
SCN9A rs6746030 Polymorphism and Pain Perception in Combat Athletes and Non-Athletes
by
 , , and
, , and
Katarzyna Leźnicka
1,*,
Maciej Pawlak
2,
Marek Sawczuk
1,3,
Agata Gasiorowska
4 and
Agata Leońska-Duniec
1,* 1
Faculty of Physical Education, Gdansk University of Physical Education and Sport, 80-336 Gdansk, Poland
2
Department of Physiology and Biochemistry, Poznan University of Physical Education, 61-871 Poznan, Poland
3
Institute of Physical Culture Sciences, University of Szczecin, 70-453 Szczecin, Poland
4
Faculty of Psychology in Wroclaw, SWPS University of Social Sciences and Humanities, Ostrowskiego 30b, 54-238 Wroclaw, Poland
*
Authors to whom correspondence should be addressed.
Genes 2023, 14(3), 733; https://doi.org/10.3390/genes14030733
Submission received: 14 February 2023
/
Revised: 9 March 2023
/
Accepted: 15 March 2023
/
Published: 16 March 2023
(This article belongs to the Section Molecular Genetics and Genomics)
Abstract
:One of the genes associated with pain perception is SCN9A, which encodes an α-subunit of the voltage gated sodium channel, NaV1.7, a crucial player in peripheral pain sensation. It has been suggested that a common missense polymorphism within SCN9A (rs6746030; G>A; R1150W) may affect nociception in the general population, but its effects of pain perception in athletes remain unknown. Therefore, the aim of the study was to investigate the association between a polymorphism within SCN9A (rs6746030) and pain perception (pain threshold and pain tolerance) in the group of combat athletes (n = 214) and students (n = 92) who did not participate in sports at a professional level. Genotyping was carried out using TaqMan Real-Time PCR method. No significant differences were found between the SCN9A genotype distributions with respect to the pain threshold. However, the probability of having a high pain threshold was higher in the combat sports group than in the control group. The probability of having a decreased pain tolerance was higher in the carriers of the GA and AA genotype than in the homozygotes of the GG genotype. Moreover, the possibility of having a high pain threshold was higher in the combat athlete group than in the control group. The results of our study suggest that the SCN9A rs6746030 polymorphism may affect pain perception. However, the additional effect of the experimental group may suggest that pain tolerance is significantly modulated by other factors, such as the systematic exposure of the athletes’ bodies to short-term high-intensity stimuli during training sessions.
1. Introduction
Pain is defined as an unpleasant sensory and emotional experience associated with actual or potential tissue damage or described in terms of such damage [1]. A negative emotional attitude towards pain occurs at every stage of life, beginning already in fetal life and manifested in neuropsychological, hormonal, and behavioral reactions to pain and non-pain stimuli [2].
By recognizing external and internal threats to the body, pain performs an extremely important function, especially task in the field of information, warning, and protection, which are crucial for the survival of each human being. Pain is admittedly considered as a subjective experience, but its quantitative and qualitative dimension is continuously modulated and modified by numerous factors such as social, psychological, religious, cultural, and personality factors, as well as physical activity [2].
Scientific research by Heneweer et al. [3] has shown that both too little and too much physical activity predisposes patients to the onset of pain. The constant or regular influx of nociceptive stimuli leads to changes in the pain processing system, including peripheral and central sensitization processes and activation of antinociceptive mechanisms. Increased pain sensitivity is associated with numerous pain disorders and is considered one of the features of central sensitization of the nervous system [4]. The phenomenon of pain is of particular interest to healthy individuals exposed to physical exertion, especially in athletes who accept pain as an integral part of their daily experience. Many hours of intensive training and frequent participation in competitions (fights) make these athletes a group that is very often exposed to intense pain [5]. On the other hand, according to theoretical studies and empirical reports, physical activity is considered one of the most important elements of pain prevention and treatment. The significant differentiation of pain response, both in patients and in physically active individuals and competitive athletes, confirms the multifactorial nature of this phenomenon [6]. One of the underlying mechanisms for individual differences in pain perception is genetic susceptibility. Multiple genes, derived from human and animal studies, have been shown to be important in modulating pain perception [7,8,9]. Literature research conducted by Wistrom et al. [7] allowed for the identification of 242 genes linked to pain-associated behaviors. Due to the function of the pain gene products, they were divided into six functional groups: (1) voltage-gated and ligand-gated ion channels (e.g., sodium channel NaV1.7—SCN9A); (2) G protein-coupled receptors—GPCRs (e.g., delta-opioid receptor—OPRD1); (3) neuropeptides, neurotransmitters, and neurotrophins (e.g., tachykinin 1—TAC1); (4) growth factors, hormones, and cytokines (e.g., interleukin 10—IL-10); (5) enzymes and enzyme-linked receptors (e.g., mitogen-activated protein kinase 1—MAPK1); and (6) transcriptional and translational control and mRNA processing (e.g., PR domain-containing protein 12—PRDM12) [7,9,10,11,12].
Recent studies have revealed that variations in voltage-gated sodium channel genes are key players in peripheral pain processing [6,7,8]. For example, inactivating mutations in SCN9A result in congenital insensitivity to pain, whereas gain-of-function mutations generate different pain syndromes such as inherited erythromelalgia, paroxysmal extreme pain disorder, and small-fiber neuropathy [13]. This gene has been extensively linked to human and animal pain perception [13,14,15,16]. Yet, despite over a decade of research into SCN9A, to the best of our knowledge, their relationship with sport skills is almost unknown. A small number of studies on association between the SCN9A genotypes and pain perception in athletes led us to interest this important issue. The management of pain by athletes and the pursuit of pain control is an integral part of sports, but it is also one of the most important skills in combat athletes. Because athletes are systematically exposed to brief periods of intense pain during training or competition, they must learn to manage pain effectively. Therefore, knowledge of the genetic determinants of the physiological and psychological aspects of pain in athletes, especially martial artists, can potentially be used in the selection of new sports talent and provides an additional source of detailed information useful in personalizing training methods and more effectively managing the sports careers of trained athletes [5].
Sodium channels are integral membrane proteins and consist of a large α-subunit that forms the voltage-sensitive and ion-selective pore and a small β-helper subunit(s) that can regulate the kinetics and voltage dependence of channel gating [17]. To date, nine isoforms of the α-subunit of the sodium channel (NaV1.1–NaV1.9) have been described, but NaV1.7 appears to play a critical role in pain perception [14,18,19]. The role of these channels is central due to the generation and repetitive firing properties of the different neurons [20].
SCN9A is a 113.5 kb gene located on the long arm of chromosome 2 (2q24.3) and consisting of 26 exons. The protein consists of 1977 amino acids organized into four domains, each with six transmembrane segments, and is expressed at high density in peripheral sensory neurons, especially in nociceptive neurons [19]. Mutations in SCN9A that alter NaV1.7 function and are associated with various channelopathies leading to electrical hyperactivity of sensory neurons in the dorsal root and low reactivity of neurons in the sympathetic ganglia have been shown to be causative factors in various human pain perceptions [21]. It has been suggested that a common missense polymorphism in exon 18 of SCN9A (rs6746030; G>A; R1150W) may influence nociception in the general population. The minor A allele has been associated with increased NaV1.7 channel activity, resulting in increased pain ratings and lower pain threshold compared with the major G allele [15].
To date, there is no gold standard for pain assessment. This is a consequence of the specificity of this multidimensional sensory phenomenon, in which peripherally registered tissue-damaging stimuli are processed in the brain and supplemented by a subjective aspect. Most often, the perceived pain is still the effect of the current psychological state, social relations, well-being, etc. In such a situation, the use of experimental pain measurements, where it should be assumed that the experimental pain response is a stable trait, provides an opportunity to investigate the use of a genetic model to predict individual differences in experimental pain in healthy young people. In the current study, we significantly reduced the number of variables and aimed to evaluate the association between SCN9A genotypes and pain perception (pain threshold and pain tolerance) in the group of combat athletes and men who do not play sports professionally. Firstly, following the literature, we expected that the proportion of participants with low pain sensitivity (high pain threshold and high pain tolerance) would be higher among combat athletes than among men who do not play sports professionally. Secondly, since the A allele seems to confer enhanced pain sensitivity, we hypothesized that the chance of having low pain sensitivity will be higher for the group of participants with the GG homozygous genotype in comparison to the AA homozygous genotype and the GA heterozygous genotype. Thirdly, we had no specific expectations concerning whether these two variables would interact with each other, that is, whether the effect of physical activity would depend on the SCN9A gene.
2. Materials and Methods
2.1. Ethics Statement
The Pomeranian Medical University Ethics Committee (Szczecin, Poland) approved the study (no. 09/KB/V/2013). The investigation protocols were conducted ethically ac-cording to the World Medical Association Declaration of Helsinki and to the Strengthening the Reporting of Genetic Association studies statement (STREGA). The participants were informed of the risks and benefits of the experimental protocols, and a written consent form was completed by each participant. All personal information and results were anonymous.
2.2. Participants
For the study, 306 healthy Caucasian men aged 18 to 32 years were recruited. The experimental group included 214 combat athletes (24.7 ± 6.6), and the control group consisted of 92 students (21.2 ± 1.8) who did not practice sports professionally. The combat athletes group consisted of athletes who had at least 5 years of experience in disciplines such as boxing (n = 101), karate (n = 85), and mixed martial arts (n = 28).
2.3. Pressure Pain Test (PPT)
Tissue pressure sensitivity was measured using an algometer from Quirumed (Spain). The device is a force gauge ranging from 0 to 10 kg with a disc-shaped tip covered with a rubber sheath with an area of 1 cm2. Pain threshold and pain tolerance were measured.
Pain threshold (PPT1) is the lowest intensity of a particular stimulus—sound, heat, touch—at which a person begins to feel pain.
Pain tolerance (PPT2) is the maximum amount of pain a person can tolerate.
The results obtained by all subjects were divided into two categories below or above 10 kg. The recording of the result in the category below and above 10.1 kg/cm2 resulted from the measurement limits of the device.
Before starting the measurement of pressure sensitivity, each participant was informed about the purpose of the study and received detailed instructions on how to perform the study and how to behave. Three test measurements were then performed to demonstrate pain-inducing pressure.
The study was performed in a sitting position on the dominant extremity. Participants were asked to place their hand on the table. Initially, the researcher determined the contact site by palpation, then placed the pressure head on the back of the hand between the thumb and forefinger at a 90° angle and applied pressure with increasing force at a rate of 100 g/s to the selected site. The measurement results were visible only to the person conducting the test.
2.4. Genotyping
Genomic DNA was extracted from the buccal cells by a Genomic Micro AX SWAB Gravity (A&A Biotechnology, Poland) according to the producer’s protocol. An allelic discrimination assay on a C1000 Touch Thermal Cycler (Bio-Rad, Feldkirchen, Germany) instrument with TaqMan® probes was applied. To discriminate the SCN9A alleles, TaqMan® Pre-Designed SNP Genotyping Assays (Applied Biosystems, Waltham, MA, USA) (assay ID: C__29330435_10), consisting of fluorescently labelled (FAM and VIC) minor groove binder (MGB) probes and two specific primers, were used. All samples were genotyped in duplicate.
2.5. Statistical Analyses
Since distributions of most of the analyzed quantitative parameters were significantly different from the normal distribution (Shapiro–Wilk test), we used the non-parametric Mann–Whitney U-test to compare them between groups. Logistic regression conducted with JAMOVI was used to compare qualitative variables (pain threshold and pain tolerance) between genotype categories and between combat athletes and the control group [22]. A chi-squared test was used to test the Hardy–Weinberg equilibrium. p < 0.05 was considered statistically significant.
3. Results
The genotype frequencies of the SCN9A rs6746030 polymorphism did not differ from the Hardy–Weinberg expectations for both athletes (GG—77.6%, GA—20.6%, AA—1.8%, chi-squared = 0.29, p = 0.86) and controls (GG—75.0%, GA—23.9%, AA—1.1%, chi-squared = 0.27, p = 0.87).
The results of anthropometric measurements are presented in Table 1. The examined athletes were older and shorter compared to the students. They did not differ significantly in body mass. Body mass index (BMI) indicated the normal weight of all study participants, and the differences between groups were mainly due to differences in body height.
There were no significant differences between the SCN9A genotype distributions and pain perception, defined as pain threshold and pain tolerance, in the group of combat athletes. However, significant differences in the control group were found. The odds ratio of having decreased pain tolerance (PPT2 > 10.00 kg/cm2) for the GA and AA genotypes was more than three times higher (3.11, CI: 95% 1.17 to 8.25, p = 0.02) than for the GG genotype homozygotes (Table 2).
In Table 2, we present the proportion of combat athletes and controls with different SCN9A genotypes who demonstrated high and low pain threshold as well as pain tolerance. The model of binominal logistic regression with the experimental group (combat athletes vs. controls) and genotype (GG vs. GA + AA) as predictors and pain threshold (PPT1) as a dependent variable was significant, χ2(2) = 37.19, p < 0.001, R2 Nagerkelke = 18.5%. The effect of the experimental group was significant, B = 3.47, SE = 1.02, Z = 3.41, p < 0.001, OR = 32.25, 95%CI (4.39, 236.92), meaning that the probability of having a high pain threshold was higher in the combat athletes’ group (probability = 0.26, SE = 0.04, 95%CI (0.20, 0.34)) than in controls (probability = 0.01, SE = 0.01, 95%CI = (0.002, 0.07)). The effect of the genotype was not significant, B = 0.03, SE = 0.37, Z = 0.07, p = 0.944, OR = 1.03, 95%CI (0.50, 2.11) (see also Figure 1). Adding an interaction between the two predictors did not improve the model significantly, χ2(1) = 2.84, p = 0.092, meaning that the difference in pain threshold between athletes and controls was independent of their genotype.
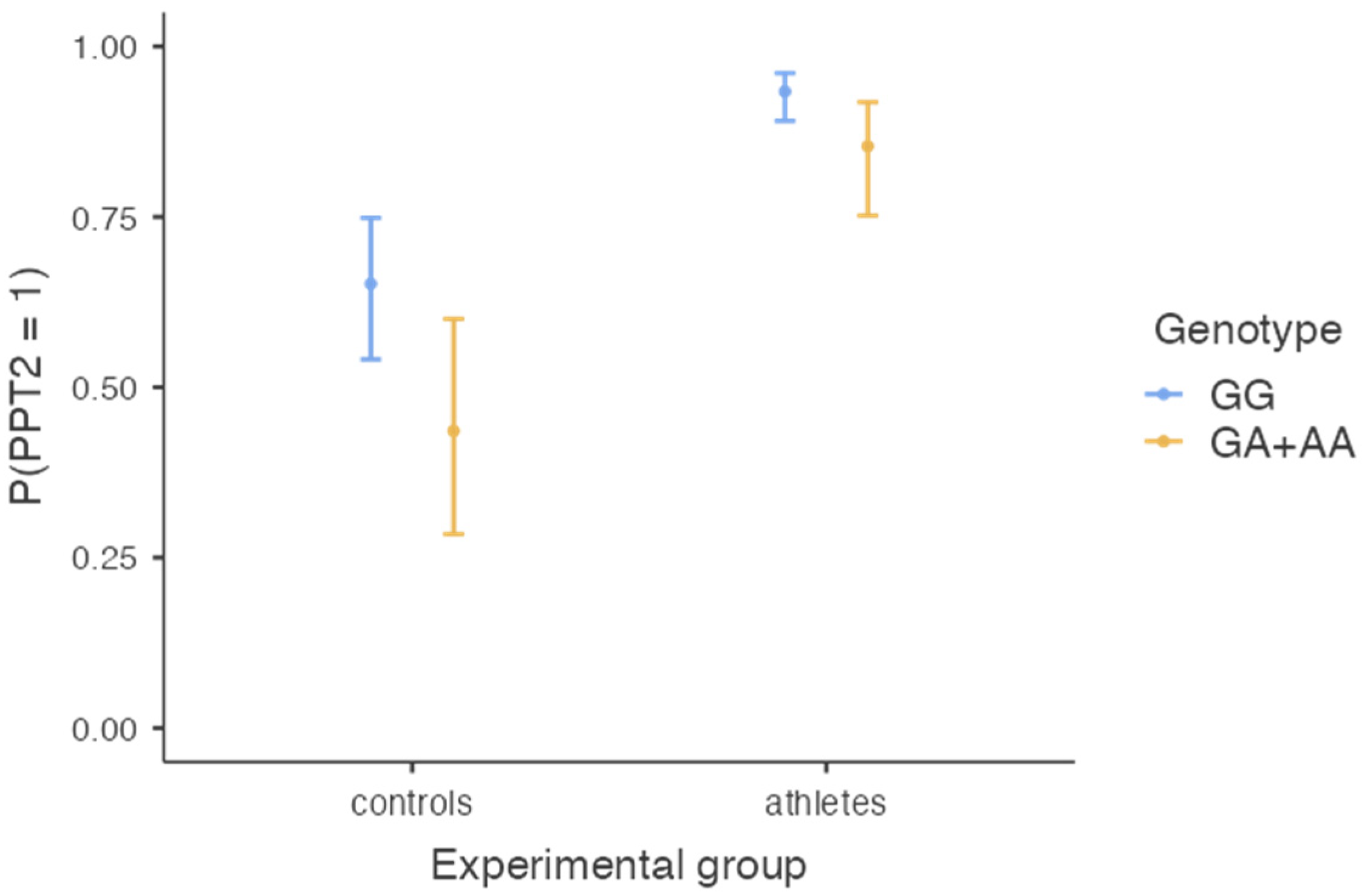
The model of binominal logistic regression with the experimental group (combat athletes vs. controls) and genotype (GG vs. GA + AA) as predictors and pain tolerance (PPT2) as a dependent variable was significant, χ2(2) = 46.55, p < 0.001, R2 Nagerkelke = 23.2%. The effect of the experimental group was significant (B = 2.02, SE = 0.331, Z = 6.10, p < 0.001, OR = 7.57, 95%CI (3.94, 14.45)), meaning that the probability of having high pain tolerance was higher in the combat athletes’ group (probability = 0.90, SE = 0.02, 95%CI (0.85, 0.94)) than in controls (probability = 0.55, SE = 0.06, 95%CI = (0.43, 0.66)). The effect of the genotype was also significant (B = −0.88, SE = 0.36, Z = −2.45, p = 0.014, OR = 0.41, 95% CI (0.20, 0.84)), meaning that those with GA and AA genotypes were less likely to have high pain tolerance (probability = 0.68, SE = 0.07, 95%CI (0.54, 0.79)) than those with the GG genotype (probability = 0.84, SE = 0.03, 95%CI (0.78, 0.88)) (see also Figure 2). Adding an interaction between the two predictors did not improve the model significantly, χ2(1) = 0.54, p = 0.463, meaning that the difference in pain tolerance between athletes and controls was independent of their genotype, and the difference between genotypes was independent of whether the participant was a combat athlete or not.
4. Discussion
The aim of our study was to determine (1) whether the proportion of participants with low pain sensitivity (PPT1 and PPT2) was higher in combat athletes than in men who do not play sports professionally and (2) whether the probability of low pain sensitivity (PPT1 and PPT2) was higher in the group of participants with the GG genotype than in the participants with AA and GA genotypes. The results showed that the probability of having increased pain threshold and pain tolerance was higher in the group of combat athletes than in the control group, and thus we confirmed the first hypothesis. Our results are consistent with the literature, which states that physically active people, including athletes, have higher pain tolerance [2,3,5,17]. This variable may be modulated by numerous factors, such as the systematic exposure of the body of athletes to short-term high-intensity stimuli during training sessions, more frequent injuries, and significantly more frequent stimulation of the nervous system compared to non-athletes. From this perspective, the intensity of physical activity appears to be inextricably linked to pain perception. For athletes, pain is an integral part of their lifestyle. Many hours of training and participation in competitions, in this case, strenuous fights, classify combat athletes in particular as a group exposed to high pain intensity [5]. Consequently, many studies have confirmed that combat athletes have a higher pain tolerance than active controls and non-combat athletes [23,24]. Participants in a sport that requires contact face the additional challenge of being intentionally injured by other players. This may present an additional external mechanical stimulus and would activate pain pathways associated with skin and muscle deformation that may or may not be associated with tissue damage, rather than those that are natural in the muscle during exercise. The combination of these experiences might indeed lead to altered pain perception [5,24,25], as we have shown in this study.
To our knowledge, the present study is the second study to investigate the association between the rs6746030 polymorphism in the SCN9A gene and pain perception in combat athletes and people with low physical activity. The SCN9A gene encodes the NaV1.7 α-subunit isoform of the sodium channel. Ion channels, which are responsible for cell excitability and thus intercellular communication, play an important role in the biological mechanisms that generate and maintain neuropathic pain. Of the three voltage-gated channels (sodium, calcium, and potassium channels) directly involved in regulating membrane potentials, sodium channels appear to be the most important targets for pain [7]. Two genome-wide association studies (GWAS) performed in primary erythromelalgia patients in 2004 and 2005 revealed gain-of-function mutations in the SCN9A gene [26,27]. This association was confirmed by Nassar et al. [28] who knocked out of the Scn9a gene in mice. They showed that the mutants had reduced thermal and mechanical sensitivity under initial and inflammatory conditions [28]. Next, Cox et al. [13] sequenced chromosomal region 2q24.3, containing the SCN9A gene, from three consanguineous families with congenital insensitivity to pain. Sequence analysis revealed that channelopathy in SCN9A is a critical mediator of nociceptive pain [21]. Given the confirmed association between the SCN9A polymorphism rs6746030 and alterations in pain signaling in humans, this genetic variant has been extensively studied in several human genetic pain syndromes [15,16,29]. However, its effect on pain perception in athletes is almost unknown. Our study showed that the difference in pain threshold between athletes and control subjects was not related to the SCN9A genotype, but the effect of genotype on pain tolerance was significant. Specifically, all participants in the experimental group with genotypes GA and AA had lower pain tolerance than those with genotype GG. The obtained results are not in agreement with our previous study performed on 101 male boxers and 332 control participants, in which no association between the SCN9A rs6746030 polymorphism and phenotypic pain variables was found [30]. This confirms that the genetic background of pain is very complex and that we are only at the beginning of understanding the role of SCN9A variations in the athletes’ sensitivity to pain.
The results obtained in the present study support previous findings suggesting that carriers of the genotypes GA and AA have lower pain tolerance. The explanation for this could be as follows. The substitution of G to A in exon 18 of SCN9A results in a switch from positively charged arginine (R) to nonpolar tryptophan (W) at residue 1150 [31]. This substitution is located in a highly conserved sequence of the C-terminus of L2, the loop connecting the II and III domains of the Chan core. Considering the high conservation of the residue of the encoded protein from different species and the presence of a positively charged residue at this site in almost all other sodium channels, Estacion et al. [14] suggested that the substitution by the tryptophan residue has a functional impact on the biophysical properties of the NaV1.7 channel, which is known to play a key role in pain perception. Current-clamp analysis showed that the A (1150 W) allele depolarizes the resting membrane potential (6 mV) and increases the firing frequency twofold in response to depolarization in neurons of the spinal ganglion [14]. This observation was confirmed by Reimann et al. [15] who described a significant association between rs6746030 and pain score in patients with osteoarthritis, sciatica, phantom limb pain, lumbar discectomy, and pancreatitis, as well as in healthy women characterized by their responses to a variety of different noxious stimuli. Finally, the genotypes GA and AA were found to be associated with increased NaV1.7 activity and consequently altered pain threshold [14,15].
Current evidence, confirmed by the results of scientific research, including ours, shows that other factors such as physical activity significantly increases the pain threshold and pain tolerance, and that this effect is independent of the effect of the SCN9A genotypes. Thus, it appears that the effect of genotype on pain sensitivity can be modified by environmental factors such as exercise. Both pain threshold and pain tolerance need not be constant values for a given individual but can be modified by among others the environment. Additionally, it needs to be highlighted that pain phenotype is a polygenic trait. To date, about 252 genes have been linked to pain-associated behaviors [7]. It is more likely that several gene loci, each with a small but significant contribution, are responsible for this genetic component. Indeed, single-marker analysis is likely to make only a limited, but still important, contribution to understanding the pain perception.
5. Limitation
We are aware that our study has some limitations. Objective pain analyses are problematic because they are subjective and it is difficult to develop reliable scales to characterize pain. Moreover, in terms of the high emotional variability of individuals, it seems almost impossible to assess the quantitative and qualitative nature of pain. The weak point of the study was the algometer used to measure sensitivity to pain, which allowed the measurement to a value of 10.1 kg/cm2. In the case of the genetic association study, the relatively small group sample was the most important limitation. We studied a relatively small sample of Polish combat athletes. However, this group had an advantage—it included the best Poles with a uniform athletic level. Therefore, it is necessary to validate our results in larger cohorts of athletes as well as of different ethnicities.
6. Conclusions
The results of our study indicate that
- -
- the combat athletes have increased pain threshold and pain tolerance;
- -
- the SCN9A rs6746030 polymorphism may affect pain perception, in particular
- -
- the GA and AA genotypes may be associated with lower pain tolerance compared with the GG genotype;
- -
- the other factors, such as regular physical activity, significantly increases the pain threshold and pain tolerance, and this effect is independent of the effect of the SCN9A genotypes.
These results support the hypothesis that pain phenotype is a complex trait, important for achieving success in combat sports. Although the SCN9A rs6746030 polymorphism seems to be an important genetic marker for pain perception in athletes, more replication studies are needed.
Author Contributions
Conceptualization, K.L. and A.L.-D.; methodology, M.S. and M.P.; software, A.G. and A.L.-D.; validation, K.L., M.S. and M.P.; formal analysis, A.G. and M.S.; investigation, A.G. and A.L.-D.; resources, M.P.; data curation, K.L.; writing—original draft preparation, K.L. and A.L.-D.; writing—review and editing, A.G., K.L. and M.S.; visualization, M.P.; supervision, A.G. and A.L.-D.; project administration, M.P. and A.L.-D.; funding acquisition, A.L.-D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The Pomeranian Medical University Ethics Committee, Poland, approved the study (no. 09/KB/V/2013). The investigation protocols were conducted ethically according to the World Medical Association Declaration of Helsinki and to the Strengthening the Reporting of Genetic Association studies statement (STREGA).
Informed Consent Statement
Informed consent was obtained from all subjects or their parents if the subject was under 18 years of age involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy/ethical restrictions.
Acknowledgments
The preliminary results were previously presented at the International Conference of Science and Methodology ‘Identification and Development of Sport Talents’, 19–21 November 2021 in Gdansk.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bonica, J.J. Editorial The need of a taxonomy. Pain 1979, 6, 247–252. [Google Scholar] [CrossRef]
- Pawlak, M.; Jażdżewska, A.; Leźnicka, K. Can physical activity modulate the pain perception during the ontogenesis? Balt. J. Health Phys. Act. 2019, 11, 90–100. [Google Scholar] [CrossRef]
- Heneweer, H.; Vanhees, L.; Picavet, H. Physical activity and low back pain: A U-shaped relation? Pain 2009, 143, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Lie, M.U.; Winsvold, B.; Gjerstad, J.; Matre, D.; Pedersen, L.M.; Heuch, I.; Zwart, J.A.; Nilsen, K.B. The association between selected genetic variants and indi-vidual differences in experimental pain. Skand. J. Pain 2020, 21, 163–173. [Google Scholar] [CrossRef]
- Leźnicka, K.; Pawlak, M.; Białecka, M.; Safranow, K.; Spieszny, M.; Klocek, T.; Cieszczyk, P. Evaluation of the pain threshold and tolerance of pain by martial arts athletes and non-athletes using a different methods and tools. Arch. Budo 2016, 12, 239–245. [Google Scholar]
- Fillingim, R.B. Individual differences in pain: Understanding the mosaic that makes pain personal. Pain 2017, 158 (Suppl. 1), S11–S18. [Google Scholar] [CrossRef] [Green Version]
- Wistrom, E.; Chase, R.; Smith, P.R.; Campbell, Z.T. A compendium of validated pain genes. WIREs Mech. Dis. 2022, 14, e1570. [Google Scholar] [CrossRef]
- Mogil, J.S.; Yu, L.; Basbaum, A.I. Pain Genes? Natural Variation and Transgenic Mutants. Annu. Rev. Neurosci. 2000, 23, 777–811. [Google Scholar] [CrossRef] [Green Version]
- Foulkes, T.; Wood, J.N. Pain genes. PLoS Genet. 2008, 4, e1000086. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Auer-Grumbach, M.; Matsukawa, S.; Zitzelsberger, M.; Themistocleous, A.C.; Strom, T.M.; Samara, C.; Moore, A.W.; Cho, L.T.-Y.; Young, G.T.; et al. Transcriptional regulator PRDM12 is essential for human pain perception. Nat. Genet. 2015, 47, 803–808. [Google Scholar] [CrossRef]
- Mollazadeh, H.; Cicero, A.F.G.; Blesso, C.N.; Pirro, M.; Majeed, M.; Sahebkar, A. Immune modulation by curcumin: The role of interleukin-10. Crit. Rev. Food Sci. Nutr. 2019, 59, 89–101. [Google Scholar] [CrossRef]
- Bagal, S.K.; Chapman, M.L.; Marron, B.E.; Prime, R.; Storer, R.I.; Swain, N.A. Recent progress in sodium channel modulators for pain. Bioorg. Med. Chem. Lett. 2014, 24, 3690–3699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, D.L.H.; Woods, C.G. Painful and painless channelopathies. Lancet Neurol. 2014, 13, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Estacion, M.; Harty, T.P.; Choi, J.S.; Tyrell, L.; Dib-Hajj, S.D.; Waxman, S.G. A sodium channel gene SCN9A polymorphism that increases nociceptor excitability. Ann. Neurol. 2009, 66, 862–866. [Google Scholar]
- Reimann, F.; Cox, J.J.; Belfer, I.; Diatchenko, L.; Zaykin, D.; McHale, D.P.; Nikolajsen, L.; Max, M.B.; Kiselycznyk, C.; Woods, C.G.; et al. Pain perception is altered by a nucleotide polymorphism in SCN9A. Proc. Natl. Acad. Sci. USA 2010, 107, 5148–5153. [Google Scholar] [CrossRef] [Green Version]
- Drenth, J.P.H.; Waxman, S.G. Mutations in sodium-channel gene SCN9A cause a spectrum of human genetic pain disorders. J. Clin. Investig. 2007, 117, 3603–3609. [Google Scholar] [CrossRef]
- Catterall, W.A. Structure and Regulation of Voltage-Gated Ca2+ Channels. Annu. Rev. Cell Dev. Biol. 2000, 16, 521–555. [Google Scholar] [CrossRef]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology. XLVII. Nomenclature and Structure-Function Relationships of Voltage-Gated Sodium Channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef]
- Toledo-Aral, J.J.; Moss, B.L.; He, Z.J.; Koszowski, A.G.; Whisenand, T.; Levinson, S.R.; Wolf, J.J.; Silos-Santiago, I.; Halegoua, S.; Mandel, G. Identification of PN1, a predominant volt-age-dependent sodium channel expressed principally in peripheral neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 1527–1532. [Google Scholar] [CrossRef] [Green Version]
- Amir, R.; Argoff, C.E.; Bennett, G.J.; Cummins, T.R.; Durieux, M.E.; Gerner, P.; Gold, M.S.; Porreca, F.; Strichartz, G.R. The Role of Sodium Channels in Chronic Inflammatory and Neuropathic Pain. J. Pain 2006, 7, S1–S29. [Google Scholar] [CrossRef]
- Cox, J.J.; Reimann, F.; Nicholas, A.K.; Thornton, G.; Roberts, E.; Springell, K.; Karbani, G.; Jafri, H.; Mannan, J.; Raashid, Y.; et al. An SCN9A channelopathy causes congenital inability to experience pain. Nature 2006, 444, 894–898. [Google Scholar] [CrossRef] [PubMed]
- The Jamovi Project 2021. Jamovi (version 2.2); Computer Software. Available online: https://www.jamovi.org (accessed on 3 February 2023).
- Tesarz, J.; Schuster, A.K.; Hartmann, M.; Gerhardt, A.; Eich, W. Pain perception in athletes compared to normally active controls: A systematic review with meta-analysis. Pain 2012, 153, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, M. Aspects of pain in sport. Trends Sport Sci. 2013, 20, 123–134. [Google Scholar]
- Van Middelkoop, M.; Rubinstein, S.; Kuijpers, T.; Verhagen, A.P.; Ostelo, R.; Koes, B.; van Tulder, M. A systematic review on the effectiveness of physical and rehabilitation interventions for chronic non-specific low back pain. Eur. Spine J. 2011, 20, 19–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dib-Hajj, S.D.; Rush, A.M.; Cummins, T.R.; Hisama, F.M.; Novella, S.; Tyrrell, L.; Marshall, L.; Waxman, S.G. Gain-of-function mutation in Nav1.7 in familial erythromelalgia induces bursting of sensory neurons. Brain 2005, 128, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, Y.; Li, S.; Xu, Z.; Li, H.; Ma, L.; Fan, J.; Bu, D.; Liu, B.; Fan, Z.; et al. Mutations in SCN9A, encoding a sodium channel α subunit, in patients with primary erythermalgia. J. Med. Genet. 2004, 41, 171–174. [Google Scholar] [CrossRef]
- Nassar, M.A.; Stirling, L.C.; Forlani, G.; Baker, M.D.; Matthews, E.A.; Dickenson, A.H.; Wood, J.N. Nociceptor-specific gene deletion reveals a major role for Na. Proc. Natl. Acad. Sci. USA 2004, 101, 12706–12711. [Google Scholar] [CrossRef] [Green Version]
- Kurzawski, M.; Rut, M.; Dziedziejko, V.; Safranow, K.; Machoy-Mokrzynska, A.; Drozdzik, M.; Białecka, M. Common Missense Variant of SCN9A Gene Is Associated with Pain Intensity in Patients with Chronic Pain from Disc Herniation. Pain Med. 2018, 19, 1010–1014. [Google Scholar] [CrossRef]
- Leźnicka, K.; Gronek, P.; Kurzawski, M.; Cięszczyk, P.; Malinowski, D.; Żmijewski, P. Is pain tolerance in boxers altered by nucleotide polymorphism rs6746030 in the SCN9A gene? Arch. Budo 2018, 14, 71–78. [Google Scholar]
- Klugbauer, N.; Lacinova, L.; Flockerzi, V.; Hofmann, F. Structure and functional expression of a new member of the tetro-dotoxinsensitive voltage-activated sodium channel family from human neuroendocrine cells. Eur. Mol. Biol. Org. J. 1995, 14, 1084–1090. [Google Scholar] [CrossRef]
Figure 1.
The chance of having high pain threshold (PPT1 > 10.0 kg/cm2) as a function of the SCN9A genotype and experimental group.
Figure 1.
The chance of having high pain threshold (PPT1 > 10.0 kg/cm2) as a function of the SCN9A genotype and experimental group.

Figure 2.
The chance of having high pain tolerance (PPT2 > 10.0 kg/cm2) as a function of the SCN9A genotype and experimental group.
Figure 2.
The chance of having high pain tolerance (PPT2 > 10.0 kg/cm2) as a function of the SCN9A genotype and experimental group.


{kind=link}
{kind=link}
Table 1.
Demographic and anthropometric data of the combat athletes and control group.
| Variables | Combat Athletes (n = 214) Mean ± SD | Control Group (n = 92) Mean ± SD | p-Value |
|---|---|---|---|
| Age (years) | 24.67 ± 6.57 | 21.21 ± 1.82 | 0.008 |
| Height (cm) | 178.6 ± 7.04 | 182.53 ± 8.04 | <0.001 |
| Body mass (kg) | 78.21 ± 13.15 | 78.25 ± 10.19 | 0.956 |
| BMI (kg/m2) | 24.46 ± 3.33 | 23.43 ± 2.06 | 0.050 |
p-values evaluated by means of the Mann–Whitney U-test. Mean and standard deviations are given.
Table 2.
The SCN9A genotype in relation to PPT measurements in combat athletes and control group.
| Pain Threshold and Pain Tolerance | Combat Athletes (n = 214) | Control Group (n = 92) | |||
|---|---|---|---|---|---|
| GG | GA + AA | GG | GA + AA | ||
| PPT1 ≤ 10.0 kg/cm2 | n | 122 | 36 | 69 | 22 |
| % | 73.5 % | 75.0% | 100.0% | 95.7% | |
| PPT1 > 10.0 kg/cm2 | n | 44 | 12 | 0 | 1 |
| % | 26.5 % | 25.0% | 0.0% | 4.3% | |
| PPT2 ≤ 10.0 kg/cm2 | n | 12 | 6 | 23 | 14 |
| % | 7.2 % | 12.5% | 33.3% | 60.9% | |
| PPT2 > 10.0 kg/cm2 | n | 154 | 42 | 46 | 9 |
| % | 92.8 % | 87.5% | 66.7% | 39.1% | |
PPT1—pain threshold; PPT2—pain tolerance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Leźnicka, K.; Pawlak, M.; Sawczuk, M.; Gasiorowska, A.; Leońska-Duniec, A. SCN9A rs6746030 Polymorphism and Pain Perception in Combat Athletes and Non-Athletes. Genes 2023, 14, 733. https://doi.org/10.3390/genes14030733
AMA Style
Leźnicka K, Pawlak M, Sawczuk M, Gasiorowska A, Leońska-Duniec A. SCN9A rs6746030 Polymorphism and Pain Perception in Combat Athletes and Non-Athletes. Genes. 2023; 14(3):733. https://doi.org/10.3390/genes14030733
Chicago/Turabian StyleLeźnicka, Katarzyna, Maciej Pawlak, Marek Sawczuk, Agata Gasiorowska, and Agata Leońska-Duniec. 2023. "SCN9A rs6746030 Polymorphism and Pain Perception in Combat Athletes and Non-Athletes" Genes 14, no. 3: 733. https://doi.org/10.3390/genes14030733
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.