
Transfer RNA Modification Enzymes with a Thiouridine Synthetase, Methyltransferase and Pseudouridine Synthase (THUMP) Domain and the Nucleosides They Produce in tRNA


Department of Materials Science and Biotechnology, Graduate School of Science and Engineering, Ehime University, Matsuyama 790-8577, Japan
Genes 2023, 14(2), 382; https://doi.org/10.3390/genes14020382
Submission received: 13 December 2022
/
Revised: 25 January 2023
/
Accepted: 27 January 2023
/
Published: 31 January 2023
(This article belongs to the Special Issue Transfer RNA Modification)
Abstract
:The existence of the thiouridine synthetase, methyltransferase and pseudouridine synthase (THUMP) domain was originally predicted by a bioinformatic study. Since the prediction of the THUMP domain more than two decades ago, many tRNA modification enzymes containing the THUMP domain have been identified. According to their enzymatic activity, THUMP-related tRNA modification enzymes can be classified into five types, namely 4-thiouridine synthetase, deaminase, methyltransferase, a partner protein of acetyltransferase and pseudouridine synthase. In this review, I focus on the functions and structures of these tRNA modification enzymes and the modified nucleosides they produce. Biochemical, biophysical and structural studies of tRNA 4-thiouridine synthetase, tRNA methyltransferases and tRNA deaminase have established the concept that the THUMP domain captures the 3′-end of RNA (in the case of tRNA, the CCA-terminus). However, in some cases, this concept is not simply applicable given the modification patterns observed in tRNA. Furthermore, THUMP-related proteins are involved in the maturation of other RNAs as well as tRNA. Moreover, the modified nucleosides, which are produced by the THUMP-related tRNA modification enzymes, are involved in numerous biological phenomena, and the defects of genes for human THUMP-related proteins are implicated in genetic diseases. In this review, these biological phenomena are also introduced.
1. Introduction
To date, more than 150 modified nucleosides have been found in RNAs from the three domains of life [1]. Transfer RNA contains numerous modified nucleosides [2,3] and the majority of modified nucleosides in tRNA are introduced by site-specific tRNA modification enzymes. Transfer RNA modification enzymes frequently contain one or more distinct domains in addition to the catalytic domain, although small tRNA methyltransferases such as TrmL [4,5] and TrmH [6,7] are mainly composed of the catalytic domain [8,9,10]. The existence of the thiouridine synthetase, methyltransferases and pseudouridine synthase (THUMP) domain was originally predicted in a bioinformatic study [11]. In this study in 2001, Aravind and Koonin reported that tRNA 4-thiouridine synthetase-like proteins, conserved RNA methyltransferases, archaeal pseudouridine synthases and several uncharacterized proteins share a predicted RNA binding domain, which adopts an α/β fold [11]. At that time, although the Escherichia coli thiI gene product had already been identified as a tRNA 4-thiouridine synthetase [12], functions of the other proteins were unknown. Furthermore, no structures for any of the proteins, including ThiI, had been reported. In 2004, the Pyrococcus abyssi PAB1283 protein was firstly identified as a tRNA methyltransferase, which contains a THUMP domain [13]. Because the PAB1283 protein possesses enzymatic activity for the formation of N2-methylguanosine (m2G) and N2, N2-dimethylguanosine (m22G) at position 10 in tRNA, nowadays, the PAB1283 protein is called archaeal Trm11 (arcTrm11). At the same time, the Saccharomyces cerevisiae tan1 gene product was found to be an essential protein for the formation of N4-acetylcytidine at position 12 (ac4C12) in tRNALeu and tRNASer [14]. Although Tan1 contains a THUMP domain, this protein itself does not possess tRNA acetyltransferase activity [14] and does not contain a catalytic domain [15]. Later, Tan1 was identified as a partner protein of S. cerevisiae tRNA acetyltransferse (Kre33) [16]. Since the prediction of the THUMP domain more than two decades ago, many tRNA modification enzymes containing a THUMP domain have been identified. Among them, in addition to tRNA 4-thiouridine synthetases, tRNA methyltransferases, tRNA pseudoridine synthases, tRNA deaminase [17] and a partner protein of tRNA acetyltransferases [16] have been identified. In this review, I focus on functions and structures of these tRNA modification enzymes and the modified nucleosides they produce. Several THUMP-related proteins are involved in not only tRNA modification but also modifications of other RNAs such as rRNA [16,18,19]. In these cases, appropriate reviews and representative articles are introduced due to the limitation of space in this review.
2. Classification of THUMP-Related tRNA Modification Enzymes
According to enzymatic activity, THUMP-related tRNA modification enzymes can be classified into five types: 4-thiouridine synthetase, deaminase, methyltransferase, a partner protein of acetyltransferase and pseudouridine synthase (Table 1). As described in a later section, although the classification of tRNA (m2G/m22G) methyltransferases is complicated, archaeal and eukaryotic Trm11-Trm112 are combined in one column in Table 1. The modification positions and structures of modified nucleosides, which are produced by THUMP-related tRNA modification enzymes, are summarized in Figure 1. TkTHUMDP1-TkNAT10 modify multiple positions in tRNA. For example, when T. kodakarensis cells were cultured at 95 °C, C12, C35 and C56 in tRNALeu were modified to ac4C12, ac4C35 and ac4C56, respectively, by TkTHUMDP1-TkNAT10 [18]. The crystal structure of human PUS10 [20] and a structural model of archaeal Pus10 [21] show that the THUMP domain-related structure is contained in the N-terminal accessory domain. The accessory domain is considerably larger than the THUMP domain in other THUMP-related tRNA modification enzymes.
The biosynthesis pathways of modified nucleosides by THUMP-related tRNA methyltransferases are summarized in Figure 2.
3. Effect of the Modified Nucleosides, Which Are Produced by THUMP-Related tRNA Modification Enzymes, on tRNA Structure
All modified nucleosides, which are produced by THUMP-related tRNA modification enzymes, are considered to stabilize the L-shaped tRNA structure.
3.1. s4U8 and s4U9
The sulfur atom in s4U strengthens the hydrophobic interaction. The melting temperature of tRNASer from an E. coli thiI gene deletion strain decreases by 4.7 °C as compared to that from the wild-type strain [32]. Therefore, at least s4U8 stabilizes the L-shaped tRNA structure. The effect of s4U9 modification on the tRNA structure is unknown.
3.2. U8
U8 is a conserved nucleoside in tRNA and forms a reverse Hoogsteen tertiary base pair with A14 [33]. Therefore, deamination from C8 to U8 is essential for maintenance of the L-shaped tRNA structure [17].
3.2.1. m2G10 and m22G10
The m2G modification does not disturb the formation of the Watson–Crick base pair with C. The O6 atom of m2G10 in the m2G10-C25 base pair forms a hydrogen bond with the amino group of G45 in S. cerevisiae tRNAPhe. Furthermore, the m2G10-C25 base pair stacks with the m22G26-A44 tertiary base pair. The methyl group in m2G10 probably stabilizes this stacking effect. In contrast, m22G cannot form a Watson–Crick base pair with C. Instead, m22G forms a non-Watson–Crick base pair with U, and the m22G10-U25 base pair can be observed in T. kodakakrensis tRNATrp [34]. The two methyl groups in m22G probably stabilize the stem structure when an m22G-U base pair is formed. Furthermore, the m22G10 modification prevents the formation of an incorrect Watson–Crick base pair in tRNA [35].
3.2.2. m2G6, m2G7 and m2G67
As described above, the m2G modification does not disturb the formation of the Watson–Crick base pair with C. Although the methyl group in these modified nucleosides probably stabilizes the aminoacyl-stem structure in tRNA, the effect has not been confirmed experimentally.
3.3. ac4C12 and ac4C Modifications in Other Positions
The ac4C modification tilts the equilibrium of ribose puckering towards the C3′ endo-form [36]. Furthermore, the ac4C modification in a stem structure increases the melting temperature of the stem [37]. Therefore, ac4C at position12 and other positions probably stabilizes the L-shaped tRNA structure and codon-anticodon interaction.
3.4. Ψ54 and Ψ55
The Ψ55 modification is highly conserved in tRNAs from the three domains of life and form a tertiary base pair with G18 in the L-shaped tRNA structure. The presence of Ψ55 enhances the affinity between the T-arm and the D-arm [38]. Although the structural effect of Ψ54 has not been confirmed experimentally, Ψ54 probably forms a tertiary base pair with A58 (or m1A58) and the Ψ54-A58 (m1A58) base pair stacks with the G53-C61 base pair in the T-stem. Thus, the Ψ54 modification probably stabilizes the tRNA structure.
4. Structures and Enzymatic Properties of THUMP-Related tRNA Modification Enzymes
In this section, the structures of THUMP-related tRNA modification enzymes and their enzymatic properties are introduced. As described below, the THUMP domain captures the 3′-end of RNA (in the case of tRNA, the CCA-terminus). This concept is proposed based on structural, biophysical and biochemical studies of ThiI and is extended to studies of other THUMP-related tRNA modification enzymes.
4.1. 4-Thiouridine Synthetase (ThiI)
When the existence of the THUMP domain was predicted [11], ThiI was the only identified tRNA modification enzyme in the list of predicted THUMP-related proteins. ThiI is a tRNA s4U synthetase [12]. s4U is found at positions 8 and 9 in tRNAs from eubacteria and archaea (Figure 1) [1,2,3]. The biosynthesis pathways of s4U are different in eubacteria and archaea [39,40,41,42]. In E. coli, the sulfur atom in L-cysteine is activated by cysteine desulfrase (IscS) and is then transferred to tRNA by ThiI in the presence of ATP [43,44,45]. Cysteine residues at positions 344 and 456 in E. coli ThiI are essential for the reaction and these residues are considered to form a disulfide bond in the catalytic turnover [46,47]. In contrast, the iscS gene is not encoded in the majority of archaea genomes [48]. In the case of Methanococcus maripuludis, ThiI contains an Fe-S cluster and S2− is used as a sulfur donor instead of L-cysteine [22,48]. However, the Fe-S cluster type thiI gene is not present in some archaea genomes and the biosynthesis pathways in these organisms are still unknown [39,48,49]. During the submission of this manuscript, it was reported that M. maripuldis and P. furiosus ThiI proteins possess a [4Fe-4S] cluster [50]. Furthermore, it has been proposed that these enzymes be renamed TtuI [50].
In 2006, the crystal structure of Bacillus anthracis ThiI (PDB code: 2C5S) was the first of the THUMP-related proteins to be reported (Figure 3A) [51]. B. anthracis ThiI contains three domains, an N-terminal ferredoxin-like domain (green), a THUMP domain (red) and a C-terminal PP-loop domain (blue) (Figure 3A). This structure revealed that the THUMP domain is composed of α-helices and β-strands as predicted. A tRNA binding model was also constructed in this study [51]. In the model, the THUMP domain of ThiI was placed near the CCA-terminus of tRNA because it was reported that the CCA-terminus was essential for the sulfur-transfer reaction of ThiI [52]. Later, this idea was experimentally verified by biochemical and structural studies of truncated tRNA [53] and ThiI-truncated tRNA complex [54]. The N-terminal ferredoxin-like domain functions to maintain the distance and angle between the THUMP and PP-loop domains. The PP-loop was originally found as a P-loop-like sequence motif, which had been observed in ATP pyrophosphatases [55]. The PP-loop domain in ThiI binds ATP and activates tRNA by adenylation [56,57]. At the same time that the crystal structure of B. anthracis ThiI was solved, the structure of Pyrococcus horikoshii PH1313 protein (PDB code: 1VBK) was released as a protein of unknown function (Figure 3B) [58]. In the Pyrococcus genera, multiple genes for ThiI homologs are often encoded in their genomes [22]. Because ThiI is involved in thiamine biosynthesis in addition to s4U modification in tRNA [12,59,60,61], the ThiI homologs in Pyrococcus may not have a dual function but instead individual proteins have single roles. Although the structure of the PH1313 protein (Figure 3B) resembles other ThiI proteins, the PH1313 protein lacks several conserved amino acid residues of ThiI proteins. To date, the enzymatic activity of the PH1313 protein has not been confirmed. Furthermore, modified nucleosides in tRNAs from P. horikoshii have not been analyzed [62]. Therefore, in this review, the PH1313 protein is described as a ThiI-like protein. The THUMP domain in the P. horikoshii ThiI-like protein is also composed of α-helices and β-strands as predicted.
Transfer RNA modification enzymes often recognize local structure(s) in tRNA [63]. Therefore, tRNA modification enzymes are frequently able to modify a truncated tRNA. For example, E. coli TrmA [64,65], E. coli TruB [66], E. coli Tgt [67,68], T. thermophilus TrmFO [69], T. thermophilus TrmI [70] and A. aeolicus TrmD [71] can modify a micro-helix RNA, which mimics the T-arm or anticodon-arm of substrate tRNA. TrmA, TruB, Tgt, TrmFO, TrmI and TrmD are tRNA (m5U54) methyltransferase [72], tRNA (Ψ55) synthase [73], tRNA guanine-transglycosylase [67,74,75,76], N5, N10-methylenetetrahydrofolate-dependent-tRNA (m5U54) methyltransferase [77], tRNA (m1A58) methyltransferase [78] and tRNA (m1G37) methyltransferase [79], respectively. Furthermore, E. coli TrmJ [80], A. aeolicus TrmB [81] and T. thermophilus TrmH [82] can methylate a truncated tRNA. TrmJ, TrmB and TrmH are tRNA (Cm32/Um32) methyltransferase [83], tRNA (m7G46) methyltransferase [84] and tRNA (Gm18) methyltransferase [6,85], respectively.
Lauhon et al. have reported that a truncated tRNAPhe (Figure 4A) is a minimum substrate for E. coli ThiI [52]. This truncated tRNAPhe is also recognized by Thermotoga maritima ThiI as a substrate [54]. The crystal structure of the complex of the minimum substrate RNA and T. maritima ThiI has been reported (Figure 4B) [54]. T. maritima ThiI forms a dimer and two minimum substrate RNAs bind to this dimer. The THUMP domain in one subunit captures the CCA terminus of one minimum substrate RNA and the PP-loop domain in this subunit accesses the modification site (U8) in another minimum substate RNA. Thus, this complex structure demonstrates that ThiI acts as a dimer. The disulfide bond, which acts in the catalytic cycle, in E. coli ThiI is formed within a single subunit [86]. Furthermore, this structure proposes a concept that the THUMP domain recognizes the 3′-end of RNA (in the case of tRNA, the CCA terminus).
4.2. Deaminase
M. kandleri is a hyper-thermophilic archaeon in which position 8 in 30 tRNA genes is encoded as C [87,88]. This C8 is modified to U8 by deamination (C to U editing) [17]. For further information about deamination in tRNA, see this review [89]. The enzyme responsible for deamination of C8 is CDAT8. CDAT8 can modify C8 in a micro-helix RNA (Figure 5A). A crystal structure of CDAT8 has been reported (Figure 5B; PDB code, 3G8Q) [17]. The domain arrangement of CDAT8 is different from that of ThiI. From the N-terminus to the C-terminus, the order of the domains is deaminase, ferredoxin-like and THUMP. However, the structure of the ferredoxin-like and THUMP domains is very similar to that of ThiI. From the model of the complex between CDAT8 and tRNA, it was predicted that the THUMP domain of CDAT8 captures the CCA terminus of substrate tRNA [17].
4.3. Methyltransferase
Of the different modified nucleosides in tRNA, methylated nucleosides are the most abundant [1,2,90]. Consistent with this, numerous tRNA methyltransferases have been identified [90]. Transfer RNA methyltransferases can be divided into two types according to the methyl group donor. The majority of tRNA methyltransferases use S-adenosyl-L-methionine as a methyl group donor whereas mnmG (previous name, GidA) [91,92,93,94,95,96] and TrmFO [69,77,97,98] are an exception and use N5, N10-methylenetetrafolare. S-adenosyl-L-methionine-dependent tRNA methyltransferases are further classified on the basis of their catalytic domain [9,90,99]. The majority of S-adenosyl-L-methionine-dependent tRNA methyltransferases possess a Rossmann fold catalytic domain [9,99]. The second group of S-adenosyl-L-methionine-dependent tRNA methyltransferases belong to a SpoU-TrmD (SPOUT) superfamily, which possess a SPOUT catalytic domain [9,100]. In addition, TrmO is an exception and has a b-barrel type catalytic domain [101].
All THUMP-related tRNA methyltransferases reported possess a Rossmann fold catalytic domain and synthesize only m2G (and m22G) (Figure 1 and Figure 2 and Table 2). Several enzymes synthesize m22G from m2G by a second methylation and act on multiple positions (Figure 2). Although classification of tRNA (m2G/m22G) methyltransferases is complicated, the THUMP-related tRNA (m2G/m22G) methyltransferases can be divided into two types according to their methylation sites (Table 2). Thus, Trm11/arcTrm11/arcTrm11-arcTrm112/TRMT11-TRMT112 act on position 10 in tRNA, whereas TrmN/Trm14/THUMPD3-TRMT112 act on position 6 and an additional site. It should be mentioned that tRNA (m2G/m22G) methyltransferases, which do not possess a THUMP domain, do exist. One major group of such tRNA (m2G/m22G) methyltransferases is the Trm1 family [102,103,104,105,106,107,108,109,110]. S. cerevisiae Trm1 catalyzes the methylation of G26 in tRNA and synthesizes m2G26 and m22G26 [102,103]. Mammalian and Aquifex aeolicus Trm1 enzymes form m2G27 and m22G27 in addition to m2G26 and m22G26 [105,107]. Crystal structures of P. horikoshii [109] and A. aeolicus [110] Trm1 proteins demonstrate that these proteins possess a distinct C-terminal domain instead of a THUMP domain.
Trm112, TRMT112 and arcTrm112 are hub-proteins (Figure 2 and Table 2), which regulate multiple methyltransferases [23,24,27,111,112,114,115,116]. In the case of human TRMT11-TRMT112, formation of the complex has been reported [111]. However, the modification, position and substrate tRNAs of human TRMT11-TRMT112 have not been experimentally confirmed. For T. kodakarensis Trm14, tRNATrp from a trm14 gene deletion strain loses the m2G67 modification [113]. However, subunit composition and enzymatic activity of T. kodakarensis Trm14 have not been confirmed with a purified enzyme. In addition, recently, RNA fragments from tRNA mixtures purified from M. Jannaschii [117], M. maripaldis, P. furiosus and Sulfolobus acidocaldarius [118] were analyzed by mass-spectrometry. m2G6 and m2G67 were observed in several tRNAs from M. Jannaschii [117], and thus Trm14 is probably involved in the formation of these modifications. Furthermore, in the case of P. furiosus, several tRNAs were shown to possess a m22G6 modification in addition to m2G6 and m2G67 modifications [118]. Therefore, archaeal Trm14 proteins may possess broader positional specificity than was previously thought.
As described in the Introduction, the P. abyssi PAB1283 protein (arcTrm11) was the first tRNA methyltransferase identified as containing a THUMP domain [13]. The THUMP domain of P. abyssi arcTrm11 has been expressed in E. coli cells, purified and analyzed [119]. This study [119] reported that the THUMP domain autonomously folds and that the affinity of the THUMP domain for tRNA is very weak. In 2005, it was reported that S. cerevisiae Trm11 requires a partner subunit, Trm112 [23]. Furthermore, the S. cerevisiae Trm11-Trm112 complex only produces m2G10 in tRNA [23] whereas arcTrm11 produces m2G10 and m22G10 [13,24,34]. Moreover, in several archaea, arcTrm11 requires arcTrm112 for enzymatic activity as seen with S. cerevisiae Trm11 [24,112].
T. thermophilus TrmN is the only eubacterial THUMP-related tRNA methyltransferase reported [25]. TrmN methylates G6 in tRNAPhe and produces m2G6 [25]. Methanococcus jannaschii Trm14 is an archaeal homolog of TrmN and produces m2G6 (and m22G6) in tRNACys [26]. Furthermore, in in vitro experiments, the second methylation from m2G6 to m22G6 in the tRNACys transcript was observed [26]. The human THUMPD3-TRMT112 complex methylates G6 and G7 in several tRNAs and produces m2G6 and m2G7 [27].
In 2012, crystal structures of P. abyssi Trm14 (Figure 6A) and T. thermophilus TrmN (Figure 6B) were reported [120]. Both enzymes methylate G6 in tRNA and produce m2G6. The crystal structures revealed that these enzymes possess a N-terminal ferredoxin-like domain, a THUMP domain, a Rossmann fold methyltransferase (methylase) domain and a linker region. In the same study, it was reported that several positively charged amino acid residues are involved in tRNA binding [120]. Furthermore, the structures of the ferredoxin-like domain and the THUMP domain of Trm14 and TrmN are remarkably similar to those of ThiI and CDAT8. In 2016, the crystal structure of T. kodakarensis arcTrm11 was solved (Figure 6C) [34]. The arrangement of the domains of arcTrm11 is the same as that of Trm14 and TrmN. However, the distance between the THUMP and methylase domains in arcTrm11 is longer than that in Trm14 and TrmN due to structural differences in the ferredoxin-like domain and the linker region. This difference is important for the selection of the modification site (G10 or G6) (Figure 6D). A site-directed mutagenesis study showed that the THUMP domain in arcTrm11 captures the CCA terminus of substrate tRNA [34]. The distance between the CCA terminus and G10 in tRNA is longer than the distance between the CCA terminus and G6 (Figure 6D). Thus, these crystal structures led to the idea that the methylation site (G6 or G10) is determined by the distance from the THUMP domain to the catalytic pocket.
Eukaryotic and some archaeal Trm11 proteins require a partner subunit (Trm112, TRMT112 or arcTrm112) for enzymatic activity [23,24,27,111,112,114,115,116]. It should be mentioned that eukaryotic Trm112 homologs activate multiple methyltransferases. For example, S. cerevisiae Trm112 activates Trm9 [121], Bud23 [122,123] and Mtq2 [124,125] in addition to Trm11. Furthermore, a human ortholog of Trm112, TRMT112 interacts with at least seven human methyltransferases (WBSCR22 (responsible for formation of 7-methylguanosine at position 1636 in 18S rRNA) [126], METTL5 (formation of N6-methyladenosine at position 1832 in 18S rRNA) [127], HEMK2 (methylation of a glutamine side chain of eRF1 protein) [128], ALKBH8 (responsible for 5-methoxycarbonylmethyluridine derivatives at position 34 in tRNA) [129,130,131,132], TRMT11 [111], THUMPD2 (function unknown) [111] and THUMPD3 (production of m2G6 and m2G7 in tRNA)) [27].
Several tRNA modification enzymes form protein complexes [90,91,96,116,133,134,135,136]. The partner subunit(s) is frequently involved in the substrate tRNA recognition. Consequently, the binding sites of these modification enzymes are often extended over the whole tRNA molecule. For example, as described in Section 4.1., bacterial tRNA (m7G46) methyltransferase (TrmB) can methylate a truncated tRNA, in which the interaction between the T-arm and D-arm is disrupted [81]. However, in contrast, eukaryotic tRNA (m7G46) methyltransferase (Trm8-Trm82) [136] requires the interaction between the T-arm and D-arm for methylation [137]. Thus, the existence of Trm82 seems to act on recognition of the L-shaped tRNA structure. In the case of S. cerevisiae Trm7, the partner subunits (Trm732 and Trm734) decide the modification positions: Trm7-Trm732 and Trm7-Trm734 catalyze 2′-O-methylations at position 32 and position 34, respectively, in tRNA [138]. The biochemical and structural studies of Trm7-Trm734 suggest that Trm734 captures the D-arm in substrate tRNA and controls the accession of the modification site (ribose at position 34) in tRNA to the catalytic pocket in Trm7 [139]. A conserved motif (RRSAGLP sequence) in Trm732 is involved in the methylation of position 32 in tRNAPhe [140]. Thus, the presence of a partner subunit is frequently involved in substrate tRNA recognition.
S. cerevisiae Trm11-Trm112 does not methylate truncated tRNAs [141]. This observation suggests that the binding sites of Trm11-Trm112 in tRNA are spread over the whole tRNA molecule. Biochemical and biophysical studies of S. cerevisiae Trm11-Trm112 resulted in the proposal of a model in whichTrm112 is accessible to the anticodon-loop region in tRNA dependent on the movement of the THUMP domain [142]. The required elements in tRNA for methylation by Trm11-Trm112 have been clarified (Figure 7A): the CCA terminus, G10-C25 base pair, regular size (5 nt) variable region and ribose-phosphate backbone around purine38 in tRNA are essential for methylation by S. cerevisiae Trm11-Trm112 [141]. Thus, the biochemical study [141] supports the model referenced [142] because the ribose-phosphate backbone around position 38 is recognized by S. cerevisiae Trm11-Trm112. Furthermore, the crystal structure of A. fulgidus arcTrm11-arcTrm112 has been reported (Figure 7B) [24]. When the THUMP domain in arcTrm11 captures the CCA terminus in substrate tRNA, arcTrm112 accesses the anticodon-loop. Therefore, tRNA recognition mechanisms of eukaryotic and archaeal Trm11-Trm112 seem to be basically common. Human THUMPD3-TRMT112 requires the CCA terminus for methylation and does not methylate a mini-helix RNA [27]. Therefore, TRMT112 in THUMPD3-TRMT112 may also be involved in the anticodon-loop recognition as per Trm11-Trm112.
4.4. Acetyltransferase
As described in the Introduction, S. cerevisiae Tan1 (human THUMPD1) contains a THUMP domain and acts as a partner protein of tRNA acetyltransferse, Kre33 (human NAT10) [16]. The Methanothermobacter thermautotrophicus Tan1 homolog is composed of N-terminal ferredoxin-like and C-terminal THUMP domains [15]. Although the crystal structure of Kre33 (or NAT10) has not been reported, a structural model (PDB code, 2ZPA) has been proposed [16] in which Kre33 (NAT10) contains DUF1726 (of unknown function), helicase, N-acetyltransferase and tRNA binding domains. In the case of T. kodakarensis TkNAT10 (the archaeal homolog of NAT10), the C-terminal region is missing [18]. Kre33 catalyzes the acetylation of 18S rRNA as well as acetylation of tRNA [16]. A random mutagenesis study of T. kodakarensis revealed that the disruption of the Tk0754 gene causes complete loss of ac4C modification in a tRNA mixture [143]. Detailed enzymatic activity of the Tk0754 gene product (TkNAT10) has been reported [18]. In this study, TkNAT10 was shown to modify multiple positions in various RNAs including tRNAs, and the rate of acetylation is increased according to increase in temperature [18]. Yeast two-hybrid system experiments have shown that Tan1 and Kre33 form a complex [16]; however, the structure of the Tan1 and Kre33 complex has not been reported. For details of acetylation of rRNA and other RNAs, see these references [16,18,19].
4.5. Pseudouridine Synthase
Pseudouridine (Ψ) is abundant in RNAs from the three domains of life [1,2,3] and is synthesized by C5-ribosyl isomerization from uridine, which is catalyzed by pseudouridine synthases [144,145,146,147,148,149,150]. Pseudouridine synthases can be classified into six families; however, PUS10 is the only THUMP-related enzyme [28,29,144,145,146,147,148,149,150]. In 2006, Ψ55 formation in tRNA catalyzed by archaeal Pus10 was reported [28]. Thus, this report demonstrates that one of the predicted THUMP-containing proteins [11] has pseudouridine synthase activity. In 2008, it was reported that archaeal Pus10 can synthesize Ψ54 in tRNA in addition to Ψ55 [29]. Furthermore, Methanocaldoccus jannaschii PUS10 can modify U54 and U55 in a micro-helix RNA, which mimics the T-arm [151].
In 2007, a crystal structure of human PUS10 was reported (Figure 8) and showed that the THUMP-related structure is contained in the N-terminal accessory domain [20]. When the CCA-terminus in tRNA is placed onto the THUMP-related structure, the modification sites (U54 and U55) have access to the catalytic pocket of the pseudouridine synthase domain [20]. However, human PUS10 can modify U54 in a tRNA transcript without a CCA terminus [30]. Because human PUS10 strongly recognizes the sequences of the aminoacyl-stem and T-arm [30], the recognition of the CCA terminus by the THUMP-related structure may be not important for pseudouridine formation. The accessory domain of human PUS10 is large compared to a typical THUMP domain. This large accessory domain was gained in the process of evolution of eukaryotic PUS10 [143]. Furthermore, tRNA recognition by human PUS10 in living cells is complicated. Human PUS10 is expressed in both the nucleus and cytoplasm [30]. Human nuclear PUS10 does not have the pseudouridine synthesis activity and inhibits the activity of TRUB1 [human tRNA (Ψ55) synthase] by binding to specific tRNAs in the nucleus [31]. In contrast, human cytoplasmic PUS10 can synthesize Ψ54 in tRNAs, which possess an AAAU sequence from position 57 to position 60 in the T-loop, in addition to Ψ55 [31]. Moreover, it has been reported that human PUS10 is involved in microRNA processing [152]. In this process, PUS10 directly binds to primary microRNA and the catalytic activity of PUS10 is not required [152]. Thus, PUS10 may act as an RNA binding subunit in microRNA processing.
Based on the crystal structure of human PUS10, a structural model of archaeal PUS10 was constructed and several amino acid residues, which are required for enzymatic activity and tRNA binding, were identified [21]. Another mutagenesis study revealed that the thumb-loop in the catalytic domain and N-terminal cysteine residues are important for the Ψ54 formation activity of M. jannaschii PUS10 [151].
5. Functions of Modified Nucleosides, Which Are Produced by THUMP-Related tRNA Modification Enzymes and Additional Information
In this section, the functions of modified nucleosides, which are produced by THUMP-related tRNA modification enzymes, are introduced. Furthermore, the relationships between the disorder of modification (or modification enzyme) and higher biological phenomena are explained.
5.1. s4U8 and s4U9
The s4U modification is observed at positions 8 and 9 in eubacterial and archaeal tRNAs [1,2,3]. The physiological roles of s4U have gradually been elucidated. The s4U modification in tRNA acts as an ultraviolet light (UV)-resistant factor [153]. Irradiation with near-UV causes crosslinking between s4U8 and C13 in tRNA [154]. Because ThiI requires the CCA terminus for the s4U modification, crosslinking by s4U occurs after the removal of the 3′-trailer sequence from precursor tRNA. This crosslinking of tRNA pauses protein synthesis and activates the DNA repair system [155,156]. Furthermore, crosslinking slows down the speed of TrmH-mediated Gm18 formation in tRNA [157]. Several archaea and bacteria live in environments in which sunlight does not reach (for example, deep sea and underground). However, these organisms also possess the s4U modification in tRNA [158], suggesting that the s4U modification functions beyond being a UV-resistant factor. As described in Section 3.1, the s4U8 modification contributes to the maintenance of the L-shaped tRNA structure. Furthermore, the s4U8 modification works as a tRNA quality control system in Vibrio cholerae in the stationary growth phase [159].
5.2. U8
Deamination from C8 to U8 performed by CDAT8 is one of the thermophile-specific tRNA modifications [17,62]. M. kandleri grows at high temperatures (more than 110 °C). Therefore, C8 in the tRNA genes may contribute to maintain the double-stranded DNA structure of the M. kandleri genome at high temperatures through an increase in the G-C content [17].
5.3. m2G6, m22G6, m2G7, m2G10, m22G10 and m2G67
The m2G modification does not disrupt formation of a Watson–Crick base pair with C, and the methyl group in m2G probably stabilizes the stem structure by hydrophobic interaction. The growth rate of a S. cerevisiae trm11 gene deletion strain is comparable to that of the wild-type strain under laboratory conditions [23]. However, a trm1-trm11 double-gene deletion strain shows an obvious growth defect [23]. Because Trm1 is the tRNA methyltransferase responsible for the formation of m22G26 [102,103], the study [23] strongly suggests that the m2G10 modification works in co-ordination with other modification(s) in tRNA. In the case of T. kodakarensis, the trm11 gene deletion strain cannot grow at high temperatures (95 °C) [113,160]. In T. thermophilus, the tRNA modification enzymes and modified nucleosides form a network in which modified nucleosides regulate the activities of other tRNA modification enzymes negatively and positively [62,63,161,162,163,164]. However, trmN gene deletion from the T. thermophilus genome does not have an effect on other modifications in tRNA [25]. This observation suggests that the m2G6 modification is a relatively late modification like dihydrouridine modification at positions 20 and 20a by DusA [165,166,167,168,169] in T. thermophilus tRNAs. In thermophiles, long and branched polyamines are produced [170,171] and have an effect on tRNA modifications [172,173]. In tRNA from the T. thermophilus speB or speD1 gene deletion strain in which long and branched polyamines are not synthesized, the m2G6 modification in tRNA is increased [174]. Therefore, long and branched polyamines may negatively regulate m2G6 formation by TrmN in T. thermophilus cells. THUMD3 knockout HEK293T cell lines show decreased protein synthesis activity and an obviously slow growth rate [27]. Thus, human THUMPD3-TRMT112 is required for cell proliferation [27]. Furthermore, absence and presence of the m2G7 modification in tRNATrp are involved in the infection of avian retrovirus [175]. Moreover, although squid tRNALys contains m2G67 [176], this modification is not explainable by the enzymatic activity of currently known eukaryotic tRNA methyltransferases.
5.4. ac4C12 and ac4C at Multiple Positions
Recent technologies, which can detect ac4C in RNAs, have shown that the ac4C modification is present in various RNAs beyond tRNA and rRNA [18,19]. As described in the Introduction, a THUMP-related protein, S. cerevisiae Tan1, was found to be an essential protein for ac4C12 modification in tRNA [14] but does not act in acetylation of 18S rRNA [14,16]. The S. cerevisiae tan1 gene deletion strain shows a decrease in tRNASer [14]. Furthermore, the S. cerevisiae tan1 and trm44 double mutant strain cannot grow at 33 °C [177]. Trm44 is a tRNA methyltransferase responsible for formation of Um44 in tRNASer [177]. Thus, these studies show that ac4C12 contributes to stabilizing tRNASer and works with other modifications such as Um44. Hypomodified tRNASer is degraded by a rapid tRNA decay pathway, which competes with the elongation factor 1A [178]. S. cerevisiae Tan1 precursor-mRNA processing requires the conserved precursor-mRNA retention and splicing complex (RES complex; Bud13, Snu17 and Pml1 complex) [179]. Thereby, the RES complex controls ac4C12 modification in tRNA [179]. In the case of T. kodkarensis, ac4C modification by TkNAT10 occurs in various RNAs including tRNAs and is increased at high temperatures [18]. The acetylation by TkNAT10 is essential for survival of T. kodakarensis at high temperatures [18,160]. Loss of function of human THUMD1 causes a syndromic neurodevelopmental disorder [180]. The expression level of THUMD1 is increased in breast cancer cells [181]. Furthermore, THUMD1 overexpression enhanced breast cancer cells’ invasion and migration [181]. Moreover, although human NAT10 localizes mainly in nucleoli of normal tissues, it is redistributed to the membrane of colon cancer cells [182]. In addition, the expression level of NAT10 is increased in liver cancer [183].
5.5. ψ 54 and ψ 55
The modifications at positions 54 and 55 in tRNA stabilize the interaction between the T-arm and D-arm. Almost all tRNAs possess U modifications at position 54 (for example, m5U54, Ψ54, m5s2U54, m1 Ψ54, Um54, m5Um54, and s2Um54) and Ψ55 [3]. The Ψ54 modification is observed in tRNAs from archaea and some eukaryotes, and the Ψ55 modification is found in tRNAs from the three domains of life. Only higher eukaryotes and archaea possess PUS10 [28,29,184]. Consequently, eubacteria and yeast possess other enzymes. In the case of E. coli, TrmA [72] and TruB [73] catalyze the formation of m5U54 and Ψ55, respectively. In the case of yeast, m5U54 and Ψ55 are produced by Trm2 [185] and PUS4 [186], respectively. In archaea and higher eukaryotes, the Ψ55 modification in tRNA is synthesized by redundant systems. In archaea, archaeal Cbf5 (or archaeal Cbf5-Gar1 complex) and arcPUS10 can synthesize the Ψ55 modification [28,184]. In humans, nuclear TRUB1, mitochondrial TRUB2 and cytoplasmic PUS10 catalyze the formation of Ψ55 [31]. Consequently, cytoplasmic tRNAs are modified by TRUB1 or PUS10. Furthermore, it has been reported that PUS1 and PUS4 can synthesize the Ψ55 modification in Cyanidioschyzon merolae [187]. Although C. merolae does not possess PUS10, the redundant Ψ55 formation in tRNA is also observed in red algae. These facts suggest the importance of the Ψ55 modification. In Haloferax volcanii and M. jannaschii, the Ψ54 modification is further modified to m1 Ψ54 by TrmY [188,189]. Furthermore, in Ignicoccus hospitalis, the m1 Ψ54 modification is modified to m1s4 Ψ54 by TtuA and TtuB [190]. TtuA and TtuB are a sulfur-transfer complex responsible for the formation of s2U54 in tRNA [40,191]. The PUS10 gene may be essential for survival of H. volcanii (the PUS10 gene deletion mutant strain could not be obtained) [192]. In humans, mutations in PUS10 gene are involved in Crohn’s disease and celiac disease (chronic intestinal inflammatory diseases) [193]. Human cytoplasmic PUS10 can synthesize Ψ54 in tRNAs, which possess an AAAU sequence from position 57 to position 60 in the T-loop, in addition to Ψ55 [30].
6. Perspective
In this review, I focus on the structures and functions of THUMP-related tRNA modification enzymes and the modified nucleosides they produce in tRNA. As described above, the studies of tRNA 4-thiouridine synthase, tRNA deaminase and tRNA methyltransferases have established the concept that the THUMP domain captures the 3′-end of RNA (the CCA-terminus of tRNA). The Tan1-Kre33 complex may have a similar recognition mechanism for substrate tRNA. However, TkTAN1-TkNAT10 modify multiple positions in tRNA. This phenomenon cannot be simply explained by our current knowledge. Furthermore, human PUS10 does not show the pseudouridine synthase activity in nucleus and is involved in processing of microRNA. Thus, functions and regulations of THUMP-related proteins in higher eukaryotes are complicated. Several THUMP-related proteins may be involved in the maturation of other RNAs beyond tRNA modifications. Moreover, there are many THUMP-related proteins for which the function is unknown. For example, the function of human THUMD2, which is predicted as a THUMP-related protein, is still unknown. Thus, further study will be necessary to clarify these issues.
Funding
This work was supported by a Grant-in-Aid for Scientific Research (20H03211 to HH) from the Japan Society for the Promotion of Science (JSPS).
Conflicts of Interest
The author declares no conflict of interest.
References
- Boccaletto, P.; Stefaniak, F.; Ray, A.; Cappannini, A.; Mukherjee, S.; Purta, E.; Kurkowska, M.; Shirvanizadeh, N.; Destefanis, E.; Groza, P.; et al. MODOMICS: A database of RNA modification pathways. 2021 update. Nucleic Acids Res. 2022, 50, D231–D235. [Google Scholar] [CrossRef]
- Jühling, F.; Mörl, M.; Hartmann, R.K.; Sprinzl, M.; Stadler, P.F.; Pütz, J. tRNAdb 2009: Compilation of tRNA sequences and tRNA genes. Nucleic Acids Res. 2009, 37, D159–D162. [Google Scholar] [CrossRef] [Green Version]
- Sajek, M.P.; Woźniak, T.; Sprinzl, M.; Jaruzelska, J.; Barciszewski, J. T-psi-C: User friendly database of tRNA sequences and structures. Nucleic Acids Res. 2020, 48, D256–D260. [Google Scholar] [CrossRef]
- Lim, K.; Zhang, H.; Tempczyk, A.; Krajewski, W.; Bonander, N.; Toedt, J.; Howard, A.; Eisenstein, E.; Herzberg, O. Structure of the YibK methyltransferase from Haemophilus influenzae (HI0766): A cofactor bound at a site formed by a knot. Proteins 2003, 51, 56–67. [Google Scholar] [CrossRef]
- Benítez-Páez, A.; Villarroya, M.; Douthwaite, S.; Gabaldón, T.; Armengod, M.E. YibK is the 2’-O-methyltransferase TrmL that modifies the wobble nucleotide in Escherichia coli tRNA(Leu) isoacceptors. RNA 2010, 16, 2131–2143. [Google Scholar] [CrossRef] [Green Version]
- Hori, H.; Suzuki, T.; Sugawara, K.; Inoue, Y.; Shibata, T.; Kuramitsu, S.; Yokoyama, S.; Oshima, T.; Watanabe, K. Identification and characterization of tRNA (Gm18) methyltransferase from Thermus thermophilus HB8: Domain structure and conserved amino acid sequence motifs. Genes Cells 2002, 7, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Nureki, O.; Watanabe, K.; Fukai, S.; Ishii, R.; Endo, Y.; Hori, H.; Yokoyama, S. Deep knot structure for construction of active site and cofactor binding site of tRNA modification enzyme. Structure 2004, 12, 593–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkaczuk, K.L.; Dunin-Horkawicz, S.; Purta, E.; Bujnicki, J.M. Structural and evolutionary bioinformatics of the SPOUT superfamily of methyltransferases. BMC Bioinform. 2007, 8, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, H. Transfer RNA methyltransferases with a SpoU-TrmD (SPOUT) fold and their modified nucleosides in tRNA. Biomolecules 2017, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Strassler, S.E.; Bowles, I.E.; Dey, D.; Jackman, J.E.; Conn, G.L. Tied up in knots: Untangling substrate recognition by the SPOUT methyltransferases. J. Biol. Chem. 2022, 298, 102393. [Google Scholar] [CrossRef]
- Aravind, L.; Koonin, E.V. THUMP—A predicted RNA-binding domain shared by 4-thiouridine, pseudouridine synthases and RNA methylases. Trends Biochem. Sci. 2001, 26, 215–217. [Google Scholar] [CrossRef]
- Mueller, E.G.; Buck, C.J.; Palenchar, P.M.; Barnhart, L.E.; Paulson, J.L. Identification of a gene involved in the generation of 4-thiouridine in tRNA. Nucleic Acids Res. 1998, 26, 2606–2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armengaud, J.; Urbonavicius, J.; Fernandez, B.; Chaussinand, G.; Bujnicki, J.M.; Grosjean, H. N2-methylation of guanosine at position 10 in tRNA is catalyzed by a THUMP domain-containing, S-adenosylmethionine-dependent methyltransferase, conserved in Archaea and Eukaryota. J. Biol. Chem. 2004, 279, 37142–37152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.J.; Byström, A.S. The Saccharomyces cerevisiae TAN1 gene is required for N4-acetylcytidine formation in tRNA. RNA 2004, 10, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.P.; Byrne, R.T.; Chechik, M.; Smits, C.; Waterman, D.G.; Antson, A.A. Expression, purification, crystallization and preliminary X-ray studies of the TAN1 orthologue from Methanothermobacter thermautotrophicus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2008, 64, 1083–1086. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Langhendries, J.L.; Watzinger, P.; Kötter, P.; Entian, K.D.; Lafontaine, D.L. Yeast Kre33 and human NAT10 are conserved 18S rRNA cytosine acetyltransferases that modify tRNAs assisted by the adaptor Tan1/THUMPD1. Nucleic Acids Res. 2015, 43, 2242–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randau, L.; Stanley, B.J.; Kohlway, A.; Mechta, S.; Xiong, Y.; Söll, D. A cytidine deaminase edits C to U in transfer RNAs in Archaea. Science 2009, 324, 657–659. [Google Scholar] [CrossRef]
- Sas-Chen, A.; Thomas, J.M.; Matzov, D.; Taoka, M.; Nance, K.D.; Nir, R.; Bryson, K.M.; Shachar, R.; Liman, G.L.S.; Burkhart, B.W.; et al. Dynamic RNA acetylation revealed by quantitative cross-evolutionary mapping. Nature 2020, 583, 638–643. [Google Scholar] [CrossRef]
- Jin, G.; Xu, M.; Zou, M.; Duan, S. The Processing, Gene Regulation, Biological Functions, and Clinical Relevance of N4-Acetylcytidine on RNA: A Systematic Review. Mol. Ther. Nucleic Acids 2020, 20, 13–24. [Google Scholar] [CrossRef]
- McCleverty, C.J.; Hornsby, M.; Spraggon, G.; Kreusch, A. Crystal structure of human Pus10, a novel pseudouridine synthase. J. Mol. Biol. 2007, 373, 1243–1254. [Google Scholar] [CrossRef]
- Kamalampeta, R.; Keffer-Wilkes, L.C.; Kothe, U. tRNA binding, positioning, and modification by the pseudouridine synthase Pus10. J. Mol. Biol. 2013, 425, 3863–3874. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, X.; Nakamura, A.; Orlando, R.; Söll, D.; Whitman, W.B. Biosynthesis of 4-thiouridine in tRNA in the methanogenic archaeon Methanococcus maripaludis. J. Biol. Chem. 2012, 20, 36683–36692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purushothaman, S.K.; Bujnicki, J.M.; Grosjean, H.; Lapeyre, B. Trm11p and Trm112p are both required for the formation of 2-methylguanosine at position 10 in yeast tRNA. Mol. Cell Biol. 2005, 25, 4359–4370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Van Tran, N.; Jactel, V.; Guérineau, V.; Graille, M. Structural and functional insights into Archaeoglobus fulgidus m2G10 tRNA methyltransferase Trm11 and its Trm112 activator. Nucleic Acids Res. 2020, 48, 11068–11082. [Google Scholar] [CrossRef] [PubMed]
- Roovers, M.; Oudjama, Y.; Fislage, M.; Bujnicki, J.M.; Versées, W.; Droogmans, L. The open reading frame TTC1157 of Thermus thermophilus HB27 encodes the methyltransferase forming N2-methylguanosine at position 6 in tRNA. RNA 2012, 18, 815–824. [Google Scholar] [CrossRef] [Green Version]
- Menezes, S.; Gaston, K.W.; Krivos, K.L.; Apolinario, E.E.; Reich, N.O.; Sowers, K.R.; Limbach, P.A.; Perona, J.J. Formation of m2G6 in Methanocaldococcus jannaschii tRNA catalyzed by the novel methyltransferase Trm14. Nucleic Acids Res. 2011, 39, 7641–7655. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.Q.; Xiong, Q.P.; Ge, J.Y.; Li, H.; Zhu, W.Y.; Nie, Y.; Lin, X.; Lv, D.; Li, J.; Lin, H.; et al. THUMPD3-TRMT112 is a m2G methyltransferase working on a broad range of tRNA substrates. Nucleic Acids Res. 2021, 49, 11900–11919. [Google Scholar] [CrossRef]
- Roovers, M.; Hale, C.; Tricot, C.; Terns, M.P.; Terns, R.M.; Grosjean, H.; Droogmans, L. Formation of the conserved pseudouridine at position 55 in archaeal tRNA. Nucleic Acids Res. 2006, 34, 4293–4301. [Google Scholar] [CrossRef]
- Gurha, P.; Gupta, R. Archaeal Pus10 proteins can produce both pseudouridine 54 and 55 in tRNA. RNA 2008, 14, 2521–2527. [Google Scholar] [CrossRef] [Green Version]
- Deogharia, M.; Mukhopadhyay, S.; Joardar, A.; Gupta, R. The human ortholog of archaeal Pus10 produces pseudouridine 54 in select tRNAs where its recognition sequence contains a modified residue. RNA 2019, 25, 336–351. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Deogharia, M.; Gupta, R. Mammalian nuclear TRUB1, mitochondrial TRUB2, and cytoplasmic PUS10 produce conserved pseudouridine 55 in different sets of tRNA. RNA 2021, 27, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Nomura, Y.; Ohno, S.; Nishikawa, K.; Yokogawa, T. Correlation between the stability of tRNA tertiary structure and the catalytic efficiency of a tRNA-modifying enzyme, archaeal tRNA-guanine transglycosylase. Genes Cells 2016, 21, 41–52. [Google Scholar] [CrossRef]
- Westhof, E.; Dumas, P.; Moras, D. Crystallographic refinement of yeast aspartic acid transfer RNA. J. Mol. Biol. 1985, 184, 119–145. [Google Scholar] [CrossRef]
- Hirata, A.; Nishiyama, S.; Tamura, T.; Yamauchi, A.; Hori, H. Structural and functional analyses of the archaeal tRNA m2G/m22G10 methyltransferase aTrm11 provide mechanistic insights into site specificity of a tRNA methyltransferase that contains common RNA-binding modules. Nucleic Acids Res. 2016, 44, 6377–6390. [Google Scholar] [CrossRef] [Green Version]
- Urbonavicius, J.; Armengaud, J.; Grosjean, H. Identity elements required for enzymatic formation of N2,N2-dimethylguanosine from N2-monomethylated derivative and its possible role in avoiding alternative conformations in archaeal tRNA. J. Mol. Biol. 2006, 357, 387–399. [Google Scholar] [CrossRef]
- Kawai, G.; Hashizume, T.; Miyazawa, T.; McCloskey, J.A.; Yokoyama, S. Conformational characteristics of 4-acetylcytidine found in tRNA. Nucleic Acids Symp. Ser. 1989, 21, 61–62. [Google Scholar]
- Bartee, D.; Nance, K.D.; Meier, J.L. Site-Specific Synthesis of N4-Acetylcytidine in RNA Reveals Physiological Duplex Stabilization. J. Am. Chem. Soc. 2022, 144, 3487–3496. [Google Scholar] [CrossRef] [PubMed]
- Nobles, K.N.; Yarian, C.S.; Liu, G.; Guenther, R.H.; Agris, P.F. Highly conserved modified nucleosides influence Mg2+-dependent tRNA folding. Nucleic Acids Res. 2002, 30, 4751–4760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čavužić, M.; Liu, Y. Biosynthesis of Sulfur-Containing tRNA Modifications: A Comparison of Bacterial, Archaeal, and Eukaryotic Pathways. Biomolecules 2017, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Shigi, N. Biosynthesis and Degradation of Sulfur Modifications in tRNAs. Int. J. Mol. Sci. 2021, 22, 11937. [Google Scholar] [CrossRef]
- Zheng, C.; Black, K.A.; Dos Santos, P.C. Diverse Mechanisms of Sulfur Decoration in Bacterial tRNA and Their Cellular Functions. Biomolecules 2017, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.Y.; Wu, Y.; Begley, T.J.; Sheng, J. Sulfur modification in natural RNA and therapeutic oligonucleotides. RSC Chem. Biol. 2021, 2, 990–1003. [Google Scholar] [CrossRef] [PubMed]
- Kambampati, R.; Lauhon, C.T. IscS is a sulfurtransferase for the in vitro biosynthesis of 4-thiouridine in Escherichia coli tRNA. Biochemistry 1999, 38, 16561–16568. [Google Scholar] [CrossRef]
- Lauhon, C.T.; Kambampati, R. The iscS gene in Escherichia coli is required for the biosynthesis of 4-thiouridine, thiamin, and NAD. J. Biol. Chem. 2000, 275, 20096–20103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauhon, C.T. Requirement for IscS in biosynthesis of all thionucleosides in Escherichia coli. J. Bacteriol. 2002, 184, 6820–6829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palenchar, P.M.; Buck, C.J.; Cheng, H.; Larson, T.J.; Mueller, E.G. Evidence that ThiI, an enzyme shared between thiamin and 4-thiouridine biosynthesis, may be a sulfurtransferase that proceeds through a persulfide intermediate. J. Biol. Chem. 2000, 275, 8283–8286. [Google Scholar] [CrossRef] [Green Version]
- Mueller, E.G.; Palenchar, P.M.; Buck, C.J. The role of the cysteine residues of ThiI in the generation of 4-thiouridine in tRNA. J. Biol. Chem. 2001, 276, 33588–33595. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Vinyard, D.J.; Reesbeck, M.E.; Suzuki, T.; Manakongtreecheep, K.; Holland, P.L.; Brudvig, G.W.; Söll, D. A [3Fe-4S] cluster is required for tRNA thiolation in archaea and eukaryotes. Proc. Natl. Acad. Sci. USA 2016, 113, 12703–12708. [Google Scholar] [CrossRef] [Green Version]
- Tomikawa, C.; Ohira, T.; Inoue, Y.; Kawamura, T.; Yamagishi, A.; Suzuki, T.; Hori, H. Distinct tRNA modifications in the thermo-acidophilic archaeon, Thermoplasma acidophilum. FEBS Lett. 2013, 587, 3537–3580. [Google Scholar] [CrossRef] [Green Version]
- He, N.; Zhou, J.; Bimai, O.; Oltmanns, J.; Ravanat, J.L.; Velours, C.; Schünemann, V.; Fontecave, M.; Golinelli-Pimpaneau, B. A subclass of archaeal U8-tRNA sulfurases requires a [4Fe-4S] cluster for catalysis. Nucleic Acids Res. 2022, 50, 12969–12978. [Google Scholar] [CrossRef]
- Waterman, D.G.; Ortiz-Lombardía, M.; Fogg, M.J.; Koonin, E.V.; Antson, A.A. Crystal structure of Bacillus anthracis ThiI, a tRNA-modifying enzyme containing the predicted RNA-binding THUMP domain. J. Mol. Biol. 2006, 356, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Lauhon, C.T.; Erwin, W.M.; Ton, G.N. Substrate specificity for 4-thiouridine modification in Escherichia coli. Substrate specificity for 4-thiouridine modification in Escherichia coli. J. Biol. Chem. 2004, 279, 23022–23029. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Yamagata, S.; Kitago, Y.; Yamada, Y.; Chimnaronk, S.; Yao, M.; Tanaka, I. Deduced RNA binding mechanism of ThiI based on structural and binding analyses of a minimal RNA ligand. RNA 2009, 15, 498–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, P.; Lakomek, K.; Naumann, P.T.; Erwin, W.M.; Lauhon, C.T.; Ficner, R. Crystal structure of a 4-thiouridine synthetase-RNA complex reveals specificity of tRNA U8 modification. Nucleic Acids Res. 2014, 42, 6673–6685. [Google Scholar] [CrossRef] [Green Version]
- Bork, P.; Koonin, E.V. A P-loop-like motif in a widespread ATP pyrophosphatase domain: Implications for the evolution of sequence motifs and enzyme activity. Proteins 1994, 20, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Mueller, E.G.; Palenchar, P.M. Using genomic information to investigate the function of ThiI, an enzyme shared between thiamin and 4-thiouridine biosynthesis. Protein Sci. 1999, 8, 2424–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambampati, R.; Lauhon, C.T. Evidence for the transfer of sulfane sulfur from IscS to ThiI during the in vitro biosynthesis of 4-thiouridine in Escherichia coli tRNA. J. Biol. Chem. 2000, 275, 10727–10730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugahara, M.; Murai, S.; Sugahara, M.; Kunishima, N. Purification, crystallization and preliminary crystallographic analysis of the putative thiamine-biosynthesis protein PH1313 from Pyrococcus horikoshii OT3. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2007, 63, 56–58. [Google Scholar] [CrossRef] [Green Version]
- Webb, E.; Claas, K.; Down, D.M. Characterization of thiI, a new gene involved in thiazole biosynthesis in Salmonella typhimurium. J. Bacteriol. 1997, 179, 4399–4402. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Gomez, N.C.; Palmer, L.D.; Vivas, E.; Roach, P.L.; Downs, D.M. The rhodanese domain of ThiI is both necessary and suf ficient for synthesis of the thiazole moiety of thiamine in Salmonella enterica. J. Bacteriol. 2011, 193, 4582–4587. [Google Scholar] [CrossRef] [Green Version]
- Rajakovich, L.J.; Tomlinson, J.; Dos Santos, P.C. Functional Analysis of Bacillus subtilis Genes Involved in the Biosynthesis of 4-Thiouridine in tRNA. J. Bacteriol. 2012, 194, 4933–4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, H.; Kawamura, T.; Awai, T.; Ochi, A.; Yamagami, R.; Tomikawa, C.; Hirata, A. Transfer RNA Modification Enzymes from Thermophiles and Their Modified Nucleosides in tRNA. Microorganisms 2018, 6, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, H. Regulatory Factors for tRNA Modifications in Extreme- Thermophilic Bacterium Thermus thermophilus. Front. Genet. 2019, 10, 204. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.R.; Santi, D.V. The T-arm of tRNA is a substrate for tRNA (m5U54)-methyltransferase. Biochemistry 1991, 30, 2999–3002. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Ivanetich, K.M.; Santi, D.V. Recognition of the T-arm of tRNA by tRNA (m5U54)-methyltransferase is not sequence specific. Biochemistry 1996, 35, 11652–11659. [Google Scholar] [CrossRef]
- Gu, X.; Yu, M.; Ivanetich, K.M.; Santi, D.V. Molecular recognition of tRNA by tRNA pseudouridine 55 synthase. Biochemistry 1998, 37, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S.; Ueda, T.; Hori, H.; Yamazaki, N.; Okada, N.; Watanabe, K. A UGU sequence in the anticodon loop is a minimum requirement for recognition by Escherichia coli tRNA-guanine transglycosylase. J. Biol. Chem. 1994, 269, 32221–32225. [Google Scholar] [CrossRef]
- Curnow, A.W.; Garcia, G.A. tRNA-guanine transglycosylase from Escherichia coli. Minimal tRNA structure and sequence requirements for recognition. J. Biol. Chem. 1995, 270, 17264–17267. [Google Scholar] [CrossRef] [Green Version]
- Yamagami, R.; Yamashita, K.; Nishimasu, H.; Tomikawa, C.; Ochi, A.; Iwashita, C.; Hirata, A.; Ishitani, R.; Nureki, O.; Hori, H. The tRNA recognition mechanism of folate/FAD-dependent tRNA methyltransferase (TrmFO). J. Biol. Chem. 2012, 287, 42480–42494. [Google Scholar] [CrossRef] [Green Version]
- Takuma, H.; Ushio, N.; Minoji, M.; Kazayama, A.; Shigi, N.; Hirata, A.; Tomikawa, C.; Ochi, A.; Hori, H. Substrate tRNA recognition mechanism of eubacterial tRNA (m1A58) methyltransferase (TrmI). J. Biol. Chem. 2015, 290, 5912–5925. [Google Scholar] [CrossRef] [Green Version]
- Takeda, H.; Toyooka, T.; Ikeuchi, Y.; Yokobori, S.; Okadome, K.; Takano, F.; Oshima, T.; Suzuki, T.; Endo, Y.; Hori, H. The substrate specificity of tRNA (m1G37) methyltransferase (TrmD) from Aquifex aeolicus. Genes Cells 2006, 11, 1353–1365. [Google Scholar] [CrossRef]
- Ny, T.; Björk, G.R. Cloning and restriction mapping of the trmA gene coding for transfer ribonucleic acid (5-methyluridine)-methyltransferase in Escherichia coli K-12. J. Bacteriol. 1980, 142, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Nurse, K.; Wrzesinski, J.; Bakin, A.; Lane, B.G.; Ofengand, J. Purification, cloning, and properties of the tRNA psi 55 synthase from Escherichia coli. RNA 1995, 1, 102–112. [Google Scholar]
- Okada, N.; Nishimura, S. Isolation and characterization of a guanine insertion enzyme, a specific tRNA transglycosylase, from Escherichia coli. J. Biol. Chem. 1979, 254, 3061–3066. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Nishimura, Y.; Hirota, Y.; Nishimura, S. Isolation and characterization of an Escherichia coli mutant lacking tRNA-guanine transglycosylase. Function and biosynthesis of queuosine in tRNA. J. Biol. Chem. 1982, 257, 6544–6550. [Google Scholar] [CrossRef] [PubMed]
- Reuter, K.; Slany, R.; Ullrich, F.; Kersten, H. Structure and organization of Escherichia coli genes involved in biosynthesis of the deazaguanine derivative queuine, a nutrient factor for eukaryotes. J. Bacteriol. 1991, 173, 2256–2264. [Google Scholar] [CrossRef] [Green Version]
- Urbonavicius, J.; Skouloubris, S.; Myllykallio, H.; Grosjean, H. Identification of a novel gene encoding a flavin-dependent tRNA:m5U methyltransferase in bacteria-evolutionary implications. Nucleic Acids Res. 2005, 33, 3955–3964. [Google Scholar] [CrossRef] [PubMed]
- Droogmans, L.; Roovers, M.; Bujnicki, J.M.; Tricot, C.; Hartsch, T.; Stalon, V.; Grosjean, H. Cloning and characterization of tRNA (m1A58) methyltransferase (TrmI) from Thermus thermophilus HB27, a protein required for cell growth at extreme temperatures. Nucleic Acids Res. 2003, 31, 2148–2156. [Google Scholar] [CrossRef] [Green Version]
- Byström, A.S.; Björk, G.R. Chromosomal location and cloning of the gene (trmD) responsible for the synthesis of tRNA (m1G) methyltransferase in Escherichia coli K-12. Mol. Gen. Genet. 1982, 188, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.J.; Long, T.; Zhou, M.; Zhou, X.L.; Wang, E.D. tRNA recognition by a bacterial tRNA Xm32 modification enzyme from the SPOUT methyltransferase superfamily. Nucleic Acids Res. 2015, 43, 7489–7503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, H.; Watanabe, K.; Ikeuchi, Y.; Suzuki, T.; Endo, Y.; Hori, H. Substrate tRNA recognition mechanism of tRNA (m7G46) methyltransferase from Aquifex aeolicus. J. Biol. Chem. 2004, 279, 49151–49159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, T.; Nishikawa, K.; Hori, H.; Ohta, T.; Miura, K.; Watanabe, K. Recognition sites of tRNA by a thermostable tRNA(guanosine-2’-)-methyltransferase from Thermus thermophilus HB27. J. Biochem. 1990, 107, 331–338. [Google Scholar] [CrossRef]
- Purta, E.; Van Vliet, F.; Tkaczuk, K.L.; Dunin-Horkawicz, S.; Mori, H.; Droogmans, L.; Bujnicki, J.M. The yfhQ gene of Escherichia coli encodes a tRNA:Cm32/Um32 methyltransferase. BMC Mol. Biol. 2006, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bie, L.G.; Roovers, M.; Oudjama, Y.; Wattiez, R.; Tricot, C.; Stalon, V.; Droogmans, L.; Bujnicki, J.M. The yggH gene of Escherichia coli encodes a tRNA (m7G46) methyltransferase. J. Bacteriol. 2003, 185, 3238–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persson, B.C.; Jäger, G.; Gustafsson, C. The spoU gene of Escherichia coli, the fourth gene of the spoT operon, is essential for tRNA (Gm18) 2’-O-methyltransferase activity. Nucleic Acids Res. 1997, 25, 4093–4097. [Google Scholar] [CrossRef] [Green Version]
- Veerareddygari, G.R.; Klusman, T.C.; Mueller, E.G. Characterization of the catalytic disulfide bond in E. coli 4-thiouridine synthetase to elucidate its functional quaternary structure. Protein Sci. 2016, 25, 1737–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.R.; Baltrus, T.; Reeve, J.N.; Daniels, C.J. Transfer RNA genes from the hyperthermophilic Archaeon, Methanopyrus kandleri. Biochim. Biophys. Acta 1992, 1132, 315–318. [Google Scholar] [CrossRef]
- Slesarev, A.I.; Mezhevaya, K.V.; Makarova, K.S.; Polushin, N.N.; Shcherbinina, O.V.; Shakhova, V.V.; Belova, G.I.; Aravind, L.; Natale, D.A.; Rogozin, I.B.; et al. The complete genome of hyperthermophile Methanopyrus kandleri AV19 and monophyly of archaeal methanogens. Proc. Natl. Acad. Sci. USA 2002, 99, 4644–4649. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.; Henderson, J.C.; Alfonzo, J.D. Multi-Substrate Specificity and the Evolutionary Basis for Interdependence in tRNA Editing and Methylation Enzymes. Front. Genet. 2019, 10, 104. [Google Scholar] [CrossRef]
- Hori, H. Methylated nucleosides in tRNA and tRNA methyltransferases. Front. Genet. 2014, 5, 144. [Google Scholar] [CrossRef] [Green Version]
- Yim, L.; Moukadiri, I.; Björk, G.R.; Armengod, M.E. Further insights into the tRNA modification process controlled by proteins MnmE and GidA of Escherichia coli. Nucleic Acids Res. 2006, 34, 5892–5905. [Google Scholar] [CrossRef] [Green Version]
- Meyer, S.; Scrima, A.; Versées, W.; Wittinghofer, A. Crystal structures of the conserved tRNA-modifying enzyme GidA: Implications for its interaction with MnmE and substrate. J. Mol. Biol. 2008, 380, 532–547. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Ito, K.; Inanaga, H.; Nureki, O.; Tomita, K.; Numata, T. Conserved cysteine residues of GidA are essential for biogenesis of 5-carboxymethylaminomethyluridine at tRNA anticodon. Structure 2009, 17, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, R.; Villarroya, M.; Ruiz-Partida, R.; Li, Y.; Proteau, A.; Prado, S.; Moukadiri, I.; Benítez-Páez, A.; Lomas, R.; Wagner, J.; et al. Structure-function analysis of Escherichia coli MnmG (GidA), a highly conserved tRNA-modifying enzyme. J. Bacteriol. 2009, 191, 7614–7619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moukadiri, I.; Prado, S.; Piera, J.; Velázquez-Campoy, A.; Björk, G.R.; Armengod, M.E. Evolutionarily conserved proteins MnmE and GidA catalyze the formation of two methyluridine derivatives at tRNA wobble positions. Nucleic Acids Res. 2009, 37, 7177–7193. [Google Scholar] [CrossRef] [Green Version]
- Moukadiri, I.; Garzón, M.J.; Björk, G.R.; Armengod, M.E. The output of the tRNA modification pathways controlled by the Escherichia coli MnmEG and MnmC enzymes depends on the growth conditions and the tRNA species. Nucleic Acids Res. 2014, 42, 2602–2623. [Google Scholar] [CrossRef]
- Nishimasu, H.; Ishitani, R.; Yamashita, K.; Iwashita, C.; Hirata, A.; Hori, H.; Nureki, O. Atomic structure of a folate/FAD-dependent tRNA T54 methyltransferase. Proc. Natl. Acad. Sci. USA 2009, 106, 8180–8185. [Google Scholar] [CrossRef] [Green Version]
- Hamdane, D.; Argentini, M.; Cornu, D.; Golinelli-Pimpaneau, B.; Fontecave, M. FAD/folate-dependent tRNA methyltransferase: Flavin as a new methyl-transfer agent. J. AM. Chem. Soc. 2012, 134, 19739–19745. [Google Scholar] [CrossRef]
- Schubert, H.G.; Blumenthal, R.M.; Cheng, X. Many paths to methyltransfer: A chronicle of convergence. Trends Biochem. Sci. 2003, 28, 329–332. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, V.; Koonin, E.V.; Aravind, L. SPOUT: A class of methyltransferases that includes spoU and trmD RNA methylase superfamilies, and novel superfamilies of predicted prokaryotic RNA methylases. J. Mol. Microbiol. Biotechnol. 2002, 4, 71–75. [Google Scholar]
- Kimura, S.; Miyauchi, K.; Ikeuchi, Y.; Thiaville, P.C.; Crécy-Lagard, V.D.; Suzuki, T. Discovery of the β-barrel-type RNA methyltransferase responsible for N6-methylation of N6-threonylcarbamoyladenosine in tRNAs. Nucleic Acids Res. 2014, 42, 9350–9365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, S.R.; Morales, M.J.; Li, J.M.; Hopper, A.K.; Martin, N.C. Isolation and characterization of the TRM1 locus, a gene essential for the N2,N2-dimethylguanosine modification of both mitochondrial and cytoplasmic tRNA in Saccharomyces cerevisiae. J. Biol. Chem. 1986, 261, 9703–9709. [Google Scholar] [CrossRef] [PubMed]
- Edqvist, J.; Blomqvist, K.; Stråby, K.B. Structural elements in yeast tRNAs required for homologous modification of guanosine-26 into dimethylguanosine-26 by the yeast Trm1 tRNA-modifying enzyme. Biochemistry 1994, 33, 9546–9551. [Google Scholar] [CrossRef] [PubMed]
- Constantinesco, F.; Benachenhou, N.; Motorin, Y.; Grosjean, H. The tRNA(guanine-26,N2-N2) methyltransferase (Trm1) from the hyperthermophilic archaeon Pyrococcus furiosus: Cloning, sequencing of the gene and its expression in Escherichia coli. Nucleic Acids Res. 1998, 26, 3753–3761. [Google Scholar] [CrossRef] [Green Version]
- Awai, T.; Kimura, S.; Tomikawa, C.; Ochi, A.; Ihsanawati; Bessho, Y.; Yokoyama, S.; Ohno, S.; Nishikawa, K.; Yokogawa, T.; et al. Aquifex aeolicus tRNA (N2,N2-guanine)-dimethyltransferase (Trm1) catalyzes transfer of methyl groups not only to guanine 26 but also to guanine 27 in tRNA. J. Biol. Chem. 2009, 284, 20467–20478. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, T.; Anraku, R.; Hasegawa, T.; Tomikawa, C.; Hori, H. Transfer RNA methyltransferases from Thermoplasma acidophilum, a thermoacidophilic archaeon. Int. J. Mol. Sci. 2014, 16, 91–113. [Google Scholar] [CrossRef]
- Dewe, J.M.; Fuller, B.L.; Lentini, J.M.; Kellner, S.M.; Fu, D. TRMT1-Catalyzed tRNA Modifications Are Required for Redox Homeostasis to Ensure Proper Cellular Proliferation and Oxidative Stress Survival. Mol. Cell Biol. 2017, 37, e00214–e00217. [Google Scholar] [CrossRef] [Green Version]
- Funk, H.M.; Zhao, R.; Thomas, M.; Spigelmyer, S.M.; Sebree, N.J.; Bales, R.O.; Burchett, J.B.; Mamaril, J.B.; Limbach, P.A.; Guy, M.P. Identification of the enzymes responsible for m2,2G and acp3U formation on cytosolic tRNA from insects and plants. PLoS ONE 2020, 15, e0242737. [Google Scholar] [CrossRef]
- Ihsanawati; Nishimoto, M.; Higashijima, K.; Shirouzu, M.; Grosjean, H.; Bessho, Y.; Yokoyama, S. Crystal structure of tRNA N2,N2-guanosine dimethyltransferase Trm1 from Pyrococcus horikoshii. J. Mol. Biol. 2008, 383, 871–884. [Google Scholar] [CrossRef]
- Awai, T.; Ochi, A.; Ihsanawati; Sengoku, T.; Hirata, A.; Bessho, Y.; Yokoyama, S.; Hori, H. Substrate tRNA recognition mechanism of a multisite-specific tRNA methyltransferase, Aquifex aeolicus Trm1, based on the X-ray crystal structure. J. Biol. Chem. 2011, 286, 35236–35246. [Google Scholar] [CrossRef] [Green Version]
- Brūmele, B.; Mutso, M.; Telanne, L.; Õunap, K.; Spunde, K.; Abroi, A.; Kurg, R. Human TRMT112-Methyltransferase Network Consists of Seven Partners Interacting with a Common Co-Factor. Int. J. Mol. Sci. 2021, 22, 13593. [Google Scholar] [CrossRef] [PubMed]
- Van Tran, N.; Muller, L.; Ross, R.L.; Lestini, R.; Létoquart, J.; Ulryck, N.; Limbach, P.A.; De Crécy-Lagard, V.; Cianférani, S.; Graille, M. Evolutionary insights into Trm112-methyltransferase holoenzymes involved in translation between archaea and eukaryotes. Nucleic Acids Res. 2018, 46, 8483–8499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, A.; Suzuki, T.; Nagano, T.; Fujii, D.; Okamoto, M.; Sora, M.; Lowe, T.M.; Kanai, T.; Atomi, H.; Suzuki, T.; et al. Distinct Modified Nucleosides in tRNATrp from the Hyperthermophilic Archaeon Thermococcus kodakarensis and Requirement of tRNA m2G10/m22G10 Methyltransferase (Archaeal Trm11) for Survival at High Temperatures. J. Bacteriol. 2019, 201, e00448–e00519. [Google Scholar] [CrossRef] [PubMed]
- Mazauric, M.H.; Dirick, L.; Purushothaman, S.K.; Björk, G.R.; Lapeyre, B. Trm112p is a 15-kDa zinc finger protein essential for the activity of two tRNA and one protein methyltransferases in yeast. J. Biol. Chem. 2010, 285, 18505–18515. [Google Scholar] [CrossRef] [Green Version]
- Bourgeois, G.; Létoquart, J.; Van Tran, N.; Graille, M. Trm112, a Protein Activator of Methyltransferases Modifying Actors of the Eukaryotic Translational Apparatus. Biomolecules 2017, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, M.P.; Phizicky, E.M. Two-subunit enzymes involved in eukaryotic post-transcriptional tRNA modification. RNA Biol. 2014, 11, 1608–1618. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Jora, M.; Solivio, B.; Thakur, P.; Acevedo-Rocha, C.G.; Randau, L.; de Crécy-Lagard, V.; Addepalli, B.; Limbach, P.A. tRNA Modification Profiles and Codon-Decoding Strategies in Methanocaldococcus jannaschii. J. Bacteriol. 2019, 201, e00690–e00718. [Google Scholar] [CrossRef] [Green Version]
- Wolff, P.; Villette, C.; Zumsteg, J.; Heintz, D.; Antoine, L.; Chane-Woon-Ming, B.; Droogmans, L.; Grosjean, H.; Westhof, E. Comparative patterns of modified nucleotides in individual tRNA species from a mesophilic and two thermophilic archaea. RNA 2020, 26, 1957–1975. [Google Scholar] [CrossRef]
- Gabant, G.; Auxilien, S.; Tuszynska, I.; Locard, M.; Gajda, M.J.; Chaussinand, G.; Fernandez, B.; Dedieu, A.; Grosjean, H.; Golinelli-Pimpaneau, B.; et al. THUMP from archaeal tRNA:m22G10 methyltransferase, a genuine autonomously folding domain. Nucleic Acids Res. 2006, 34, 2483–2494. [Google Scholar] [CrossRef] [Green Version]
- Fislage, M.; Roovers, M.; Tuszynska, I.; Bujnicki, J.M.; Droogmans, L.; Versées, W. Crystal structures of the tRNA:m2G6 methyltransferase Trm14/TrmN from two domains of life. Nucleic Acids Res. 2012, 40, 5149–5161. [Google Scholar] [CrossRef] [Green Version]
- Kalhor, H.R.; Clarke, S. Novel methyltransferase for modified uridine residues at the wobble position of tRNA. Mol. Cell. Biol. 2003, 23, 9283–9292. [Google Scholar] [CrossRef] [Green Version]
- White, J.; Li, Z.; Sardana, R.; Bujnicki, J.M.; Marcotte, E.M.; Johnson, A.W. Bud23 methylates G1575 of 18S rRNA and is required for efficient nuclear export of pre-40S subunits. Mol. Cell Biol. 2008, 28, 3151–3161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figaro, S.; Wacheul, L.; Schillewaert, S.; Graille, M.; Huvelle, E.; Mongeard, R.; Zorbas, C.; Lafontaine, D.L.; Heurgué-Hamard, V. Trm112 is required for Bud23-mediated methylation of the 18S rRNA at position G1575. Mol. Cell Biol. 2012, 32, 2254–2267. [Google Scholar] [CrossRef] [Green Version]
- Heurgué-Hamard, V.; Champ, S.; Mora, L.; Merkulova-Rainon, T.; Kisselev, L.L.; Buckingham, R.H. The glutamine residue of the conserved GGQ motif in Saccharomyces cerevisiae release factor eRF1 is methylated by the product of the YDR140w gene. J. Biol. Chem. 2005, 280, 2439–2445. [Google Scholar] [CrossRef] [Green Version]
- Polevoda, B.; Span, L.; Sherman, F. The yeast translation release factors Mrf1p and Sup45p (eRF1) are methylated, respectively, by the methyltransferases Mtq1p and Mtq2p. J. Biol. Chem. 2006, 281, 2562–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haag, S.; Kretschmer, J.; Bohnsack, M.T. WBSCR22/Merm1 is required for late nuclear pre-ribosomal RNA processing and mediates N7-methylation of G1639 in human 18S rRNA. RNA 2015, 21, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Van Tran, N.; Ernst, F.G.M.; Hawley, B.R.; Zorbas, C.; Ulryck, N.; Hackert, P.; Bohnsack, K.E.; Bohnsack, M.T.; Jaffrey, S.R.; Graille, M.; et al. The human 18S rRNA m6A methyltransferase METTL5 is stabilized by TRMT112. Nucleic Acids Res. 2019, 47, 7719–7733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figaro, S.; Scrima, N.; Buckingham, R.H.; Heurgué-Hamard, V. HemK2 protein, encoded on human chromosome 21, methylates translation termination factor eRF1. FEBS Lett. 2008, 582, 2352–2356. [Google Scholar] [CrossRef] [Green Version]
- Songe-Møller, L.; Van Den Born, E.; Leihne, V.; Vågbø, C.B.; Kristoffersen, T.; Krokan, H.E.; Kirpekar, F.; Falnes, P.Ø.; Klungland, A. Mammalian ALKBH8 possesses tRNA methyltransferase activity required for the biogenesis of multiple wobble uridine modifications implicated in translational decoding. Mol. Cell Biol. 2010, 30, 1814–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Brophy, J.A.; Chan, C.T.; Atmore, K.A.; Begley, U.; Paules, R.S.; Dedon, P.C.; Begley, T.J.; Samson, L.D. Human AlkB homolog ABH8 Is a tRNA methyltransferase required for wobble uridine modification and DNA damage survival. Mol. Cell Biol. 2010, 30, 2449–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Dai, Q.; Zhang, W.; Ren, J.; Pan, T.; He, C. The AlkB domain of mammalian ABH8 catalyzes hydroxylation of 5-methoxycarbonylmethyluridine at the wobble position of tRNA. Angew Chem. Int. Ed. Engl. 2010, 49, 8885–8888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Born, E.; Vågbø, C.B.; Songe-Møller, L.; Leihne, V.; Lien, G.F.; Leszczynska, G.; Malkiewicz, A.; Krokan, H.E.; Kirpekar, F.; Klungland, A.; et al. ALKBH8-mediated formation of a novel diastereomeric pair of wobble nucleosides in mammalian tRNA. Nat. Commun. 2011, 2, 172. [Google Scholar] [CrossRef] [Green Version]
- Yokogawa, T.; Nomura, Y.; Yasuda, A.; Ogino, H.; Hiura, K.; Nakada, S.; Oka, N.; Ando, K.; Kawamura, T.; Hirata, A.; et al. Identification of a radical SAM enzyme involved in the synthesis of archaeosine. Nat. Chem. Biol. 2019, 15, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Jin, M.; Zhang, W. Conservation and Diversification of tRNA t6A-Modifying Enzymes across the Three Domains of Life. Int. J. Mol. Sci. 2022, 23, 13600. [Google Scholar] [CrossRef]
- Krutyhołowa, R.; Zakrzewski, K.; Glatt, S. Charging the code—tRNA modification complexes. Curr. Opin. Struct. Biol. 2019, 55, 138–146. [Google Scholar] [CrossRef]
- Alexandrov, A.; Martzen, M.R.; Phizicky, E.M. Two proteins that form a complex are required for 7-methylguanosine modification of yeast tRNA. RNA 2002, 8, 1253–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, K.; Toyooka, T.; Tomikawa, C.; Ochi, A.; Takano, Y.; Takayanagi, N.; Endo, Y.; Hori, H. RNA recognition mechanism of eukaryote tRNA (m7G46) methyltransferase (Trm8-Trm82 complex). FEBS Lett. 2007, 581, 1599–1604. [Google Scholar] [CrossRef] [Green Version]
- Guy, M.P.; Podyma, B.M.; Preston, M.A.; Shaheen, H.H.; Krivos, K.L.; Limbach, P.A.; Hopper, A.K.; Phizicky, E.M. Yeast Trm7 interacts with distinct proteins for critical modifications of the tRNAPhe anticodon loop. RNA 2012, 18, 1921–1933. [Google Scholar] [CrossRef] [Green Version]
- Hirata, A.; Okada, K.; Yoshii, K.; Shiraishi, H.; Saijo, S.; Yonezawa, K.; Shimizu, N.; Hori, H. Structure of tRNA methyltransferase complex of Trm7 and Trm734 reveals a novel binding interface for tRNA recognition. Nucleic Acids Res. 2019, 47, 10942–10955. [Google Scholar] [CrossRef] [Green Version]
- Funk, H.M.; DiVita, D.J.; Sizemore, H.E.; Wehrle, K.; Miller, C.L.W.; Fraley, M.E.; Mullins, A.K.; Guy, A.R.; Phizicky, E.M.; Guy, M.P. Identification of a Trm732 Motif Required for 2’-O-methylation of the tRNA Anticodon Loop by Trm7. ACS Omega 2022, 7, 13667–13675. [Google Scholar] [CrossRef]
- Nishida, Y.; Ohmori, S.; Kakizono, R.; Kawai, K.; Namba, M.; Okada, K.; Yamagami, R.; Hirata, A.; Hori, H. Required Elements in tRNA for Methylation by the Eukaryotic tRNA (Guanine-N2-) Methyltransferase (Trm11-Trm112 Complex). Int. J. Mol. Sci. 2022, 23, 4046. [Google Scholar] [CrossRef]
- Bourgeois, G.; Marcoux, J.; Saliou, J.M.; Cianférani, S.; Graille, M. Activation mode of the eukaryotic m2G10 tRNA methyltransferase Trm11 by its partner protein Trm112. Nucleic Acids Res. 2017, 45, 1971–1982. [Google Scholar] [PubMed] [Green Version]
- Fitzek, E.; Joardar, A.; Gupta, R.; Geisler, M. Evolution of Eukaryal and Archaeal Pseudouridine Synthase Pus10. J. Mol. Evol. 2018, 86, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Ofengand, J. Ribosomal RNA pseudouridines and pseudouridine synthases. FEBS Lett. 2002, 514, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Hamma, T.; Ferré-D’Amaré, A.R. Pseudouridine synthases. Chem. Biol. 2006, 13, 1125–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spenkuch, F.; Motorin, Y.; Helm, M. Pseudouridine: Still mysterious, but never a fake (uridine)! RNA Biol. 2014, 11, 1540–1554. [Google Scholar] [CrossRef] [Green Version]
- Rintala-Dempsey, A.C.; Kothe, U. Eukaryotic stand-alone pseudouridine synthases—RNA modifying enzymes and emerging regulators of gene expression? RNA Biol. 2017, 14, 1185–1196. [Google Scholar] [CrossRef] [Green Version]
- Borchardt, E.K.; Martinez, N.M.; Gilbert, W.V. Regulation and Function of RNA Pseudouridylation in Human Cells. Annu. Rev. Genet. 2020, 54, 309–336. [Google Scholar] [CrossRef]
- Kaya, Y.; Ofengand, J. A novel unanticipated type of pseudouridine synthase with homologs in bacteria, archaea, and eukarya. RNA 2003, 9, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Gray, M.W. Evolutionary appearance of genes encoding proteins associated with box H/ACA snoRNAs: cbf5p in Euglena gracilis, an early diverging eukaryote, and candidate Gar1p and Nop10p homologs in archaebacteria. Nucleic Acids Res. 2000, 28, 2342–2352. [Google Scholar] [CrossRef] [Green Version]
- Joardar, A.; Jana, S.; Fitzek, E.; Gurha, P.; Majumder, M.; Chatterjee, K.; Geisler, M.; Gupta, R. Role of forefinger and thumb loops in production of Ψ54 and Ψ55 in tRNAs by archaeal Pus10. RNA 2013, 19, 1279–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Zhuang, Y.; Zhu, C.; Meng, H.; Lu, B.; Xie, B.; Peng, J.; Li, M.; Yi, C. Differential roles of human PUS10 in miRNA processing and tRNA pseudouridylation. Nat. Chem. Biol. 2020, 16, 160–169. [Google Scholar] [CrossRef]
- Ramabhadran, T.V.; Fossum, T.; Jagger, J. Escherichia coli mutant lacking 4-thiouridine in its transfer ribonucleic acid. J. Bacteriol. 1976, 128, 671–672. [Google Scholar] [CrossRef] [Green Version]
- Favre, A.; Yaniv, M.; Michelson, A.M. The photochemistry of 4-thiouridine in Escherichia coli t-RNAVał1. Biochem. Biophys. Res. Commun. 1969, 37, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Caldeira de Araujo, A.; Favre, A. Induction of size reduction in Escherichia coli by near-ultraviolet light. Eur. J. Biochem. 1985, 146, 605–610. [Google Scholar] [CrossRef]
- Caldeira de Araujo, A.; Favre, A. Near ultraviolet DNA damage induces the SOS responses in Escherichia coli. EMBO J. 1986, 5, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Saneyoshi, M.; Kumagai, I.; Miura, K.; Watanabe, K. Effects of modification of 4-thiouridine in E. coli tRNA(fMet) on its methyl acceptor activity by thermostable Gm-methylases. J. Biochem. 1989, 106, 798–802. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, J.A.; Graham, D.E.; Zou, S.; Crain, P.F.; Ibba, M.; Konisky, J.; Soll, D.; Olsen, G.J. Post-transcriptional modification in archaeal tRNAs: Identities and phylogenetic relations of nucleotides from mesophilic and hyperthermophilic Methanococcales. Nucleic Acids Res. 2001, 29, 4299–4706. [Google Scholar] [CrossRef]
- Kimura, S.; Waldor, M.K. The RNA degradosome promotes tRNA quality control through clearance of hypomodified tRNA. Proc. Natl. Acad. Sci. USA 2019, 116, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Orita, I.; Futatsuishi, R.; Adachi, K.; Ohira, T.; Kaneko, A.; Minowa, K.; Suzuki, M.; Tamura, T.; Nakamura, S.; Imanaka, T.; et al. Random mutagenesis of a hyperthermophilic archaeon identified tRNA modifications associated with cellular hyperthermotolerance. Nucleic Acids Res. 2019, 47, 1964–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigi, N.; Suzuki, T.; Terada, T.; Shirouzu, M.; Yokoyama, S.; Watanabe, K. Temperature-dependent biosynthesis of 2-thioribothymidine of Thermus thermophilus tRNA. J. Biol. Chem. 2006, 281, 2104–2113. [Google Scholar] [CrossRef] [Green Version]
- Tomikawa, C.; Yokogawa, T.; Kanai, T.; Hori, H. N7-Methylguanine at position 46 (m7G46) in tRNA from Thermus thermophilus is required for cell viability at high temperatures through a tRNA modification network. Nucleic Acids Res. 2010, 38, 942–957. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Kunibayashi, T.; Tomikawa, C.; Ochi, A.; Kanai, T.; Hirata, A.; Iwashita, C.; Hori, H. Pseudouridine at position 55 in tRNA controls the contents of other modified nucleotides for low-temperature adaptation in the extreme-thermophilic eubacterium Thermus thermophilus. Nucleic Acids Res. 2011, 39, 1304–2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagami, R.; Tomikawa, C.; Shigi, N.; Kazayama, A.; Asai, S.; Takuma, H.; Hirata, A.; Fourmy, D.; Asahara, H.; Watanabe, K.; et al. Folate-/FAD-dependent tRNA methyltransferase from Thermus thermophilus regulates other modifications in tRNA at low temperatures. Genes Cells 2016, 21, 740–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, D.F.; De Crécy-Lagard, V.; Bishop, A.C. Molecular determinants of dihydrouridine synthase activity. FEBS Lett. 2006, 580, 5198–5202. [Google Scholar] [CrossRef] [Green Version]
- Bou-Nader, C.; Montémont, H.; Guérineau, V.; Jean-Jean, O.; Brégeon, D.; Hamdane, D. Unveiling structural and functional divergences of bacterial tRNA dihydrouridine synthases: Perspectives on the evolution scenario. Nucleic Acids Res. 2018, 46, 1386–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finet, O.; Yague-Sanz, C.; Marchand, F.; Hermand, D. The Dihydrouridine landscape from tRNA to mRNA: A perspective on synthesis, structural impact and function. RNA Biol. 2022, 19, 735–750. [Google Scholar] [CrossRef]
- Kusuba, H.; Yoshida, T.; Iwasaki, E.; Awai, T.; Kazayama, A.; Hirata, A.; Tomikawa, C.; Yamagami, R.; Hori, H. In vitro dihydrouridine formation by tRNA dihydrouridine synthase from Thermus thermophilus, an extreme-thermophilic eubacterium. J. Biochem. 2015, 158, 513–521. [Google Scholar]
- Brégeon, D.; Pecqueur, L.; Toubdji, S.; Sudol, C.; Lombard, M.; Fontecave, M.; De Crécy-Lagard, V.; Motorin, Y.; Helm, M.; Hamdane, D. Dihydrouridine in the Transcriptome: New Life for This Ancient RNA Chemical Modification. ACS Chem Biol. 2022, 17, 1638–1657. [Google Scholar] [CrossRef] [PubMed]
- Oshima, T.; Moriya, T.; Terui, Y. Identification, chemical synthesis, and biological functions of unusual polyamines produced by extreme thermophiles. Methods Mol. Biol. 2011, 720, 81–111. [Google Scholar] [PubMed]
- Hamana, K.; Tanaka, T.; Hosoya, R.; Niitsu, M.; Itoh, T. Cellular polyamines of the acidophilic, thermophilic and thermoacidophilic archaebacteria, Acidilobus, Ferroplasma, Pyrobaculum, Pyrococcus, Staphylothermus, Thermococcus, Thermodiscus and Vulcanisaeta. J. Gen. Appl. Microbiol. 2003, 49, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayrapetyan, A.; Grosjean, H.; Helm, M. Effect of a quaternary pentamine on RNA stabilization and enzymatic methylation. Biol. Chem. 2009, 390, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Terui, Y.; Nakamoto, C.; Iwashita, C.; Ochi, A.; Watanabe, K.; Oshima, T. Effects of polyamines from Thermus thermophilus, an extreme-thermophilic eubacterium, on tRNA methylation by tRNA (Gm18) methyltransferase (TrmH). J. Biochem. 2016, 159, 509–517. [Google Scholar] [CrossRef]
- Nakashima, M.; Yamagami, R.; Tomikawa, C.; Ochi, Y.; Moriya, T.; Asahara, H.; Fourmy, D.; Yoshizawa, S.; Oshima, T.; Hori, H. Long and branched polyamines are required for maintenance of the ribosome, tRNAHis and tRNATyr in Thermus thermophilus cells at high temperatures. Genes Cells 2017, 22, 628–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keith, G.; Heyman, T. Heterogeneities in vertebrate tRNAs(Trp) avian retroviruses package only as a primer the tRNA(Trp) lacking modified m2G in position 7. Nucleic Acids Res. 1990, 18, 703–710. [Google Scholar] [CrossRef]
- Matsuo, M.; Yokogawa, T.; Nishikawa, K.; Watanabe, K.; Okada, N. Highly specific and efficient cleavage of squid tRNALys catalyzed by magnesium ions. J. Biol. Chem. 1995, 270, 10097–10104. [Google Scholar] [CrossRef] [Green Version]
- Kotelawala, L.; Grayhack, E.J.; Phizicky, E.M. Identification of yeast tRNA Um(44) 2’-O-methyltransferase (Trm44) and demonstration of a Trm44 role in sustaining levels of specific tRNA(Ser) species. RNA 2008, 14, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Dewe, J.M.; Whipple, J.M.; Chernyakov, I.; Jaramillo, L.N.; Phizicky, E.M. The yeast rapid tRNA decay pathway competes with elongation factor 1A for substrate tRNAs and acts on tRNAs lacking one or more of several modifications. RNA 2012, 18, 1886–1896. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chen, C.; Johansson, M.J. The pre-mRNA retention and splicing complex controls tRNA maturation by promoting TAN1 expression. Nucleic Acids Res. 2013, 41, 5669–5678. [Google Scholar] [CrossRef] [Green Version]
- Broly, M.; Polevoda, B.V.; Awayda, K.M.; Tong, N.; Lentini, J.; Besnard, T.; Deb, W.; O’Rourke, D.; Baptista, J.; Ellard, S.; et al. THUMPD1 bi-allelic variants cause loss of tRNA acetylation and a syndromic neurodevelopmental disorder. Am. J. Hum. Genet. 2022, 109, 587–600. [Google Scholar] [CrossRef]
- Zang, X.; Jiang, G.; Sun, M.; Zhou, H.; Miao, Y.; Liang, M.; Wang, E.; Zhang, Y. Cytosolic THUMPD1 promotes breast cancer cells invasion and metastasis via the AKT-GSK3-Snail pathway. Oncotarget 2017, 8, 13357–13366. [Google Scholar] [CrossRef] [Green Version]
- Zang, H.; Hou, W.; Wang, H.-L.; Liu, H.-J.; Jia, X.-Y.; Zheng, X.-Z.; Zou, Y.-X.; Li, X.; Hou, L.; McNutt, M.A.; et al. GSK-3β-regulated N-acetyltransferase 10 is involved in colorectal cancer invasion. Clin. Cancer Res. 2014, 20, 4717–4729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, J.; Yan, S.; Huang, K.; Bai, Y.; Zheng, S. High expression of N-acetyltransferase 10: A novel independent prognostic marker of worse outcome in patients with hepatocellular carcinoma. Int. J. Exp. Pathol. 2015, 8, 14765–14771. [Google Scholar]
- Gurha, P.; Joardar, A.; Chaurasia, P.; Gupta, R. Differential roles of archaeal box H/ACA proteins in guide RNA-dependent and independent pseudouridine formation. RNA Biol. 2007, 4, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Nordlund, M.E.; Johansson, J.O.; Von Pawel-Rammingen, U.; Byström, A.S. Identification of the TRM2 gene encoding the tRNA(m5U54)methyltransferase of Saccharomyces cerevisiae. RNA 2000, 6, 844–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, H.F.; Motorin, Y.; Planta, R.J.; Grosjean, H. The yeast gene YNL292w encodes a pseudouridine synthase (Pus4) catalyzing the formation of psi55 in both mitochondrial and cytoplasmic tRNAs. Nucleic Acids Res. 1997, 25, 4493–4499. [Google Scholar] [CrossRef]
- Nagato, Y.; Tomikawa, C.; Yamaji, H.; Soma, A.; Takai, K. Intron-Dependent or Independent Pseudouridylation of Precursor tRNA Containing Atypical Introns in Cyanidioschyzon merolae. Int. J. Mol. Sci. 2022, 23, 12058. [Google Scholar] [CrossRef]
- Chatterjee, K.; Blaby, I.K.; Thiaville, P.C.; Majumder, M.; Grosjean, H.; Yuan, Y.A.; Gupta, R.; De Crécy-Lagard, V. The archaeal COG1901/DUF358 SPOUT-methyltransferase members, together with pseudouridine synthase Pus10, catalyze the formation of 1-methylpseudouridine at position 54 of tRNA. RNA 2012, 18, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Wurm, J.P.; Griese, M.; Bahr, U.; Held, M.; Heckel, A.; Karas, M.; Soppa, J.; Wöhnert, J. Identification of the enzyme responsible for N1-methylation of pseudouridine 54 in archaeal tRNAs. RNA 2012, 18, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Rose, S.; Auxilien, S.; Havelund, J.F.; Kirpekar, F.; Huber, H.; Grosjean, H.; Douthwaite, S. The hyperthermophilic partners Nanoarchaeum and Ignicoccus stabilize their tRNA T-loops via different but structurally equivalent modifications. Nucleic Acids Res. 2020, 48, 6906–6918. [Google Scholar] [CrossRef]
- Shigi, N.; Sakaguchi, Y.; Suzuki, T.; Watanabe, K. Identification of two tRNA thiolation genes required for cell growth at extremely high temperatures. J. Biol. Chem. 2006, 281, 14296–14306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaby, I.K.; Majumder, M.; Chatterjee, K.; Jana, S.; Grosjean, H.; De Crécy-Lagard, V.; Gupta, R. Pseudouridine formation in archaeal RNAs: The case of Haloferax volcanii. RNA 2011, 17, 1367–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festen, E.A.; Goyette, P.; Green, T.; Boucher, G.; Beauchamp, C.; Trynka, G.; Dubois, P.C.; Lagacé, C.; Stokkers, P.C.; Hommes, D.W.; et al. A meta-analysis of genome-wide association scans identifies IL18RAP, PTPN2, TAGAP, and PUS10 as shared risk loci for Crohn’s disease and celiac disease. PLoS Genet. 2011, 7, e1001283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Structures of modified nucleosides, which are produced by THUMP-related tRNA modification enzymes, and their positions in tRNA. (A) Structures of modified nucleosides, which are produced by THUMP-related tRNA modification enzymes. Modifications are indicated in red. Because uridine is produced by deamination of cytidine, the 4-O atom is colored in red. Because pseudouridine is synthesized by isomerization of uridine, the uracil base is enclosed in a red circle. (B) The typical tRNA structure is represented as a cloverleaf model. The numbers show the positions in tRNA. Conserved residues in tRNA are shown as letters: abbreviations, R, purine; Y, pyrimidine. Position 8 is conserved as U (red) in almost all tRNAs; however, in the case of M. kandleri, position 8 in precursor tRNA is C (orange). The colors correspond to the modified nucleosides in A: blue, m2G (and m22G); red, s4U; orange, U; cyan, ac4C; purple, Ψ. T. kodakarensis NAT10 homolog acetylates multiple positions in tRNA as described in the main text. (C) The modification positions are mapped on the L-shaped yeast tRNAPhe structure.
Figure 1.
Structures of modified nucleosides, which are produced by THUMP-related tRNA modification enzymes, and their positions in tRNA. (A) Structures of modified nucleosides, which are produced by THUMP-related tRNA modification enzymes. Modifications are indicated in red. Because uridine is produced by deamination of cytidine, the 4-O atom is colored in red. Because pseudouridine is synthesized by isomerization of uridine, the uracil base is enclosed in a red circle. (B) The typical tRNA structure is represented as a cloverleaf model. The numbers show the positions in tRNA. Conserved residues in tRNA are shown as letters: abbreviations, R, purine; Y, pyrimidine. Position 8 is conserved as U (red) in almost all tRNAs; however, in the case of M. kandleri, position 8 in precursor tRNA is C (orange). The colors correspond to the modified nucleosides in A: blue, m2G (and m22G); red, s4U; orange, U; cyan, ac4C; purple, Ψ. T. kodakarensis NAT10 homolog acetylates multiple positions in tRNA as described in the main text. (C) The modification positions are mapped on the L-shaped yeast tRNAPhe structure.

Figure 2.
The modification pathways of THUMP-related tRNA methyltransferases. Eukaryotic, archaeal and bacterial enzymes are colored in red, blue and orange, respectively. The modification sites and modified nucleosides are enclosed by squares. (A) ArcTrm11 from P. abyssi and T. kodakarensis produces m2G10 and m22G10. The m22G10 modification is produced by the second methylation from m2G10. (B) S. cerevisiae Trm11 required a partner protein (Trm112) for the methylation and produces only m2G10. (C) ArcTrm11 from A. fulgidus and Halloferax volcanii requires a partner protein (arcTrm112) and produces both m2G10 and m22G10. (D) TrmN produces m2G6 from G6. (E) Trm14 produces m2G6 from G6. “?” means that T. kodakarensis Trm14 may produce m2G67 as well as m2G6; this modification has not been confirmed by purified protein. (F) Human THUMP3-TRMT112 complex produces m2G6 and m2G7 from G6 and G7, respectively.
Figure 2.
The modification pathways of THUMP-related tRNA methyltransferases. Eukaryotic, archaeal and bacterial enzymes are colored in red, blue and orange, respectively. The modification sites and modified nucleosides are enclosed by squares. (A) ArcTrm11 from P. abyssi and T. kodakarensis produces m2G10 and m22G10. The m22G10 modification is produced by the second methylation from m2G10. (B) S. cerevisiae Trm11 required a partner protein (Trm112) for the methylation and produces only m2G10. (C) ArcTrm11 from A. fulgidus and Halloferax volcanii requires a partner protein (arcTrm112) and produces both m2G10 and m22G10. (D) TrmN produces m2G6 from G6. (E) Trm14 produces m2G6 from G6. “?” means that T. kodakarensis Trm14 may produce m2G67 as well as m2G6; this modification has not been confirmed by purified protein. (F) Human THUMP3-TRMT112 complex produces m2G6 and m2G7 from G6 and G7, respectively.

Figure 3.
Structures of B. anthracis ThiI and P. horikoshii ThiI-like (PH1313) protein. (A) Structure of B. anthracis ThiI (PDB code: 2C5S) is represented by a cartoon model. Ferredoxin-like, THUMP and PP-loop domains are colored in green, red and blue, respectively. N and C show the N- and C-termini, respectively. Bound AMP is shown as a stick model. (B) Structure of P. horokoshii ThiI-like (PH1313) protein (PDB code: 1VBK) is shown by a cartoon model. Although this protein structure was solved as a dimer, only one subunit is shown. Ferredoxin-like, THUMP and PP-loop domains are colored in green, red and blue, respectively. The size of the PP-loop domain of this protein is smaller than that of B. anthracis ThiI due to the deletion of the C-terminal region.
Figure 3.
Structures of B. anthracis ThiI and P. horikoshii ThiI-like (PH1313) protein. (A) Structure of B. anthracis ThiI (PDB code: 2C5S) is represented by a cartoon model. Ferredoxin-like, THUMP and PP-loop domains are colored in green, red and blue, respectively. N and C show the N- and C-termini, respectively. Bound AMP is shown as a stick model. (B) Structure of P. horokoshii ThiI-like (PH1313) protein (PDB code: 1VBK) is shown by a cartoon model. Although this protein structure was solved as a dimer, only one subunit is shown. Ferredoxin-like, THUMP and PP-loop domains are colored in green, red and blue, respectively. The size of the PP-loop domain of this protein is smaller than that of B. anthracis ThiI due to the deletion of the C-terminal region.

Figure 4.
(A) Secondary structure of minimum substrate RNA for ThiI. The modification position (U8) is colored in red. This RNA is a truncated RNA of E. coli tRNAPhe. The secondary structure is based on the complex of minimum substrate RNA and ThiI shown in panel B. (B) Crystal structure of the complex of the minimum substrate and T. maritima ThiI (PDB code: 4KR6). ThiI forms a dimer structure. To distinguish between the two subunits, one subunit is colored in pale green. The ferredoxin-like, THUMP and PP-loop domains in one subunit are colored in yellow, magenta and pale blue, respectively. The THUMP domain captures the CCA terminus of one minimum substrate RNA. The PP-loop domain in this subunit accesses U8 (red) in another minimum substrate RNA.
Figure 4.
(A) Secondary structure of minimum substrate RNA for ThiI. The modification position (U8) is colored in red. This RNA is a truncated RNA of E. coli tRNAPhe. The secondary structure is based on the complex of minimum substrate RNA and ThiI shown in panel B. (B) Crystal structure of the complex of the minimum substrate and T. maritima ThiI (PDB code: 4KR6). ThiI forms a dimer structure. To distinguish between the two subunits, one subunit is colored in pale green. The ferredoxin-like, THUMP and PP-loop domains in one subunit are colored in yellow, magenta and pale blue, respectively. The THUMP domain captures the CCA terminus of one minimum substrate RNA. The PP-loop domain in this subunit accesses U8 (red) in another minimum substrate RNA.

Figure 5.
(A) Structure of mini-helix RNA. The modification position (C8) is colored in red. (B) Crystal structure of CDAT8. CDAT8 forms a dimer structure. To distinguish subunits, one subunit is colored in pale green. Deaminase, ferredoxin-like and THUMP domains in the other subunit are colored in pale blue, yellow and magenta, respectively.
Figure 5.
(A) Structure of mini-helix RNA. The modification position (C8) is colored in red. (B) Crystal structure of CDAT8. CDAT8 forms a dimer structure. To distinguish subunits, one subunit is colored in pale green. Deaminase, ferredoxin-like and THUMP domains in the other subunit are colored in pale blue, yellow and magenta, respectively.

Figure 6.
Structures of P. abyssi Trm14 ((A): PDB code, 3TM4), T. thermophilus TrmN ((B): PDB code, 3TMA) and T. kodakarensis arcTrm11 ((C): PDB code, 5E71) are compared. The N-terminal ferredoxin-like domain, THUMP domain, Rossmann fold methyltransferase (methylase) domain and linker region are colored in yellow, red, blue and orange, respectively. Trm14 and TrmN modify G6 in tRNA while arcTrm11 modifies G10. The modification sites (G6 and G10) are mapped onto the L-shaped tRNA structure (D). G6, G10 and CCA terminus are highlighted as stick models. The distance between the THUMP and methylase domains of Trm14 and TrmN is shorter than that seen in arcTrm11. Because the THUMP domain captures the CCA terminus in tRNA, this short distance between the THUMP and methylase domains of Trm14 and TrmN enables the catalytic pocket in the methylase domain to access the modification site G6. In contrast, the longer distance between the THUMP and methylase domains of arcTrm11 is required for the positioning of the catalytic pocket with respect to the modification site G10. Thus, the N-terminal ferredoxin-like domain and linker region are important for the maintenance of the distance and angle between the THUMP and methylase domains, which decides the modification site in tRNA.
Figure 6.
Structures of P. abyssi Trm14 ((A): PDB code, 3TM4), T. thermophilus TrmN ((B): PDB code, 3TMA) and T. kodakarensis arcTrm11 ((C): PDB code, 5E71) are compared. The N-terminal ferredoxin-like domain, THUMP domain, Rossmann fold methyltransferase (methylase) domain and linker region are colored in yellow, red, blue and orange, respectively. Trm14 and TrmN modify G6 in tRNA while arcTrm11 modifies G10. The modification sites (G6 and G10) are mapped onto the L-shaped tRNA structure (D). G6, G10 and CCA terminus are highlighted as stick models. The distance between the THUMP and methylase domains of Trm14 and TrmN is shorter than that seen in arcTrm11. Because the THUMP domain captures the CCA terminus in tRNA, this short distance between the THUMP and methylase domains of Trm14 and TrmN enables the catalytic pocket in the methylase domain to access the modification site G6. In contrast, the longer distance between the THUMP and methylase domains of arcTrm11 is required for the positioning of the catalytic pocket with respect to the modification site G10. Thus, the N-terminal ferredoxin-like domain and linker region are important for the maintenance of the distance and angle between the THUMP and methylase domains, which decides the modification site in tRNA.

Figure 7.
(A) Recognition sites of S. cerevisiae Trm11-Trm112 are marked on the L-shaped tRNA structure. The modification site (G10) and other recognition sites are colored in red and magenta, respectively. S. cerevisiae Trm11-Trm112 methylates standard tRNAs, which possess a regular size (5 nt) variable region, G10-C25 base pair and purine38 in addition to the CCA terminus. (B) Crystal structure of A. fulgidus arcTrm11-arcTrm112 (PDB code, 6ZXW) is represented by a cartoon model. The ferredoxin-like domain, THUMP domain, Rossmann fold methylase domain, and linker region are colored in yellow, red, blue and orange, respectively. Archaeal Trm112 is colored in green.
Figure 7.
(A) Recognition sites of S. cerevisiae Trm11-Trm112 are marked on the L-shaped tRNA structure. The modification site (G10) and other recognition sites are colored in red and magenta, respectively. S. cerevisiae Trm11-Trm112 methylates standard tRNAs, which possess a regular size (5 nt) variable region, G10-C25 base pair and purine38 in addition to the CCA terminus. (B) Crystal structure of A. fulgidus arcTrm11-arcTrm112 (PDB code, 6ZXW) is represented by a cartoon model. The ferredoxin-like domain, THUMP domain, Rossmann fold methylase domain, and linker region are colored in yellow, red, blue and orange, respectively. Archaeal Trm112 is colored in green.

Figure 8.
Structure of human PUS10 (PDB code, 2V9K) is represented by a cartoon model. N-terminal accessory and C-terminal pseudouridine synthase domains are colored in green and pale blue, respectively. The THUMP-related structure in the accessory domain is enclosed by a red circle. One Zn atom (magenta) is bound in the accessory domain.
Figure 8.
Structure of human PUS10 (PDB code, 2V9K) is represented by a cartoon model. N-terminal accessory and C-terminal pseudouridine synthase domains are colored in green and pale blue, respectively. The THUMP-related structure in the accessory domain is enclosed by a red circle. One Zn atom (magenta) is bound in the accessory domain.


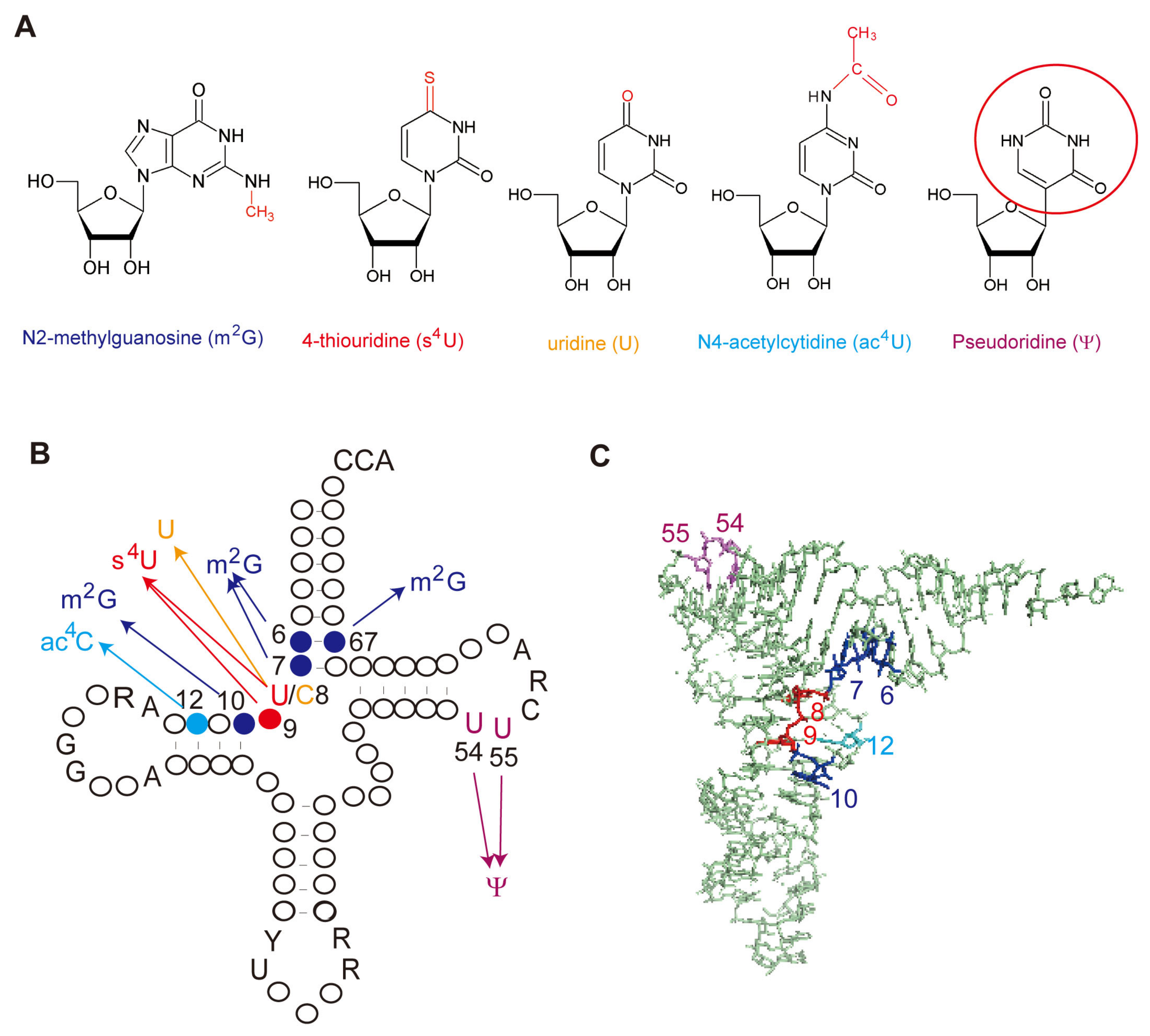{kind=link}
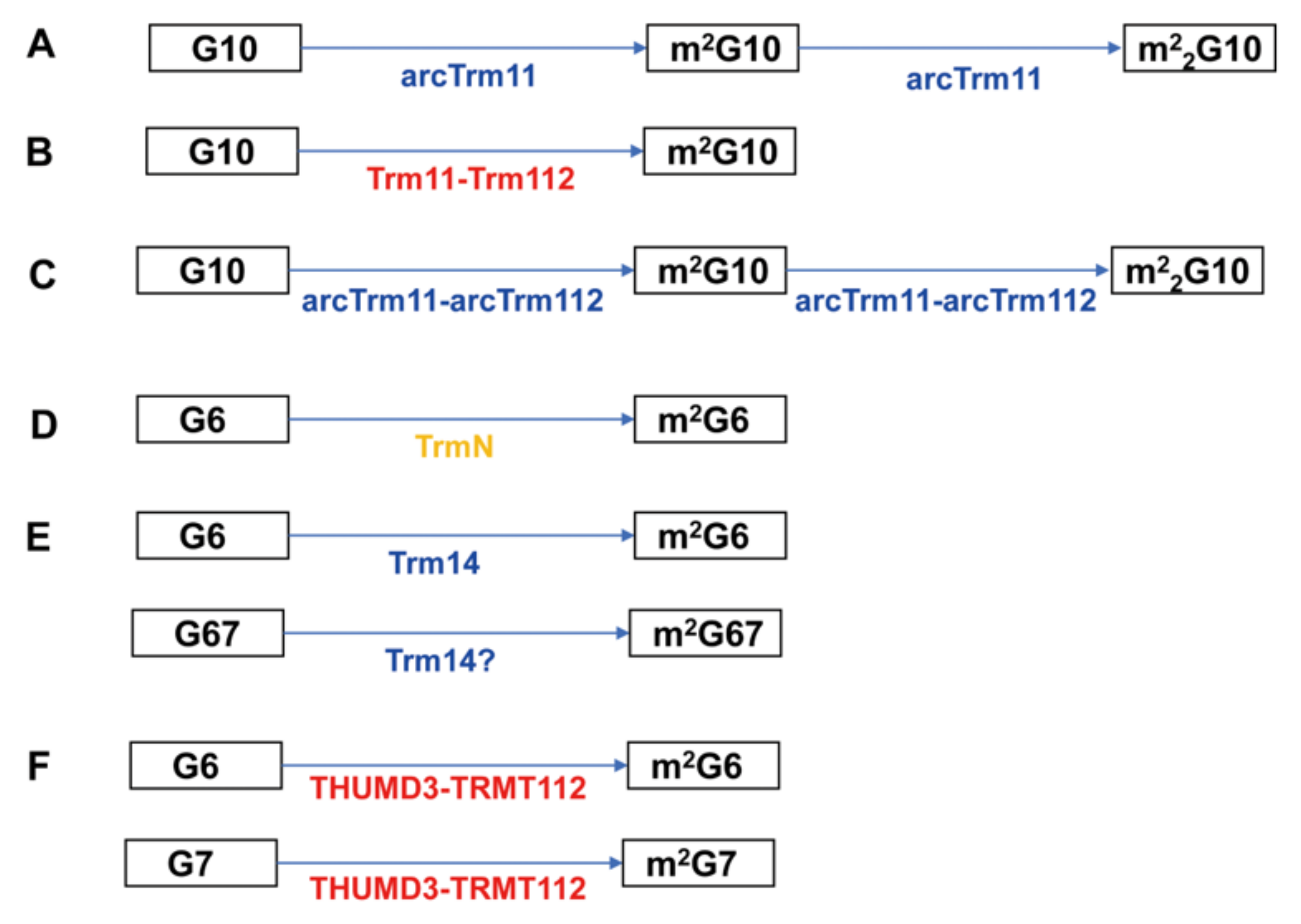{kind=link}
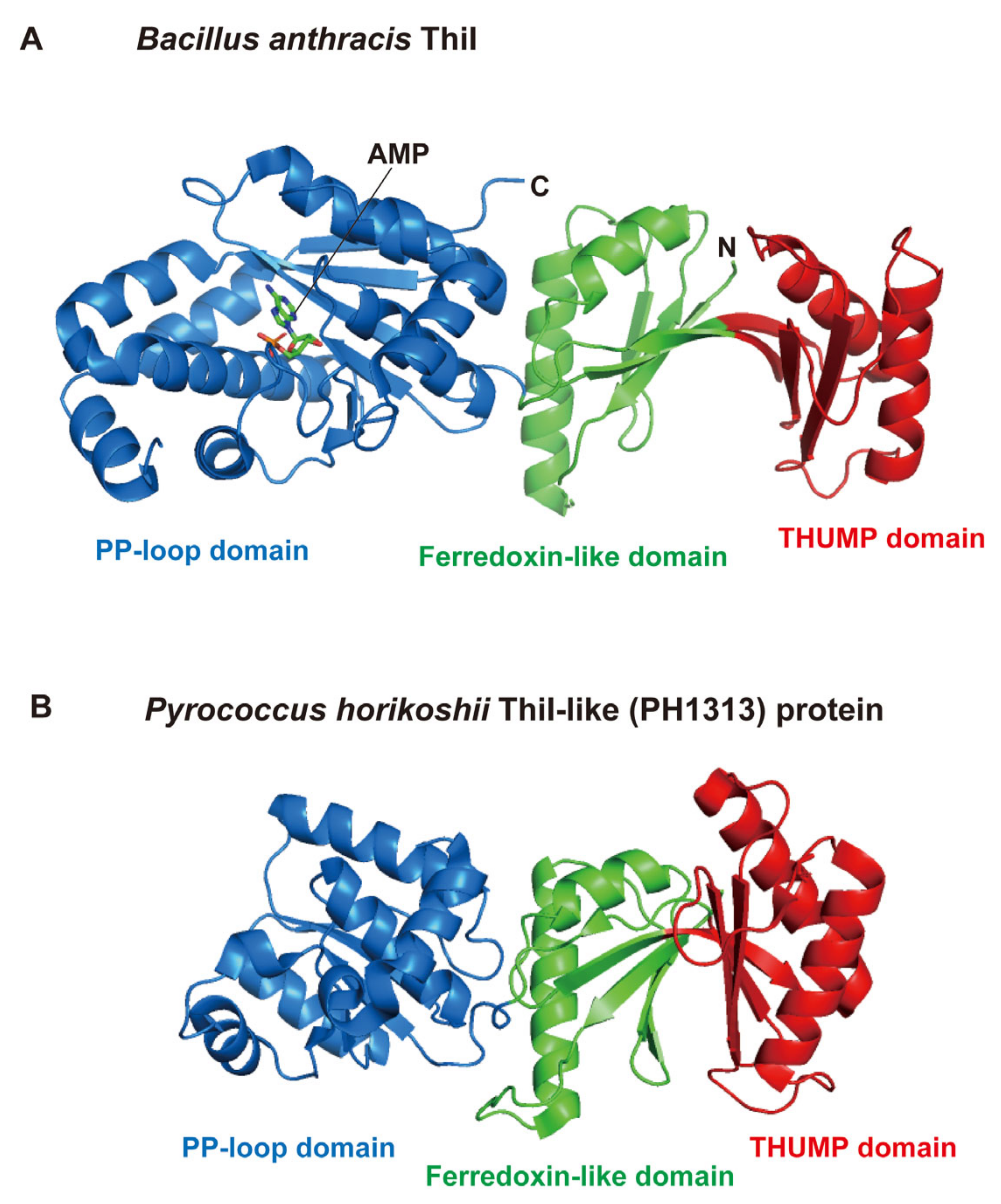{kind=link}
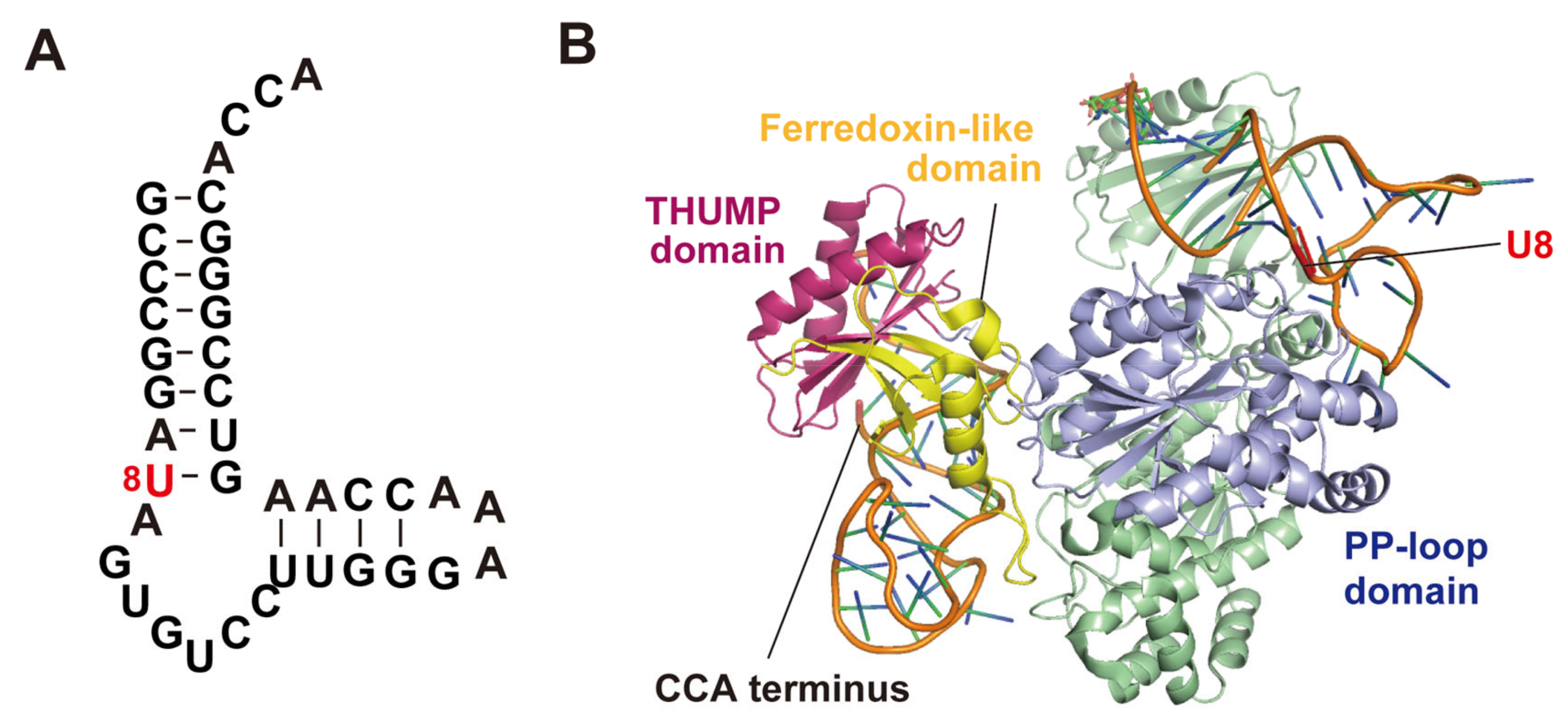{kind=link}
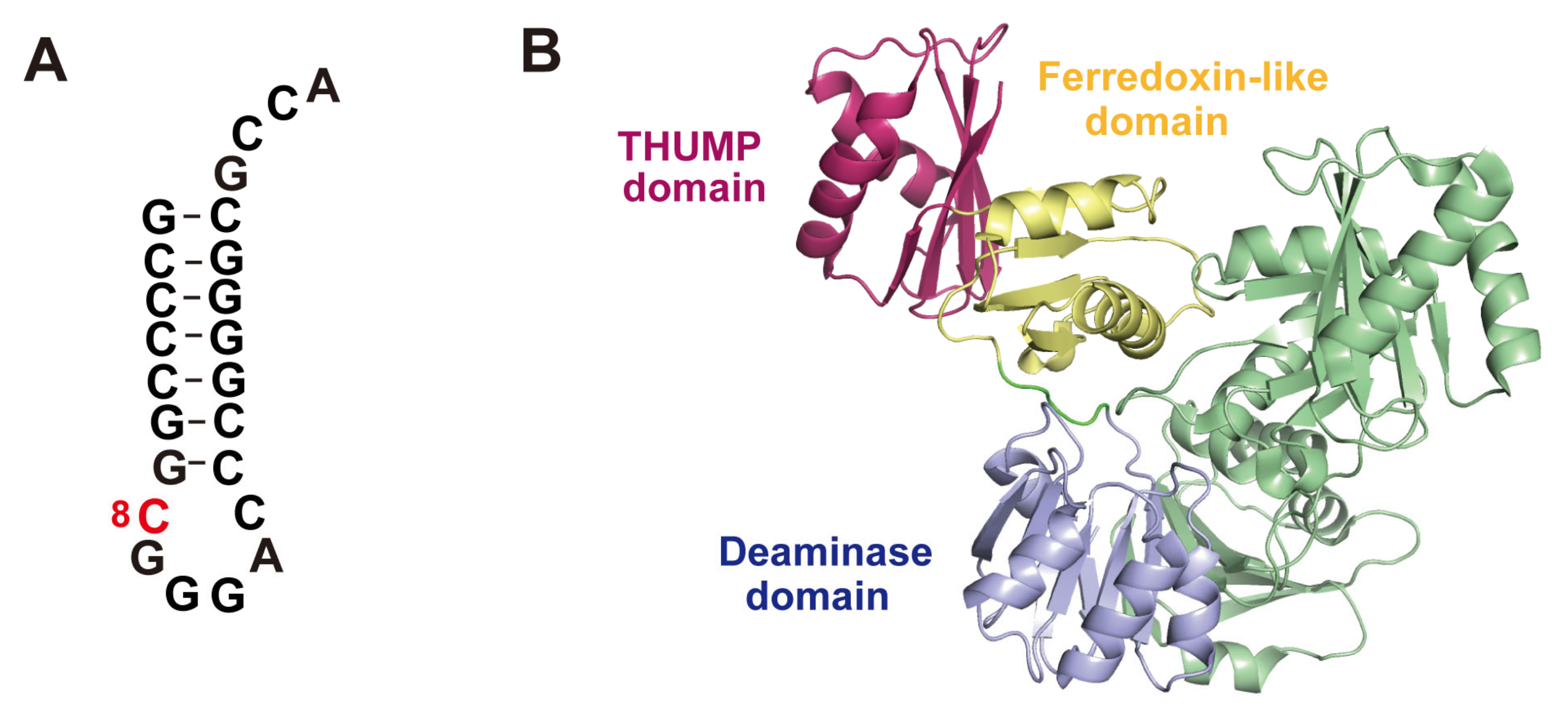{kind=link}
{kind=link}
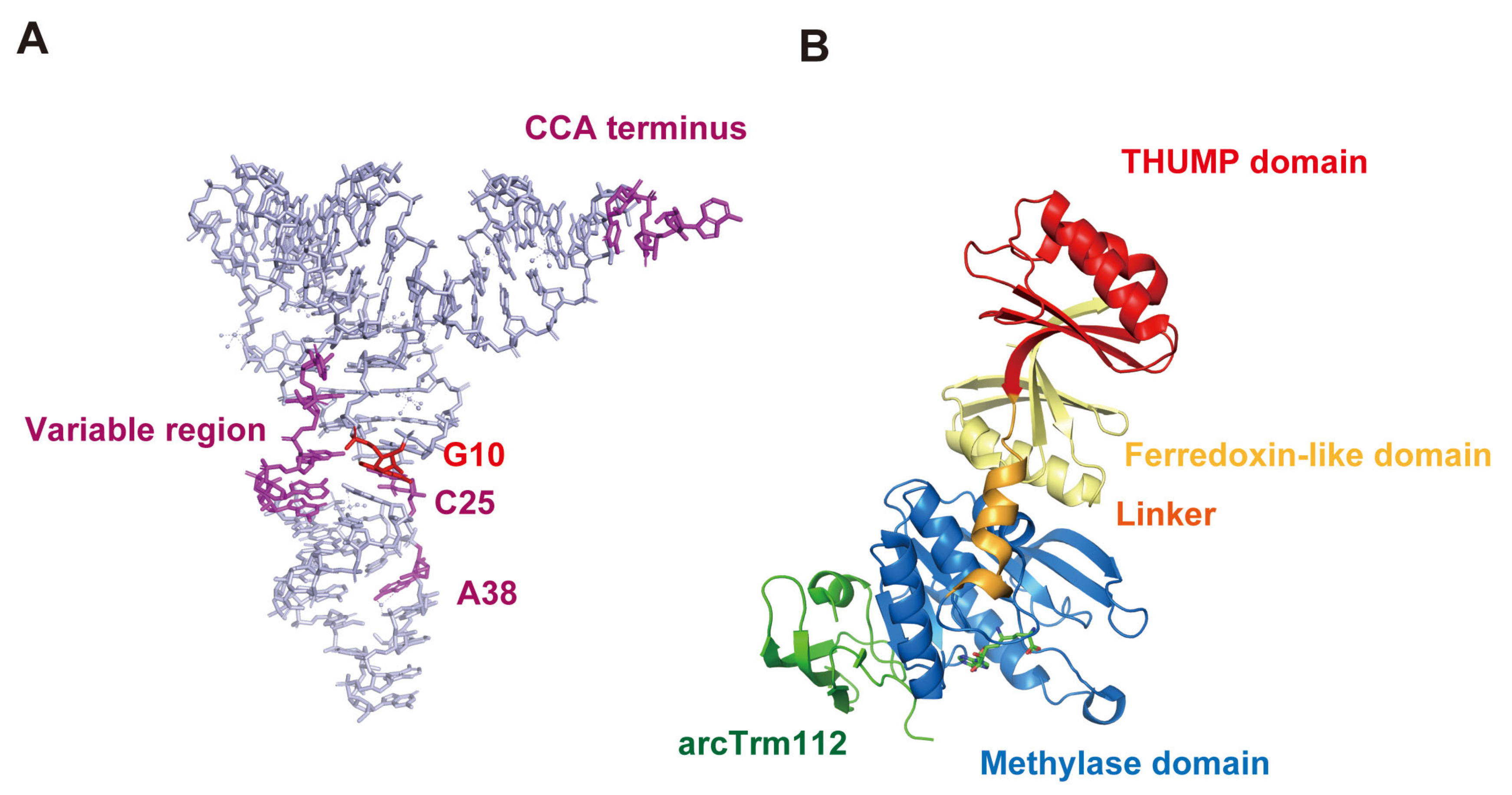{kind=link}
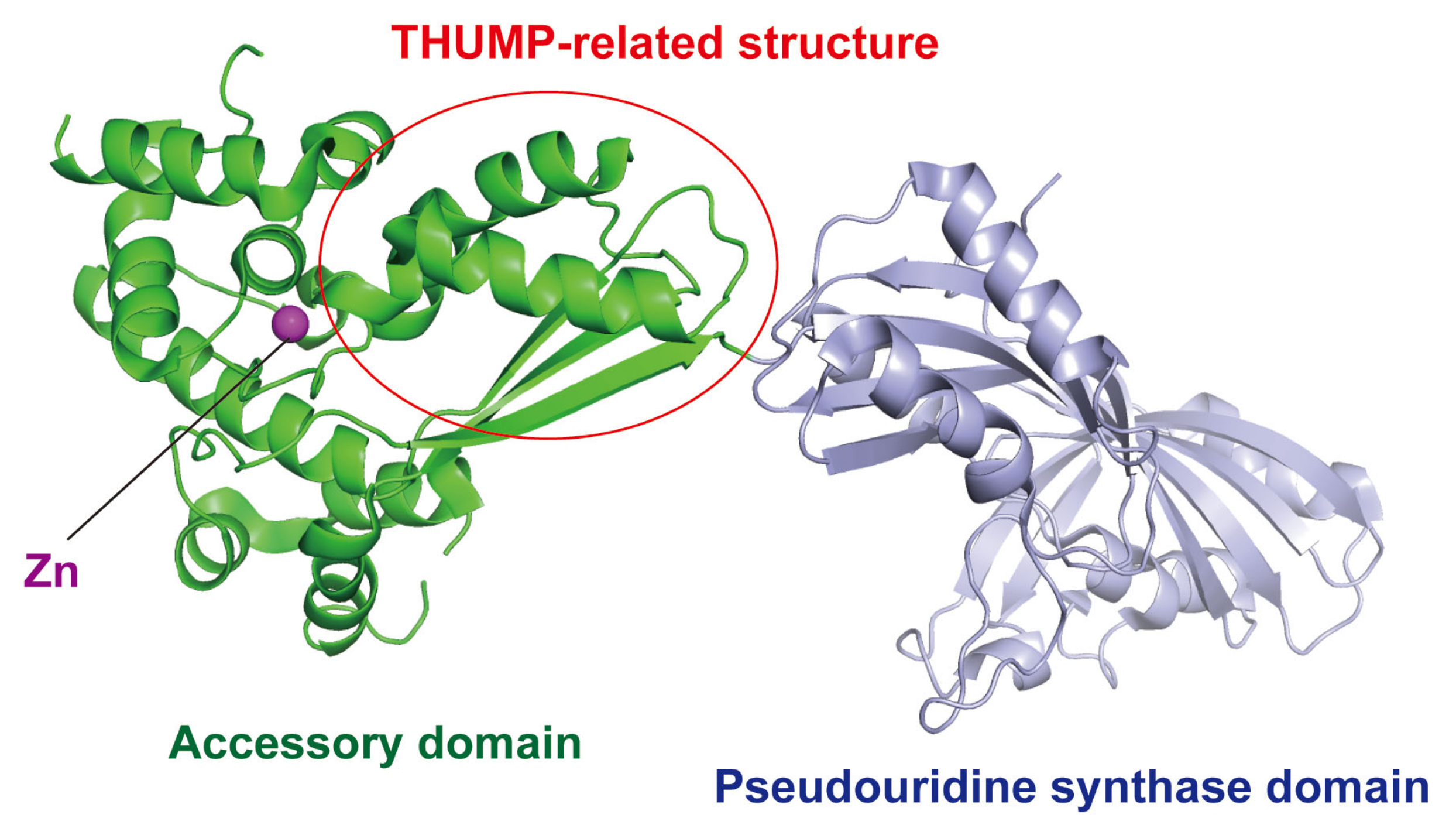{kind=link}
Table 1.
Classification of tRNA modification enzymes with a THUMP domain.
| Enzyme Type | Name | Modification and Position(s) in tRNA | References |
|---|---|---|---|
| 4-thiouridine synthetase | E. coli and Methanococcus maripaludis ThiI | s4U8 and s4U9 | [12,22] |
| deaminase | Methanopyrus kandleri CDAT8 | U8 | [17] |
| methyltransferase | P. abyssi Trm11 (arcTrm11) | m2G10 and m22G10 | [13] |
| S. cerevisiae Trm11-Trm112 and Archaeoglobus fulgidus arcTrm11-arcTrm112 | m2G10 (and m22G10) | [23,24] | |
| Thermus thermophilus TrmN | m2G6 | [25] | |
| Methanocaldococcus jannaschii Trm14 | m2G6 and m2G67 | [26] | |
| Homo sapiens THUMPD3-TRM112 | m2G6 and m2G7 | [27] | |
| Partner protein of acetyltransferase | S. cerevisiae Tan1-Kre33 | ac4C12 | [16] |
| H. sapiens THUMPD1-NAT10 | ac4C12 | [16] | |
| Thermococcus kodakarensis TkTHUMDP1-TkNAT10 | ac4C (multiple positions) | [18] | |
| Pseudouridine synthase | Pyrococcus furiosus and M. jannaschii Archaeal Pus10 (arcPus10) | Ψ54 and Ψ55 | [28,29] |
| H. sapiens PUS10 | Ψ54 and Ψ55 | [30,31] |
Table 2.
THUMP-related tRNA methyltransferases.
| Enzyme Type | Organism | Subunit Composition | Modification(s) | Reference(s) |
|---|---|---|---|---|
| Trm11/arcTrm11/arcTrm11-arcTrm112/TRMT11-TRMT112 | S. cerevisiae | Trm11-Trm112 | m2G10 | [23] |
| H. sapiens | TRMT11-TRMT112 | m2G10? | [111] | |
| A. fulgidus | arcTrm11-arcTrm112 | m2G10 and m22G10 | [24] | |
| H. volcanii | arcTrm11-arcTrm112 | m2G10 and m22G10 | [112] | |
| P. abyssi | arcTrm11 | m2G10 and m22G10 | [13] | |
| T. kodakarensis | arcTrm11 | m2G10 and m22G10 | [34,113] | |
| TrmN/Trm14/THUMPD3-TRMT112 | T. thermophilus | TrmN | m2G6 | [25] |
| M. jannaschii | Trm14 | m2G6 and m2G67? | [26] | |
| T. kodakarensis | Trm14 | m2G6 and m2G67? | [113] | |
| H. sapiens | THUMPD3-TRMT112 | m2G6 and m2G7 | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hori, H. Transfer RNA Modification Enzymes with a Thiouridine Synthetase, Methyltransferase and Pseudouridine Synthase (THUMP) Domain and the Nucleosides They Produce in tRNA. Genes 2023, 14, 382. https://doi.org/10.3390/genes14020382
AMA Style
Hori H. Transfer RNA Modification Enzymes with a Thiouridine Synthetase, Methyltransferase and Pseudouridine Synthase (THUMP) Domain and the Nucleosides They Produce in tRNA. Genes. 2023; 14(2):382. https://doi.org/10.3390/genes14020382
Chicago/Turabian StyleHori, Hiroyuki. 2023. "Transfer RNA Modification Enzymes with a Thiouridine Synthetase, Methyltransferase and Pseudouridine Synthase (THUMP) Domain and the Nucleosides They Produce in tRNA" Genes 14, no. 2: 382. https://doi.org/10.3390/genes14020382
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.