
Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae) †


School of Health, Medical & Applied Sciences, Central Queensland University, Rockhampton, QLD 4701, Australia
†
Presented at the 2nd International Electronic Conference on Diversity (IECD 2022)—New Insights into the Biodiversity of Plants, Animals and Microbes, 15–31 March 2022; Available online: https://sciforum.net/event/IECD2022 .
Biol. Life Sci. Forum 2022, 15(1), 5; https://doi.org/10.3390/IECD2022-12383
Published: 14 March 2022
(This article belongs to the Proceedings of The 2nd International Electronic Conference on Diversity (IECD 2022)—New Insights into the Biodiversity of Plants, Animals and Microbes)
Abstract
:Cuticular hydrocarbons (CHCs) have been used as a chemotaxonomic tool to support the classification and identification of various insect species for decades. However, there have been limited research performed on the CHC profiles of weevils (Coleoptera: Curculionidae), despite the extensive diversity and ecological significance of this family. In this study, CHCs were extracted from fifteen Gonipterini weevil specimens from eastern Australia, comprising five species from three genera. Analysis by gas chromatography-mass spectrometry (GC-MS) revealed the presence of over 90 compounds, with the most abundant compounds including nonacosane, 7-methylheptacosane, heptacosane and hexacosane. Principal component analysis revealed Bryachus squamicollis to be the most dissimilar species in terms of its CHC profile, while the two Oxyops species showed relatively similar CHC profiles. The results may support the use of CHC profiling as a chemotaxonomic tool for the identification and delineation of various Gonipterini genera and species.
1. Introduction
The Gonipterini tribe of weevils (Coleoptera: Curculionidae) contains a number of economically important species, including several species of Gonipterus which have become international pests of Eucalyptus plantations [1,2] and Oxyops vitiosa, which has been used for the biocontrol of the invasive species Melaleuca quinquenervia in the Florida everglades [3]. Consequently, the accurate identification of species from this tribe is of utmost importance. However, Gonipterus in particular contains a number of cryptic species, the identification of which typically requires molecular analysis and/or dissection of male genitalia [2].
However, the emergence of cuticular hydrocarbon (CHC) profiling has emerged as a useful chemotaxonomic tool over the past few decades [4]. These compounds are endogenously synthesized by nearly all insects and exported to the cuticle [5,6], with one of their major functions being to mediate communication with other insects. Consequently, there is a wide range of diversity in CHC profiles, which is reflective of the genetic diversity of the species in question. Although CHC profiling has been used to support the classification and discrimination of other beetle species [7,8], there have been limited studies to date on the weevil family, despite the large number of species present.
Recently, Souza, et al. [9] demonstrated that the differences in CHC profiles between a number of Gonipterus species concurred with molecular sequencing data and morphological features, demonstrating that this method may be used for the rapid classification of species from this genus. However, these authors did not examine the CHC profiles of other Gonipterini genera in detail, only using Oxyops (one or more unidentified species) as an outgroup. Specifically, there has been no work to date looking at the CHC profiles of different Oxyops or Bryachus species. In order to fill this research gap, the present study aimed to extend the use of CHC profiling to different genera and species from the Gonipterini tribe.
2. Materials and Methods
In February 2021, 15 weevils, comprising three genera and five species, were collected from Eucalyptus populnea saplings on a Central Queensland grazing property (23°46′ S, 150°21′ E). Details of the specimens are provided in Table 1, while scale photographs are given in Figure 1. CHCs were extracted from each weevil using 300 μL of hexane with 4 min of agitation, following the methods of Souza, et al. [9]. The hexane extracts were analysed by gas chromatography-mass spectrometry (GC-MS), following the methods of Souza, et al. [9], but using a single quadrupole Shimadzu QP2010 Plus system fitted with a Shimadzu SH-Rxi-5Sil MS column (29 m × 0.25 mm i.d. × 0.25 µm thickness; Columbia, MD, USA). Compounds were identified from comparison of their mass spectra and linear retention indices (LRIs; calculated from a series of C8–C40 alkanes run under the same conditions; Sigma Aldrich; Castle Hill, NSW, Australia) to literature values [10,11,12]. Data analysis was conducted in R studio [13].
3. Results and Discussion
3.1. CHC Profiles
A total of 97 compounds were found across the five weevil species, with 59 of these identified from their mass spectra and LRI, and a further 24 compounds tentatively identified from their mass spectra where their LRIs could not be found in the literature. Table 2 lists the identified compounds, along with their relative abundance in each of the weevil species. This number of compounds was significantly more than the 31 CHCs identified by Souza, et al. [9], likely due to the wider range of genetic diversity sampled here (i.e., three different Gonipterini genera instead of two). In addition, compounds eluting earlier than icosane (C20) were considered in this study.
The most abundant compounds across all five species were nonacosane and 7-methylheptacosane. Notably, Souza, et al. [9] had not reported these CHCs from their work on Gonipterus species. As both of these compounds were identified from their mass spectra and LRIs, a high level of confidence can be had in the identities reported here.
B. squamicollis also contained high levels of heptacosane, while Oxyops sp. contained higher concentrations of 2-methyloctacosane, and both Gonipterus species contained high levels of hexacosane, comparative to the levels reported by Souza, et al. [9] for several Gonipterus species. Oxyops sp. also contained much lower concentrations of 7-methylheptacosane compared to the other species.
Notably, very low levels of 2-methylhexacosane were found in the species studied here; whereas Souza, et al. [9] found relatively high concentrations of this compound in their species of Gonipterus (although it was not detected in Oxyops).
3.2. Chemometric Analysis
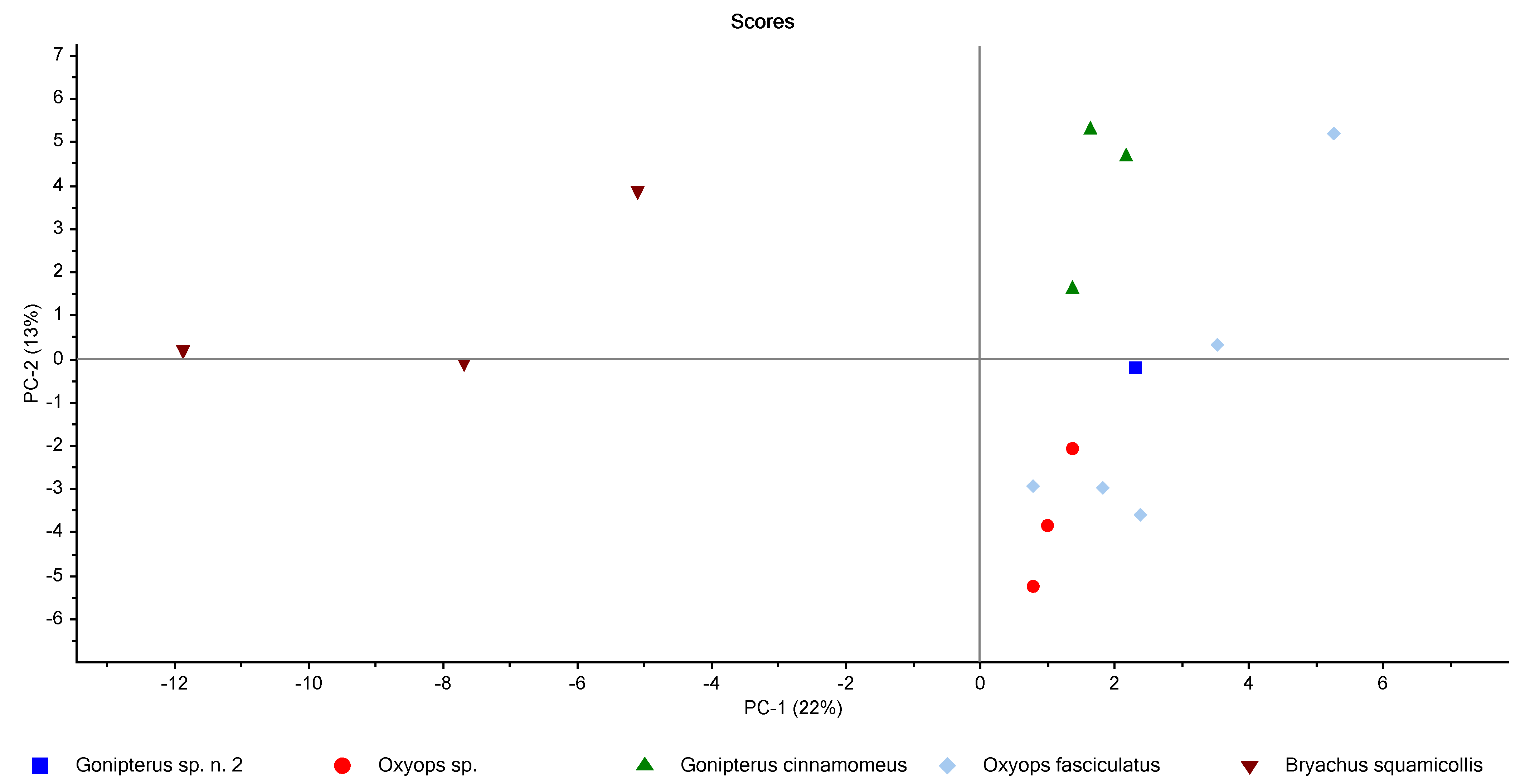
In order to visualize the broad differences in CHC composition between species, principal component analysis (PCA) was performed on the normalized CHC dataset. As can be seen from the scores plot in Figure 2, B. squamicollis was well separated from the remaining species, while the Gonipterus specimens were located toward the centre of the plot. The undescribed Oxyops species showed some overlap with O. fasciculatus, although it was slightly more separated across the third principal component (data not shown).
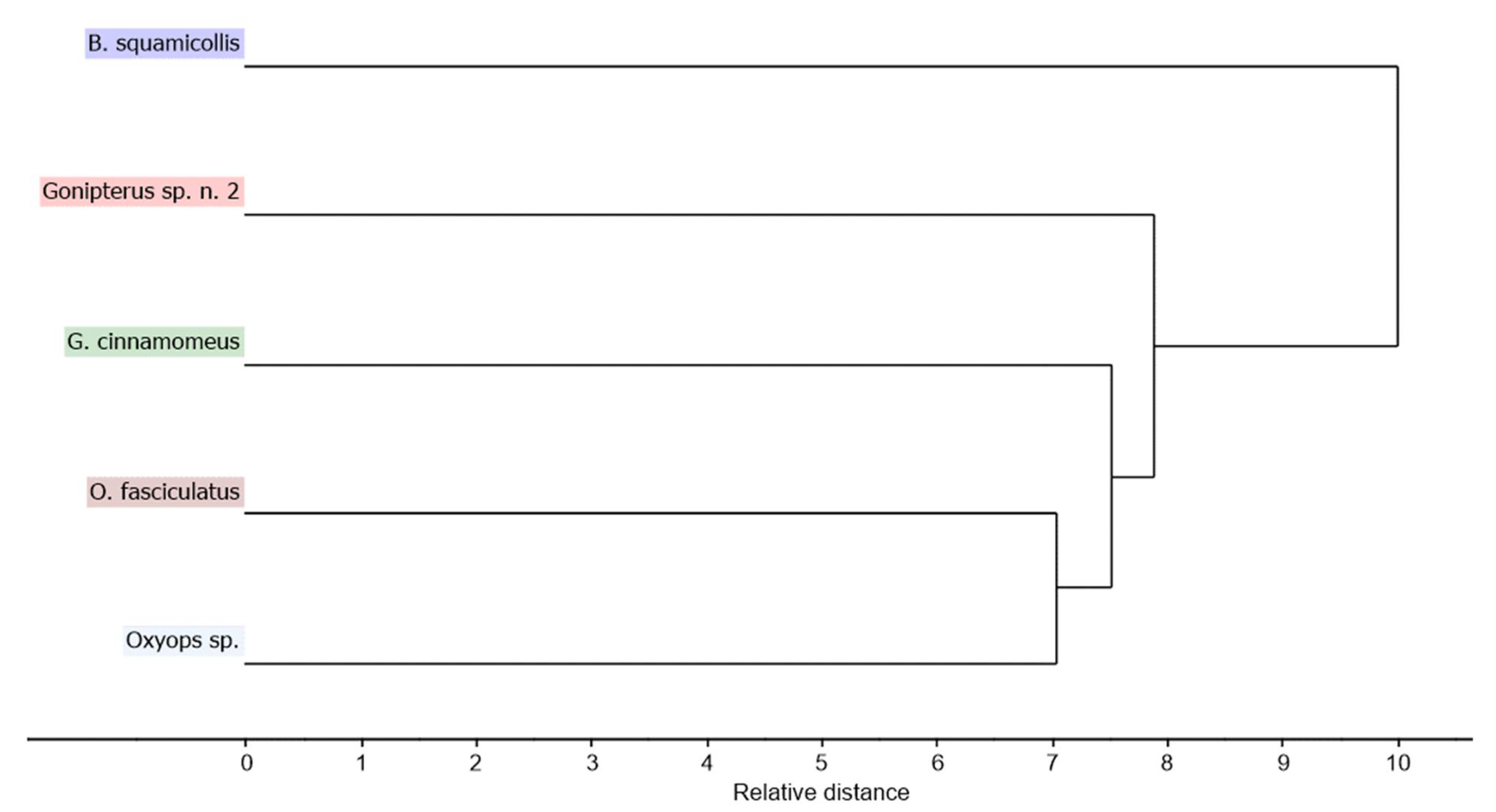
Hierarchical cluster analysis, performed on the normalized GC-MS data using the Euclidean distance with Ward’s method of clustering, displayed similar results. Bryachus squamicollis was again identified as the outlier taxon, while the two Oxyops species were most closely related in terms of their CHC composition (Figure 3).
4. Conclusions
The results of this proof-of-concept study support the use of CHC profiling as a potential chemotaxonomic method for discriminating between different genera and species of Gonipterini weevil. Around three times as many compounds were identified and quantified compared to previous work focusing on Gonipterus [9], with some of identities of the major CHCs differing from those found in previous work on the Gonipterini. As suggested by Souza, et al. [9], it may be possible to extract CHCs from live specimens in the future, further increasing the usefulness of this technique. However, further investigation with a larger number of specimens is recommended.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Thanks to Tania Collins for demonstrating how to use the GC-MS instrumentation.
Conflicts of Interest
The author declares no conflict of interest.
References
- Schröder, M.L.; Slippers, B.; Wingfield, M.J.; Hurley, B.P. Invasion history and management of Eucalyptus snout beetles in the Gonipterus scutellatus species complex. J. Pest Sci. 2020, 93, 11–25. [Google Scholar] [CrossRef]
- Mapondera, T.S.; Burgess, T.; Matsuki, M.; Oberprieler, R.G. Identification and molecular phylogenetics of the cryptic species of the Gonipterus scutellatus complex (Coleoptera: Curculionidae: Gonipterini). Aust. J. Entomol. 2012, 51, 175–188. [Google Scholar] [CrossRef]
- Wheeler, G.S.; Massey, L.M.; Southwell, I.A. Antipredator Defense of Biological Control Agent Oxyops vitiosa Is Mediated by Plant Volatiles Sequestered from the Host Plant Melaleuca quinquenervia. J. Chem. Ecol. 2002, 28, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Drijfhout, F. A Review of Ant Cuticular Hydrocarbons. J. Chem. Ecol. 2009, 35, 1151. [Google Scholar] [CrossRef] [PubMed]
- Holze, H.; Schrader, L.; Buellesbach, J. Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation. Heredity 2021, 126, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, P.P.; Hartke, J.; Schmitt, T.; Menzel, F.; Feldmeyer, B. Candidate genes involved in cuticular hydrocarbon differentiation between cryptic, parabiotic ant species. G3 2021, 11, jkab078. [Google Scholar] [CrossRef] [PubMed]
- Page, M.; Nelson, L.J.; Haverty, M.I.; Blomquist, G.J. Cuticular Hydrocarbons as Chemotaxonomic Characters for Bark Beetles: Dendroctonus ponderosae, D. jeffreyi, D. brevicomis, and D. frontalis (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 1990, 83, 892–901. [Google Scholar] [CrossRef]
- Jacob, J.; Hanssen, H.-P. Distribution and variability of cuticular hydrocarbons within the Coleoptera. Biochem. Syst. Ecol. 1986, 14, 207–210. [Google Scholar] [CrossRef]
- Souza, N.M.; Schröder, M.L.; Hayes, R.A.; Bello, J.E.; Nahrung, H.F. Cuticular hydrocarbons of Gonipterus weevils: Are there species differences? Chemoecology. 2021, 31, 159–167. [Google Scholar] [CrossRef]
- Lapointe, S.L.; Hunter, W.B.; Alessandro, R.T. Cuticular hydrocarbons on elytra of the Diaprepes root weevil Diaprepes abbreviatus (L.) (Coleoptera: Curculionidae). Agric. For. Entomol. 2004, 6, 251–257. [Google Scholar] [CrossRef]
- Mackley, J.W.; Carlson, D.A.; Butler, J.F. Identification of the cuticular hydrocarbons of the horn fly and assays for attraction. J. Chem. Ecol. 1981, 7, 669–683. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M. The Pherobase: Database of Insect Pheromones and Semiochemicals. Available online: https://www.pherobase.com/ (accessed on 7 July 2021).
- R Core Team. R: A Language and Environment for Statistical Computing (Version 4.0.2); R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
Figure 1.
The five Gonipterini weevil species investigated in this study. Photographs are to scale.

Figure 2.
Scores plot showing the results of PCA performed on the normalized CHC data.

Figure 3.
Hierarchical cluster analysis performed on the normalized CHC data.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Details of the Gonipterini weevil specimens analysed in this study.
| Species | No. Specimens | Length (mm) | Width (mm) |
|---|---|---|---|
| Oxyops fasciculatus | 5 | 6.4 ± 0.4 | 3.4 ± 0.2 |
| Oxyops sp. (undescribed) | 3 | 7.6 ± 0.5 | 4.1 ± 0.3 |
| Gonipterus sp. n. 2 (tentative ID) | 1 | 7.8 | 4.2 |
| Gonipterus cinnamomeus | 3 | 6.6 ± 0.2 | 3.7 ± 0.2 |
| Bryachus squamicollis | 3 | 9.9 ± 0.8 | 5.0 ± 0.5 |
Table 2.
Compounds identified from the Gonipterini specimens using GC-MS. The abundance of each compound was quantified as percentage of the total peak area from the total ion chromatogram for each sample.
Table 2.
Compounds identified from the Gonipterini specimens using GC-MS. The abundance of each compound was quantified as percentage of the total peak area from the total ion chromatogram for each sample.
| No. | Compound | LRI | M+ (m/z) | Ident. ^ | B. squamicollis (n = 3) | G. cinnamomeus (n = 3) | G. sp. n. 2 (n = 1) | O. fasciculatus (n = 5) | Oxyops sp. (n = 3) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 3-hexanone | 787 | 100 | MS, LRI | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.08 | 0.05 ± 0.04 | 0.04 ± 0.00 |
| 2 | 2-hexanone | 791 | 100 | MS, LRI | 0.02 ± 0.02 | 0.07 ± 0.01 | 0.07 | 0.06 ± 0.04 | 0.04 ± 0.01 |
| 3 | 2,4-dimethylheptane | 820 | 128 | MS, LRI | 0.02 ± 0.01 | 0.02 ± 0.02 | 0.05 | 0.03 ± 0.02 | 0.02 ± 0.00 |
| 4 | Heptanal | 902 | 114 | MS, LRI | 0.03 ± 0.01 | 0.03 ± 0.02 | 0 | 0 | 0 |
| 5 | Octanal | 1002 | 128 | MS, LRI | 0.01 ± 0.01 | 0 | 0 | 0 | 0 |
| 6 | Eucalyptol | 1033 | 154 | MS, LRI | 0 | 0 | 0.10 | 0.01 ± 0.02 | 0 |
| 7 | 3,6-dimethyldecane | 1055 | 170 | MS, LRI | 0.03 ± 0.01 | 0.06 ± 0.02 | 0.06 | 0.05 ± 0.03 | 0.03 ± 0.01 |
| 8 | 2,6,8-trimethyldecane | 1099 | 184 | MS, LRI | 0.02 ± 0.02 | 0.02 ± 0.03 | 0 | 0.01 ± 0.03 | 0.01 ± 0.01 |
| 9 | Nonanal | 1104 | 142 | MS, LRI | 0.13 ± 0.05 | 0 | 0 | 0 | 0 |
| 10 | Decanal | 1205 | 156 | MS, LRI | 0.01 ± 0.01 | 0.02 ± 0.04 | 0 | 0 | 0 |
| 11 | Exo-2-hydroxycineole | 1228 | 170 | MS, LRI | 0.04 ± 0.02 | 0.08 ± 0.04 | 0 | 0.02 ± 0.02 | 0 |
| 12 | Tentative: 2,6,10-trimethylundecane | 1275 | 198 | MS, LRI | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.05 | 0.06 ± 0.03 | 0.03 ± 0 |
| 13 | 10-undecenal | 1282 | 168 | MS, LRI | 0.14 ± 0.17 | 0 | 0 | 0 | 0 |
| 14 | Tentative: carvacrol | 1297 | 150 | MS, LRI | 2.42 ± 3.83 | 0.04 ± 0.07 | 0 | 0.02 ± 0.04 | 0 |
| 15 | Tentative: isoascaridole | 1312 | 168 | MS, LRI | 0.01 ± 0.01 | 0 | 0 | 0 | 0 |
| 16 | Tentative: 4a-methyldecahydro-1-naphthalenol | 1319 | 168 | MS, LRI | 0.21 ± 0.23 | 0 | 0 | 0 | 0 |
| 17 | 4,6-dimethyldodecane | 1321 | 198 | MS, LRI | 0.03 ± 0.01 | 0.02 ± 0.03 | 0 | 0.03 ± 0.03 | 0.01 ± 0.02 |
| 18 | Tentative: cis-p-menth-1-en-3,8-diol | 1358 | 170 | MS, LRI | 0.02 ± 0.03 | 0 | 0 | 0 | 0 |
| 19 | (+)-cis,trans-nepetalactone | 1364 | 166 | MS, LRI | 0.02 ± 0.03 | 0 | 0 | 0 | 0 |
| 20 | Dodecanal | 1408 | 184 | MS, LRI | 0 | 0.02 ± 0.03 | 0 | 0.01 ± 0.01 | 0.01 ± 0.02 |
| 21 | Aromadendrene | 1444 | 204 | MS, LRI | 0 | 0 | 0 | 0.01 ± 0.03 | 0 |
| 22 | Unidentified alkane 1 | 1488 | - | MS | 0.01 ± 0.01 | 0.01 ± 0.02 | 0.06 | 0.05 ± 0.02 | 0.03 ± 0.00 |
| 23 | Bicyclogermacrene | 1501 | 204 | MS, LRI | 0.03 ± 0.05 | 0 | 0 | 0.02 ± 0.02 | 0 |
| 24 | Tentative: 2,6,10-trimethyltridecane | 1534 | 226 | MS, LRI | 0.01 ± 0.01 | 0.01 ± 0.02 | 0 | 0 | 0 |
| 25 | Globulol | 1592 | 222 | MS, LRI | 0 | 0 | 0.07 | 0.01 ± 0.03 | 0 |
| 26 | Tetradecanal | 1612 | 212 | MS, LRI | 0.04 ± 0.02 | 0 | 0.05 | 0.01 ± 0.02 | 0.02 ± 0.03 |
| 27 | Heptadecane | 1699 | 240 | MS, LRI | 0.03 ± 0.01 | 0.05 ± 0.01 | 0.05 | 0.04 ± 0.03 | 0.02 ± 0.02 |
| 28 | Phytane | 1743 | 282 | MS, LRI | 0.01 ± 0.02 | 0 | 0 | 0 | 0 |
| 29 | Cis-9-hexadecenal | 1795 | 220 | MS, LRI | 0.05 ± 0.01 | 0 | 0 | 0 | 0 |
| 30 | Hexadecanal | 1816 | 240 | MS, LRI | 0.17 ± 0.05 | 0.09 ± 0.04 | 0.10 | 0.16 ± 0.13 | 0.13 ± 0.08 |
| 31 | 6,10,14-trimethyl-2-pentadecanone | 1840 | 268 | MS, LRI | 0 | 0 | 0 | 0 | 0.01 ± 0.02 |
| 32 | 2-heptadecanone | 1899 | 254 | MS, LRI | 0.09 ± 0.02 | 0.07 ± 0.03 | 0.14 | 0.04 ± 0.05 | 0.06 ± 0.02 |
| 33 | Tentative: 2,2-dimethyloctadecane | 1910 | 282 | MS, LRI | 0.04 ± 0.02 | 0.06 ± 0.06 | 0.09 | 0.03 ± 0.03 | 0.02 ± 0.03 |
| 34 | Heptadecanal | 1918 | 254 | MS, LRI | 0.05 ± 0.01 | 0 | 0 | 0 | 0 |
| 35 | Tentative: 3-ethyl-3-methylheptadecane | 1953 | 283 | MS, LRI | 0.03 ± 0.02 | 0.06 ± 0.02 | 0.05 | 0.06 ± 0.03 | 0.03 ± 0.00 |
| 36 | 9-octadecanone | 1990 | 268 | MS, LRI | 0 | 0 | 0 | 0.1 ± 0.1 | 0 |
| 37 | Cis-13-octadecenal | 1995 | 266 | MS, LRI | 0.12 ± 0.03 | 0 | 0 | 0 | 0 |
| 38 | Tentative: cis-9-octadecenal | 2014 | 266 | MS, LRI | 0 | 0 | 0 | 0.03 ± 0.07 | 0 |
| 39 | Octadecanal | 2019 | 268 | MS, LRI | 0.61 ± 0.14 | 0.07 ± 0.04 | 0.13 | 0.19 ± 0.18 | 0.16 ± 0.13 |
| 40 | Cis-2-octadecen-1-ol acetate | 2074 | 310 | MS, LRI | 0 | 0 | 0 | 0.01 ± 0.01 | 0 |
| 41 | 2-nonadecanone | 2098 | 282 | MS, LRI | 0.09 ± 0.02 | 0.04 ± 0.04 | 0.18 | 0.02 ± 0.05 | 0.08 ± 0.05 |
| 42 | Nonadecanal | 2117 | 282 | MS, LRI | 0.06 ± 0.01 | 0 | 0 | 0 | 0 |
| 43 | Unidentified alkane 2 | 2128 | - | MS | 0.23 ± 0.21 | 0.25 ± 0.43 | 0 | 0.03 ± 0.07 | 0.12 ± 0.22 |
| 44 | Unidentified alkane 3 | 2139 | - | MS | 0 | 0.35 ± 0.60 | 0 | 0 | 0.23 ± 0.40 |
| 45 | Unidentified alkane 4 | 2148 | - | MS | 0.48 ± 0.46 | 0.42 ± 0.73 | 0 | 0.06 ± 0.14 | 0.28 ± 0.48 |
| 46 | Unidentified alkane 5 | 2160 | - | MS | 0.10 ± 0.08 | 0.14 ± 0.24 | 0.07 | 0.04 ± 0.02 | 0.01 ± 0.02 |
| 47 | Unidentified alkane 6 | 2168 | - | MS | 0.08 ± 0.08 | 0.09 ± 0.15 | 0 | 0 | 0.28 ± 0.49 |
| 48 | Docosane | 2197 | 310 | MS, LRI | 0.02 ± 0.02 | 0 | 0 | 0.06 ± 0.05 | 0 |
| 49 | Eicosanal | 2222 | 296 | MS, LRI | 0.09 ± 0.04 | 0 | 0.71 | 0.03 ± 0.07 | 0 |
| 50 | Unidentified alkane 7 a | 2260 | - | MS | 0 | 0 | 0 | 0 | 0.03 ± 0.03 |
| 51 | Tricosane | 2297 | 324 | MS, LRI | 0.74 ± 0.19 | 0.37 ± 0.09 | 0 | 1.35 ± 0.54 | 0.27 ± 0.06 |
| 52 | Unidentified ketone 1 | 2304 | - | MS | 0 | 0 | 0.09 | 0 | 0 |
| 53 | Henicosanal | 2326 | 310 | MS, LRI | 0 | 0 | 0.50 | 0 | 0 |
| 54 | 11-methyltricosane | 2331 | 338 | MS, LRI | 0.20 ± 0.34 | 0 | 0 | 0 | 0.02 ± 0.04 |
| 55 | Unidentified aldehyde | 2367 | - | MS | 0.02 ± 0.01 | 0 | 0 | 0 | 0 |
| 56 | 3-methyltricosane | 2374 | 339 | MS, LRI | 0.02 ± 0.03 | 0 | 0 | 0 | 0 |
| 57 | Tetracosane | 2400 | 338 | MS, LRI | 0.03 ± 0.02 | 0.07 ± 0.01 | 0.16 | 0.09 ± 0.05 | 0.02 ± 0.02 |
| 58 | Docosanal | 2430 | 324 | MS, LRI | 0.01 ± 0.01 | 0.58 ± 0.31 | 1.12 | 0 | 0 |
| 59 | Tentative: 9-methyltetracosane | 2437 | 352 | MS, LRI | 0.01 ± 0.02 | 0 | 0 | 0 | 0 |
| 60 | 2-methyltetracosane | 2473 | 352 | MS, LRI | 0 | 0.94 ± 1.64 | 0 | 0 | 0.69 ± 1.19 |
| 61 | Tentative: x-pentacosene | 2479 | 352 | MS, LRI | 0 | 0 | 0 | 0.03 ± 0.07 | 0 |
| 62 | Pentacosane | 2499 | 352 | MS, LRI | 1.67 ± 0.93 | 1.83 ± 0.37 | 1.82 | 4.26 ± 2.02 | 0.97 ± 0.88 |
| 63 | Unidentified ketone 2 b | 2509 | - | MS | 0 | 0.05 ± 0.09 | 0.13 | 0 | 0 |
| 64 | Tentative: 7-methylpentacosane | 2522 | 367 | MS, LRI | 0 | 0.27 ± 0.46 | 0 | 0.48 ± 0.42 | 0 |
| 65 | Tentative: 11-methylpentacosane | 2530 | 395 | MS, LRI | 0 | 0.15 ± 0.26 | 0 | 0.11 ± 0.14 | 0 |
| 66 | Tentative: 13-methylpentacosane | 2569 | 367 | MS, LRI | 0.03 ± 0.05 | 0 | 0 | 0 | 0 |
| 67 | 3-methylpentacosane | 2574 | 366 | MS, LRI | 0.26 ± 0.23 | 1.09 ± 0.64 | 0.27 | 0 | 0 |
| 68 | Tentative: 11,15-dimethylpentacosane | 2584 | 409 | MS, LRI | 0 | 0 | 0 | 0.70 ± 1.57 | 0 |
| 69 | Hexacosane | 2600 | 366 | MS, LRI | 9.04 ± 6.87 | 13.26 ± 7.6 | 14.70 | 8.30 ± 7.10 | 11.01 ± 8.76 |
| 70 | Tetracosanal | 2637 | 352 | MS, LRI | 1.95 ± 2.56 | 4.22 ± 0.91 | 0 | 0 | 0.16 ± 0.27 |
| 71 | 2-methylhexacosane | 2663 | 380 | MS, LRI | 0 | 0 | 0 | 0.03 ± 0.07 | 0 |
| 72 | Unidentified alkane 8 | 2672 | - | MS | 0 | 0.51 ± 0.46 | 0 | 0 | 0.15 ± 0.15 |
| 73 | Tentative: 13-methylhexacosane | 2682 | 381 | MS, LRI | 0.65 ± 1.12 | 0.02 ± 0.04 | 0 | 0.88 ± 1.37 | 0 |
| 74 | Unidentified alkane 9 | 2690 | - | MS | 0.36 ± 0.62 | 0 | 0 | 0 | 0 |
| 75 | Heptacosane | 2704 | 380 | MS, LRI | 11.32 ± 2.95 | 4.3 ± 4.94 | 2.27 | 1.54 ± 3.44 | 0.36 ± 0.62 |
| 76 | Tentative: 7-methylheptacosane | 2712 | 395 | MS, LRI | 21.49 ± 5.33 | 28.94 ± 7.24 | 25.45 | 27.58 ± 6.29 | 16.77 ± 6.68 |
| 77 | Unidentified ketone 3 | 2723 | - | MS | 0 | 0.75 ± 0.31 | 0 | 0 | 0 |
| 78 | 13-methylheptacosane | 2737 | 394 | MS, LRI | 0.32 ± 0.06 | 0.07 ± 0.13 | 0 | 0.08 ± 0.15 | 0.73 ± 0.24 |
| 79 | Unidentified alkane 10 | 2755 | - | MS | 1.14 ± 1.2 | 0 | 0 | 0.01 ± 0.02 | 0.10 ± 0.17 |
| 80 | Tentative: 11-methylheptacosane | 2759 | 395 | MS, LRI | 0 | 4.60 ± 4.16 | 0 | 1.18 ± 2.49 | 0.14 ± 0.23 |
| 81 | 2-methylheptacosane | 2764 | 394 | MS, LRI | 0 | 0 | 0 | 0 | 0.08 ± 0.07 |
| 82 | Docosyl pentyl ether | 2770 | 396 | MS, LRI | 0 | 0 | 0 | 0 | 0.27 ± 0.47 |
| 83 | 3-methylheptacosane | 2774 | 394 | MS, LRI | 0 | 3.14 ± 1.61 | 2.82 | 0 | 0 |
| 84 | Tentative: 5,15- or 5,17-dimethylheptacosane | 2777 | 409 | MS, LRI | 1.06 ± 1.07 | 0 | 0 | 0 | 0 |
| 85 | Tentative: 5,11-dimethylheptacosane | 2784 | 409 | MS, LRI | 0.36 ± 0.62 | 1.93 ± 3.35 | 0 | 3.93 ± 7.21 | 0.79 ± 1.37 |
| 86 | Octacosane | 2800 | 394 | MS, LRI | 6.89 ± 2.27 | 3.55 ± 0.89 | 19.10 | 4.16 ± 0.85 | 4.43 ± 1.34 |
| 87 | Squalene | 2811 | 384 | MS, LRI | 0.38 ± 0.65 | 0.36 ± 0.37 | 0.71 | 0.43 ± 0.20 | 0.27 ± 0.08 |
| 88 | Tentative: 12-methyloctacosane | 2830 | 409 | MS, LRI | 0.22 ± 0.14 | 0.05 ± 0.08 | 0 | 0 | 0 |
| 89 | Hexacosanal | 2837 | 380 | MS, LRI | 0.94 ± 0.24 | 0.94 ± 0.3 | 0 | 0 | 0 |
| 90 | Tentative: x-methyloctacosane | 2858 | 408 | MS, LRI | 0.05 ± 0.05 | 0 | 0.11 | 0 | 0 |
| 91 | 2-methyloctacosane | 2865 | 408 | MS, LRI | 0.24 ± 0.14 | 0.93 ± 0.22 | 0.96 | 0 | 8.28 ± 6.05 |
| 92 | Nonacosene | 2881 | 407 | MS, LRI | 1.03 ± 0.40 | 0 | 0 | 0 | 0 |
| 93 | 1-hexacosanol | 2890 | 382 | MS, LRI | 0 | 0.21 ± 0.20 | 0 | 0 | 0 |
| 94 | Nonacosane | 2918 | 408 | MS, LRI | 26.54 ± 5.05 | 17.34 ± 3.08 | 24.90 | 42.99 ± 14.09 | 51.41 ± 13.58 |
| 95 | Triacontane | 2982 | 422 | MS, LRI | 5.72 ± 0.63 | 7.31 ± 3.07 | 2.79 | 0.51 ± 0.37 | 1.38 ± 0.68 |
| 96 | Tentative: x,12-dimethylnonacosane | 3002 | 437 | MS, LRI | 0.30 ± 0.50 | 0 | 0 | 0.01 ± 0.02 | 0 |
| 97 | Tentative: 2-methyltriacontane | 3039 | 437 | MS, LRI | 1.38 ± 0.43 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Johnson, J.B. Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae). Biol. Life Sci. Forum 2022, 15, 5. https://doi.org/10.3390/IECD2022-12383
AMA Style
Johnson JB. Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae). Biology and Life Sciences Forum. 2022; 15(1):5. https://doi.org/10.3390/IECD2022-12383
Chicago/Turabian StyleJohnson, Joel B. 2022. "Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae)" Biology and Life Sciences Forum 15, no. 1: 5. https://doi.org/10.3390/IECD2022-12383