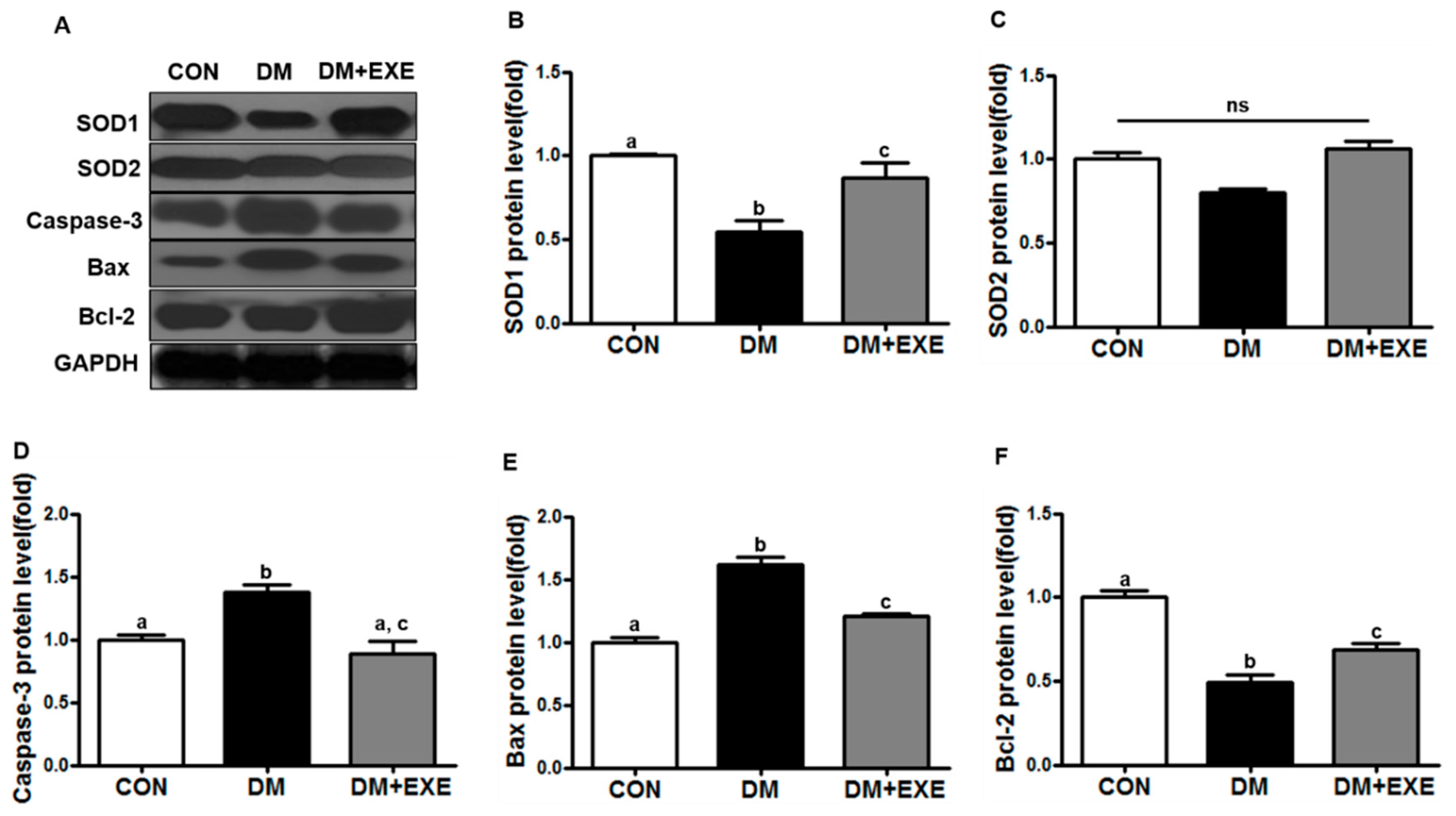
3.4. Resistance Exercise Improved Antioxidant Enzyme and Apoptotic Protein Levels
4. Discussion
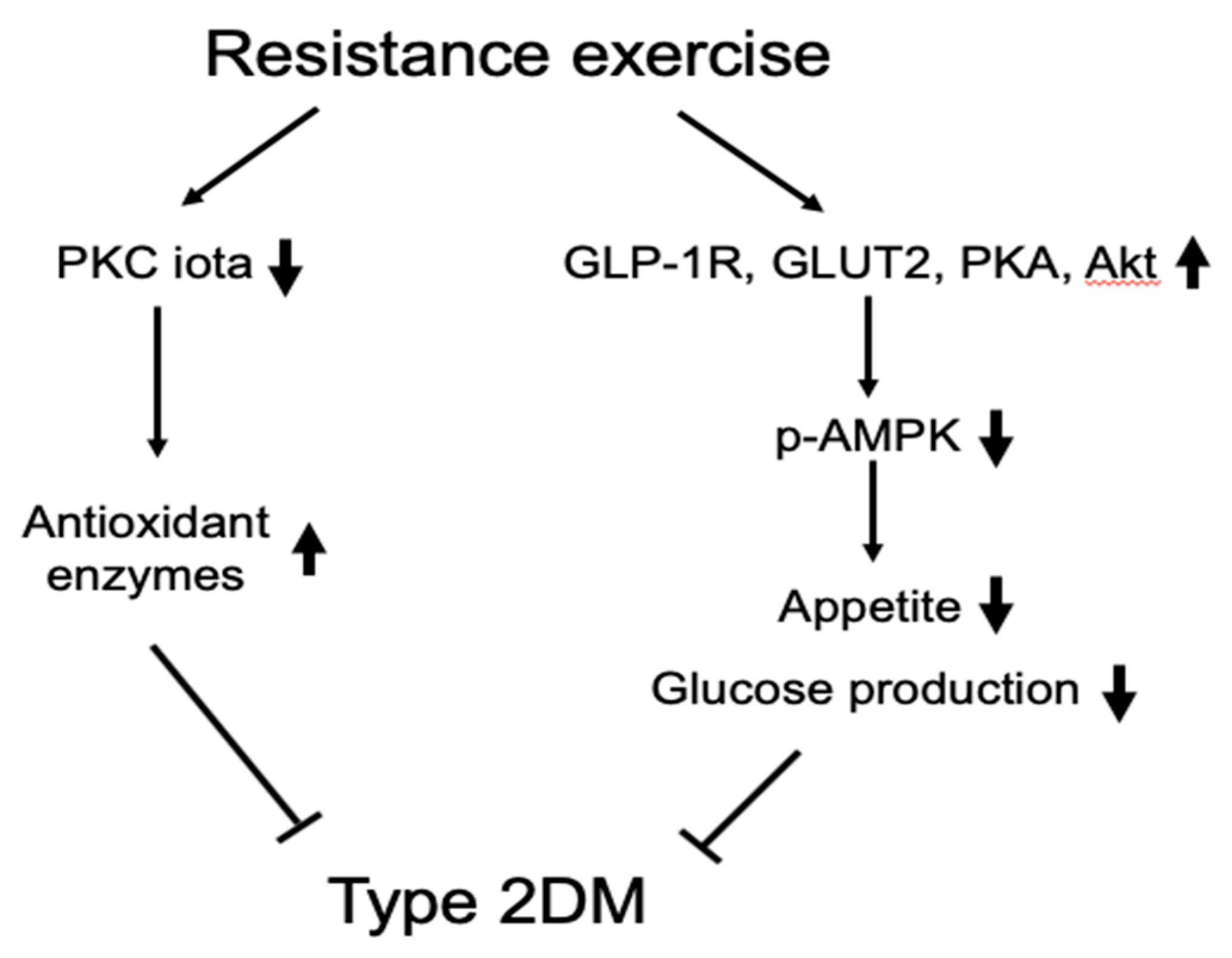
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Marques, C.; Mega, C.; Goncalves, A.; Rodrigues-Santos, P.; Teixeira-Lemos, E.; Teixeira, F.; Fontes-Ribeiro, C.; Reis, F.; Fernandes, R. Sitagliptin prevents inflammation and apoptotic cell death in the kidney of type 2 diabetic animals. Mediat. Inflamm. 2014, 2014, 538737. [Google Scholar] [CrossRef] [PubMed]
- Defronzo, R.A. Banting Lecture. From the triumvirate to the ominous octet: A new paradigm for the treatment of type 2 diabetes mellitus. Diabetes 2009, 58, 773–795. [Google Scholar] [CrossRef] [PubMed]
- Khunti, K.; Wolden, M.L.; Thorsted, B.L.; Andersen, M.; Davies, M.J. Clinical inertia in people with type 2 diabetes: A retrospective cohort study of more than 80,000 people. Diabetes Care 2013, 36, 3411–3417. [Google Scholar] [CrossRef] [PubMed]
- Smokovski, I.; Milenkovic, T.; Trapp, C.; Mitov, A. Diabetes Care in Republic of Macedonia: Challenges and Opportunities. Ann. Glob. Health 2015, 81, 792–802. [Google Scholar] [CrossRef] [PubMed]
- George, C.M.; Brujin, L.L.; Will, K.; Howard-Thompson, A. Management of Blood Glucose with Noninsulin Therapies in Type 2 Diabetes. Am. Fam. Physician 2015, 92, 27–34. [Google Scholar] [PubMed]
- American Diabetes Association. 4. Prevention or Delay of Type 2 Diabetes. Diabetes Care 2016, 39, S36–S38. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Seeley, R.J.; Tschop, M.H.; Woods, S.C.; Morton, G.J.; Myers, M.G.; D’Alessio, D. Cooperation between brain and islet in glucose homeostasis and diabetes. Nature 2013, 503, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, A.P.; Yeo, G.S. The hypothalamus and metabolism: Integrating signals to control energy and glucose homeostasis. Curr. Opin. Pharmacol. 2013, 13, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, V.A.; Zinman, B.; Nauck, M.A.; Goldfine, A.B.; Plutzky, J. Confronting the type 2 diabetes epidemic: The emerging role of incretin-based therapies. Am. J. Med. 2010, 123, S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Flock, G.; Baggio, L.L.; Longuet, C.; Drucker, D.J. Incretin receptors for glucagon-like peptide 1 and glucose-dependent insulinotropic polypeptide are essential for the sustained metabolic actions of vildagliptin in mice. Diabetes 2007, 56, 3006–3013. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pang, Z.P. Glucagon-like peptide-1 drives energy metabolism on the synaptic highway. FEBS J. 2016, 283, 4413–4423. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, D.A.; Bagnol, D.; Woods, S.C.; D’Alessio, D.A.; Seeley, R.J. Arcuate glucagon-like peptide 1 receptors regulate glucose homeostasis but not food intake. Diabetes 2008, 57, 2046–2054. [Google Scholar] [CrossRef] [PubMed]
- Koole, C.; Pabreja, K.; Savage, E.E.; Wootten, D.; Furness, S.G.; Miller, L.J.; Christopoulos, A.; Sexton, P.M. Recent advances in understanding GLP-1R (glucagon-like peptide-1 receptor) function. Biochem. Soc. Trans. 2013, 41, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmeister, M.A.; Ayala, J.; Drucker, D.J.; Ayala, J.E. Central glucagon-like peptide 1 receptor-induced anorexia requires glucose metabolism-mediated suppression of AMPK and is impaired by central fructose. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E677–E685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Lee, K.; Martin, R.J. Overexpression of glucose transporter 2 in GT1-7 cells inhibits AMP-activated protein kinase and agouti-related peptide expression. Brain Res. 2006, 1118, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Vilsboll, T.; Christensen, M.; Junker, A.E.; Knop, F.K.; Gluud, L.L. Effects of glucagon-like peptide-1 receptor agonists on weight loss: Systematic review and meta-analyses of randomised controlled trials. BMJ 2012, 344, d7771. [Google Scholar] [CrossRef] [PubMed]
- Gallwitz, B. Anorexigenic effects of GLP-1 and its analogues. Handb. Exp. Pharmacol. 2012, 209, 185–207. [Google Scholar]
- Hayes, M.R.; Leichner, T.M.; Zhao, S.; Lee, G.S.; Chowansky, A.; Zimmer, D.; De Jonghe, B.C.; Kanoski, S.E.; Grill, H.J.; Bence, K.K. Intracellular signals mediating the food intake-suppressive effects of hindbrain glucagon-like peptide-1 receptor activation. Cell Metab. 2011, 13, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Barrera, J.G.; D’Alessio, D.A.; Drucker, D.J.; Woods, S.C.; Seeley, R.J. Differences in the central anorectic effects of glucagon-like peptide-1 and exendin-4 in rats. Diabetes 2009, 58, 2820–2827. [Google Scholar] [CrossRef] [PubMed]
- Pannacciulli, N.; Le, D.S.; Salbe, A.D.; Chen, K.; Reiman, E.M.; Tataranni, P.A.; Krakoff, J. Postprandial glucagon-like peptide-1 (GLP-1) response is positively associated with changes in neuronal activity of brain areas implicated in satiety and food intake regulation in humans. Neuroimage 2007, 35, 511–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajan, S.; Dickson, L.M.; Mathew, E.; Orr, C.M.; Ellenbroek, J.H.; Philipson, L.H.; Wicksteed, B. Chronic hyperglycemia downregulates GLP-1 receptor signaling in pancreatic beta-cells via protein kinase A. Mol. Metab. 2015, 4, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of incretin hormone action. Cell Metab. 2013, 17, 819–837. [Google Scholar] [CrossRef] [PubMed]
- Francois, M.E.; Durrer, C.; Pistawka, K.J.; Halperin, F.A.; Little, J.P. Resistance-based interval exercise acutely improves endothelial function in type 2 diabetes. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1258–H1267. [Google Scholar] [CrossRef] [PubMed]
- Talebi-Garakani, E.; Safarzade, A. Resistance training decreases serum inflammatory markers in diabetic rats. Endocrine 2013, 43, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Kawano, K.; Hirashima, T.; Mori, S.; Saitoh, Y.; Kurosumi, M.; Natori, T. Spontaneous long-term hyperglycemic rat with diabetic complications. Otsuka Long-Evans Tokushima Fatty (OLETF) strain. Diabetes 1992, 41, 1422–1428. [Google Scholar] [CrossRef] [PubMed]
- Karimian, J.; Khazaei, M.; Shekarchizadeh, P. Effect of Resistance Training on Capillary Density Around Slow and Fast Twitch Muscle Fibers in Diabetic and Normal Rats. Asian J. Sports Med. 2015, 6, e24040. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.Y.; Lee, S.R.; Kwak, H.B.; Seo, K.W.; McGregor, R.A.; Yeo, J.Y.; Ko, T.H.; Bolorerdene, S.; Kim, N.; Ko, K.S.; et al. Voluntary stand-up physical activity enhances endurance exercise capacity in rats. Korean J. Physiol. Pharmacol. 2016, 20, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Patel, Y.R.; Kirkman, M.S.; Considine, R.V.; Hannon, T.S.; Mather, K.J. Changes in Weight and Glucose Can Protect Against Progression in Early Diabetes Independent of Improvements in beta-Cell Function. J. Clin. Endocrinol. Metab. 2016, 101, 4076–4084. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; So, B.; Son, J.S.; Song, H.S.; Oh, S.L.; Seong, J.K.; Lee, H.; Song, W. Resistance training inhibits the elevation of skeletal muscle derived-BDNF level concomitant with improvement of muscle strength in zucker diabetic rat. J. Exerc. Nutr. Biochem. 2015, 19, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozo, M.; Claret, M. Hypothalamic Control of Systemic Glucose Homeostasis: The Pancreas Connection. Trends Endocrinol. Metab. 2018, 29, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Jun, H.S. Anti-Inflammatory Effects of GLP-1-Based Therapies beyond Glucose Control. Mediat. Inflamm. 2016, 2016, 3094642. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.I.; Park, M.J.; Choi, J.H.; Lim, S.K.; Choi, H.J.; Park, S.H. Hyperglycemia-induced GLP-1R downregulation causes RPE cell apoptosis. Int. J. Biochem. Cell Biol. 2015, 59, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Kaneto, H.; Laybutt, D.R.; Duvivier-Kali, V.F.; Trivedi, N.; Suzuma, K.; King, G.L.; Weir, G.C.; Bonner-Weir, S. Downregulation of GLP-1 and GIP receptor expression by hyperglycemia: Possible contribution to impaired incretin effects in diabetes. Diabetes 2007, 56, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Mensberg, P.; Nyby, S.; Jorgensen, P.G.; Storgaard, H.; Jensen, M.T.; Sivertsen, J.; Holst, J.J.; Kiens, B.; Richter, E.A.; Knop, F.K.; et al. Near-normalization of glycaemic control with glucagon-like peptide-1 receptor agonist treatment combined with exercise in patients with type 2 diabetes. Diabetes Obes. Metab. 2017, 19, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Heiskanen, M.A.; Motiani, K.K.; Mari, A.; Saunavaara, V.; Eskelinen, J.J.; Virtanen, K.A.; Koivumaki, M.; Loyttyniemi, E.; Nuutila, P.; Kalliokoski, K.K.; et al. Exercise training decreases pancreatic fat content and improves beta cell function regardless of baseline glucose tolerance: A randomised controlled trial. Diabetologia 2018, 61, 1817–1828. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Kulseng, B.; King, N.A.; Holst, J.J.; Blundell, J.E. The effects of exercise-induced weight loss on appetite-related peptides and motivation to eat. J. Clin. Endocrinol. Metab. 2010, 95, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Yoo, J.H.; So, Y.S. Effect of the low- versus high-intensity exercise training on endoplasmic reticulum stress and GLP-1 in adolescents with type 2 diabetes mellitus. J. Phys. Ther. Sci. 2015, 27, 3063–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasyurek, H.M.; Altunbas, H.A.; Balci, M.K.; Sanlioglu, S. Incretins: Their physiology and application in the treatment of diabetes mellitus. Diabetes Metab. Res. Rev. 2014, 30, 354–371. [Google Scholar] [CrossRef] [PubMed]
- Combs, C.K. Are GLP-1 receptor agonists useful against traumatic brain injury? J. Neurochem. 2015, 135, 1059–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deacon, C.F.; Mannucci, E.; Ahren, B. Glycaemic efficacy of glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors as add-on therapy to metformin in subjects with type 2 diabetes—A review and meta analysis. Diabetes Obes. Metab. 2012, 14, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Millan, C.; Martinez, F.; Cortes-Campos, C.; Lizama, I.; Yanez, M.J.; Llanos, P.; Reinicke, K.; Rodriguez, F.; Peruzzo, B.; Nualart, F.; et al. Glial glucokinase expression in adult and post-natal development of the hypothalamic region. ASN Neuro 2010, 2, e00035. [Google Scholar] [CrossRef] [PubMed]
- Bady, I.; Marty, N.; Dallaporta, M.; Emery, M.; Gyger, J.; Tarussio, D.; Foretz, M.; Thorens, B. Evidence from glut2-null mice that glucose is a critical physiological regulator of feeding. Diabetes 2006, 55, 988–995. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, I.S.; Zemdegs, J.C.; de Souza, A.P.; Watanabe, R.L.; Telles, M.M.; Nascimento, C.M.; Oyama, L.M.; Ribeiro, E.B. Diet-induced obesity impairs hypothalamic glucose sensing but not glucose hypothalamic extracellular levels, as measured by microdialysis. Nutr. Diabetes 2015, 5, e162. [Google Scholar] [CrossRef] [PubMed]
- Mace, O.J.; Schindler, M.; Patel, S. The regulation of K- and L-cell activity by GLUT2 and the calcium-sensing receptor CasR in rat small intestine. J. Physiol. 2012, 590, 2917–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, M. AMPK Wars: The VMH Strikes Back, Return of the PVH. Trends Endocrinol. Metab. 2018, 29, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Tomaz, L.M.; Barbosa, M.R.; Farahnak, Z.; Lagoeiro, C.G.; Magosso, N.S.; Lavoie, J.M.; Perez, S.E. GLUT2 proteins and PPARgamma transcripts levels are increased in liver of ovariectomized rats: Reversal effects of resistance training. J. Exerc. Nutr. Biochem. 2016, 20, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Kiraly, M.A.; Bates, H.E.; Kaniuk, N.A.; Yue, J.T.; Brumell, J.H.; Matthews, S.G.; Riddell, M.C.; Vranic, M. Swim training prevents hyperglycemia in ZDF rats: Mechanisms involved in the partial maintenance of beta-cell function. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E271–E283. [Google Scholar] [CrossRef] [PubMed]
- Kjobsted, R.; Pedersen, A.J.; Hingst, J.R.; Sabaratnam, R.; Birk, J.B.; Kristensen, J.M.; Hojlund, K.; Wojtaszewski, J.F. Intact Regulation of the AMPK Signaling Network in Response to Exercise and Insulin in Skeletal Muscle of Male Patients With Type 2 Diabetes: Illumination of AMPK Activation in Recovery From Exercise. Diabetes 2016, 65, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Bertram, S.; Brixius, K.; Brinkmann, C. Exercise for the diabetic brain: How physical training may help prevent dementia and Alzheimer’s disease in T2DM patients. Endocrine 2016, 53, 350–363. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.H.; Yoon, J.H.; Seo, D.Y.; Kim, T.N.; Ko, J.R.; Han, J. Resistance Exercise Training Attenuates the Loss of Endogenous GLP-1 Receptor in the Hypothalamus of Type 2 Diabetic Rats. Int. J. Environ. Res. Public Health 2019, 16, 830. https://doi.org/10.3390/ijerph16050830
Park SH, Yoon JH, Seo DY, Kim TN, Ko JR, Han J. Resistance Exercise Training Attenuates the Loss of Endogenous GLP-1 Receptor in the Hypothalamus of Type 2 Diabetic Rats. International Journal of Environmental Research and Public Health. 2019; 16(5):830. https://doi.org/10.3390/ijerph16050830
Chicago/Turabian StylePark, Se Hwan, Jin Hwan Yoon, Dae Yun Seo, Tae Nyun Kim, Jeong Rim Ko, and Jin Han. 2019. "Resistance Exercise Training Attenuates the Loss of Endogenous GLP-1 Receptor in the Hypothalamus of Type 2 Diabetic Rats" International Journal of Environmental Research and Public Health 16, no. 5: 830. https://doi.org/10.3390/ijerph16050830