
Micropropagation of Vaccinium corymbosum L.: An Alternative Procedure for the Production of Secondary Metabolites

 ,
,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Shoot Proliferation and Biomass Production in Solid and TIS Culture Systems
2.2. Ultrasound-Assisted Extraction of Total Phenolic Compounds
2.3. Quantitative and Qualitative Analysis of Total Phenolic Compounds by HPLC-DAD-ESI-MS
2.4. Extraction of Carotenoid and Chlorophyll Compounds
2.5. Quantitative and Qualitative Analysis of Carotenoids and Chlorophylls by HPLC-DAD-ESI-MS
2.6. Data Analysis
3. Results and Discussions
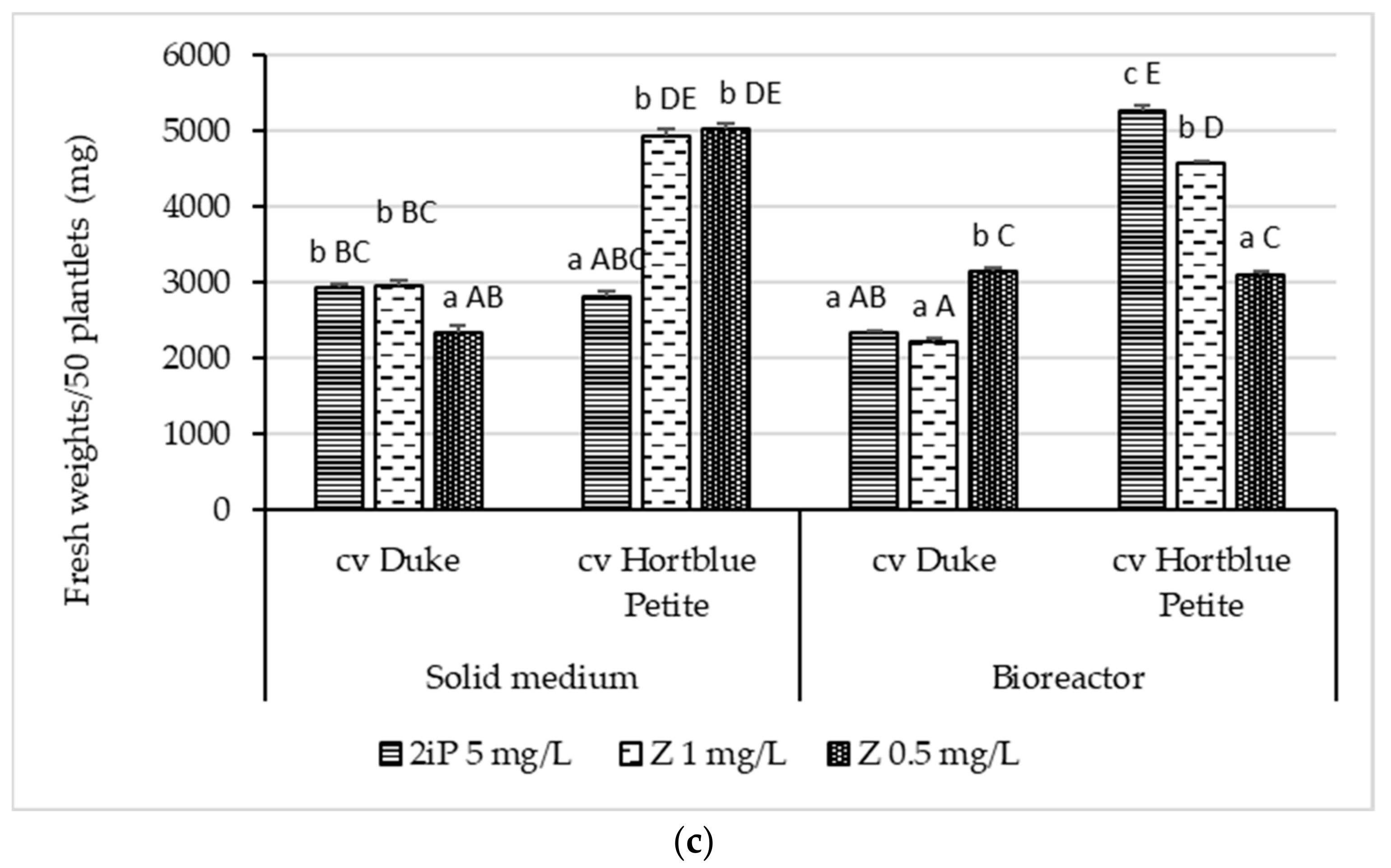
3.1. Shoot Proliferation and Biomass Production (Shoots) in Solid and TIS Culture Systems
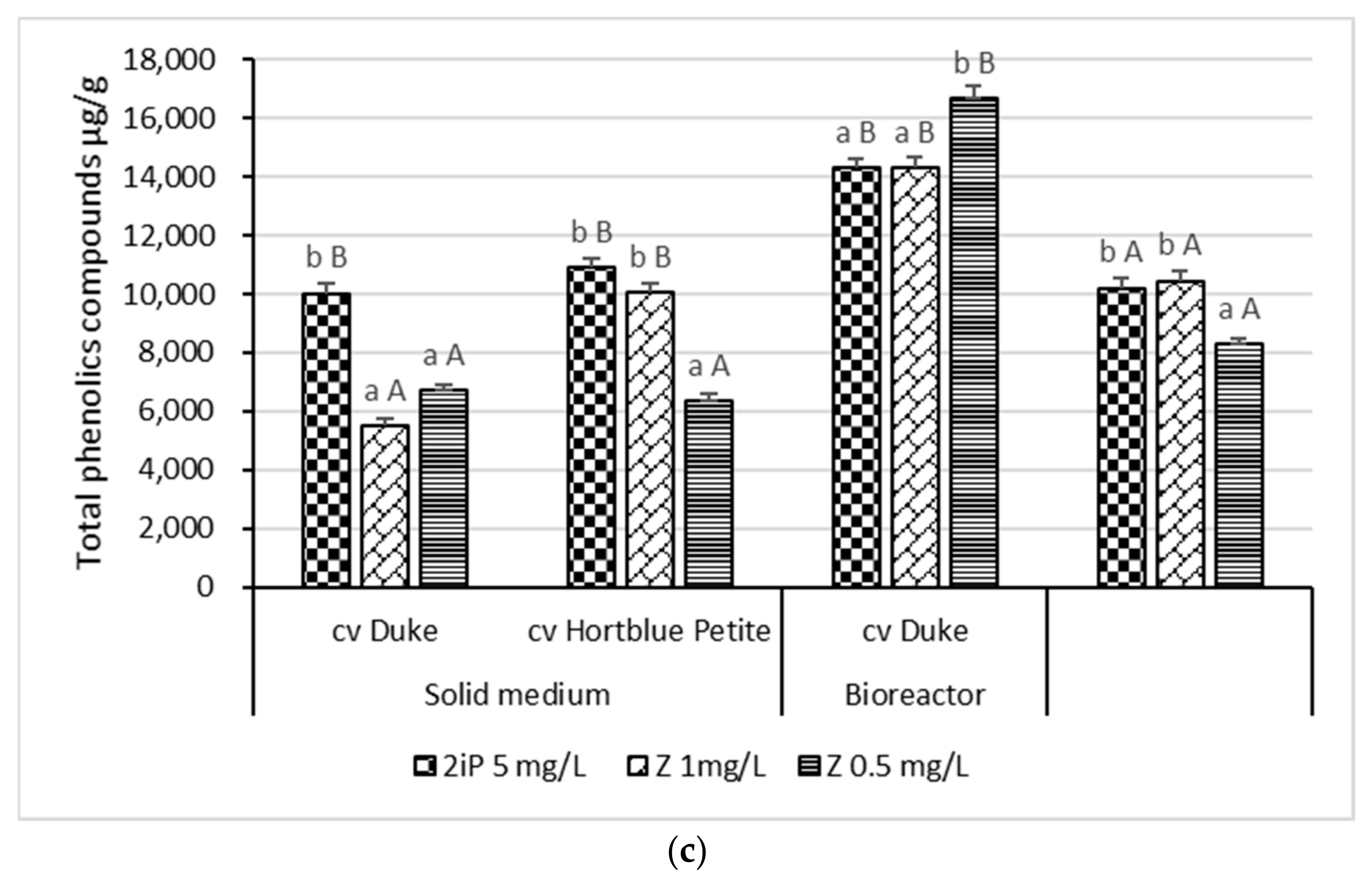
3.2. Content of Phenolic Compounds
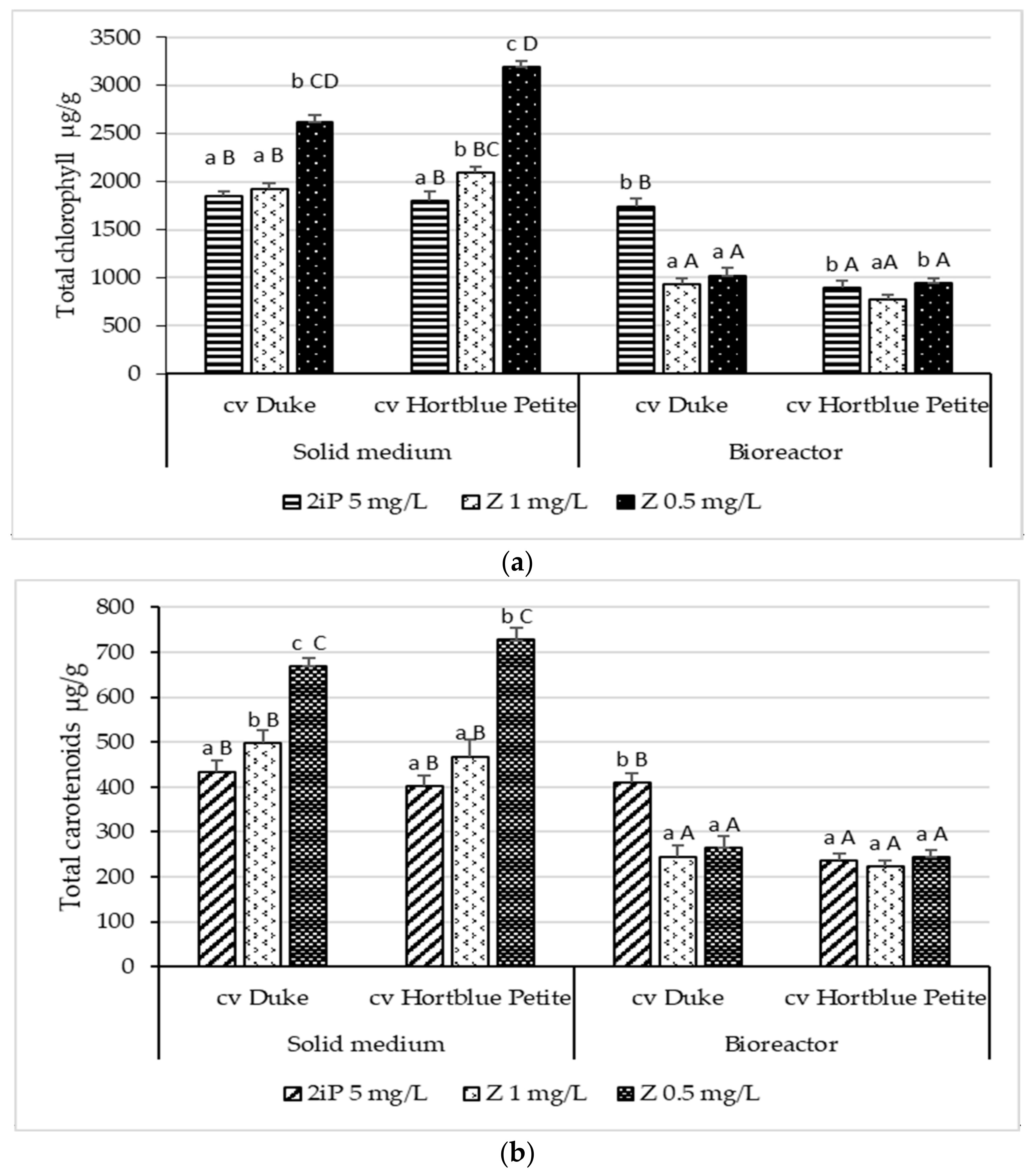
3.3. Content of Carotenoids and Chlorophylls
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ștefănescu, B.-E.; Călinoiu, L.F.; Ranga, F.; Fetea, F.; Mocan, A.; Vodnar, D.C.; Crișan, G. Chemical Composition and Biological Activities of the Nord-West Romanian Wild Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinium vitis-idaea L.) Leaves. Antioxidants 2020, 9, 495. [Google Scholar] [CrossRef] [PubMed]
- Ștefănescu, B.-E.; Călinoiu, L.F.; Ranga, F.; Fetea, F.; Mocan, A.; Vodnar, D.C.; Crișan, G. The Chemical and Biological Profiles of Leaves from Commercial Blueberry Varieties. Plants 2020, 9, 1193. [Google Scholar] [CrossRef] [PubMed]
- Ștefănescu, B.E.; Szabo, K.; Mocan, A.; Crişan, G. Phenolic Compounds from Five Ericaceae Species Leaves and Their Related Bioavailability and Health Benefits. Molecules 2019, 24, 2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, J.C.; de Oliveira, M.E.B.; Cardoso, F.D.C. Advances and Challenges on the in vitro Production of Secondary Metabolites from Medicinal Plants. Hortic. Bras. 2019, 37, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Martău, G.A.; Călinoiu, L.-F.; Vodnar, D.C. Bio-Vanillin: Towards a Sustainable Industrial Production. Trends Food Sci. Technol. 2021, 109, 579–592. [Google Scholar] [CrossRef]
- Ferlemi, A.-V.; Lamari, F.N. Berry Leaves: An Alternative Source of Bioactive Natural Products of Nutritional and Medicinal Value. Antioxidants 2016, 5, 17. [Google Scholar] [CrossRef]
- Wang, T.; Guo, N.; Wang, S.-X.; Kou, P.; Zhao, C.-J.; Fu, Y.-J. Ultrasound-Negative Pressure Cavitation Extraction of Phenolic Compounds from Blueberry Leaves and Evaluation of Its DPPH Radical Scavenging Activity. Food Bioprod. Process. 2018, 108, 69–80. [Google Scholar] [CrossRef]
- Ferlemi, A.-V.; Makri, O.E.; Mermigki, P.G.; Lamari, F.N.; Georgakopoulos, C.D. Quercetin Glycosides and Chlorogenic Acid in Highbush Blueberry Leaf Decoction Prevent Cataractogenesis in vivo and in vitro: Investigation of the Effect on Calpains, Antioxidant and Metal Chelating Properties. Exp. Eye Res. 2016, 145, 258–268. [Google Scholar] [CrossRef]
- Pervin, M.; Hasnat, M.A.; Lim, B.O. Antibacterial and Antioxidant Activities of Vaccinium corymbosum L. Leaf Extract. Asian Pac. J. Trop. Dis. 2013, 3, 444–453. [Google Scholar] [CrossRef]
- Ribera-Fonseca, A.; Jiménez, D.; Leal, P.; Riquelme, I.; Roa, J.C.; Alberdi, M.; Peek, R.M.; Reyes-Díaz, M. The Anti-Proliferative and Anti-Invasive Effect of Leaf Extracts of Blueberry Plants Treated with Methyl Jasmonate on Human Gastric Cancer In Vitro Is Related to Their Antioxidant Properties. Antioxidants 2020, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Vodnar, D.-C.; Mitrea, L.; Teleky, B.-E.; Szabo, K.; Călinoiu, L.-F.; Nemeş, S.-A.; Martău, G.-A. Coronavirus Disease (COVID-19) Caused by (SARS-CoV-2) Infections: A Real Challenge for Human Gut Microbiota. Front. Cell. Infect. Microbiol. 2020, 10, 786. [Google Scholar] [CrossRef] [PubMed]
- Debnath, S.C. Propagation of Vaccinium in Vitro. Int. J. Fruit Sci. 2007, 6, 47–71. [Google Scholar] [CrossRef]
- Harris, C.S.; Burt, A.J.; Saleem, A.; Le, P.M.; Martineau, L.C.; Haddad, P.S.; Bennett, S.A.L.; Arnason, J.T. A Single HPLC-PAD-APCI/MS Method for the Quantitative Comparison of Phenolic Compounds Found in Leaf, Stem, Root and Fruit Extracts of Vaccinium Angustifolium. Phytochem. Anal. 2007, 18, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Maya-Cano, D.A.; Arango-Varela, S.; Santa-Gonzalez, G.A. Phenolic Compounds of Blueberries (Vaccinium Spp) as a Protective Strategy against Skin Cell Damage Induced by ROS: A Review of Antioxidant Potential and Antiproliferative Capacity. Heliyon 2021, 7, e06297. [Google Scholar] [CrossRef] [PubMed]
- Ramata-Stunda, A.; Valkovska, V.; Borodušķis, M.; Livkiša, D.; Kaktiņa, E.; Silamiķele, B.; Borodušķe, A.; Pentjušs, A.; Rostoks, N. Development of Metabolic Engineering Approaches to Regulate the Content of Total Phenolics, Antiradical Activity and Organic Acids in Callus Cultures of the Highbush Blueberry (Vaccinium corymbosum L.). Agron. Res. 2020, 18, 1860–1872. [Google Scholar] [CrossRef]
- Ștefănescu, B.E.; Nemes, S.-A.; Teleky, B.-E.; Călinoiu, L.F.; Mitrea, L.; Martău, G.A.; Szabo, K.; Mihai, M.; Vodnar, D.C.; Crișan, G. Microencapsulation and Bioaccessibility of Phenolic Compounds of Vaccinium Leaf Extracts. Antioxidants 2022, 11, 674. [Google Scholar] [CrossRef]
- Vodnar, D.C.; Călinoiu, L.F.; Mitrea, L.; Precup, G.; Bindea, M.; Păcurar, A.M.; Szabo, K.; Ştefănescu, B.E. 15-A New Generation of Probiotic Functional Beverages Using Bioactive Compounds From Agro-Industrial Waste. In Functional and Medicinal Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 483–528. ISBN 978-0-12-816397-9. [Google Scholar]
- Bakhshipour, M.; Mafakheri, M.; Kordrostami, M.; Zakir, A.; Rahimi, N.; Feizi, F.; Mohseni, M. In Vitro Multiplication, Genetic Fidelity and Phytochemical Potentials of Vaccinium arctostaphylos L.: An Endangered Medicinal Plant. Ind. Crops Prod. 2019, 141, 111812. [Google Scholar] [CrossRef]
- Yue, W.; Ming, Q.; Lin, B.; Rahman, K.; Zheng, C.-J.; Han, T.; Qin, L. Medicinal Plant Cell Suspension Cultures: Pharmaceutical Applications and High-Yielding Strategies for the Desired Secondary Metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef]
- Rao, S.R.; Ravishankar, G.A. Plant Cell Cultures: Chemical Factories of Secondary Metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Debnath, M.; Malik, C.P.; Bisen, P.S. Micropropagation: A Tool for the Production of High Quality Plant-Based Medicines. Curr. Pharm. Biotechnol. 2006, 7, 33–49. [Google Scholar] [CrossRef] [Green Version]
- Ekiert, H.; Szewczyk, A.; Kuś, A. Free Phenolic Acids in Ruta graveolens L. in Vitro Culture. Die Pharm.-Int. J. Pharm. Sci. 2009, 64, 692–694. [Google Scholar] [CrossRef]
- Szopa, A.; Ekiert, H. In Vitro Cultures of Schisandra Chinensis (Turcz.) Baill. (Chinese Magnolia Vine)—A Potential Biotechnological Rich Source of Therapeutically Important Phenolic Acids. Appl. Biochem. Biotechnol. 2012, 166, 1941–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiem, B.; Kikowska, M.; Krawczyk, A.; Więckowska, B.; Sliwinska, E. Phenolic Acid and DNA Contents of Micropropagated Eryngium planum L. Plant Cell Tiss Organ Cult. 2013, 114, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Szopa, A.; Ekiert, H. Production of Biologically Active Phenolic Acids in Aronia Melanocarpa (Michx.) Elliott in Vitro Cultures Cultivated on Different Variants of the Murashige and Skoog Medium. Plant Growth Regul. 2014, 72, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Piątczak, E.; Grzegorczyk-Karolak, I.; Wysokińska, H. Micropropagation of Rehmannia Glutinosa Libosch.: Production of Phenolics and Flavonoids and Evaluation of Antioxidant Activity. Acta Physiol. Plant. 2014, 36, 1693–1702. [Google Scholar] [CrossRef] [Green Version]
- Mamdouh, D.; Mahgoub, H.A.M.; Gabr, A.M.M.; Ewais, E.A.; Smetanska, I. Genetic Stability, Phenolic, Flavonoid, Ferulic Acid Contents, and Antioxidant Activity of Micropropagated Lycium Schweinfurthii Plants. Plants 2021, 10, 2089. [Google Scholar] [CrossRef]
- Park, H.-Y.; Kim, K.-S.; Ak, G.; Zengin, G.; Cziáky, Z.; Jekő, J.; Adaikalam, K.; Song, K.; Kim, D.-H.; Sivanesan, I. Establishment of a Rapid Micropropagation System for Kaempferia Parviflora Wall. Ex Baker: Phytochemical Analysis of Leaf Extracts and Evaluation of Biological Activities. Plants 2021, 10, 698. [Google Scholar] [CrossRef]
- Twardovska, M.O.; Konvalyuk, I.I.; Lystvan, K.V.; Andreev, I.O.; Kunakh, V.A. THE CONTENT OF PHENOLIC COMPOUNDS AND FLAVONOIDS IN Deschampsia Antarctica TISSUE CULTURE. Biotechnol. Acta 2021, 14, 59–66. [Google Scholar] [CrossRef]
- Kikowska, M.; Chanaj-Kaczmarek, J.; Derda, M.; Budzianowska, A.; Thiem, B.; Ekiert, H.; Szopa, A. The Evaluation of Phenolic Acids and Flavonoids Content and Antiprotozoal Activity of Eryngium Species Biomass Produced by Biotechnological Methods. Molecules 2022, 27, 363. [Google Scholar] [CrossRef]
- Santos-Tierno, R.; Garcia, R.; Fonseca, E.; Faleiro, F.; Moreira, D.; Pacheco, G.; Mansur, E. Flavonoid Content and Antioxidant Potential of Leaf Extracts of Passiflora Setacea Cv BRS Prola Do Cerrado, a New Wild Passion Fruit Cultivar. JMPR 2022, 16, 26–34. [Google Scholar] [CrossRef]
- Ružić, D.; Vujović, T.; Libiakova, G.; Cerović, R.; Gajdošova, A. Micropropagation in Vitro of Highbush Blueberry (Vaccinium corymbosum L.). J. Berry Res. 2012, 2, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Vescan, L.; Pamfil, D.; Clapa, D.; Fira, A.; Sisea, C.; Pop, I.; Petricele, I.; Ciuzan, O.; Pop, R. Efficient Micropropagation Protocol for Highbush Blueberry (Vaccinium corymbosum L.) Cv. “Elliot.”. Rom. Biotechnol. Lett. 2012, 17, 6893–6902. [Google Scholar]
- Borsai, O.; Hârța, M.; Pamfil, D.; Clapa, D. The Effect of Cytokinins on Micropropagation Success of Highbush Blueberry (Vaccinium corymbosum L.). Agricultura 2019, 111, 93–100. [Google Scholar] [CrossRef]
- Debnath, S.C.; Arigundam, U. In Vitro Propagation Strategies of Medicinally Important Berry Crop, Lingonberry (Vaccinium Vitis-Idaea L.). Agronomy 2020, 10, 744. [Google Scholar] [CrossRef]
- Georgieva, L.M.; Kondakova, V. In Vitro Propagation of Vaccinium corymbosum L. Available online: https://www.semanticscholar.org/paper/In-vitro-propagation-of-Vaccinium-corymbosum-L-L.-eorgieva/72d1a184be09813f1b35f44915a165f29ed8ee79 (accessed on 14 April 2022).
- Gajdosova, A.; Ostrolucka, M.G.; Libiakova, G.; Ondruskova, E.; Simala, D. Microclonal Propagation of Vaccinium Sp. and Rubus Sp. and Detection of Genetic Variability in Culture in Vitro. J. Fruit Ornam. Plant Res. 2006, 14, 1–5. [Google Scholar]
- Tetsumura, T.; Matsumoto, Y.; Sato, M.; Honsho, C.; Yamashita, K.; Komatsu, H.; Sugimoto, Y.; Kunitake, H. Evaluation of Basal Media for Micropropagation of Four Highbush Blueberry Cultivars. Sci. Hortic. 2008, 119, 72–74. [Google Scholar] [CrossRef]
- Sedlak, J.; Paprstein, F. In vitro multiplication of highbush blueberry (Vaccinium corymbosum l.) Cultivars. Acta Hortic. 2009, 810, 575–580. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially-Feasible Micropropagation of Mountain Laurel, Kalmia Latifolia, by Use of Shoot-Tip Culture. Comb. Proc. Int. Plant Propagators Soc. 1980, 30, 421–427. [Google Scholar]
- A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures-Murashige-1962-Physiologia Plantarum-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/10.1111/j.1399-3054.1962.tb08052.x (accessed on 29 March 2022).
- Anderson, W.C. Tissue culture propagation of red and black raspberries, rubus idaeus and r. Occidentalis. Acta Hortic. 1980, 112, 13–20. [Google Scholar] [CrossRef]
- Reed, B.M.; Abdelnour-Esquivel, A. The Use of Zeatin to Initiate in Vitro Cultures of Vaccinium Species and Cultivars. HortScience 1991, 26, 3. Available online: https://journals.ashs.org/hortsci/view/journals/hortsci/26/10/article-p1320.xml (accessed on 14 April 2022). [CrossRef] [Green Version]
- Ostrolucka, M.G.; Libiakova, G.; Ondruskova, E.; Gajdosova, A. In Vitro Propagation of Vaccinium Species. Acta Univ. Latv. Biol. 2005, 676, 207–212. [Google Scholar]
- Wang, Y.; Dong, X.; Huang, H.-Y.; Wang, Y.-Z.; Wang, Y.; Dong, X.; Huang, H.-Y.; Wang, Y.-Z. Establishment of Efficient Adventitious Shoots Induction System and Ex Vitro Rooting in Vaccinium Corymbosum (Ericaceae). Bot. Sci. 2019, 97, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Clapa, D.; Hârța, M.; Borsai, O.; Pamfil, D. Micropropagation of Vaccinium corymbosum L. and Corylus avellana L. Using a Temporary Immersion Bioreactor System. Agric.-Rev. Știință Pract. Agric. 2019, 28, 101–108. [Google Scholar]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.-H.; Toşa, M.I. Total Phenolic Contents, Antioxidant Activities, and Lipid Fractions from Berry Pomaces Obtained by Solid-State Fermentation of Two Sambucus Species with Aspergillus Niger. J. Agric. Food Chem. 2015, 63, 3489–3500. [Google Scholar] [CrossRef]
- Bunea, A.; Rugină, D.; Sconţa, Z.; Pop, R.M.; Pintea, A.; Socaciu, C.; Tăbăran, F.; Grootaert, C.; Struijs, K.; VanCamp, J. Anthocyanin Determination in Blueberry Extracts from Various Cultivars and Their Antiproliferative and Apoptotic Properties in B16-F10 Metastatic Murine Melanoma Cells. Phytochemistry 2013, 95, 436–444. [Google Scholar] [CrossRef]
- Parlog, R.; Vodnar, D.; Francisc, D.; Leopold, L.; Socaciu, C. HPLC-PDA and UV-VIS Spectrometry Analysis Used to Fingerprint Sea Buckthorn (Hippophae rhamnoides L.) Berries Comparatively with Leaves and Seeds Extracts. Bull. UASVM Agric. 2009, 66, 409–414. [Google Scholar] [CrossRef]
- Pintea, A.; Varga, A.; Stepnowski, P.; Socaciu, C.; Culea, M.; Diehl, H.A. Chromatographic Analysis of Carotenol Fatty Acid Esters in Physalis Alkekengi and Hippophae Rhamnoides. Phytochem. Anal. 2005, 16, 188–195. [Google Scholar] [CrossRef]
- Molnar, S.; Clapa, D.; Mitre, V. Response of the Five Highbush Blueberry Cultivars to In Vitro Induced Drought Stress by Polyethylene Glycol. Agronomy 2022, 12, 732. [Google Scholar] [CrossRef]
- Arencibia, A.D.; Gómez, A.; Mora, P.; Orellana, F.; Alarcón, J.E. Photomixotrophic Cultures of Blueberries (Vaccinium corymbosum) Accumulate or Release Phenylpropanoids via Inductive Treatments. Acta Physiol. Plant. 2018, 40, 1–5. [Google Scholar] [CrossRef]
- Nielsen, E.; Temporiti, M.E.E.; Cella, R. Improvement of Phytochemical Production by Plant Cells and Organ Culture and by Genetic Engineering. Plant Cell Rep. 2019, 38, 1199–1215. [Google Scholar] [CrossRef]
- Wang, L.-J.; Wu, J.; Wang, H.-X.; Li, S.-S.; Zheng, X.-C.; Du, H.; Xu, Y.-J.; Wang, L.-S. Composition of Phenolic Compounds and Antioxidant Activity in the Leaves of Blueberry Cultivars. J. Funct. Foods 2015, 16, 295–304. [Google Scholar] [CrossRef]
- Wang, H.; Guo, X.; Hu, X.; Li, T.; Fu, X.; Liu, R.H. Comparison of Phytochemical Profiles, Antioxidant and Cellular Antioxidant Activities of Different Varieties of Blueberry (Vaccinium Spp.). Food Chem. 2017, 217, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Arencibia, A.D.; Vergara, C.; Quiroz, K.; Carrasco, B.; Bravo, C.; Garcia-Gonzales, R. An Approach for Micropropagation of Blueberry (Vaccinium corymbosum L.) Plants Mediated by Temporary Immersion Bioreactors (TIBs). Am. J. Plant Sci. 2013, 4, 1022–1028. [Google Scholar] [CrossRef] [Green Version]
- Arencibia, A.D.; Bernal, A.; Yang, L.; Cortegaza, L.; Carmona, E.R.; Pérez, A.; Hu, C.-J.; Li, Y.-R.; Zayas, C.M.; Santana, I. New Role of Phenylpropanoid Compounds during Sugarcane Micropropagation in Temporary Immersion Bioreactors (TIBs). Plant Sci. 2008, 175, 487–496. [Google Scholar] [CrossRef]
- Bernal, A.; Machado, P.; Cortegaza, L.; Carmona, E.R.; Rivero, O.; Zayas, C.M.; Nodarse, O.; Perez, A.; Santana, I.; Arencibia, A.D. Priming and Biopriming Integrated into the Sugarcane Micropropagation Technology by Temporary Immersion Bioreactors (TIBS). Sugar Tech 2008, 10, 42–47. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Quinic Acid | Neochlorogenic Acid | Chloroge Nic Acid | Catechin | Epicate Chin | Caffeic Acid | ECG | Feruroylquinic Acid | Quercetin Glucoside | Kaemferol Glucoside | Quercetin |
|---|---|---|---|---|---|---|---|---|---|---|---|
| DK 0.5 mg/L Z agar | 1382.466 ± 2.63 b | 35.528 ± 0.34 a | 300.148 ± 1.54 a | 1005.697 ± 8.54 c | 869.839 ± 2.01 b | 371.429 ± 2.15 a | 2237.107 ± 1.78 b | 331.029 ± 2.88 a | 176.430 ± 1.58 a | 0.000 | 1.404 ± 0.11 a |
| DK 1 mg/L Z agar | 978.196 ± 3.42 c | 27.499 ± 0.27 b | 255.332 ± 1.38 c | 1098.291 ± 8.39 b | 625.820 ± 1.69 c | 312.996 ± 1.51 c | 1920.981 ± 1.73 c | 187.467 ± 1.63 c | 98.994 ±1.14 b | 0.000 | 0.151 ± 0.01 c |
| DK 5 mg/L 2iP agar | 1543.237 ± 4.01 a | 34.533 ± 0.31 a | 281.310 ± 1.41 b | 2246.689 ± 8.12 a | 1308.753 ± 0.52 a | 332.997 ± 2.26 b | 3866.141 ± 3.86 a | 319.550 ± 1.88 b | 93.055 ± 1.09 b | 0.000 | 1.081 ± 0.09 b |
| DK 0.5 mg/L Z TIS | 2640.141 ± 1.53 a | 149.138 ± 1.29 a | 1020.510 ± 1.09 a | 3347.069 ± 3.41 a | 1721.375 ± 2.27 a | 752.859 ± 3.98 a | 5324.222 ± 0.53 a | 1553.827 ± 2.19 c | 152.952 ± 0.92 b | 0.000 | 3.523 ± 0.29 b |
| DK 1 mg/L Z TIS | 2330.046 ± 1.21 b | 132.169 ± 1.33 a | 918.432 ± 7.81 c | 2808.755 ± 3.65 b | 1694.765 ± 2.13 a | 438.434 ± 2.12 c | 4196.936 ± 0.88 b | 1652.177 ± 1.98 b | 142.669 ± 0.86 b | 0.000 | 3.028 ± 0.25 b |
| DK 5 mg/L 2iP TIS | 2213.671 ± 1.63 c | 112.085 ± 1.24 b | 952.229 ± 1.14 b | 2734.314 ± 3.52 b | 1431.193 ± 1.87 b | 524.608 ± 2.71 b | 4107.055 ± 0.71 b | 2050.486 ± 3.08 a | 164.153 ±1.01 a | 0.000 | 4.509 ± 0.37 a |
| HP 0.5 mg/L Z agar | 642.250 ± 1.99 c | 93.707 ± 0.38 a | 545.274 ± 1.68 a | 881.729 ± 3.13 c | 600.083 ± 1.51 c | 413.909 ± 2.62 a | 1678.137 ± 2.28 b | 1051.008 ± 2.16 a | 249.367 ± 0.22 b | 158.447 ± 1.28 a | 56.438 ± 0.31 b |
| HP 1 mg/L Z agar | 998.923 ± 2.45 b | 47.324 ± 0.34 c | 212.932 ± 0.51 b | 2383.382 ± 1.39 a | 1682.349 ± 3.16 b | 311.495 ± 0.63 b | 3185.247 ± 1.91 a | 737.267 ± 1.83 c | 302.407 ± 0.34 a | 122.569± 1.12 b | 56.768 ± 0.24 b |
| HP 5 mg/L 2iP agar | 1300.584 ± 3.31 a | 75.218 ± 0.26 b | 183.185 ± 0.65 c | 2188.658 ± 1.11 b | 2207.689 ± 1.29 a | 292.779 ± 0.49 b | 3168.711 ± 2.01 a | 968.351 ± 2.31 b | 294.522 ± 0.31 a | 103.902 ± 1.33 c | 106.941 ± 0.22 a |
| HP 0.5 mg/L Z TIS | 842.677 ± 2.09 c | 111.915 ± 1.27 a | 386.031 ± 0.95 b | 1655.987 ± 2.12 c | 1512.319 ± 2.04 c | 261.306 ± 1.01 b | 2835.774 ± 1.96 c | 399.163 ± 0.82 b | 196.743 ± 1.14 b | 41.275 ± 0.33 c | 27.170 ± 0.25 a |
| HP 1 mg/L Z TIS | 1166.982 ± 1.39 b | 119.642 ± 0.71 a | 432.270 ± 0.74 a | 2157.515 ± 1.85 b | 1955.351 ± 1.26 a | 309.360 ± 0.63 a | 3455.840 ± 2.68 a | 565.243 ± 1.06 a | 228.391 ± 1.16 a | 53.644 ± 0.32 a | 4.412 ± 0.35 b |
| HP 5 mg/L 2iP TIS | 1369.291 ± 1.21 a | 75.055 ± 0.46 b | 285.923 ± 1.52 c | 2231.319 ± 1.36 a | 1896.649 ± 1.49 b | 277.908 ± 1.53 b | 3213.485 ± 2.41 b | 580.403 ± 1.22 a | 227.659 ± 1.14 a | 45.010 ± 0.31 b | 4.513 ± 0.38 b |
| Sample | Chlorophyll b | Chlorophyll a | Pheophytin b | Pheophytin a | Lutein | Zeaxanthin | βCarotene |
|---|---|---|---|---|---|---|---|
| DK 0.5 mg/L Z agar | 644.062 ± 1.86 a | 1477.052 ± 4.46 a | 11.911 ± 0.34 a | 491.115 ± 1.09 a | 188.620 ± 0.68 a | 219.501 ± 1.45 a | 260.840 ± 1.43 a |
| DK 1 mg/L Z agar | 469.433 ± 1.68 b | 1067.194 ± 3.04 b | 7.356 ± 0.32 c | 377.915 ± 1.4 b | 139.483 ± 1.25 b | 163.189 ± 0.72 b | 195.700 ± 0.84 b |
| DK 5 mg/L 2iP agar | 459.969 ± 1.24 b | 1001.148 ± 3.02 c | 7.684 ± 0.35 b | 376.419 ± 1.17 b | 121.156 ± 0.91 c | 143.927 ± 0.88 c | 168.881 ± 1.13 c |
| DK 0.5 mg/L Z TIS | 259.848 ± 1.77 b | 451.440 ± 1.48 b | 6.376 ± 0.3 b | 305.701 ± 1.84 b | 71.805 ± 0.51 b | 92.652 ± 0.63 b | 100.477 ± 0.71 b |
| DK 1 mg/L Z TIS | 238.211 ± 1.36 c | 458.142 ± 1.36 b | 5.319 ± 0.25 c | 231.642 ± 1.23 c | 63.867 ± 0.37 c | 85.171 ± 0.56 c | 96.221 ± 0.62 b |
| DK 5 mg/L 2iP TIS | 411.889 ± 1.22 a | 775.069 ± 1.74 a | 12.515 ± 0.44 a | 543.771 ± 1.64 a | 111.326 ± 1.14 a | 138.635 ± 0.92 a | 160.107 ± 1.1 a |
| HP 0.5 mg/L Z agar | 746.370 ± 1.91 a | 1325.398 ± 5.32 a | 22.775 ± 0.19 a | 1097.240 ± 1.74 a | 224.295 ± 1.68 a | 256.210 ± 1.44 a | 247.204 ± 1.42 a |
| HP 1 mg/L Z agar | 508.559 ± 1.63 b | 983.638 ± 5.12 b | 11.835 ± 0.46 b | 584.039 ± 2.14 b | 127.022 ± 1.05 b | 157.510± 1.09 b | 183.069 ± 1.46 b |
| HP 5 mg/L 2iP agar | 416.239 ± 1.37 c | 810.501 ± 4.59 c | 10.707 ± 0.39 b | 563.313 ± 3.49 c | 103.812 ± 0.73 c | 131.853 ± 0.81 c | 166.514± 1.31 c |
| HP 0.5 mg/L Z TIS | 229.551 ± 1.71 a | 526.567 ± 2.36 b | 6.565 ± 0.23 a | 131.904 ± 0.68 b | 71.208 ± 0.12 a | 83.581 ± 0.11 a | 89.815 ± 0.17 b |
| HP 1 mg/L Z TIS | 205.921 ± 1.78 c | 479.281 ± 2.87 c | 5.776 ± 0.19 b | 81.926 ± 0.17 c | 64.666 ± 0.13 b | 77.565 ± 0.1 b | 80.958 ± 0.25 c |
| HP 5 mg/L 2iP TIS | 255.609 ± 1.54 b | 536.385 ± 2.2 a | 6.960 ± 0.21 a | 140.627 ± 0.4 a | 65.755 ± 0.07 b | 78.300 ± 0.16 b | 91.844 ± 0.28 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clapa, D.; Nemeș, S.-A.; Ranga, F.; Hârța, M.; Vodnar, D.-C.; Călinoiu, L.-F. Micropropagation of Vaccinium corymbosum L.: An Alternative Procedure for the Production of Secondary Metabolites. Horticulturae 2022, 8, 480. https://doi.org/10.3390/horticulturae8060480
Clapa D, Nemeș S-A, Ranga F, Hârța M, Vodnar D-C, Călinoiu L-F. Micropropagation of Vaccinium corymbosum L.: An Alternative Procedure for the Production of Secondary Metabolites. Horticulturae. 2022; 8(6):480. https://doi.org/10.3390/horticulturae8060480
Chicago/Turabian StyleClapa, Doina, Silvia-Amalia Nemeș, Floricuța Ranga, Monica Hârța, Dan-Cristian Vodnar, and Lavinia-Florina Călinoiu. 2022. "Micropropagation of Vaccinium corymbosum L.: An Alternative Procedure for the Production of Secondary Metabolites" Horticulturae 8, no. 6: 480. https://doi.org/10.3390/horticulturae8060480