

Single Nucleotide Polymorphisms Associated with AA-Amyloidosis in Siamese and Oriental Shorthair Cats
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals
2.3. Histological and Immunohistochemical Examination
2.4. Genotyping on the Illumina Infinium Feline 63 K iSelect DNA Array
2.5. Genome-Wide Association Studies
2.6. Putative Candidate Gene Identification
3. Results
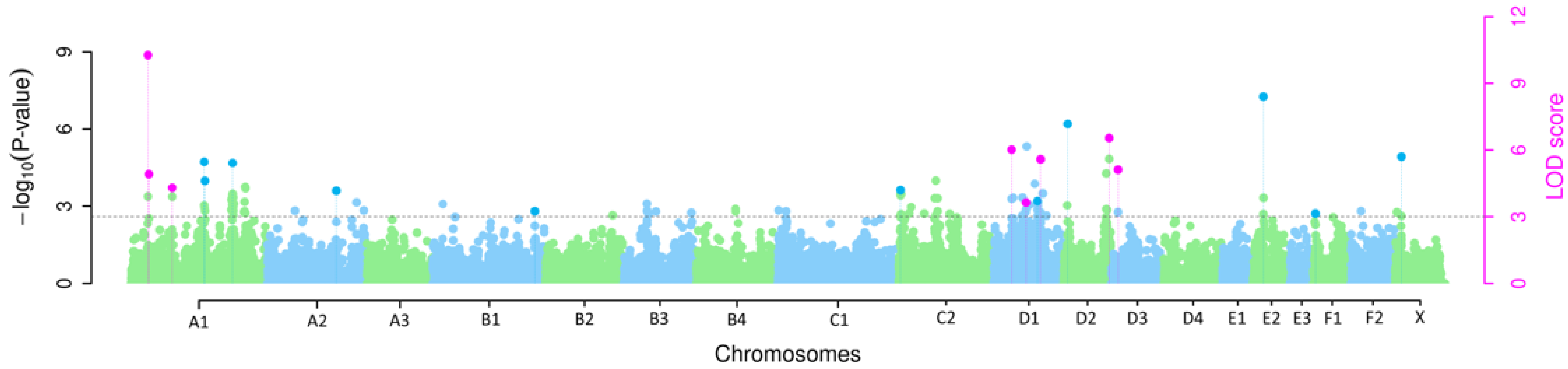
3.1. Multi-Locus GWAS
3.2. Candidate Gene Search
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kumar, V.; Abbas, A.; Fausto, N.; Aster, J. Diseases of the Immune System, Robbins and Cotran Pathologic Basis of Disease; Saunders Elsevier: Philadelphia, PA, USA, 2010; Volume 8, pp. 249–255. [Google Scholar]
- Glenner, G.G.; Eanes, E.D.; Page, D.L. The relation of the properties of Congo red-stained amyloid fibrils to the -conformation. J. Histochem. Cytochem. 1972, 20, 821–826. [Google Scholar] [CrossRef]
- Johnson, K.H.; Sletten, K.; Werdin, R.E.; Westermark, G.T.; O’Brien, T.D.; Westermark, P. Amino acid sequence variations in protein AA of cats with high and low incidences of AA amyloidosis. Comp. Biochem. Physiol. B 1989, 94, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Husebekk, A.; Skogen, B.; Husby, G.; Marhaug, G. Transformation of amyloid precursor SAA to Protein AA and incorporation in amyloid fibrils in vivo. Scand. J. Immunol. 1985, 21, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.T.; DiBartola, S.P.; Chew, D.J.; Gasper, P.W. Familial renal amyloidosis in Abyssinian cats. Vet. Pathol. 1984, 21, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Van der Linde-Sipman, J.S.; Niewold, T.A.; Tooten, P.C.J.; de Neijs-Backer, M.; Gruys, E. Generalized AA-amyloidosis in Siamese and Oriental cats. Vet. Immunol. Immunopathol. 1997, 56, 1–10. [Google Scholar] [CrossRef]
- Beatty, J.A.; Barrs, V.R.; Martin, P.A.; Nicoll, R.G.; France, M.P.; Foster, S.F.; Lamb, W.A.; Malik, R. Spontaneous hepatic rupture in six cats with systemic amyloidosis. J. Small Anim. Pract. 2002, 43, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Zuber, R.M. Systemic amyloidosis in Oriental and Siamese Cats. Aust. Vet. Practit. 1993, 23, 66–70. [Google Scholar]
- DiBartola, S.P.; Tarr, M.J.; Benson, M.D. Tissue distribution of amyloid deposits in Abyssinian cats with familial amyloidosis. J. Comp. Pathol. 1986, 96, 387–398. [Google Scholar] [CrossRef]
- Chew, D.J.; DiBartola, S.P.; Boyce, J.T.; Gasper, P.W. Renal amyloidosis in related Abyssinian cats. J. Am. Vet. Med. Assoc. 1982, 181, 139–142. [Google Scholar]
- Struck, A.K.; Klotz, D.; Hülskötter, K.; Wohlsein, P.; Schmitz, J.; Bräsen, J.H.; Distl, O. Complex segregation analysis of familial amyloidosis in Oriental shorthair cats. Vet. J. 2020, 265, 105552. [Google Scholar] [CrossRef]
- Godfrey, D.R.; Day, M.J. Generalised amyloidosis in two Siamese cats: Spontaneous liver haemorrhage and chronic renal failure. J. Small. Anim. Pract. 1998, 39, 442–447. [Google Scholar] [CrossRef]
- Niewold, T.A.; Van der Linde-Sipman, J.S.; Murphy, C.; Tooten, P.C.; Gruys, E. Familial amyloidosis in cats: Siamese and Abyssinian AA proteins differ in primary sequence and pattern of deposition. Amyloid 1999, 6, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Genova, F.; Nonnis, S.; Maffioli, E.; Tedeschi, G.; Strillacci, M.G.; Carisetti, M.; Sironi, G.; Cupaioli, F.A.; Di Nanni, N.; Mezzelani, A.; et al. Multi-omic analyses in Abyssinian cats with primary renal amyloid deposits. Sci. Rep. 2021, 11, 8339. [Google Scholar] [CrossRef]
- Connors, L.H.; Lim, A.; Prokaeva, T.; Roskens, V.A.; Costello, C.E. Tabulation of human transthyretin (TTR) variants, 2003. Amyloid 2003, 10, 160–184. [Google Scholar] [CrossRef] [PubMed]
- Van Broeckhoven, C.; Haan, J.; Bakker, E.; Hardy, J.A.; Van Hul, W.; Wehnert, A.; Vegter-Van der Vlis, M.; Roos, R.A.C. Amyloid β; Protein precursor gene and hereditary cerebral hemorrhage with amyloidosis (dutch). Science 1990, 248, 1120–1122. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, L.; van Duijn, C.M.; Cras, P.; Cruts, M.; Van Hul, W.; van Harskamp, F.; Warren, A.; McInnis, M.G.; Antonarakis, S.E.; Martin, J.J.; et al. Presenile dementia and cerebral haemorrhage linked to a mutation at codon 692 of the β–amyloid precursor protein gene. Nat. Genet. 1992, 1, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Kamino, K.; Orr, H.T.; Payami, H.; Wijsman, E.M.; Alonso, M.E.; Pulst, S.M.; Anderson, L.; O’Dahl, S.; Nemens, E.; White, J.A.; et al. Linkage and mutational analysis of familial Alzheimer disease kindreds for the APP gene region. Am. J. Hum. Genet. 1992, 51, 998–1014. [Google Scholar]
- Sikora, J.; Kmochová, T.; Mušálková, D.; Pohludka, M.; Přikryl, P.; Hartmannová, H.; Hodaňová, K.; Trešlová, H.; Nosková, L.; Mrázová, L.; et al. A mutation in the SAA1 promoter causes hereditary amyloid A amyloidosis. Kidney Int. 2022, 101, 349–359. [Google Scholar] [CrossRef]
- Saleem, A.N.; Chen, Y.-H.; Baek, H.J.; Hsiao, Y.-W.; Huang, H.-W.; Kao, H.-J.; Liu, K.-M.; Shen, L.-F.; Song, I.w.; Tu, C.-P.D.; et al. Mice with alopecia, osteoporosis, and systemic amyloidosis due to mutation in Zdhhc13, a gene coding for palmitoyl acyltransferase. PLoS Genet. 2010, 6, e1000985. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Tamba, C.L.; Wen, Y.J.; Li, P.; Ren, W.L.; Ni, Y.L.; Gao, J.; Zhang, Y.M. mrMLM v4.0.2: An R Platform for multi-locus genome-wide association studies. Genom. Proteom. Bioinform. 2020, 18, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-B.; Feng, J.-Y.; Ren, W.-L.; Huang, B.; Zhou, L.; Wen, Y.-J.; Zhang, J.; Dunwell, J.M.; Xu, S.; Zhang, Y.-M. Improving power and accuracy of genome-wide association studies via a multi-locus mixed linear model methodology. Sci. Rep. 2016, 6, 19444. [Google Scholar] [CrossRef]
- Tamba, C.L.; Zhang, Y.M. A fast mrMLM algorithm for multi-locus genome-wide association studies. biorXiv 2018, 341784. [Google Scholar] [CrossRef]
- Wen, Y.J.; Zhang, H.; Ni, Y.L.; Huang, B.; Zhang, J.; Feng, J.Y.; Wang, S.B.; Dunwell, J.M.; Zhang, Y.M.; Wu, R. Methodological implementation of mixed linear models in multi-locus genome-wide association studies. Brief. Bioinform. 2018, 19, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, J.Y.; Ni, Y.L.; Wen, Y.J.; Niu, Y.; Tamba, C.L.; Yue, C.; Song, Q.; Zhang, Y.M. pLARmEB: Integration of least angle regression with empirical Bayes for multilocus genome-wide association studies. Heredity 2017, 118, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Tamba, C.L.; Ni, Y.L.; Zhang, Y.M. Iterative sure independence screening EM-Bayesian LASSO algorithm for multi-locus genome-wide association studies. PLoS Comput. Biol. 2017, 13, e1005357. [Google Scholar] [CrossRef]
- Neyman, J.; Pearson, E.S. On the use and interpretation of certain test criteria for purposes of statistical inference: Part I. Biometrika 1928, 20A, 175–240. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, T.; Zhou, Y.; Yin, S.; Li, P.; Liu, J.; Xu, S.; Yang, Z.; Xu, C. Genome-wide association mapping of starch pasting properties in maize using single-locus and multi-locus models. Front. Plant Sci. 2018, 9, 1311. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Safran, M.; Dalah, I.; Alexander, J.; Rosen, N.; Iny Stein, T.; Shmoish, M.; Nativ, N.; Bahir, I.; Doniger, T.; Krug, H.; et al. GeneCards version 3: The human gene integrator. Database 2010, 2010, baq020. [Google Scholar] [CrossRef]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed]
- Mez, J.; Chung, J.; Jun, G.; Kriegel, J.; Bourlas, A.P.; Sherva, R.; Logue, M.W.; Barnes, L.L.; Bennett, D.A.; Buxbaum, J.D.; et al. Two novel loci, COBL and SLC10A2, for Alzheimer’s disease in African Americans. Alzheimers. Dement. 2017, 13, 119–129. [Google Scholar] [CrossRef]
- Marzi, C.; Albrecht, E.; Hysi, P.G.; Lagou, V.; Waldenberger, M.; Tönjes, A.; Prokopenko, I.; Heim, K.; Blackburn, H.; Ried, J.S.; et al. Genome-Wide Association Study identifies two novel regions at 11p15.5-p13 and 1p31 with major impact on acute-phase Serum Amyloid A. PLoS Genet. 2010, 6, e1001213. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Zhang, C.; Yu, W.; Li, L.; Zhang, Z.; Liu, T.; Zhang, Y.; Fan, G.; Huangfu, H. Receptor Type Protein Tyrosine Phosphatase Epsilon (PTPRE) plays an oncogenic role in thyroid carcinoma by activating the AKT and ERK1/2 signaling pathway. Curr. Cancer Drug Targets 2023, 23, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Aschner, Y.; Downey, G.P. The importance of tyrosine phosphorylation control of cellular signaling pathways in respiratory disease: pY and pY not. Am. J. Respir. Cell Mol. Biol. 2018, 59, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Rettenberger, G.; Klett, C.; Zechner, U.; Bruch, J.; Just, W.; Vogel, W.; Hameister, H. ZOO-FISH analysis: Cat and human karyotypes closely resemble the putative ancestral mammalian karyotype. Chromosome Res. 1995, 3, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Abrami, L.; Leppla, S.H.; van der Goot, F.G. Receptor palmitoylation and ubiquitination regulate anthrax toxin endocytosis. J. Cell Biol. 2006, 172, 309–320. [Google Scholar] [CrossRef]
- Boyer, J.L. Bile formation and secretion. Compr. Physiol. 2013, 3, 1035–1078. [Google Scholar] [CrossRef]
- Li, C.; Fu, Y.; Sun, R.; Wang, Y.; Wang, Q. Single-locus and multi-locus genome-wide association studies in the genetic dissection of fiber quality traits in upland cotton (Gossypium hirsutum L.). Front. Plant Sci. 2018, 9, 1083. [Google Scholar] [CrossRef]


{kind=link}
| SNP-ID | FCA | Position | MAF | LOD Score | PVE (%) | Method |
|---|---|---|---|---|---|---|
| CHRA1.33307498 | A1 | 29,635,172 | 0.36 | 12.65 | 26.61 | mrMLM |
| 0.35 | 5.40 | 28.50 | FASTmrMLM | |||
| 0.35 | 10.27 | 16.75 | FASTmrEMMA | |||
| CHRA1.35096216 | A1 | 31,080,887 | 0.29 | 3.78 | 17.36 | pLARmEB |
| 0.29 | 6.05 | 18.54 | ISIS EM-BLASSO | |||
| CHRA1.87876182 | A1 | 74,262,275 | 0.39 | 7.21 | 12.22 | mrMLM |
| 0.40 | 4.30 | 4.21 | FASTmrMLM | |||
| 0.40 | 3.88 | 3.74 | FASTmrEMMA | |||
| CHRUN5.1424878 | D1 | 30,498,477 | 0.31 | 5.20 | 15.65 | FASTmrMLM |
| 0.31 | 6.84 | 15.35 | ISIS EM-BLASSO | |||
| CHRD1.83154354 | D1 | 52,625,246 | 0.45 | 3.07 | 3.23 | FASTmrEMMA |
| 0.45 | 4.20 | 8.47 | pLARmEB | |||
| CHRD1.112359805 | D1 | 75,099,182 | 0.23 | 6.34 | 11.86 | mrMLM |
| 0.23 | 3.49 | 1.96 | FASTmrMLM | |||
| 0.23 | 5.59 | 5.01 | pLARmEB | |||
| CHRD2.113529492 | D2 | 84,125,513 | 0.35 | 6.32 | 14.73 | FASTmrMLM |
| 0.35 | 6.77 | 10.81 | FASTmrEMMA | |||
| 0.35 | 6.29 | 16.56 | pLARmEB | |||
| 0.35 | 13.35 | 27.57 | ISIS EM-BLASSO | |||
| CHRA1.170535359 | D3 | 12,578,042 | 0.44 | 3.32 | 5.39 | mrMLM |
| 0.44 | 6.91 | 10.35 | ISIS-EM-BLASSO |
| SNP-ID | FCA | Gene | Start | End | Related Functions |
|---|---|---|---|---|---|
| CHRA1.87876182 | A1 | SLC10A2 | 73,755,036 | 73,774,055 | Late onset Alzheimer’s disease [34] |
| CHRD1.112359805 | D1 | SAA1 | 74,193,158 | 74,196,364 | AA-amyloidosis [19] |
| HPS5 GTF2H1 LDHA LDHC | 74,220,513 74,280,919 74,338,285 74,353,740 | 74,280,965 74,318,855 74,349,667 74,397,837 | Major impact on SAA [35] | ||
| ZDHHC13 | 74,760,625 | 74,812,445 | AA-/AL-amyloidosis [20] | ||
| CHRD2.113529492 | D2 | PTPRE | 83,094,722 | 83,258,667 | Thyroid carcinoma [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esders, S.L.; Hülskötter, K.; Schreiner, T.; Wohlsein, P.; Schmitz, J.; Bräsen, J.H.; Distl, O. Single Nucleotide Polymorphisms Associated with AA-Amyloidosis in Siamese and Oriental Shorthair Cats. Genes 2023, 14, 2126. https://doi.org/10.3390/genes14122126
Esders SL, Hülskötter K, Schreiner T, Wohlsein P, Schmitz J, Bräsen JH, Distl O. Single Nucleotide Polymorphisms Associated with AA-Amyloidosis in Siamese and Oriental Shorthair Cats. Genes. 2023; 14(12):2126. https://doi.org/10.3390/genes14122126
Chicago/Turabian StyleEsders, Stella L., Kirsten Hülskötter, Tom Schreiner, Peter Wohlsein, Jessica Schmitz, Jan H. Bräsen, and Ottmar Distl. 2023. "Single Nucleotide Polymorphisms Associated with AA-Amyloidosis in Siamese and Oriental Shorthair Cats" Genes 14, no. 12: 2126. https://doi.org/10.3390/genes14122126