
In Vitro Analysis of Biological Activity of Circulating Cell-Free DNA Isolated from Blood Plasma of Schizophrenic Patients and Healthy Controls—Part 2: Adaptive Response
 ,
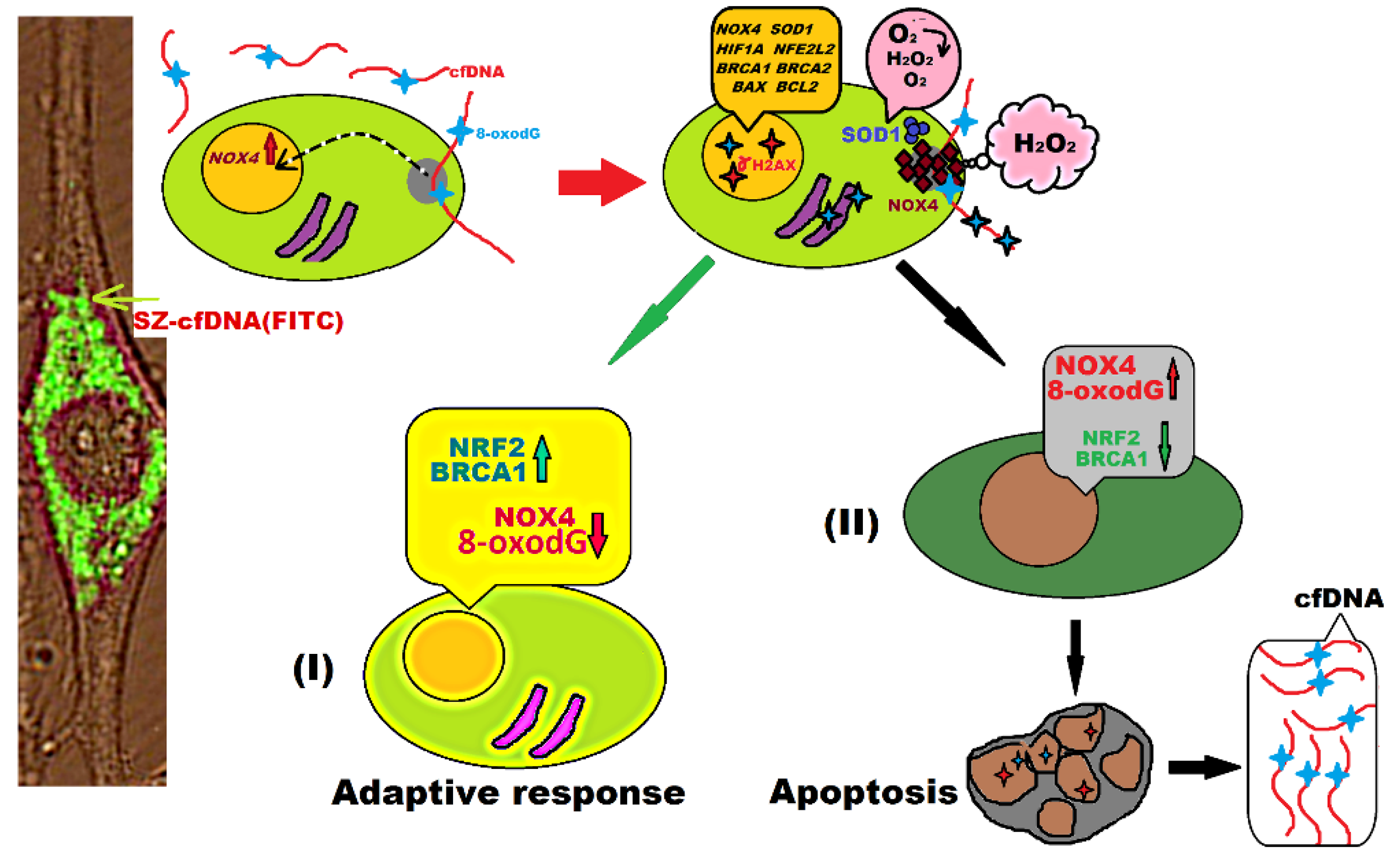
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. DNA Fragments
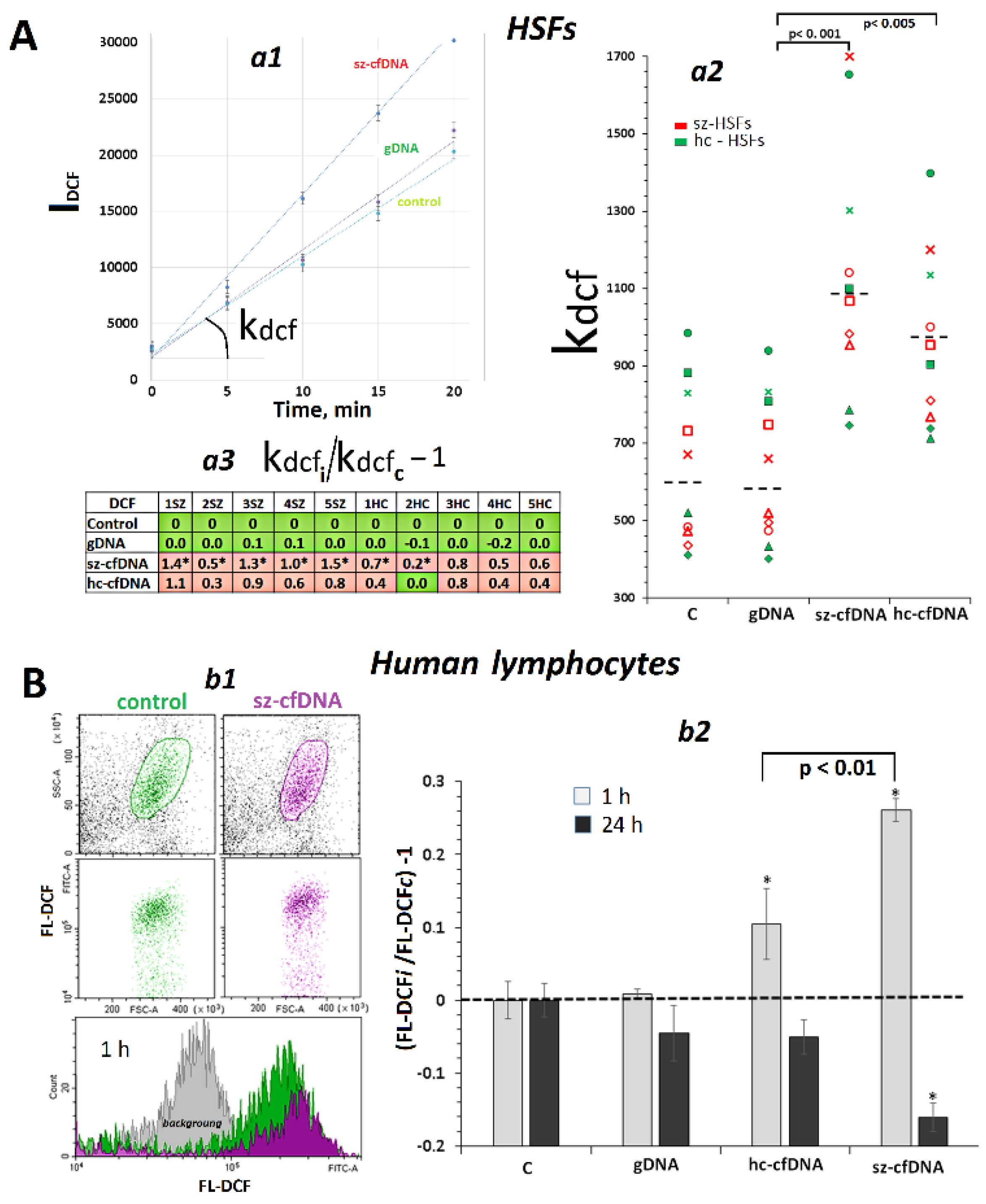
2.2. Exposure to (hc,sz)-cfDNA Induced an Increase in ROS Production
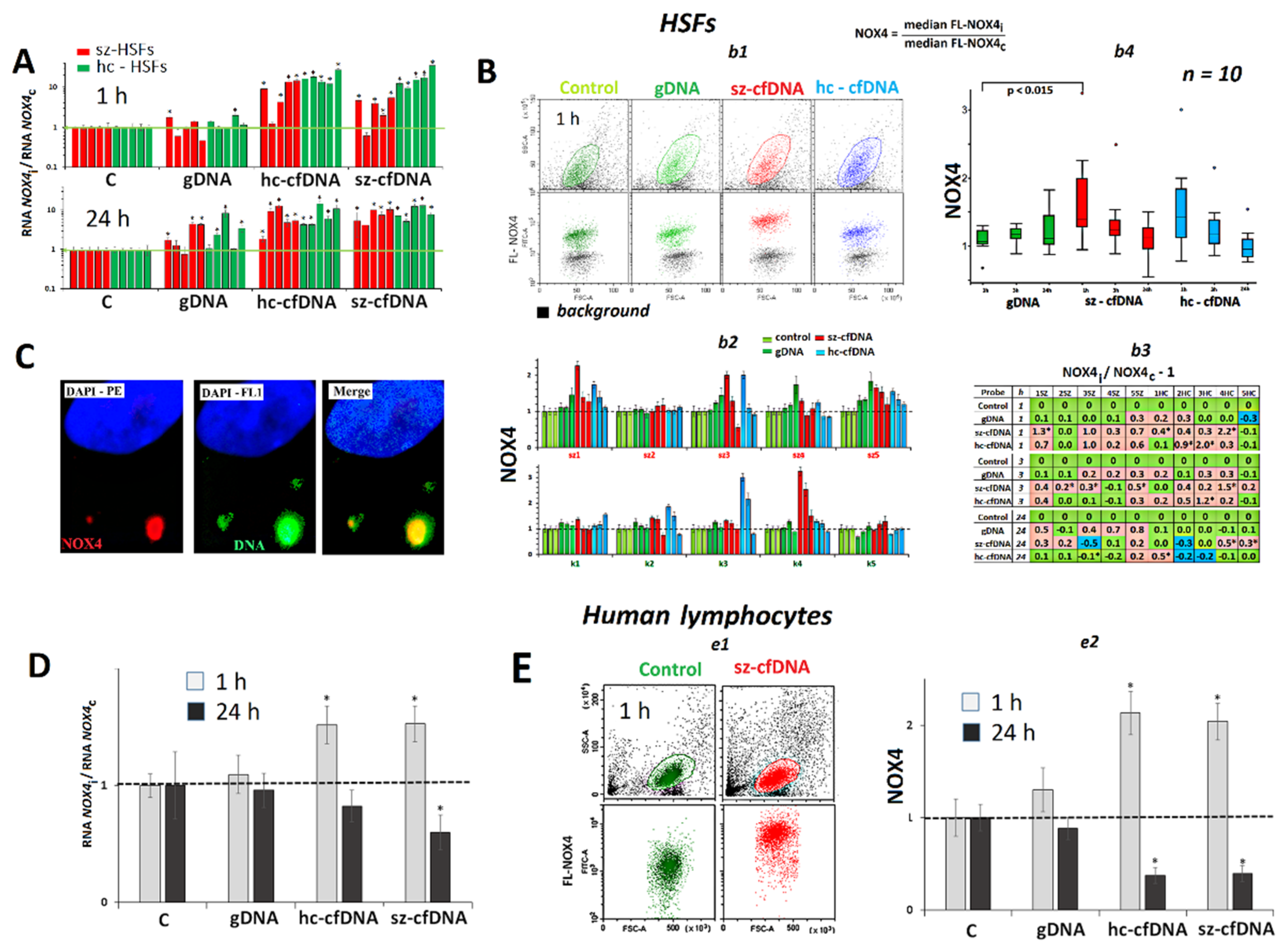
2.3. NOX4 Expression
2.3.1. In HSFs
2.3.2. In PBL
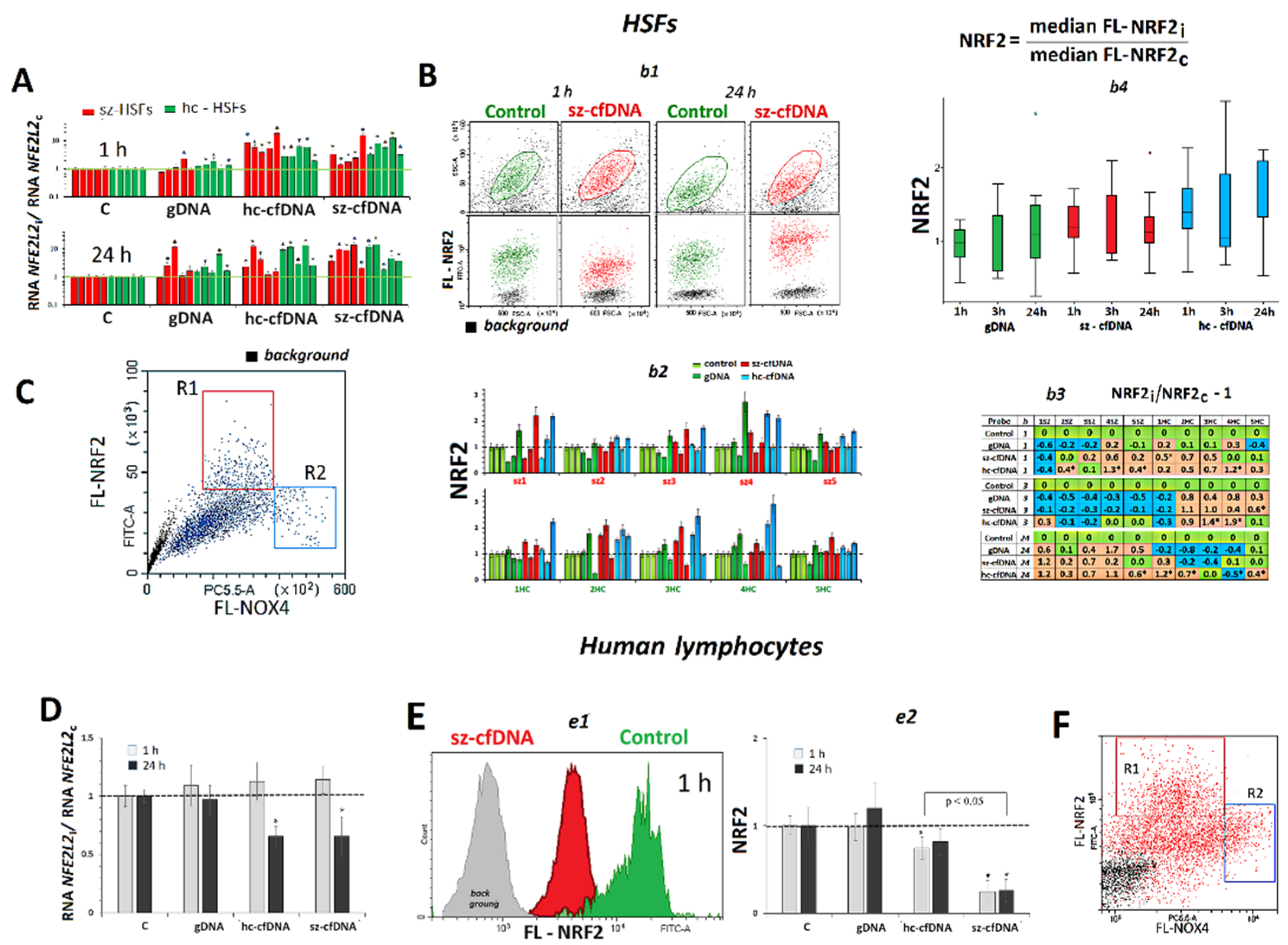
2.4. NFE2L2 Expression
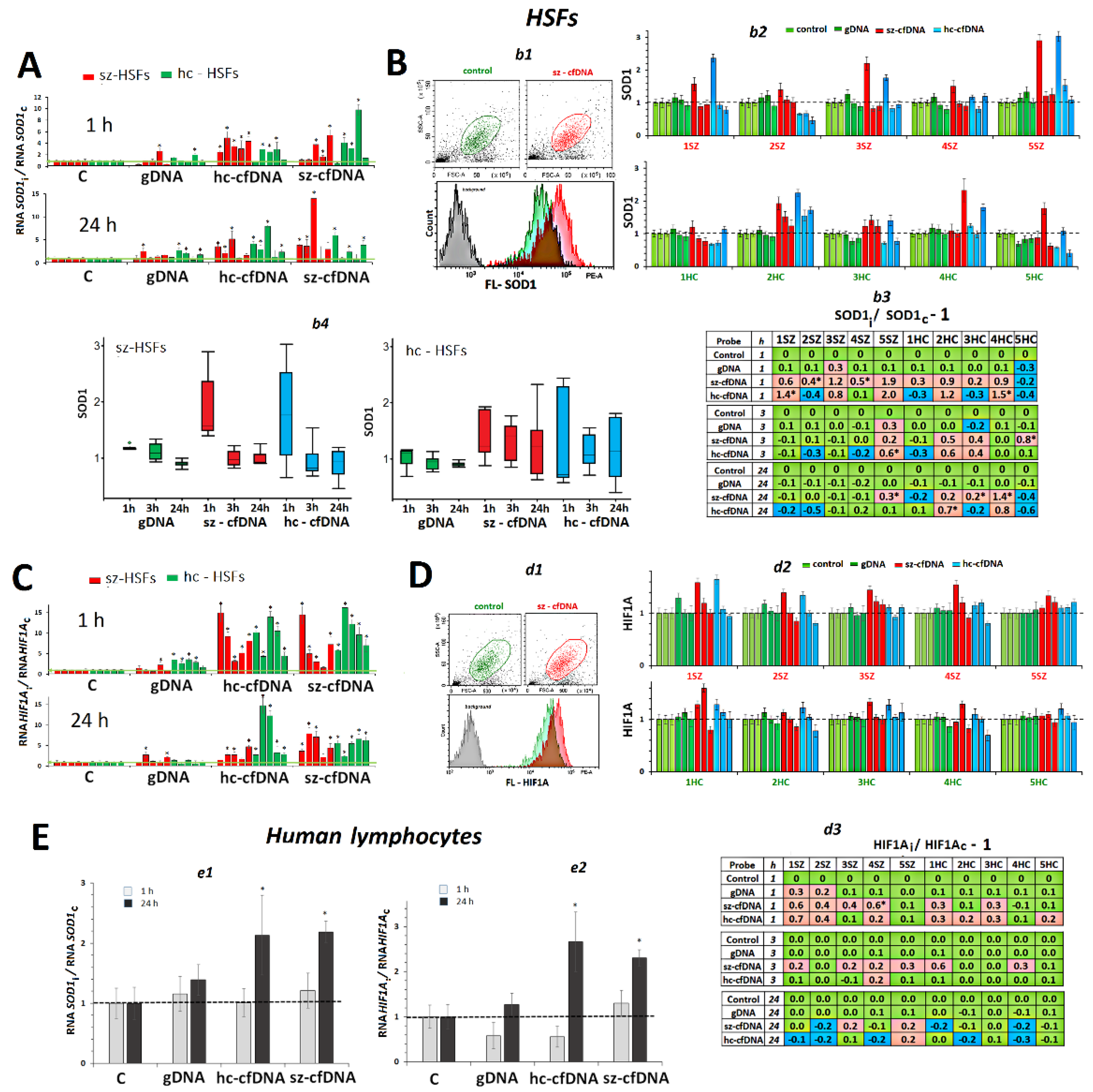
2.5. SOD1 Expression
2.6. HIF1A Expression
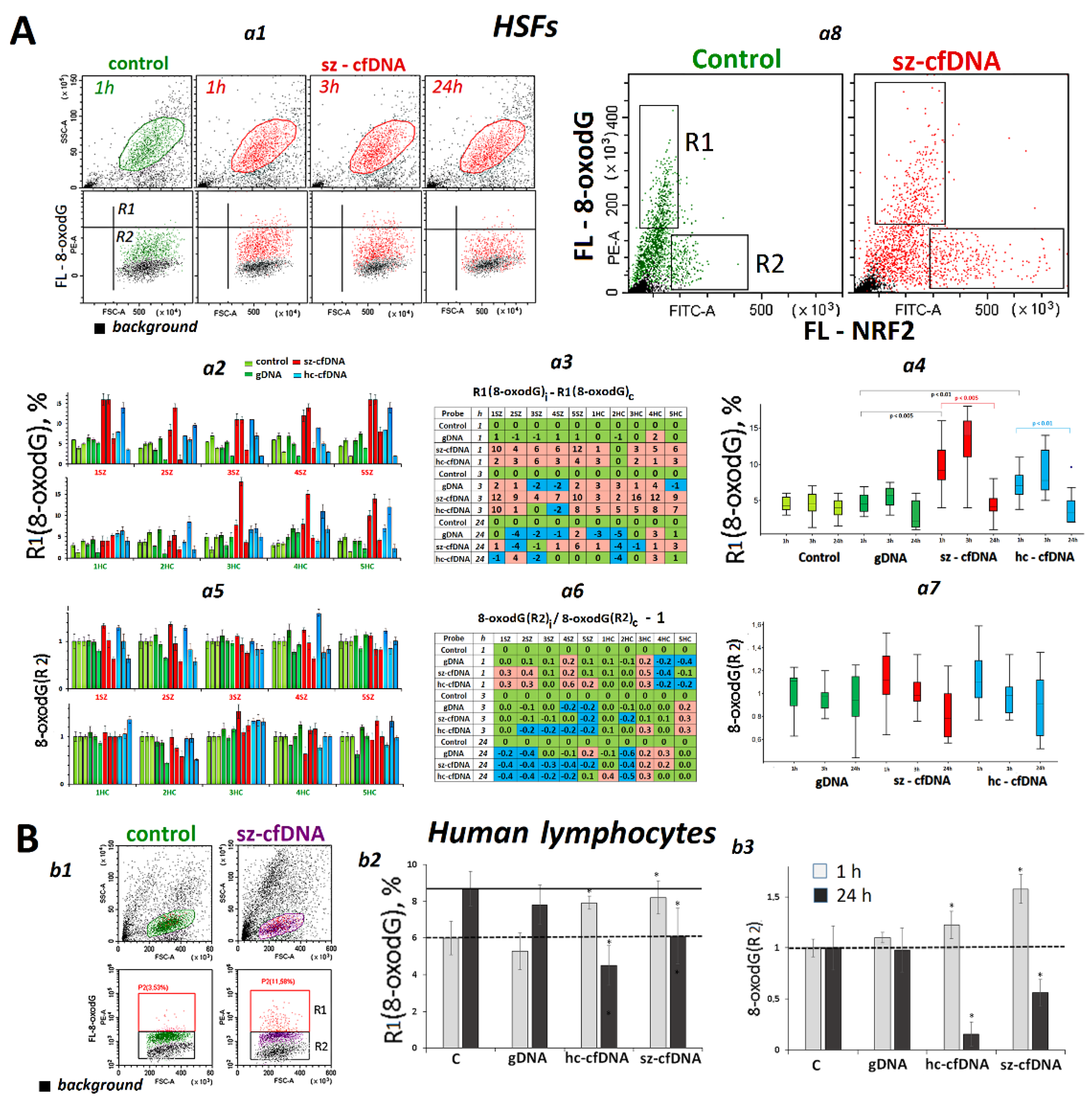
2.7. Changes in 8-OxodG Content
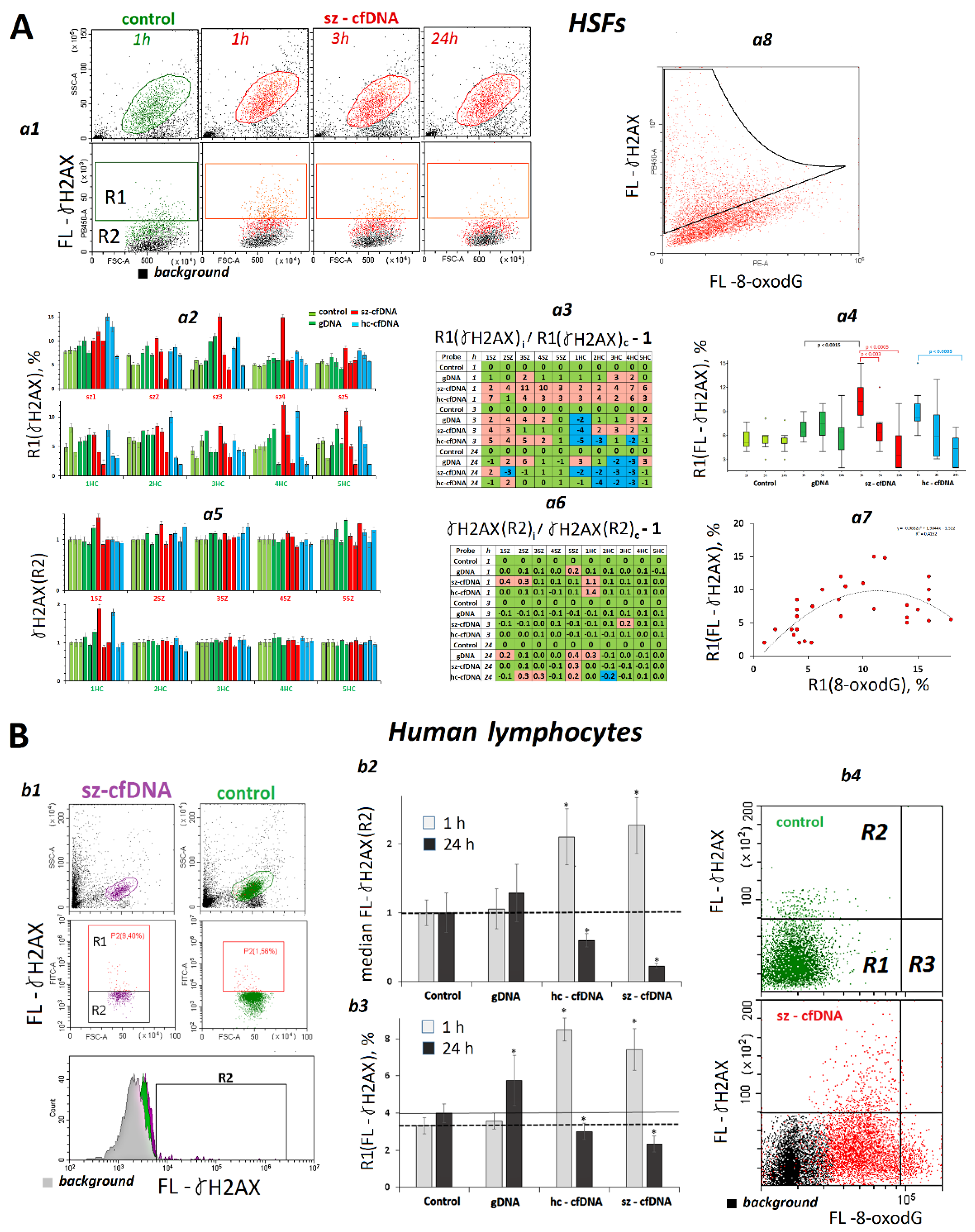
2.8. Changes in γH2AX Content
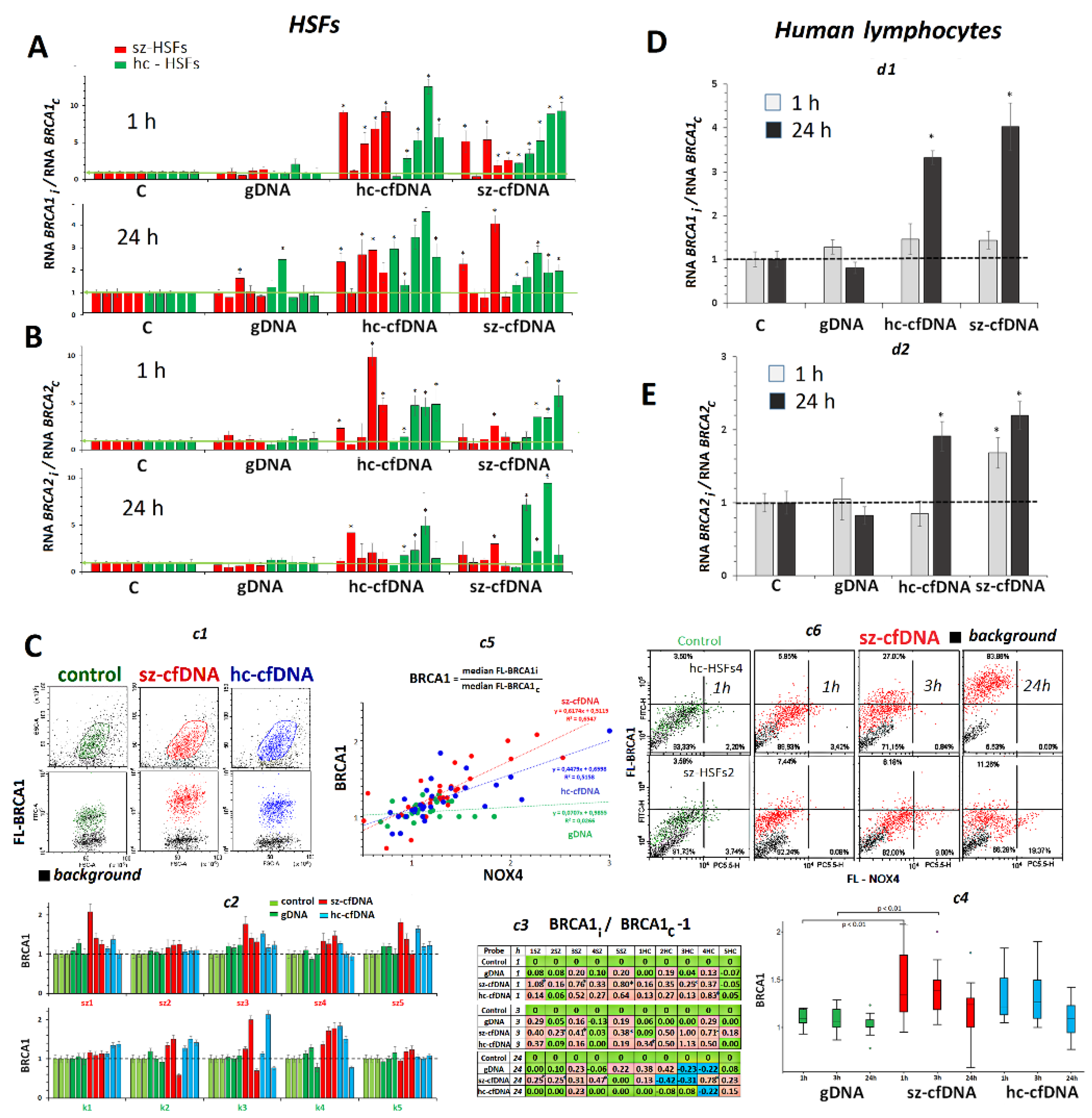
2.9. BRCA1 and BRCA2 Expression
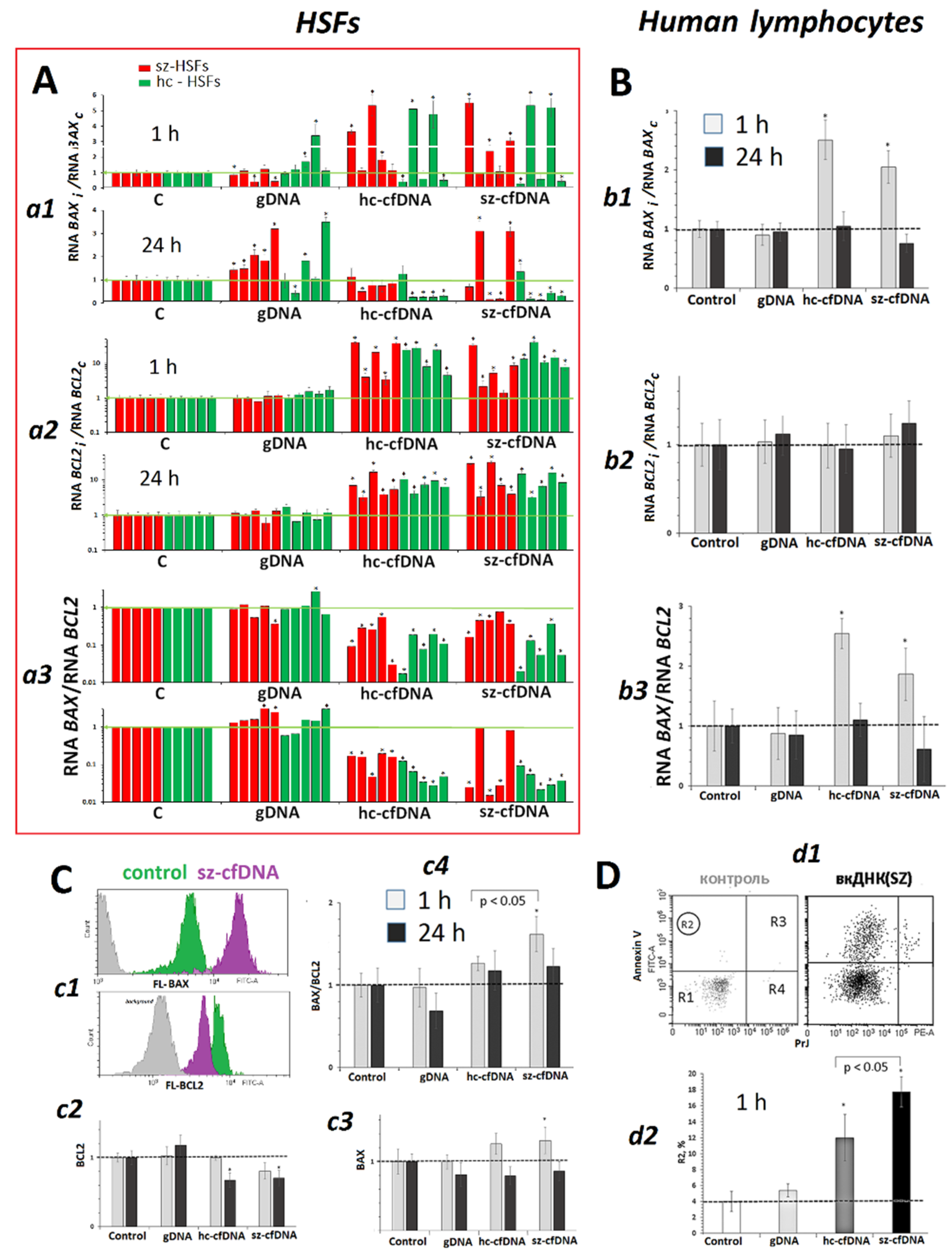
2.10. BAX and BCL2 Expression
2.11. Correlation Analysis of Changes in the ROS, DNA Damage, RNA and Protein Levels in HSFs
3. Discussion
3.1. Hc-cfDNA and sz-cfDNA Induced a Transient Oxidative Stress and DNA Damage in HSFs and PBL
3.2. Hc-cfDNA and sz-cfDNA Induced an Antioxidant Response in HSFs and PBL
3.3. Hc-cfDNA and sz-cfDNA Induced b HSFs and PBL DNA Damage Response or Apoptosis
3.4. Heterogeneity of HSFs and PBL Pools and HSF Cell Lines by the Type of Their Response to the Action of hc-cfDNA and sz-cfDNA
4. Materials and Methods
4.1. Experimental Design and Participants
4.2. Cell Cultures
4.2.1. Human Skin Fibroblasts (HSFs)
4.2.2. Peripheral Blood Lymphocyte (PBL)
4.3. Ethical Approval for Operating with Cultured and Primary (Blood Leukocytes) Human Cells
4.4. Synthesis of Fluorescently Labeled sz-cfDNA Probe
4.5. ROS Assay on HSFs
4.6. Flow Cytometry Analysis (FCA)
4.6.1. Cell Cultivation
4.6.2. Fixation and Quantification
4.6.3. PBL ROS Assay
4.7. Real-Time PCR Assay
4.8. Fluorescence Microscopy
4.9. Statistical Data Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bronkhorst, A.J.; Ungerer, V.; Oberhofer, A.; Gabriel, S.; Polatoglou, E.; Randeu, H.; Uhlig, C.; Pfister, H.; Mayer, Z.; Holdenrieder, S. New Perspectives on the Importance of Cell-Free DNA Biology. Diagnostics 2022, 2, 2147. [Google Scholar] [CrossRef] [PubMed]
- Han, D.S.C.; Lo, Y.M.D. The Nexus of cfDNA and Nuclease Biology. Trends Genet. 2021, 37, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Szilágyi, M.; Pös, O.; Márton, É.; Buglyó, G.; Soltész, B.; Keserű, J.; Penyige, A.; Szemes, T.; Nagy, B. Circulating Cell-Free Nucleic Acids: Main Characteristics and Clinical Application. Int. J. Mol. Sci. 2020, 21, 6827. [Google Scholar] [CrossRef] [PubMed]
- Kustanovich, A.; Schwartz, R.; Peretz, T.; Grinshpun, A. Life and death of circulating cell-free DNA. Cancer Biol. Ther. 2019, 20, 1057–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Chen, H.; Long, Y.; Li, P.; Gu, Y. The main sources of circulating cell-free DNA: Apoptosis, necrosis and active secretion. Crit. Rev. Oncol. Hematol. 2021, 157, 103166. [Google Scholar] [CrossRef] [PubMed]
- Rostami, A.; Lambie, M.; Yu, C.W.; Stambolic, V.; Waldron, J.N.; Bratman, S.V. Senescence, Necrosis, and Apoptosis Govern Circulating Cell-free DNA Release Kinetics. Cell Rep. 2020, 31, 107830. [Google Scholar] [CrossRef]
- Grabuschnig, S.; Bronkhorst, A.J.; Holdenrieder, S.; Rosales Rodriguez, I.; Schliep, K.P.; Schwendenwein, D.; Ungerer, V.; Sensen, C.W. Putative Origins of Cell-Free DNA in Humans: A Review of Active and Passive Nucleic Acid Release Mechanisms. Int. J. Mol. Sci. 2020, 21, 8062. [Google Scholar] [CrossRef]
- Cheng, A.P.; Cheng, M.P.; Loy, C.J.; Lenz, J.S.; Chen, K.; Smalling, S.; Burnham, P.; Timblin, K.M.; Orejas, J.L.; Silverman, E.; et al. Cell-free DNA profiling informs all major complications of hematopoietic cell transplantation. Proc. Natl. Acad. Sci. USA 2022, 119, e2113476118. [Google Scholar] [CrossRef]
- Song, P.; Wu, L.R.; Yan, Y.H.; Zhang, J.X.; Chu, T.; Kwong, L.N.; Patel, A.A.; Zhang, D.Y. Limitations and opportunities of technologies for the analysis of cell-free DNA in cancer diagnostics. Nat. Biomed. Eng. 2022, 6, 232–245. [Google Scholar] [CrossRef]
- Lannoo, L.; Lenaerts, L.; Van Den Bogaert, K.; Che, H.; Brison, N.; Devriendt, K.; Amant, F.; Vermeesch, J.R.; Van Calsteren, K. Non-invasive prenatal testing suggesting a maternal malignancy: What do we tell the prospective parents in Belgium? Prenat. Diagn. 2021, 41, 1264–1272. [Google Scholar] [CrossRef]
- Ranucci, R. Cell-Free DNA: Applications in Different Diseases. Methods Mol. Biol. 2019, 1909, 3–12. [Google Scholar] [PubMed]
- Hummel, E.M.; Hessas, E.; Müller, S.; Beiter, T.; Fisch, M.; Eibl, A.; Wolf, O.T.; Giebel, B.; Platen, P.; Kumsta, R.; et al. Cell-free DNA release under psychosocial and physical stress conditions. Transl. Psychiatry 2018, 8, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.Q.; Liang, D.Y.; Lou, X.L.; Zhang, M.; Zhang, Z.H.; Zhang, L.R. Branched DNA-based Alu quantitative assay for cell-free plasma DNA levels in patients with sepsis or systemic inflammatory response syndrome. J. Crit. Care 2016, 31, 90–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitbach, S.; Tug, S.; Simon, P. Circulating cell-free DNA: An up-coming molecular marker in exercise physiology. Sports Med. 2012, 42, 565–586. [Google Scholar] [CrossRef]
- Pinzani, P.; Salvianti, F.; Pazzagli, M.; Orlando, C. Circulating nucleic acids in cancer and pregnancy. Methods 2010, 50, 302–307. [Google Scholar] [CrossRef]
- Ershova, E.S.; Jestkova, E.M.; Chestkov, I.V.; Porokhovnik, L.N.; Izevskaya, V.L.; Kutsev, S.I.; Veiko, N.N.; Shmarina, G.; Dolgikh, O.; Kostyuk, S.V. Quantification of cell-free DNA in blood plasma and DNA damage degree in lymphocytes to evaluate dysregulation of apoptosis in schizophrenia patients. J. Psychiatr. Res. 2017, 87, 15–22. [Google Scholar] [CrossRef]
- Ershova, E.S.; Jestkova, E.M.; Martynov, A.V.; Shmarina, G.V.; Umriukhin, P.E.; Bravve, L.V.; Zakharova, N.V.; Kostyuk, G.P.; Saveliev, D.V.; Orlova, M.D.; et al. Accumulation of Circulating Cell-Free CpG-Enriched Ribosomal DNA Fragments on the Background of High Endonuclease Activity of Blood Plasma in Schizophrenic Patients. Int. J. Genom. 2019, 2019, 8390585. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Chen, X.; Sun, L.; Qing, Y.; Yang, X.; Hu, X.; Yang, C.; Xu, T.; Wang, J.; Wang, P.; et al. Analysis of the concentrations and size distributions of cell-free DNA in schizophrenia using fluorescence correlation spectroscopy. Transl. Psychiatry 2018, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Y.; Qi, J.; Xu, H.L.; Lin, X.Y.; Sun, Y.J.; Ju, S.Q. The Value of Serum Cell-Free DNA Levels in Patients with Schizophrenia. Front. Psychiatry 2021, 12, 637789. [Google Scholar] [CrossRef]
- Deng, J.; Pan, W.; Ji, N.; Liu, N.; Chen, Q.; Chen, J.; Sun, Y.; Xie, L.; Chen, Q. Cell-Free DNA Promotes Inflammation in Patients with Oral Lichen Planus via he STING Pathway. Front. Immunol. 2022, 13, 838109. [Google Scholar] [CrossRef]
- Korabecna, M.; Zinkova, A.; Brynychova, I.; Chylikova, B.; Prikryl, P.; Sedova, L.; Neuzil, P.; Seda, O. Cell-free DNA in plasma as an essential immune system regulator. Sci. Rep. 2020, 10, 17478. [Google Scholar] [CrossRef] [PubMed]
- Shmarina, G.V.; Ershova, E.S.; Simashkova, N.V.; Nikitina, S.G.; Chudakova, J.M.; Veiko, N.N.; Porokhovnik, L.N.; Basova, A.Y.; Shaposhnikova, A.F.; Pukhalskaya, D.A.; et al. Oxidized cell-free DNA as a stress-signaling factor activating the chronic inflammatory process in patients with autism spectrum disorders. J. Neuroinflamm. 2020, 17, 212. [Google Scholar] [CrossRef] [PubMed]
- Anunobi, R.; Boone, B.A.; Cheh, N.; Tang, D.; Kang, R.; Loux, T.; Lotze, M.T.; Zeh, H.J. Extracellular DNA promotes colorectal tumor cell survival after cytotoxic chemotherapy. J. Surg. Res. 2018, 226, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Waldvogel Abramowski, S.; Tirefort, D.; Lau, P.; Guichebaron, A.; Taleb, S.; Modoux, C.; Lemoine Chaduc, C.; Bruyere Cerdan, P.; Roux Lombard, P.; Lecompte, T.; et al. Cell-free nucleic acids are present in blood products and regulate genes of innate immune response. Transfusion 2018, 58, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Zhang, W.; Wang, X.; Cai, P.; Jia, Q.; Zhao, W. Cell-free DNA induced apoptosis of granulosa cells by oxidative stress. Clin. Chim. Acta 2017, 473, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Műzes, G.; Kiss, A.L.; Tulassay, Z.; Sipos, F. Cell-free DNA-induced alteration of autophagy response and TLR9-signaling: Their relation to amelioration of DSS-colitis. Comp. Immunol. Microbiol. Infect. Dis. 2017, 52, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Gravina, S.; Sedivy, J.M.; Vijg, J. The dark side of circulating nucleic acids. Aging Cell 2016, 15, 398–399. [Google Scholar] [CrossRef] [Green Version]
- Basak, R.; Nair, N.K.; Mittra, I. Evidence for cell-free nucleic acids as continuously arising endogenous DNA mutagens. Mutat. Res. 2016, 793–794, 15–21. [Google Scholar] [CrossRef]
- Ershova, E.S.; Shmarina, G.V.; Porokhovnik, L.N.; Zakharova, N.V.; Kostyuk, G.P.; Umriukhin, P.E.; Kutsev, S.I.; Sergeeva, V.A.; Veiko, N.N.; Kostyuk, S.V. In Vitro Analysis of Biological Activity of Circulating Cell-Free DNA Isolated from Blood Plasma of Schizophrenic Patients and Healthy Controls. Genes 2022, 13, 551. [Google Scholar] [CrossRef]
- Ermakov, A.V.; Konkova, M.S.; Kostyuk, S.V.; Egolina, N.A.; Efremova, L.V.; Veiko, N.N. Oxidative stress as a significant factor for development of an adaptive response in irradiated and nonirradiated human lymphocytes after inducing the bystander effect by low-dose X-radiation. Mutat. Res. 2009, 669, 155–161. [Google Scholar] [CrossRef]
- Kostyuk, S.V.; Ermakov, A.V.; Alekseeva, A.Y.; Smirnova, T.D.; Glebova, K.V.; Efremova, L.V.; Baranova, A.; Veiko, N.N. Role of extracellular DNA oxidative modification in radiation induced bystander effects in human endotheliocytes. Mutat. Res. 2012, 729, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, S.V.; Tabakov, V.J.; Chestkov, V.V.; Konkova, M.S.; Glebova, K.V.; Baydakova, G.V.; Ershova, E.S.; Izhevskaya, V.L.; Baranova, A.; Veiko, N.N. Oxidized DNA induces an adaptive response in human fibroblasts. Mutat. Res. 2013, 747–748, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, S.V.; Konkova, M.S.; Ershova, E.S.; Alekseeva, A.Y.; Smirnova, T.D.; Stukalov, S.V.; Kozhina, E.A.; Shilova, N.V.; Zolotukhina, T.V.; Markova, Z.G.; et al. An exposure to the oxidized DNA enhances both instability of genome and survival in cancer cells. PLoS ONE 2013, 8, e77469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostyuk, S.; Smirnova, T.; Kameneva, L.; Porokhovnik, L.; Speranskij, A.; Ershova, E.; Stukalov, S.; Izevskaya, V.; Veiko, N. GC-Rich Extracellular DNA Induces Oxidative Stress, Double-Strand DNA Breaks, and DNA Damage Response in Human Adipose-Derived Mesenchymal Stem Cells. Oxidative Med. Cell. Longev. 2015, 2015, 782123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergeeva, V.A.; Ershova, E.S.; Veiko, N.N.; Malinovskaya, E.M.; Kalyanov, A.A.; Kameneva, L.V.; Stukalov, S.V.; Dolgikh, O.A.; Konkova, M.S.; Ermakov, A.V.; et al. Low-Dose Ionizing Radiation Affects Mesenchymal Stem Cells via Extracellular Oxidized Cell-Free DNA: A Possible Mediator of Bystander Effect and Adaptive Response. Oxidative Med. Cell. Longev. 2017, 2017, 9515809. [Google Scholar] [CrossRef] [Green Version]
- Filev, A.D.; Shmarina, G.V.; Ershova, E.S.; Veiko, N.N.; Martynov, A.V.; Borzikova, M.A.; Poletkina, A.A.; Dolgikh, O.A.; Veiko, V.P.; Bekker, A.A.; et al. Oxidized Cell-Free DNA Role in the Antioxidant Defense Mechanisms under Stress. Oxidative Med. Cell. Longev. 2019, 2019, 1245749. [Google Scholar] [CrossRef]
- Kozhina, E.A.; Ershova, E.S.; Okorokova, N.A.; Veiko, V.P.; Malinovskaya, E.M.; Sergeeva, V.A.; Konkova, M.S.; Kutsev, S.I.; Veiko, N.N.; Kostyuk, S.V. Extracellular DNA Containing (dG)n Motifs Penetrates into MCF7 Breast Cancer Cells, Induces the Adaptive Response, and Can Be Expressed. Oxidative Med. Cell. Longev. 2019, 2019, 7853492. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Chen, X. The human Nox4: Gene, structure, physiological function and pathological significance. J. Drug Target 2015, 23, 888–896. [Google Scholar] [CrossRef]
- Ershova, E.S.; Shmarina, G.V.; Martynov, A.V.; Zakharova, N.V.; Veiko, R.V.; Umriukhin, P.E.; Kostyuk, G.P.; Kutsev, S.I.; Veiko, N.N.; Kostyuk, S.V. NADPH-oxidase 4 gene over-expression in peripheral blood lymphocytes of the schizophrenia patients. PLoS ONE 2022, 17, e0269130. [Google Scholar] [CrossRef]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by Mitochondrial Reactive Oxygen Species in Physiology and Pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef]
- Mir, S.; Golden, B.D.O.; Griess, B.J.; Vengoji, R.; Tom, E.; Kosmacek, E.A.; Oberley-Deegan, R.E.; Talmon, G.A.; Band, V.; Teoh-Fitzgerald, M.L. Upregulation of Nox4 induces a pro-survival Nrf2 response in cancer-associated fibroblasts that promotes tumorigenesis and metastasis, in part via Birc5 induction. Breast Cancer Res. 2022, 24, 48. [Google Scholar] [CrossRef] [PubMed]
- Milani, P.; Ambrosi, G.; Gammoh, O.; Blandini, F.; Cereda, C. SOD1 and DJ-1 converge at Nrf2 pathway: A clue for antioxidant therapeutic potential in neurodegeneration. Oxidative Med. Cell. Longev. 2013, 2013, 836760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwa, J.; Yamada, S.; Ishigaki, S.; Sone, J.; Takahashi, M.; Katsuno, M.; Tanaka, F.; Doyu, M.; Sobue, G. Disulfide bond mediates aggregation, toxicity, and ubiquitylation of familial amyotrophic lateral sclerosis-linked mutant SOD1. J. Biol. Chem. 2007, 282, 28087–28095. [Google Scholar] [CrossRef] [Green Version]
- Bi, Z.; Zhang, Q.; Fu, Y.; Wadgaonkar, P.; Zhang, W.; Almutairy, B.; Xu, L.; Rice, M.; Qiu, Y.; Thakur, C.; et al. Nrf2 and HIF1α converge to arsenic-induced metabolic reprogramming and the formation of the cancer stem-like cells. Theranostics 2020, 10, 4134–4149. [Google Scholar] [CrossRef]
- Yu, H.; Chen, B.; Ren, Q. Baicalin relieves hypoxia-aroused H9c2 cell apoptosis by activating Nrf2/HO-1-mediated HIF1α/BNIP3 pathway. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3657–3663, Erratum in Artif. Cells Nanomed. Biotechnol. 2021, 49, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Gregorio, J.; Cilenti, L.; Ambivero, C.T.; Andl, T.; Liao, R.; Zervos, A.S. UBXN7 cofactor of CRL3KEAP1 and CRL2VHL ubiquitin ligase complexes mediates reciprocal regulation of NRF2 and HIF-1α proteins. Biochim. Biophys. Acta Mol. Cell. Res. 2021, 1868, 118963. [Google Scholar] [CrossRef] [PubMed]
- Pilger, A.; Rüdiger, H.W. 8-Hydroxy-2′-deoxyguanosine as a marker of oxidative DNA damage related to occupational and environmental exposures. Int. Arch. Occup. Environ. Health 2006, 80, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, M.K.; Ames, B.N. Assays for 8-hydroxy-2′-deoxyguanosine: A biomarker of in vivo oxidative DNA damage. Free Radic. Biol. Med. 1991, 10, 211–216. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Giunta, S.; Fenech, M.; Neri, M.; Bonassi, S. γH2AX as a marker of DNA double strand breaks and genomic instability in human population studies. Mutat. Res. 2013, 753, 24–40. [Google Scholar] [CrossRef]
- Duncan, J.A.; Reeves, J.R.; Cooke, T.G. BRCA1 and BRCA2 proteins: Roles in health and disease. Mol. Pathol. 1998, 51, 237–247. [Google Scholar] [CrossRef]
- Yoshida, K.; Miki, Y. Role of BRCA1 and BRCA2 as regulators of DNA repair, transcription, and cell cycle in response to DNA damage. Cancer Sci. 2004, 95, 866–871. [Google Scholar] [CrossRef]
- Hardwick, J.M.; Soane, L. Multiple functions of BCL-2 family proteins. Cold Spring Harb. Perspect. Biol. 2013, 5, a008722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltvai, Z.N.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell 1993, 74, 609–619. [Google Scholar] [CrossRef]
- Korsmeyer, S.J.; Shutter, J.R.; Veis, D.J.; Merry, D.E.; Oltvai, Z.N. Bcl-2/Bax: A rheostat that regulates an anti-oxidant pathway and cell death. Semin. Cancer Biol. 1993, 4, 327–332. [Google Scholar]
- Porokhovnik, L.N.; Passekov, V.P.; Gorbachevskaya, N.L.; Sorokin, A.B.; Veiko, N.N.; Lyapunova, N.A. Active ribosomal genes, translational homeostasis and oxidative stress in the pathogenesis of schizophrenia and autism. Psychiatr. Genet. 2015, 25, 79–87. [Google Scholar] [CrossRef]
- Alekseeva, A.Y.; Kameneva, L.V.; Kostyuk, S.V.; Veiko, N.N. Multiple Ways of cfDNA Reception and Following ROS Production in Endothelial Cells. Adv. Exp. Med. Biol. 2016, 924, 127–131. [Google Scholar] [PubMed]
- Kostyuk, S.V.; Mordkovich, N.N.; Okorokova, N.A.; Veiko, V.P.; Malinovskaya, E.M.; Ershova, E.S.; Konkova, M.S.; Savinova, E.A.; Borzikova, M.A.; Muzaffarova, T.A.; et al. Increased Transfection of the Easily Oxidizable GC-Rich DNA Fragments into the MCF7 Breast Cancer Cell. Oxidative Med. Cell. Longev. 2019, 2019, 2348165. [Google Scholar] [CrossRef] [Green Version]
- Malinovskaya, E.M.; Ershova, E.S.; Okorokova, N.A.; Veiko, V.P.; Konkova, M.S.; Kozhina, E.A.; Savinova, E.A.; Porokhovnik, L.N.; Kutsev, S.I.; Veiko, N.N.; et al. Ribosomal DNA as DAMPs Signal for MCF7 Cancer Cells. Front. Oncol. 2019, 9, 445. [Google Scholar] [CrossRef] [Green Version]
- Tamkovich, S.; Laktionov, P. Cell-surface-bound circulating DNA in the blood: Biology and clinical application. IUBMB Life 2019, 71, 1201–1210. [Google Scholar] [CrossRef]
- Chestkov, I.V.; Jestkova, E.M.; Ershova, E.S.; Golimbet, V.G.; Lezheiko, T.V.; Kolesina, N.Y.; Dolgikh, O.A.; Izhevskaya, V.L.; Kostyuk, G.P.; Kutsev, S.I.; et al. ROS-Induced DNA Damage Associates with Abundance of Mitochondrial DNA in White Blood Cells of the Untreated Schizophrenic Patients. Oxidative Med. Cell. Longev. 2018, 2018, 8587475. [Google Scholar] [CrossRef] [Green Version]
- Lacher, S.E.; Levings, D.C.; Freeman, S.; Slattery, M. Identification of a functional antioxidant response element at the HIF1A locus. Redox Biol. 2018, 19, 401–411. [Google Scholar] [CrossRef]
- Konkova, M.; Abramova, M.; Kalianov, A.; Ershova, E.; Dolgikh, O.; Umriukhin, P.; Izhevskaya, V.; Kutsev, S.; Veiko, N.; Kostyuk, S. Mesenchymal Stem Cells Early Response to Low-Dose Ionizing Radiation. Front. Cell Dev. Biol. 2020, 8, 584497. [Google Scholar] [CrossRef]
- Bulicheva, N.; Fidelina, O.; Mkrtumova, N.; Neverova, M.; Bogush, A.; Bogush, M.; Roginko, O.; Veiko, N. Effect of cell-free DNA of patients with cardiomyopathy and rDNA on the frequency of contraction of electrically paced neonatal rat ventricular myocytes in culture. Ann. N. Y. Acad. Sci. 2008, 1137, 273–277. [Google Scholar] [CrossRef]
- Efremova, L.V.; Alekseeva, A.Y.; Konkova, M.S.; Kostyuk, S.V.; Ershova, E.S.; Smirnova, T.D.; Konorova, I.L.; Veiko, N.N. Extracellular DNA affects NO content in human endothelial cells. Bull. Exp. Biol. Med. 2010, 149, 196–200. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostyuk, S.V.; Ershova, E.S.; Martynov, A.V.; Artyushin, A.V.; Porokhovnik, L.N.; Malinovskaya, E.M.; Jestkova, E.M.; Zakharova, N.V.; Kostyuk, G.P.; Izhevskaia, V.L.; et al. In Vitro Analysis of Biological Activity of Circulating Cell-Free DNA Isolated from Blood Plasma of Schizophrenic Patients and Healthy Controls—Part 2: Adaptive Response. Genes 2022, 13, 2283. https://doi.org/10.3390/genes13122283
Kostyuk SV, Ershova ES, Martynov AV, Artyushin AV, Porokhovnik LN, Malinovskaya EM, Jestkova EM, Zakharova NV, Kostyuk GP, Izhevskaia VL, et al. In Vitro Analysis of Biological Activity of Circulating Cell-Free DNA Isolated from Blood Plasma of Schizophrenic Patients and Healthy Controls—Part 2: Adaptive Response. Genes. 2022; 13(12):2283. https://doi.org/10.3390/genes13122283
Chicago/Turabian StyleKostyuk, Svetlana V., Elizaveta S. Ershova, Andrey V. Martynov, Andrey V. Artyushin, Lev N. Porokhovnik, Elena M. Malinovskaya, Elizaveta M. Jestkova, Natalia V. Zakharova, George P. Kostyuk, Vera L. Izhevskaia, and et al. 2022. "In Vitro Analysis of Biological Activity of Circulating Cell-Free DNA Isolated from Blood Plasma of Schizophrenic Patients and Healthy Controls—Part 2: Adaptive Response" Genes 13, no. 12: 2283. https://doi.org/10.3390/genes13122283