

Mapping of QTLs Associated with Biological Nitrogen Fixation Traits in Peanuts (Arachis hypogaea L.) Using an Interspecific Population Derived from the Cross between the Cultivated Species and Its Wild Ancestors
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotyping of CSSL Population for Traits Associated with BNF
2.3. Statistical Analysis and QTL Mapping
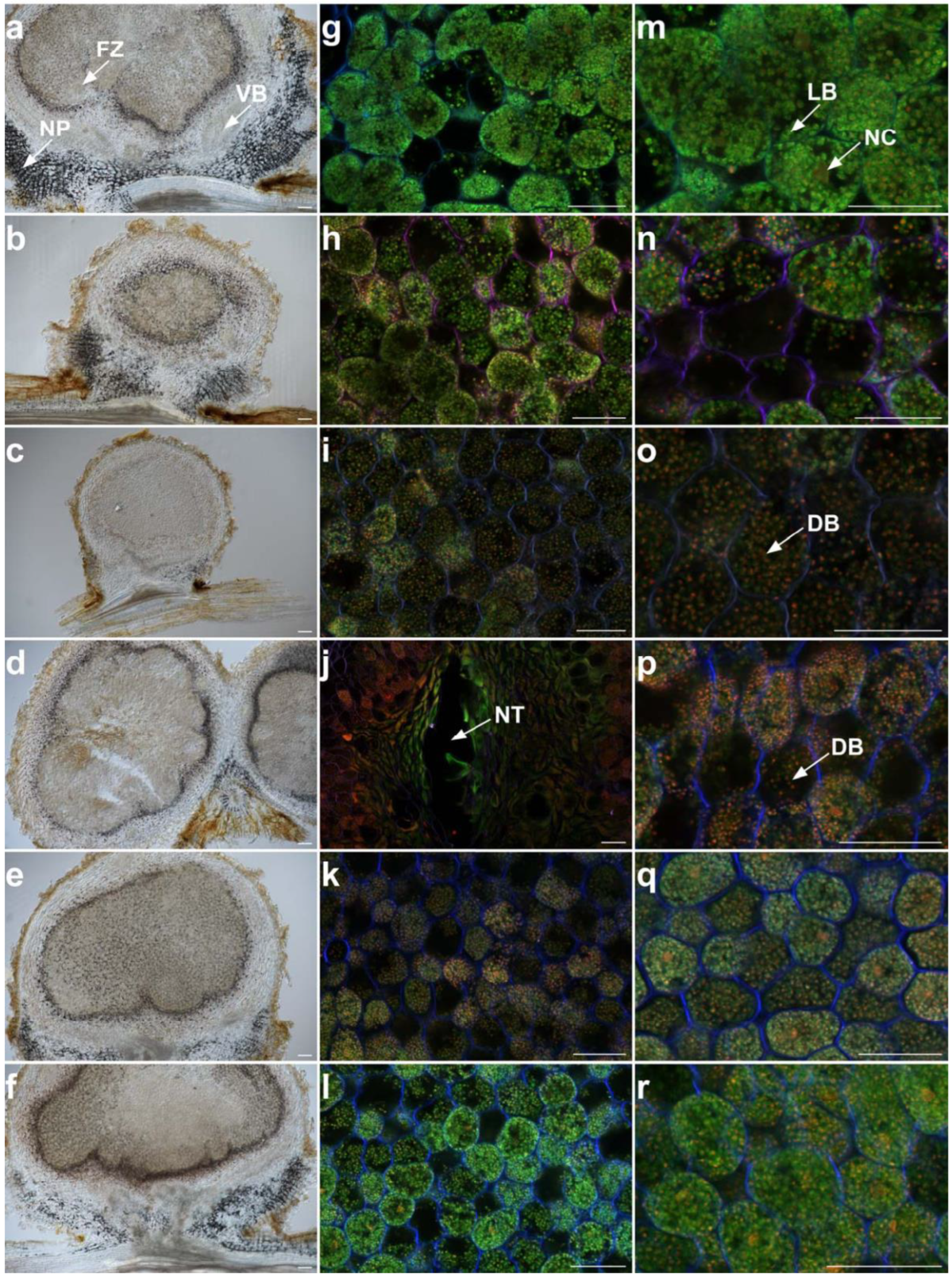
2.4. Cytological Analysis of Nodules
3. Results
3.1. Phenotypic Variation in BNF in the CSSL Population under Shade-House Conditions
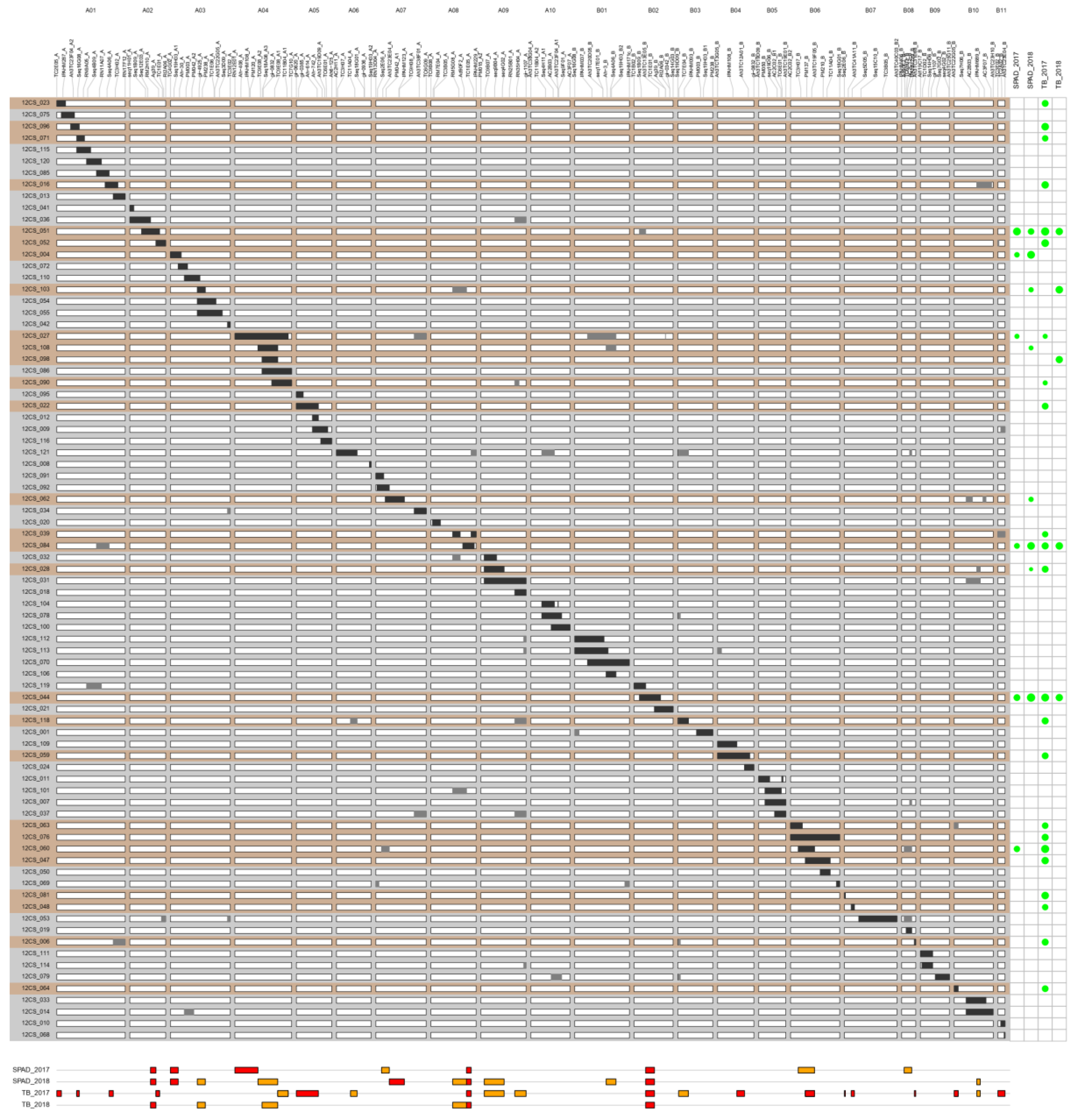
3.2. QTL Mapping
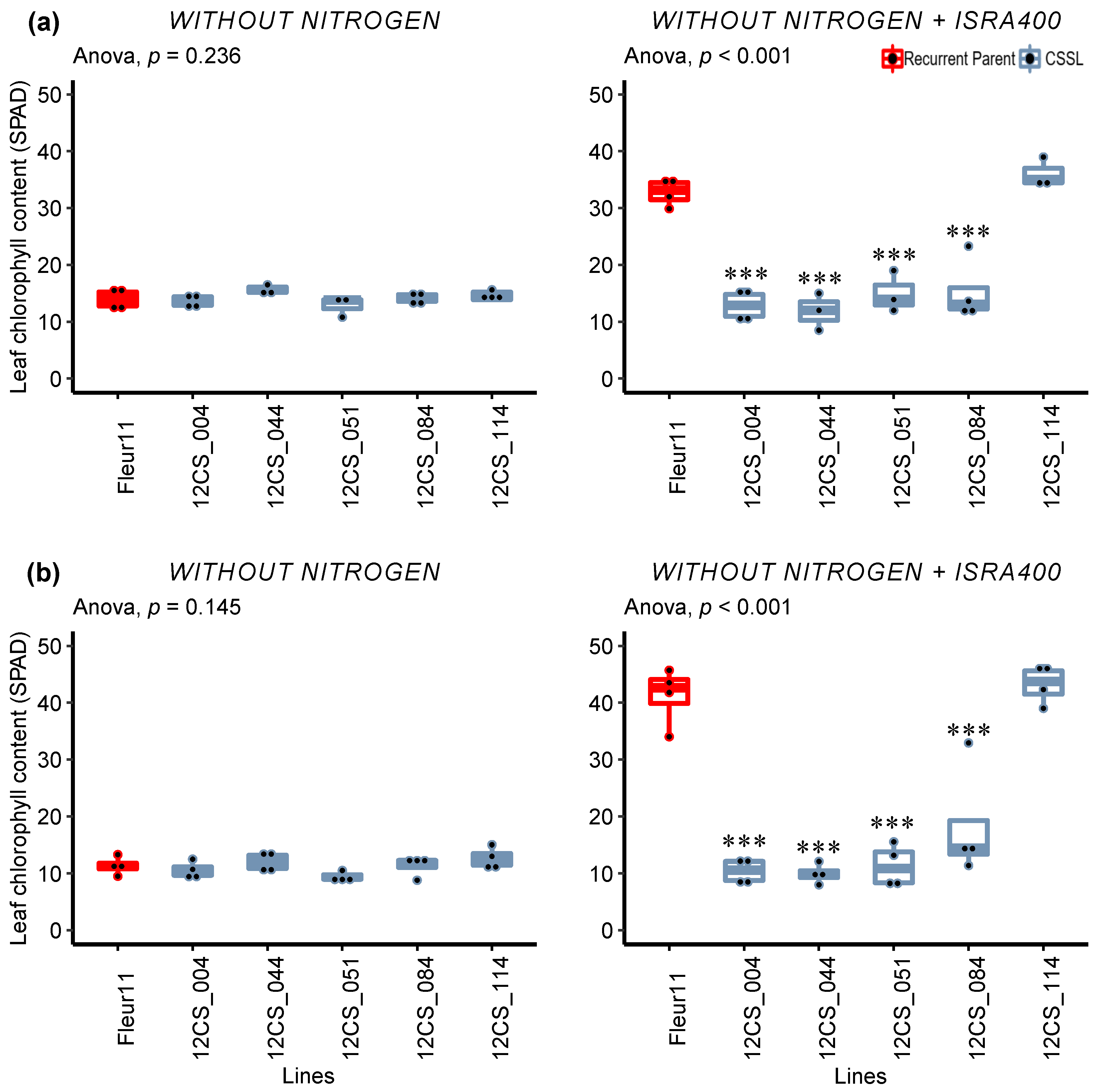
3.2.1. Chlorophyll Content
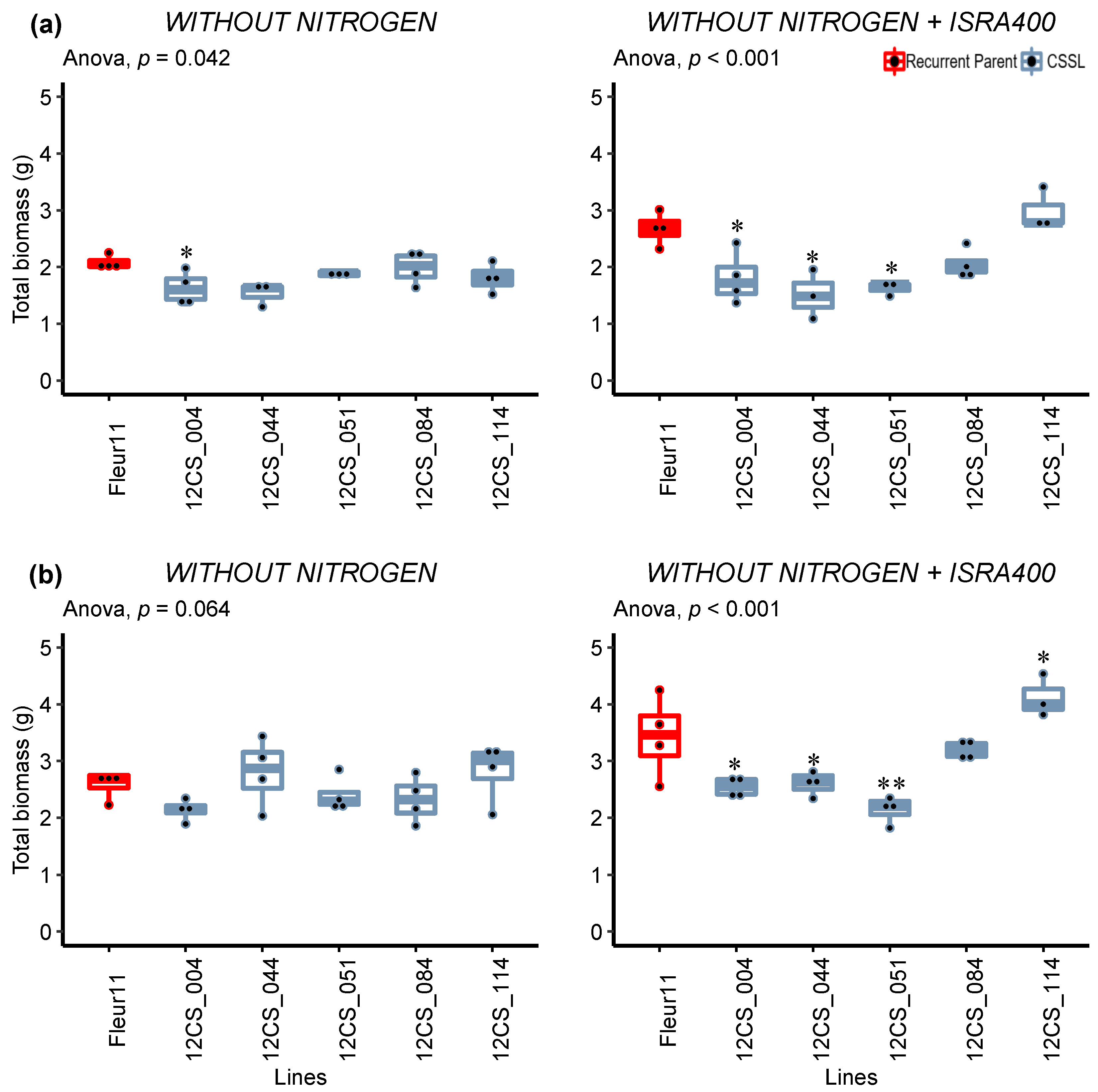
3.2.2. Total Biomass (TB)
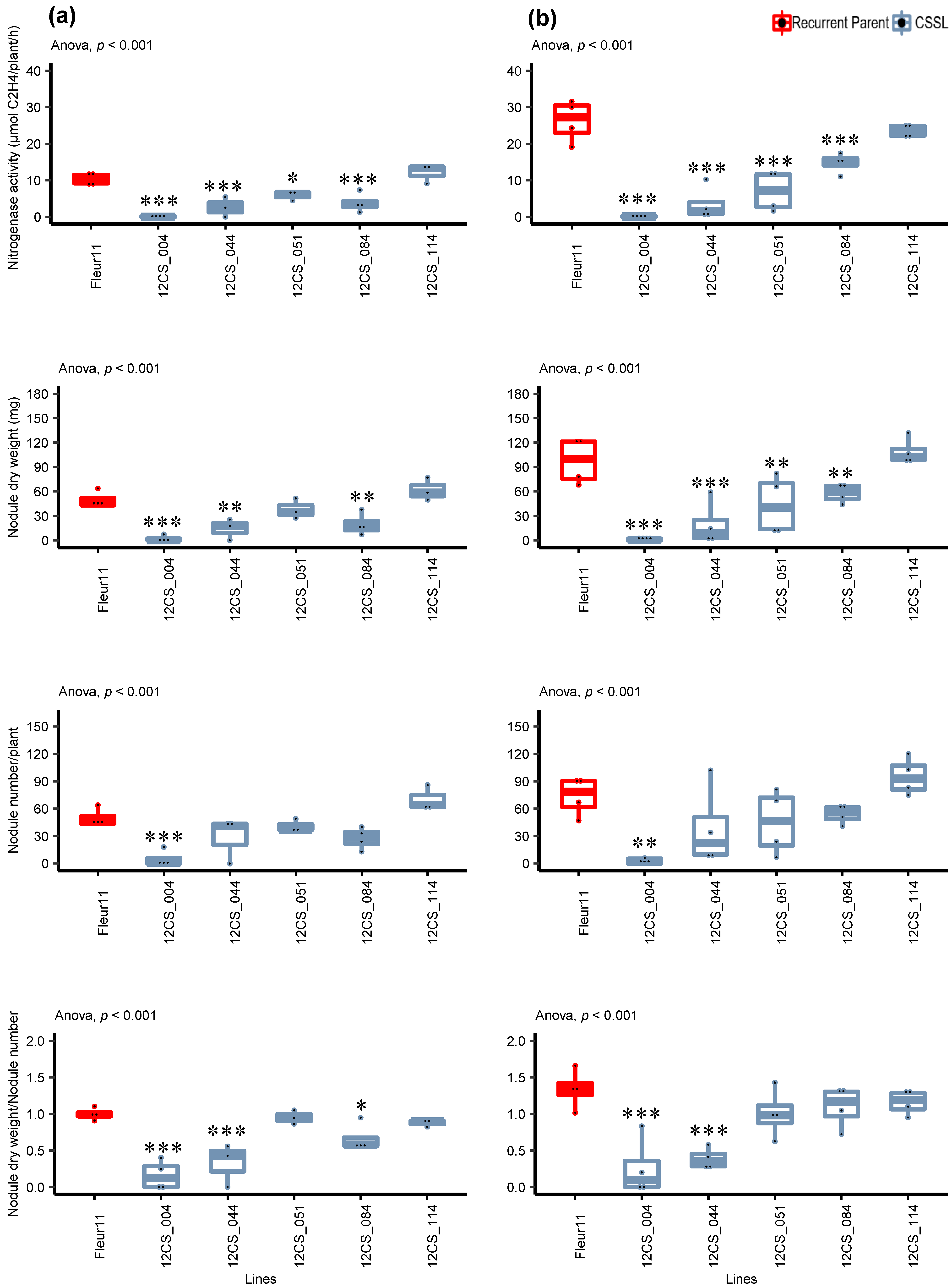
3.3. Validation of QTLs in Controlled Conditions
Detailed Analysis of the Response to Inoculation
4. Discussion
4.1. Variability in Traits Related to BNF and QTL Mapping
4.2. QTL Action on Nodulation Traits
4.3. Wild Species, Domestication, and BNF
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Udvardi, M.; Poole, P.S. Transport and Metabolism in Legume-Rhizobia Symbioses. Annu. Rev. Plant Biol. 2013, 64, 781–805. [Google Scholar] [CrossRef] [Green Version]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global Inputs of Biological Nitrogen Fixation in Agricultural Systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.-F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.M.; Jones, H.G.; Karley, A.J.; et al. Improving Intercropping: A Synthesis of Research in Agronomy, Plant Physiology and Ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Gepts, P. Crop Domestication as a Long-Term Selection Experiment. Plant Breed. Rev. 2004, 24, 1–44. [Google Scholar]
- Hufford, M.B.; Berny Mier y Teran, J.C.; Gepts, P. Crop Biodiversity: An Unfinished Magnum Opus of Nature. Annu. Rev. Plant Biol. 2019, 70, 727–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, N.; Liu, A.; Kan, L.; Li, M.-W.; Lam, H.-M. Potential Uses of Wild Germplasms of Grain Legumes for Crop Improvement. Int. J. Mol. Sci. 2017, 18, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, S.S.; Sachs, J.L. Agriculture and the Disruption of Plant–Microbial Symbiosis. Trends Ecol. Evol. 2020, 35, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, J.P.; Mendes, R.; Raaijmakers, J. Impact of Plant Domestication on Rhizosphere Microbiome Assembly and Functions. Plant Mol. Biol. 2015, 90, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Mutch, L.A.; Young, J.P.W. Diversity and Specificity of Rhizobium Leguminosarum Biovar Viciae on Wild and Cultivated Legumes. Mol. Ecol. 2004, 13, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Kaashyap, M.; Rathore, A.; Das, R.R.; Parupalli, S.; Upadhyaya, H.D.; Gopalakrishnan, S.; Gaur, P.M.; Singh, S.; Kaur, J.; et al. Phylogenetic Diversity of Mesorhizobium in Chickpea. J. Biosci. 2014, 39, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Chen, W.; Luo, S.; Ma, L.; Li, X.; Tian, C. Rhizosphere Microbiota Assemblage Associated with Wild and Cultivated Soybeans Grown in Three Types of Soil Suspensions. Arch. Agron. Soil Sci. 2019, 65, 74–87. [Google Scholar] [CrossRef]
- Kiers, E.T.; Hutton, M.G.; Denison, R.F. Human Selection and the Relaxation of Legume Defences against Ineffective Rhizobia. Proc. R. Soc. B Biol. Sci. 2007, 274, 3119–3126. [Google Scholar] [CrossRef] [Green Version]
- Provorov, N.A.; Tikhonovich, I.A. Genetic Resources for Improving Nitrogen Fixation in Legume-Rhizobia Symbiosis. Genet. Resour. Crop Evol. 2003, 50, 89–99. [Google Scholar] [CrossRef]
- Hufford, M.B.; Lubinksy, P.; Pyhäjärvi, T.; Devengenzo, M.T.; Ellstrand, N.C.; Ross-Ibarra, J. The Genomic Signature of Crop-Wild Introgression in Maize. PLoS Genet. 2013, 9, e1003477. [Google Scholar] [CrossRef]
- Liu, J.; Yu, X.; Qin, Q.; Dinkins, R.D.; Zhu, H. The Impacts of Domestication and Breeding on Nitrogen Fixation Symbiosis in Legumes. Front. Genet. 2020, 11, 973. [Google Scholar] [CrossRef]
- Muñoz, N.; Qi, X.; Li, M.-W.; Xie, M.; Gao, Y.; Cheung, M.-Y.; Wong, F.-L.; Lam, H.-M. Improvement in Nitrogen Fixation Capacity Could Be Part of the Domestication Process in Soybean. Heredity 2016, 117, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.K.; Msimbira, L.A.; Dakora, F.D. Phylogenetically Diverse Group of Native Bacterial Symbionts Isolated from Root Nodules of Groundnut (Arachis hypogaea L.) in South Africa. Syst. Appl. Microbiol. 2017, 40, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Zaiya, Z.A.; Fonceka, D.; Fall, S.; Fabra, A.; Ibañez, F.; Pignoly, S.; Diouf, A.; Touré, O.; Faye, M.N.; Hocher, V.; et al. Genetic Diversity and Symbiotic Efficiency of Rhizobial Strains Isolated from Nodules of Peanut (Arachis hypogaea L.) in Senegal. Agric. Ecosyst. Environ. 2018, 265, 384–391. [Google Scholar] [CrossRef]
- Janila, P.; Nigam, S.N.; Pandey, M.K.; Nagesh, P.; Varshney, R.K. Groundnut Improvement: Use of Genetic and Genomic Tools. Front. Plant Genet. Genom. 2013, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wynne, J.C.; Elkan, G.H.; Schneeweis, T.J. Increasing Nitrogen Fixation of the Groundnut by Strain and Host Selection. In Proceedings of the International Workshop on Groundnuts, Patancheru, India, 13–17 October 1980; ICRISAT Center: Hyderabad, India; pp. 95–109. [Google Scholar]
- Arrenddell, S.; Wynne, J.C.; Elkan, G.H.; Isleib, T.G. Variation for Nitrogen Fixation Among Progenies of a Virginia ✕ Spanish Peanut Cross1. Crop Sci. 1985, 25, 865–869. [Google Scholar] [CrossRef]
- Nigam, S.N.; Dwivedi, S.L.; Nambiar, P.T.C.; Gibbons, R.W.; Dart, P.J. Combining Ability Analysis of N2-Fixation and Related Traits in Peanut. Peanut Sci. 1985, 12, 55–57. [Google Scholar] [CrossRef] [Green Version]
- Pimratch, S.; Jogloy, S.; Toomsan, B.; Jaisil, P.; Sikhinarum, J.; Kesmala, T.; Patanothai, A. Evaluation of Seven Peanut Genotypes for Nitrogen Fixation and Agronomic Traits. Songklanakarin J. Sci. Technol. 2004, 26, 295–304. [Google Scholar]
- Dutta, M.; Reddy, L.J. Further Studies on Genetics of Nonnodulation in Peanut. Crop Sci. 1988, 28, 60–62. [Google Scholar] [CrossRef]
- Gallo-Meagher, M.; Dashiell, K.E.; Gorbet, D.W. Parental Effects in the Inheritance of Nonnodulation in Peanut. J. Hered. 2001, 92, 86–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Tan, L.; Lopez, Y.; Maku, J.; Liu, F.; Zhou, H.; Tseng, Y.-C.; Yang, X.; Hsieh, Y.; Song, J.; et al. Morphological and Genetic Characterization of Non-Nodulating Peanut Recombinant Inbred Lines. Crop Sci. 2018, 58, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Griesmann, M.; Chang, Y.; Liu, X.; Song, Y.; Haberer, G.; Crook, M.B.; Billault-Penneteau, B.; Lauressergues, D.; Keller, J.; Imanishi, L.; et al. Phylogenomics reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 2018, 361, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karmakar, K.; Kundu, A.; Rizvi, A.Z.; Dubois, E.; Severac, D.; Czernic, P.; Cartieaux, F.; DasGupta, M. Transcriptomic Analysis With the Progress of Symbiosis in ‘Crack-Entry’ Legume Arachis hypogaea Highlights Its Contrast With ‘Infection Thread’ Adapted Legumes. MPMI 2018, 32, 271–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [Green Version]
- Raul, B.; Bhattacharjee, O.; Ghosh, A.; Upadhyay, P.; Tembhare, K.; Singh, A.; Shaheen, T.; Ghosh, A.K.; Torres-Jerez, I.; Krom, N.; et al. Microscopic and Transcriptomic Analyses of Dalbergoid Legume Peanut Reveal a Divergent Evolution Leading to Nod-Factor-Dependent Epidermal Crack-Entry and Terminal Bacteroid Differentiation. MPMI 2022, 35, 131–145. [Google Scholar] [CrossRef]
- Sprent, J.I.; James, E.K. Legume Evolution: Where Do Nodules and Mycorrhizas Fit In? Plant Physiol. 2007, 144, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, F.; Wall, L.; Fabra, A. Starting Points in Plant-Bacteria Nitrogen-Fixing Symbioses: Intercellular Invasion of the Roots. J. Exp. Bot. 2017, 68, 1905–1918. [Google Scholar] [CrossRef] [Green Version]
- Quilbé, J.; Montiel, J.; Arrighi, J.-F.; Stougaard, J. Molecular Mechanisms of Intercellular Rhizobial Infection: Novel Findings of an Ancient Process. Front. Plant Sci. 2022, 13, 922982. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The Genome Sequences of Arachis duranensis and Arachis ipaensis, the Diploid Ancestors of Cultivated Peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Fonceka, D.; Hodo-Abalo, T.; Rivallan, R.; Faye, I.; Sall, M.N.; Ndoye, O.; Fávero, A.P.; Bertioli, D.J.; Glaszmann, J.-C.; Courtois, B.; et al. Genetic Mapping of Wild Introgressions into Cultivated Peanut: A Way toward Enlarging the Genetic Basis of a Recent Allotetraploid. BMC Plant Biol. 2009, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Mallikarjuna, N.; Senthilvel, S.; Hoisington, D. Development of new sources of tetraploid Arachis to broaden the genetic base of cultivated groundnut (Arachis hypogaea L.). Genet. Resour. Crop. Evol. 2010, 58, 889–907. [Google Scholar] [CrossRef] [Green Version]
- Fonceka, D.; Tossim, H.-A.; Rivallan, R.; Vignes, H.; Faye, I.; Ndoye, O.; Moretzsohn, M.C.; Bertioli, D.J.; Glaszmann, J.-C.; Courtois, B.; et al. Fostered and left behind alleles in peanut: Interspecific QTL mapping reveals footprints of domestication and useful natural variation for breeding. BMC Plant Biol. 2012, 12, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonceka, D.; Tossim, H.-A.; Rivallan, R.; Vignes, H.; Lacut, E.; de Bellis, F.; Faye, I.; Ndoye, O.; Leal-Bertioli, S.C.M.; Valls, J.F.M.; et al. Construction of Chromosome Segment Substitution Lines in Peanut (Arachis hypogaea L.) Using a Wild Synthetic and QTL Mapping for Plant Morphology. PLoS ONE 2012, 7, e48642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khera, P.; Pandey, M.K.; Mallikarjuna, N.; Sriswathi, M.; Roorkiwal, M.; Janila, P.; Sharma, S.; Shilpa, K.; Sudini, H.; Guo, B.; et al. Genetic imprints of domestication for disease resistance, oil quality, and yield component traits in groundnut (Arachis hypogaea L.). Mol. Genet. Genom. 2018, 294, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tossim, H.-A.; Nguepjop, J.R.; Diatta, C.; Sambou, A.; Seye, M.; Sane, D.; Rami, J.-F.; Fonceka, D. Assessment of 16 Peanut (Arachis hypogaea L.) CSSLs Derived from an Interspecific Cross for Yield and Yield Component Traits: QTL Validation. Agronomy 2020, 10, 583. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.M. A Manual for the Practical Study of the Root-Nodule Bacteria; Blackwell Scientific Publications: Hoboken, NJ, USA, 1970. [Google Scholar]
- Kung, F.H.; Bey, C.F. Heritability Construction for Provenance and Family Selection; Department of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1977.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Takebe, I. Cell Wall Regeneration and Cell Division in Isolated Tobacco Mesophyll Protoplasts. Planta 1970, 92, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Sinharoy, S.; DasGupta, M. RNA Interference Highlights the Role of CCaMK in Dissemination of Endosymbionts in the Aeschynomeneae Legume Arachis. MPMI 2009, 22, 1466–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gully, D.; Gargani, D.; Bonaldi, K.; Grangeteau, C.; Chaintreuil, C.; Fardoux, J.; Nguyen, P.; Marchetti, R.; Nouwen, N.; Molinaro, A.; et al. A Peptidoglycan-Remodeling Enzyme Is Critical for Bacteroid Differentiation in Bradyrhizobium spp. During Legume Symbiosis. MPMI 2016, 29, 447–457. [Google Scholar] [CrossRef] [Green Version]
- Songwattana, P.; Chaintreuil, C.; Wongdee, J.; Teulet, A.; Mbaye, M.; Piromyou, P.; Gully, D.; Fardoux, J.; Zoumman, A.M.A.; Camuel, A.; et al. Identification of Type III Effectors Modulating the Symbiotic Properties of Bradyrhizobium vignae Strain ORS3257 with Various Vigna Species. Sci. Rep. 2021, 11, 4874. [Google Scholar] [CrossRef]
- Sahrawat, K.L.; Srinivasa Rao, B.; Nambiar, P.T.C. Macro- and Micronutrient Uptake by Nodulating and Non-Nodulating Peanut Lines. Plant Soil 1988, 109, 291–293. [Google Scholar] [CrossRef] [Green Version]
- Vollmann, J.; Walter, H.; Sato, T.; Schweiger, P. Digital Image Analysis and Chlorophyll Metering for Phenotyping the Effects of Nodulation in Soybean. Comput. Electron. Agric. 2011, 75, 190–195. [Google Scholar] [CrossRef]
- Ramaekers, L.; Galeano, C.H.; Garzón, N.; Vanderleyden, J.; Blair, M.W. Identifying Quantitative Trait Loci for Symbiotic Nitrogen Fixation Capacity and Related Traits in Common Bean. Mol. Breed. 2013, 31, 163–180. [Google Scholar] [CrossRef]
- Hamawaki, R.L.; Kantartzi, S.K. Di-Nitrogen Fixation at the Early and Late Growth Stages of Soybean. Acta Scientiarum. Agron. 2018, 40, e36372. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wu, L.; Baddeley, J.A.; Watson, C.A. Models of biological nitrogen fixation of legumes. A review. Agron. Sustain. Dev. 2011, 31, 155–172. [Google Scholar] [CrossRef]
- Herridge, D.F.; Robertson, M.J.; Cocks, B.; Peoples, M.B.; Holland, J.F.; Heuke, L. Low nodulation and nitrogen fixation of mungbean reduce biomass and grain yields. Aust. J. Exp. Agric. 2005, 45, 269–277. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Nakabayashi, K.; Mohapatra, P.K.; Thompson, J.; Fujita, K. Effect of Nitrogen Deficiency on Biomass Production, Photosynthesis, Carbon Partitioning, and Nitrogen Nutrition Status of Melaleuca and Eucalyptus Species. Soil Sci. Plant Nutr. 2003, 49, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.A.; Boscariol, R.L.; Moon, D.H.; Camargo, L.E.A.; Tsai, S.M. Effects of Phaseolus Vulgaris QTL in Controlling Host-Bacteria Interactions under Two Levels of Nitrogen Fertilization. Genet. Mol. Biol. 2000, 23, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Bourion, V.; Rizvi, S.M.H.; Fournier, S.; de Larambergue, H.; Galmiche, F.; Marget, P.; Duc, G.; Burstin, J. Genetic dissection of nitrogen nutrition in pea through a QTL approach of root, nodule, and shoot variability. Theor. Appl. Genet. 2010, 121, 71–86. [Google Scholar] [CrossRef]
- Nicolás, M.F.; Hungria, M.; Arias, C.A.A. Identification of Quantitative Trait Loci Controlling Nodulation and Shoot Mass in Progenies from Two Brazilian Soybean Cultivars. Field Crops Res. 2006, 95, 355–366. [Google Scholar] [CrossRef]
- Santos, M.A.; Geraldi, I.O.; Garcia, A.A.F.; Bortolatto, N.; Schiavon, A.; Hungria, M. Mapping of QTLs Associated with Biological Nitrogen Fixation Traits in Soybean. Hereditas 2013, 150, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Ni, H.; Peng, Y.; Wang, J.; Wang, J.; Yuan, Y.; Fu, T.; Zhu, Z.; Zhang, J.; Pan, X.; Cui, Z.; et al. Mapping of Quantitative Trait Loci Underlying Nodule Traits in Soybean (Glycine max (L.) Merr.) and Identification of Genes Whose Expression Is Affected by the Sinorhizobium Fredii HH103 Effector Proteins NopL and NopT. Agronomy 2022, 12, 946. [Google Scholar] [CrossRef]
- Wang, J.; Ma, C.; Ma, S.; Zheng, H.; Tian, H.; Wang, X.; Wang, Y.; Jiang, H.; Wang, J.; Zhang, Z.; et al. Genetic Variation in GmCRP Contributes to Nodulation in Soybean (Glycine max Merr.). Crop J. 2022; in press. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, H.; Tan, L.; Shu, H.; Varshney, R.K.; Zhou, Z.; Zhao, Z.; Luo, Z.; Chitikineni, A.; Wang, L.; et al. Natural Polymorphisms in a Pair of NSP2 Homoeologs Can Cause Loss of Nodulation in Peanut. J. Exp. Bot. 2021, 72, 1104–1118. [Google Scholar] [CrossRef] [PubMed]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can Less Yield More? Is Reducing Nutrient Input into the Environment Compatible with Maintaining Crop Production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef]
- Damiani, I.; Pauly, N.; Puppo, A.; Brouquisse, R.; Boscari, A. Reactive Oxygen Species and Nitric Oxide Control Early Steps of the Legume—Rhizobium Symbiotic Interaction. Front. Plant Sci. 2016, 7, 454. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.D.; Liu, C.-W.; Chen, Y.; Miller, A.J. Nitrogen Sensing in Legumes. J. Exp. Bot. 2017, 68, 1919–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, M.R. Some Observations on Infection of Arachis hypogaea L. by Rhizobium. J. Exp. Bot. 1978, 29, 749–755. [Google Scholar] [CrossRef]
- Boogerd, F.C.; van Rossum, D. Nodulation of groundnut by Bradyrhizobium: A simple infection process by crack entry. FEMS Microbiol. Rev. 1997, 21, 5–27. [Google Scholar] [CrossRef]
- Guinel, F.C. Getting around the Legume Nodule: I. The Structure of the Peripheral Zone in Four Nodule Types. Botany 2009, 87, 1117–1138. [Google Scholar] [CrossRef]
- Bonaldi, K.; Gherbi, H.; Franche, C.; Bastien, G.; Fardoux, J.; Barker, D.; Giraud, E.; Cartieaux, F. The Nod Factor–Independent Symbiotic Signaling Pathway: Development of Agrobacterium rhizogenes–Mediated Transformation for the Legume Aeschynomene indica. MPMI 2010, 23, 1537–1544. [Google Scholar] [CrossRef] [Green Version]
- Kiers, E.T.; Rousseau, R.A.; West, S.A.; Denison, R.F. Host Sanctions and the Legume–Rhizobium Mutualism. Nature 2003, 425, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Oono, R.; Denison, R.F.; Kiers, E.T. Controlling the Reproductive Fate of Rhizobia: How Universal Are Legume Sanctions? New Phytol. 2009, 183, 967–979. [Google Scholar] [CrossRef]
- Oono, R.; Anderson, C.G.; Denison, R.F. Failure to Fix Nitrogen by Non-Reproductive Symbiotic Rhizobia Triggers Host Sanctions That Reduce Fitness of Their Reproductive Clonemates. Proc. R. Soc. B Biol. Sci. 2011, 278, 2698–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teulet, A.; Busset, N.; Fardoux, J.; Gully, D.; Chaintreuil, C.; Cartieaux, F.; Jauneau, A.; Comorge, V.; Okazaki, S.; Kaneko, T.; et al. The rhizobial type III effector ErnA confers the ability to form nodules in legumes. Proc. Natl. Acad. Sci. USA 2019, 116, 21758–21768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundu, A.; DasGupta, M. Silencing of Putative Cytokinin Receptor Histidine Kinase1 Inhibits Both Inception and Differentiation of Root Nodules in Arachis hypogaea. Mol. Plant Microbe Interact. 2018, 31, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Bouznif, B.; Guefrachi, I.; Rodríguez de la Vega, R.C.; Hungria, M.; Mars, M.; Alunni, B.; Shykoff, J.A. Phylogeography of the Bradyrhizobium spp. Associated With Peanut, Arachis hypogaea: Fellow Travelers or New Associations? Front. Microbiol. 2019, 10, 2041. [Google Scholar] [CrossRef]
- Zaiya, Z.A. Etude de La Composante Symbiotique Bactérienne Impliquée Dans l’adaptation de l’arachide Cultivée (Arachis hypogaea L.) Au Sénégal. Ph.D. Thesis, Cheikh Anta Diop de Dakar, Dakar, Senegal, 2018. [Google Scholar]
- Greenlon, A.; Chang, P.L.; Damtew, Z.M.; Muleta, A.; Carrasquilla-Garcia, N.; Kim, D.; Nguyen, H.P.; Suryawanshi, V.; Krieg, C.P.; Yadav, S.K.; et al. Global-Level Population Genomics Reveals Differential Effects of Geography and Phylogeny on Horizontal Gene Transfer in Soil Bacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 15200–15209. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, O.M.; Riva, O.; Peltzer, E. Analysis of Rhizobium etli and of its symbiosis with wild Phaseolus vulgaris supports coevolution in centers of host diversification. Proc. Natl. Acad. Sci. USA 2004, 101, 13548–13553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, O.M.; Collavino, M.M.; Mancini, U. Nodulation competitiveness and diversification of symbiosis genes in common beans from the American centers of domestication. Sci. Rep. 2022, 12, 4591. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T.L.; Wood, C.W.; Borges, I.L.; Stinchcombe, J.R. No evidence for adaptation to local rhizobial mutualists in the legume Medicago lupulina. Ecol. Evol. 2017, 7, 4367–4376. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Treatment | 2017 | 2018 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | F | Pr | h2 | Mean ± SD | F | Pr | h2 | ||
| SPAD | −N | 26.2 ± 7.82 | 1.017 | 0.456 | 0 | 14.8 ± 5.10 | 1 | 0.583 | 0 |
| +N | 43.5 ± 3.79 | 1.742 | 0.001 ** | 0.43 | 40.6 ± 2.06 | 2 | <0.001 *** | 0.50 | |
| −N+ISRA400 | 41.6 ± 6.42 | 3.843 | <0.001 *** | 0.74 | 39.8 ± 6.48 | 11.358 | <0.001 *** | 0.91 | |
| TB | −N | 2.88 ± 1.26 | 1.611 | 0.005 ** | 0.38 | 2.69 ± 0.96 | 2.486 | <0.001 *** | 0.60 |
| +N | 5.75 ± 1.62 | 1.310 | 0.073 | 0.24 | 7.98 ± 2.07 | 1.446 | 0.023 * | 0.30 | |
| −N+ISRA400 | 4.36 ± 1.63 | 2.324 | <0.001 *** | 0.57 | 3.90 ± 1.47 | 1.979 | <0.001 *** | 0.49 | |
| Trait | CSSL | LG | 2017 | 2018 | ||
|---|---|---|---|---|---|---|
| QTL Effect (%) | Conf. Int (cM) | QTL Effect (%) | Conf. Int (cM) | |||
| SPAD | 12CS_051 | A02 | −65.29 *** | 43.85–55.35 | −43.26 *** | 43.85–55.35 |
| 12CS_004 | A03 | −29.23 *** | 0–16.6 | −64.77 *** | 0–16.6 | |
| 12CS_103 | A03 | ns | −26.76 *** | 57.2–74.2 | ||
| 12CS_027 | A04 | −24.61 * | 0–48.975 | ns | ||
| 12CS_108 | A04 | ns | −22.79 *** | 48.975–90.975 | ||
| 12CS_060 | A07 | −41.77 *** | 12.1–28.525 | ns | ||
| 12CS_062 | A07 | ns | −26.32 *** | 28.525–60.0 | ||
| 12CS_084 | A08 | −33.79 *** | 75.525–85.375 | −61.67 *** | 75.525–85.375 | |
| 12CS_103 | A08 | ns | −26.76 *** | 46.15–75.525 | ||
| 12CS_028 | A09 | ns | −15.17 ** | 7.125–49.375 | ||
| 12CS_108 | B01 | ns | −22.79 *** | 66.725–87.525 | ||
| 12CS_044 | B02 | −47.89 *** | 24.55–43.25 | −73.49 *** | 24.55–43.25 | |
| 12CS_060 | B06 | −41.77 *** | 16.275–50.95 | ns | ||
| 12CS_060 | B08 | −41.77 *** | 5.25–21.25 | ns | ||
| 12CS_028 | B10 | ns | −15.17 ** | 48.025–55.525 | ||
| TB | 12CS_023 | A01 | −48.76 ** | 0–9.675 | ns | |
| 12CS_096 | A01 | −61.13 *** | 42.325–47.7 | ns | ||
| 12CS_071 | A01 | −41.65 * | 42.325–47.7 | ns | ||
| 12CS_016 | A01 | −58.32 ** | 111.125–119.975 | ns | ||
| 12CS_051 | A02 | −71.51 *** | 55.35–63.125 | −56.43 * | 43.85–55.35 | |
| 12CS_052 | A02 | −62.55 *** | 55.35–63.125 | ns | ||
| 12CS_103 | A03 | ns | −60.58 ** | 57.2–74.2 | ||
| 12CS_027 | A04 | −47.38 ** | 90.975–113.15 | ns | ||
| 12CS_090 | A04 | −45.74 * | 90.975–113.15 | ns | ||
| 12CS_098 | A04 | ns | −55.10 * | 57.625–90.975 | ||
| 12CS_022 | A05 | −47.87 * | 0–46.8 | ns | ||
| 12CS_118 | A06 | −52.02 * | 29.825–43.925 | ns | ||
| 12CS_084 | A08 | −53.28 ** | 75.525–85.375 | −58.35 * | 75.525–85.375 | |
| 12CS_103 | A08 | ns | −60.58 ** | 46.15–75.525 | ||
| 12CS_028 | A09 | −49.2 * | 7.125–49.375 | ns | ||
| 12CS_118 | A09 | −52.02 * | 72.1–96.1 | ns | ||
| 12CS_044 | B02 | −66.09 *** | 24.55–43.25 | −55.36 ** | 24.55–43.25 | |
| 12CS_118 | B03 | −52.02 * | 1.725–22.575 | ns | ||
| 12CS_059 | B04 | −49.12 ** | 40.875–57.225 | ns | ||
| 12CS_076 | B06 | −52.02 ** | 31.3–50.95 | ns | ||
| 12CS_060 | B06 | −69.63 *** | 31.3–50.95 | ns | ||
| 12CS_047 | B06 | −57.13 *** | 31.3–50.95 | ns | ||
| 12CS_081 | B07 | −60.27 ** | 0–2.05 | ns | ||
| 12CS_048 | B07 | −41.04 * | 14.5–21.275 | ns | ||
| 12CS_006 | B08 | −50.57 * | 26.95–29.8 | ns | ||
| 12CS_063 | B10 | −44.63 ** | 0–8.5 | ns | ||
| 12CS_064 | B10 | −47.1** | 0–8.5 | ns | ||
| 12CS_028 | B10 | −49.2 * | 48.025–55.525 | ns | ||
| 12CS_039 | B11 | −39.73 * | 0–15.1 | ns | ||
| Trait 1 | Trait 2 | Corr. Coef. |
|---|---|---|
| Leaf chlorophyll content | Nitrogenase activity | 0.92 *** |
| Nodule dry weight | 0.89 *** | |
| Nodule number | 0.81 ** | |
| Nodule dry weight and number ratio | 0.72 ** | |
| Total biomass | Nitrogenase activity | 0.84 *** |
| Nodule dry weight | 0.80 ** | |
| Nodule number | 0.74 ** | |
| Nodule dry weight and number ratio | 0.62 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nzepang, D.T.; Gully, D.; Nguepjop, J.R.; Zaiya Zazou, A.; Tossim, H.-A.; Sambou, A.; Rami, J.-F.; Hocher, V.; Fall, S.; Svistoonoff, S.; et al. Mapping of QTLs Associated with Biological Nitrogen Fixation Traits in Peanuts (Arachis hypogaea L.) Using an Interspecific Population Derived from the Cross between the Cultivated Species and Its Wild Ancestors. Genes 2023, 14, 797. https://doi.org/10.3390/genes14040797
Nzepang DT, Gully D, Nguepjop JR, Zaiya Zazou A, Tossim H-A, Sambou A, Rami J-F, Hocher V, Fall S, Svistoonoff S, et al. Mapping of QTLs Associated with Biological Nitrogen Fixation Traits in Peanuts (Arachis hypogaea L.) Using an Interspecific Population Derived from the Cross between the Cultivated Species and Its Wild Ancestors. Genes. 2023; 14(4):797. https://doi.org/10.3390/genes14040797
Chicago/Turabian StyleNzepang, Darius T., Djamel Gully, Joël R. Nguepjop, Arlette Zaiya Zazou, Hodo-Abalo Tossim, Aissatou Sambou, Jean-François Rami, Valerie Hocher, Saliou Fall, Sergio Svistoonoff, and et al. 2023. "Mapping of QTLs Associated with Biological Nitrogen Fixation Traits in Peanuts (Arachis hypogaea L.) Using an Interspecific Population Derived from the Cross between the Cultivated Species and Its Wild Ancestors" Genes 14, no. 4: 797. https://doi.org/10.3390/genes14040797