The Sex-Specific Splicing of Doublesex in Brine Shrimp Artemia franciscana
by
 , , and
, , and
Dung Nguyen Viet
1,2, 
Olivier Christiaens
3,
Stephanie De Vos
2,
Guy Smagghe
3 and
Peter Bossier
2,* 1
Research Institute for Aquaculture No. 2, 116 Nguyen Dinh Chieu Street, Dakao Ward, District 1, Ho Chi Minh City 71007, Vietnam
2
Laboratory of Aquaculture & Artemia Reference Center, Ghent University, Coupure Links 653, 9000 Ghent, Belgium
3
Laboratory of Agrozoology, Department of Plants and Crops, Faculty of Bioscience Engineering, Ghent University, Coupure Links 653, 9000 Ghent, Belgium
*
Author to whom correspondence should be addressed.
Genes 2022, 13(11), 1997; https://doi.org/10.3390/genes13111997
Submission received: 25 August 2022
/
Revised: 18 September 2022
/
Accepted: 24 October 2022
/
Published: 1 November 2022
(This article belongs to the Section Animal Genetics and Genomics)
Abstract
:The understanding of sex determination and differentiation in animals has recently made remarkable strides through the use of advanced research tools. At the gene level, the Mab-3-related transcription factor (Dmrt) gene family, which encodes for the typical DNA-binding doublesex/Mab-3 (DM) domain in their protein, is known for its contribution to sex determination and differentiation in insects. In this study, DNA-binding DM domain screening has identified eight transcripts from Artemia franciscana transcriptomic that encode proteins containing one conserved DNA-binding DM domain. The genome mapping confirmed that these eight transcripts are transcribed from six different loci on the A. franciscana genome assembly. One of those loci, the Af.dsx-4 locus, is closely related to Doublesex, a gene belonging to the Dmrt gene family. This locus could be transcribed into three alternative transcripts, namely Af.dsx4, Af.dsxF and Af.dsxM. While Af.dsx4 and Af.dsxF could putatively be translated to form an identical Af.dsxF protein of 186 aa long, Af.dsxM translates for an Af.dsxM protein of 289 aa long but shares a DNA-binding DM domain. Interestingly, Af.dsxF and Af.dsxM are confirmed as sex-specific transcripts, Af.dsxF is only present in females, and Af.dsxM is only present in male individuals. The results suggest that the sex-specific splicing mechanism of the doublesex described in insects is also present in A. franciscana. Af.dxs-4 locus can be used in further studies to clarify the sex determination pathways in A. fracnciscana.
1. Introduction
The Artemia genus contains species with sexual as well as parthenogenetic reproduction modes. Artemia has a short life cycle, and is cultured easily under laboratory conditions. This species is being used as an animal model to discover genes involved in biological processes. Artemia has a WZ-ZZ sex-determining system which means that males are homogametic (ZZ), while females are heterogametic (ZW) [1,2] such as other crustacean species including Cherax quadricarinatus [3], Penaeus monodon [4], Macrobrachium rosenbergii [5] and Eriocheir sinensis [6]. In an effort to explore the molecular mechanism of sex determination in Artemia, a few molecular biology techniques have been employed to understand this mechanism. The Z-chromosome of Artemia. franciscana has recently been characterized by detecting genomic regions that show lower genomic coverage in female than in male samples, and regions that harbor an excess of female-specific SNPs [7]. Eight sex-linked AFLP marker alleles have been reported in A. franciscana that are inherited from the female parent [1]. Some genes related to sexual differentiation in A. franciscana have been identified and shown as sex-biased gene expression [8]. Masculinizer gene (Ar-Masc) was identified in A. franciscana [9], the MASC protein encoded in the Z chromosome activates a sex-determining gene that regulates gene splicing of Doublesex (dsx) into the male-specific isoform in the silkworm Bombyx mori [10]. Until now, the sex-determining genes in A. franciscana have not yet been identified and no genetic sex-specific marker is reported.
Doublesex and Mab-3-related transcription factor (Dmrt) genes family are reported to be very highly conserved, as well as essential for sexual development in mammal, fish, and insects. In Dmrt genes family, Dmrt1, Dmrt7, and Dmrt8 are only found in vertebrate; dmrt2a/2b, dmrt3, dmrt4/5 and dmrt93B are commonly present in invertebrate bilateria; whereas Dmrt1 and dsx have suggested independent evolution for sex determination and differentiation in the Dmrt gene family [11]. Doublesex bearing a doublesex/Mab-3 (DM) domain, regulates the sexual differentiation in Drosophila melanogaster, was the first Dmrt family member to be discovered [12]. Dsx and another two Dmrt genes including Dmrt99B and Dmrt11E could only be found in arthropod. Dsx gene is a critical transcription factor considered to be at the end of the sex determination cascade in many other insect species as a master gene in genotypic sex determination pathway. Dmrt genes family are involved in the formation of sex-linked traits including sexual behavior, gonadal development and sex-specific morphology in many insects [13]. In D. melanogaster, dsx gene is transcribed to produce a common primary transcript that is alternatively spliced and polyadenylated to yield male (dsxM) and female (dsxF) specific transcript [12]. Dsx and Dmrt have been characterized in a few crustacean species including M. rosenbergii [14,15,16,17], M. nipponense [18], E. sinensis [19], F. chinensis [20], Litopenaeus. vannamei [21], and four Daphnia species [22]. In F. chinensis, Dsx exhibited a sex-biased expression pattern in different tissues and its expression level increased along with developmental stages. Its binding site was identified on the promoter region of insulin-like androgenic gland gene [20]. Two dsx genes have been identified in D. magna (DapmaDsx1 and DapmaDsx2), which are expressed in males but not in females. DapmaDsx1 has been reported as a sex determination gene which is responsible for the male trait development [22]. Recently, Sagmariasus verreauxi, a heterogametic sex-linked iDMY/iDmrt1 has been recently reported, in which the dominant negative suppression of iDMY over its autosomal iDmrt1 paralogue might play an important key in the mechanism of determines sex this species [23]. These findings suggest the Doublesex and Mab-3-related transcription factor genes is good clues to look for the sex-determining genes in Artemia. In this study, the Dsx/Dmrt gene family is a target candidate for studying the sex determination mechanism in A. franciscana. We are looking for sex-specific splicing of Dsx/Dmrt gene family in the transcriptome database of A. franciscana. The expression profile of those genes was evaluated in both males and females using RT-PCR.
2. Materials and Methods
2.1. Animal Culture
A. franciscana cysts (Great Salt Lake, UT, USA) were hydrated in autoclaved seawater with strong aeration at 28 °C until hatching (approximately 20–24 h). The nauplii (larvae) were then transferred to a 5 L plastic tank with autoclaved seawater for rearing. The larvae were cultured at 28 °C in autoclaved seawater with aeration under fluorescent light and were fed with live marine microalgae Tetraselmis suecica. At 15–20 DAH (days after hatching), the juvenile females (before egg-sac formation) were identified by their brown-red color. At the juvenile stage, 20 individuals (10 males and 10 females) were analyzed separately to confirm sex-specific transcripts by RT-PCR. Adult males and females are morphologically completely different 20–25 DAH with the hooked grasper phenotype in males and the egg-sac in females. At the adult stage, pooled samples of the whole body of 10 adult females or 10 adult males were used to confirm sex-specific transcripts by RT-PCR.
2.2. Total RNA Extraction and cDNA Synthesis
The total RNA of the samples (the hydrated cysts, nauplii, juveniles and adults) was prepared using the RNeasy mini kit (Qiagen, Hilden, Germany). The total RNA was used to synthesize cDNA in a 20 μL reaction by using poly-T primer and RevertAid H Minus First Strand cDNA Synthesis Kit (Thermal Scientific, Waltham, MA, USA). The 20 μL of reaction mixture contained 4 μL of 5X reaction buffer, 2 μL of 10 μM dNTPs mix, 20 units of ribonuclease inhibitor, 200 units of RevertAid H Minus M-MuLV Reverse Transcriptase, 1 μL of oligo-dT primer and 500 ng of total RNA. Subsequently, the reaction mixture was incubated for 70 min at 42 °C. The reaction was terminated by heating at 70 °C for 5 min and then cooled to 4 °C. Complementary deoxyribonucleic acids (cDNA) were then used as a template in PCR for further steps.
2.3. Identification of Putative of Dsx/Dmrt Transcripts
The primary amino acid sequences of doublesex from Daphnia magna were used for BLAST against the A. franciscana transcriptome database of the Laboratory of Aquaculture & Artemia Reference Center, Ghent University, Ghent, Belgium [24]. The identity of the selected hit contigs was subsequently verified using the BLASTX algorithm with default parameters on the nucleotide database of the National Center for Biotechnology Information (NCBI) [25]. Contigs matching Dsx/Dmrt gene family were selected for coding DNA sequence (CDS) analysis. The amino acid sequence of the selected contigs and open reading frame (ORF) was determined by the translation tool from the website Expasy (http://web.expasy.org/translate accessed on 26 January 2017). For contigs containing a full coding sequence (CDS), PCR primers were designed for isolation and sequencing. In contrast, for contigs containing only a partial CDS, their respective full-length cDNAs were identified by 3′ and 5′ rapid amplification of cDNA ends (RACE).
2.4. Isolation Full-Length of Dsx/Dmrt Transcript
Based on contigs containing only a partial CDS, gene-specific primers (Table 1) were designed to isolate the UTR (UnTranslated Region) using the SMARTer RACE Kit (Clontech, Palo Alto, CA, USA) with a modified protocol. The cDNA of each transcript from different lifecycle stages was used as a template for the PCR reaction to increase the chance of transcript amplification. Briefly, the 20 µL cDNA reaction for the isolation of the 5′UTR contained 4 µL 5X First-Strand Buffer, 0.5 µL DTT (100 mM), 1 µL dNTPs (20 mM) 5′-CDS Primer A, 10 μL total RNA, 1 µL of the SMARTer II A Oligonucleotide, 0.5 µL RNase Inhibitor (40 U/µL) and 2 µL SMARTScribe Reverse Transcriptase (100 U). The mixture was incubated at 42 °C for 70 min. The cDNA reaction for the isolation of the 3′UTR followed the same protocol as used for the isolation of the 5′UTR, except that SMARTer II A Oligonucleotide was not added, and the 5′-CDS Primer was replaced by 3′-CDS Primer from the kit. Subsequently, a PCR reaction (50 μL) was carried out to amplify the UTR sequence: 1 μL cDNA (for 5′ end UTR cDNA or 3′end UTR cDNA), 5 μL PCR buffer, 1 μL dNTPs (10 mM), 5 μL 10X UMP primer, 0.5 mM gene-specific primer (for 5′UTR or 3′UTR) and 0.25 μL DreamTaq DNA polymerase. The PCR thermal cycle was: denaturation for 3 min at 95 °C, followed by 35 cycles of amplification; 30 s at 95 °C; 30 s at 56 °C; and 2 min at 72 °C. The reaction was extended for 10 min at 72 °C and then cooled to 4 °C. The UTR PCR products were purified from agarose gel using the Wizard® SV gel and PCR Clean-up System (Promega, Mandison, WI, USA) for subsequent, direct sequencing. The full length of the UTR sequence was assembled by Vector NTI software (Invitrogen, Waltham, MA, USA). For mapping analysis, the full length of the isolated transcript was mapped to the genome of A. franciscana (Unit of Polar Genomics, Korea Polar Research Institute, version 1.0, http://antagen.kopri.re.kr/project/genome_info_iframe.php?Code=AF01 accessed on 26 June 2022) using BLASTN with default parameters.
2.5. Phylogenetic Analysis
The phylogenetic tree was generated using website NGPhylogeny (https://ngphylogeny.fr, accessed on 17 September 2022), a phylogeny software based on the maximum-likelihook [26], wherein the usual bootstrapping procedure is replaced by a fast approximate likelihood ratio test (aLRT), which is proven to be a good alternative to the (time-consuming) bootstrap analysis. The putative polypeptide sequence from the isolated transcripts and known polypeptide sequences of homologs from NCBI were introduced under FASTA format.
2.6. Transcription Proof of Dsx/Dmrt Transcript
Gene-specific PCR primers was designed for each Dsx/Dmrt gene to amplify the complete transcript (Table 1). The applied PCR conditions were the same for all genes in this study. In addition, the PCR reaction contained 150 ng cDNA, 5 μL reaction buffer, 1 μL dNTPs (10 mM), 0.5 mM gene-specific primers, (Table 1) and 0.25 μL DreamTaq DNA polymerase (Thermal Scientific, Waltham, MA, USA). The PCR reaction was carried out in a thermal cycler (2720 Thermal Cycler (Thermal Scientific, Waltham, MA, USA) programmed for 94 °C for 4 min; 35 cycles at 94 °C for 30 s, 55 °C for 60 s, and 72 °C for 5 min; followed by 72 °C for 10 min. The amplicons were then separated on agarose gel and cleaned using Wizard® SV Gel and PCR Clean-Up System (Promega, Madison, WI, USA). These cleaned amplicons were used directly for sequencing by the Sanger sequencing method using the same PCR primers.
3. Results
3.1. Identification of Dsx/Dmrt Transcripts
Six contigs encoding for polypeptides with a Doublesex/Mab-3 domain [27] were found in the A. franciscana transcriptome. These contigs were named doublesex-1, doublesex-2, doublesex-3, doublesex-4, doublesex-5 and doublesex-6. Based on the nucleotide sequence, only doublesex-3, doublesex-5 and doublesex-6 contained a full CDS encoding for the full putative polypeptide of the protein, while the others contained only a partial CDS. Therefore, the full doublesex-1, doublesex-2, and doublesex-4 cDNA sequences were then identified by the RACE method. Interestingly, RACE results revealed three different isoform from the doublesex-4 contig, namely doublesex-4α isoform A of 920 bp, doublesex-4α isoform B of 1010 bp; and doublesex-4β isoform C of 1156 bp. Their nucleotide sequences were submitted to NCBI GenBank as doublesex-1 (dsx-1) (MF287957), doublesex-2 (dsx-2) (MF287958), doublesex-3 (dsx-3) (MF287959), doublesex-4α isoform A (dsx-4αA) (MF287960), doublesex-4α isoform B (dsx-4αB) (MF993793), doublesex-4-β isoform C (dsx-4βC) (MF287961), doublesex-5 (dsx-5) (MF287962) and doublesex-6 (dsx-6) (MF287963).
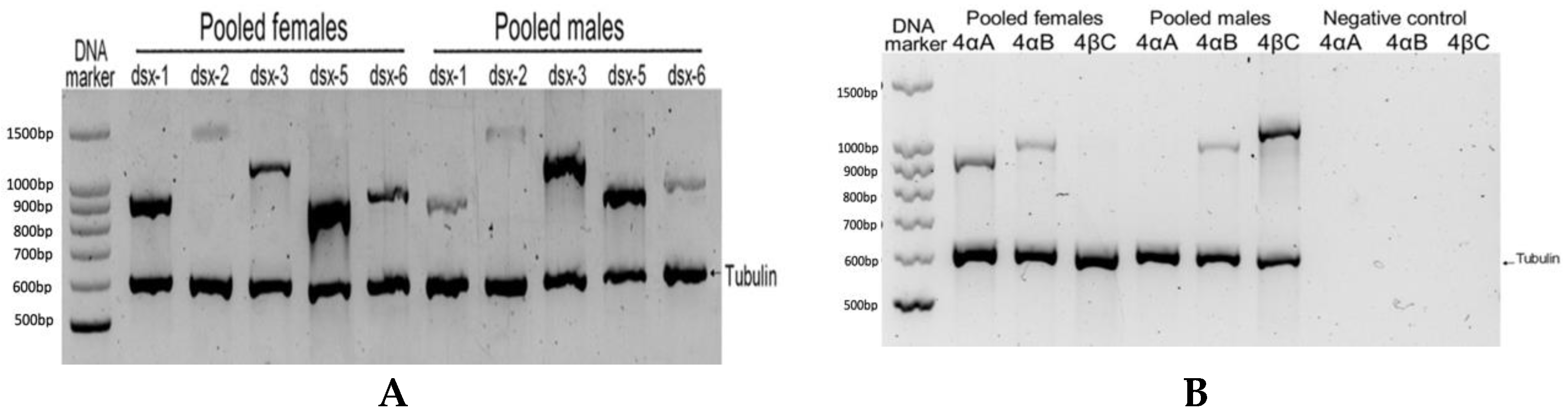
The presence of the isolated Dsx/Dmrt transcripts were confirmed in adult A. franciscana animals by PCR method. The sex specific PCR analysis was conducted on cDNA from whole body RNA extraction of pooled male or female adults showed the presence of Dsx-1, Dsx-2, Dsx-3, Dsx-5 and Dsx-6 transcripts in both sexes (Figure 1A). However, dsx-4βC transcript could only be seen in the pooled males, while dsx-4αA transcript could only be found in the pooled females. Interestingly, dsx-4αB transcript was present in both sexes (Figure 1B).
3.2. Genome Mapping and Putative Protein Translation
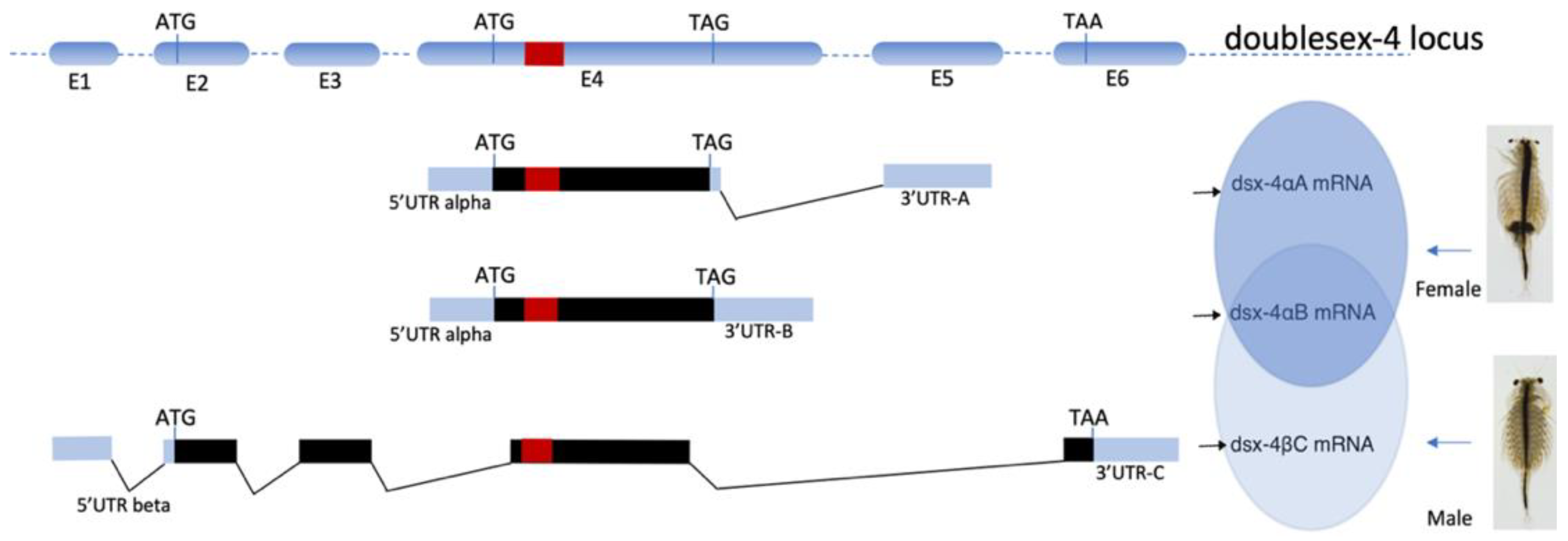
The nucleotide sequence BLAST results of isolated Dsx/Dmrt transcripts against the A. franciscana genome assembly (NCBI Bioproject PRJNA589114) confirmed these stranscripts are transcribed from different loci on six diferent scaffolds. These loci encoded putatively different DSX proteins bearing one DM domain [28,29]. Among those, the DSX protein that is translated by dsx-2 transcript beared an additional ubiquitin—associated domain (UBA domain) at its C-terminal, which is a common domain found in DSX proteins in insect species [28]. Genome mapping of the full-length of three transcripts from putative dsx-4 contig to the genome of A. franciscana revealed a DNA scaffold (scaffold901_size138921) bearing an intact locus, namely Af.dsx-4 locus, which contained six different exons and its arrangement on this scaffold. The organization of six exons on Af.dsx-4 locus and the sequence of dsx-4αB, dsx-4αA, and dsx-4βC transcripts suggest that Af.dsx-4 locus could produces three alternative transcripts including dsx-4αB, dsx-4αA, and dsx-4βC as namely Af.dsx4, Af.dsxF and Af.dsxM, respectively. Therefore, these three transcripts of Af.dsx-4 locus are a probable consequence of a splicing event (Figure 2).
The open reading frame prediction of Af.dsxF and Af.dsx4 transcripts showed that they only contained different 3′UTR sequences which are namely 3′UTR-A and 3′UTR-B, respectively. Therefore, they are predicted to encode for an identical putative polypeptide of 186 aa long, namely Af.DSX-F. In contrast, the Af.dsxM transcript is predicted to encode a polypeptide of 289 aa long, namely Af.DSX-M. The sequence at the 5′end (consisting of a 5′ untranslated region (5′UTR) and a partial CDS encoding for the N-terminal) of Af.dsxM and Af.dsxF transcripts are completely different sequences. Additionally, the Af.dsxM transcript had a different 3′UTR sequence, namely 3′UTR-C, in comparison to the 3′UTR-A and 3′UTR-B sequence of Af.dsxF and Af.dsx4 transcripts, respectively. The Af.dsx4, Af.dsxF and Af.dsxM transcripts shared a sequence of 477 nucleotides encoding a polypeptide, including the DM domain. The polypeptide alignment suggested that Af.DSX-M is probably an isoform of Af.DSX-F but bears an extended polypeptide at its N-terminal (Figure 3).
3.3. Phylogenetic Tree
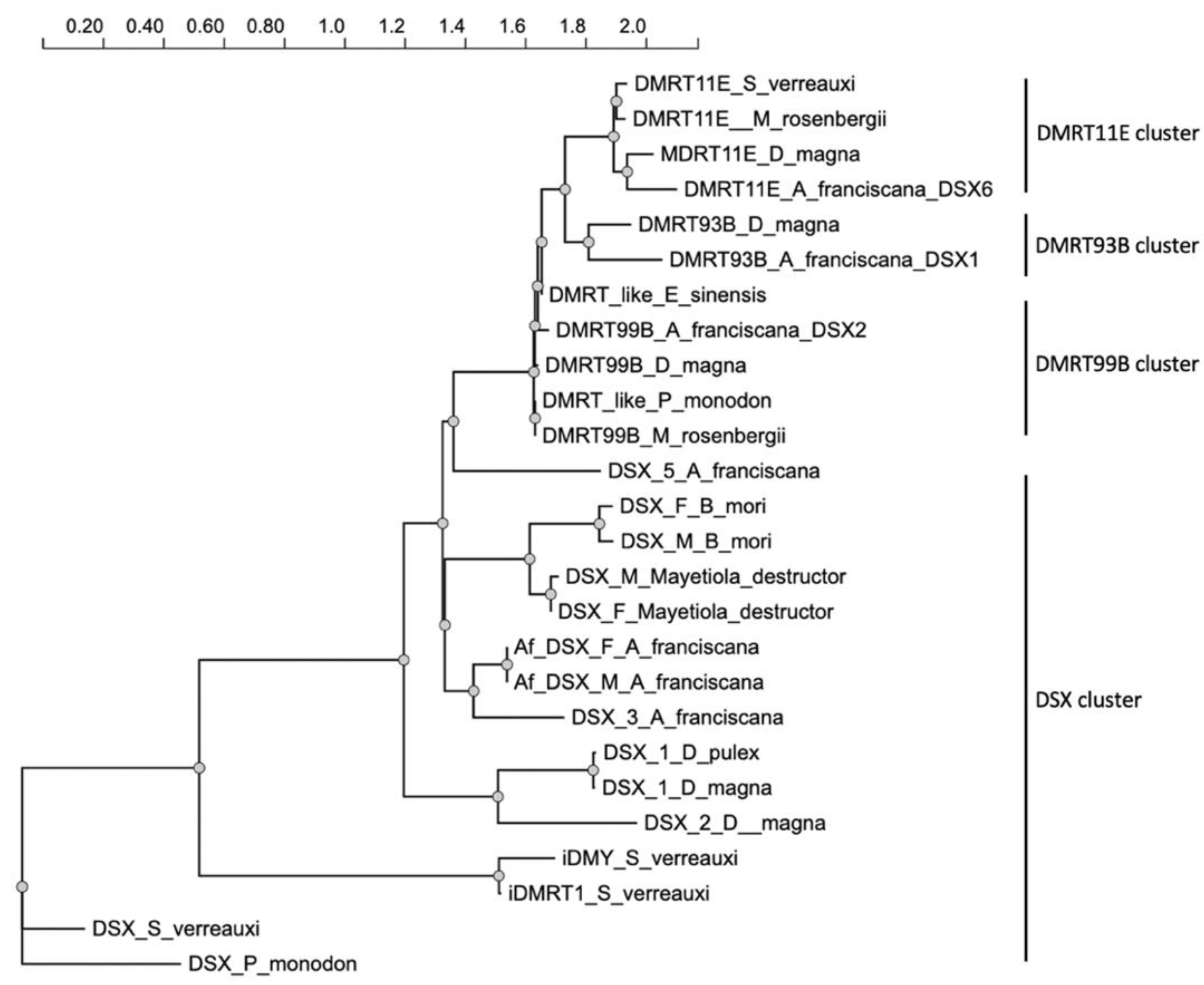
Eight transcripts were identified encoding seven putative DSX proteins in A. franciscana. The polypeptide phylogenetic tree showed that A. franciscana DSX-1, DSX-2 and DSX-6 are related to DMRT93B, DMRT99B and DMRT11E protein from crustaceans D. magna, E. sinensis, M. rosenbergii and S. verreauxi. In contrast, putative A. franciscana DSX-3, Af.DSX-F, Af.DSX-F and DSX-5 polypeptides are closely related to the sex-specific DSX proteins cluster (Figure 4).
3.4. Sex-Specific Splicing of Doublesex Gene
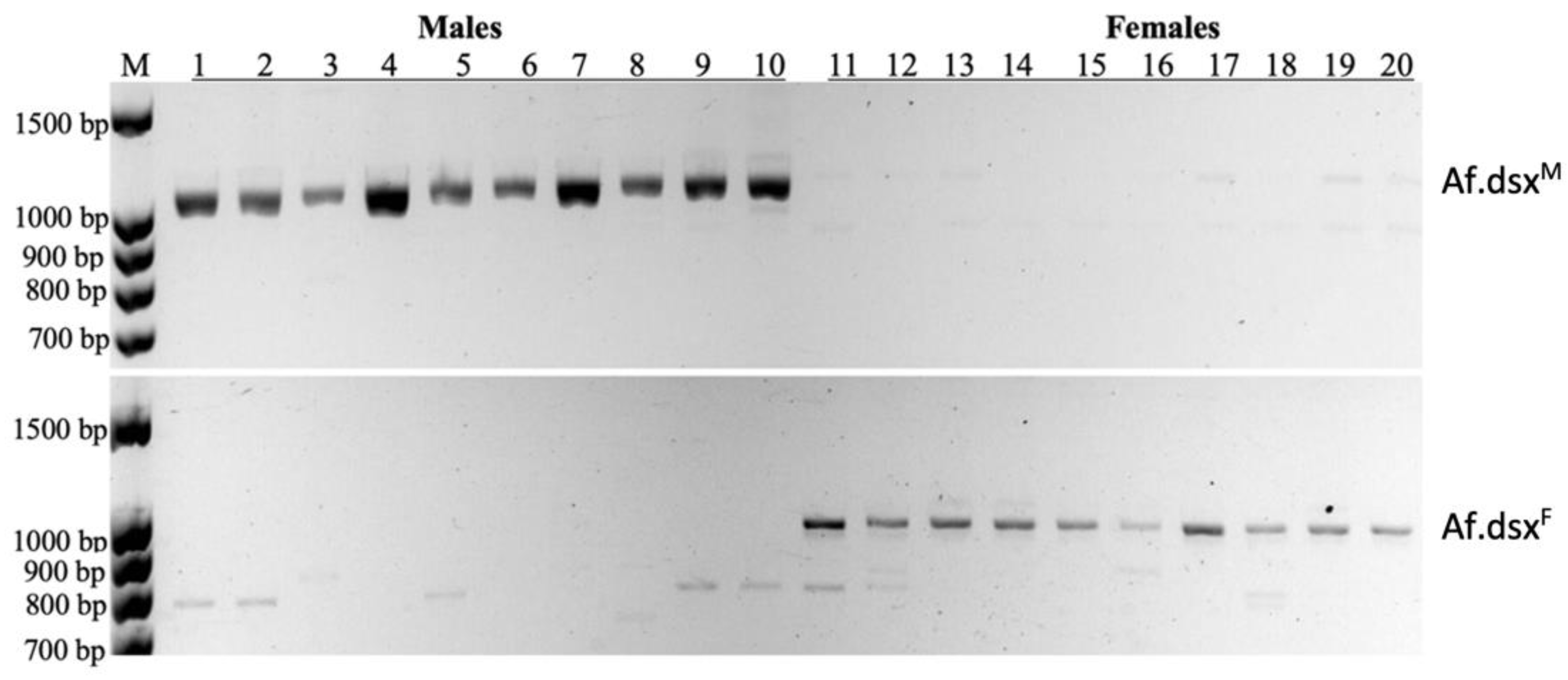
The presence of sex-specific transcripts originating from Af.dsx-4 locus was searched on A. franciscana individuals, which were reared separately from the juvenile until the adult stage (full maturity). The sex-specific PCR analysis of Af.dsxF and Af.dsxF transcripts at the juvenile stage was performed on twenty juvenile individuals separately. It again confirmed that Af.dsxF and Af.dsxM were female and male-specific transcripts, respectively (Figure 5).
4. Discussion
Extensive studies of the sex determination mechanism in invertebrate animal models such as D. melanogaster have shown that sex is determined in the embryonic stage by the involvement of so-called sex-determining genes. Particularly, dsx genes have been identified in many insects to be linked to the sex determination pathway [13]. In A. franciscana, the available RNAseq databases suggest that this species has many genes that are similar to sex-determining genes in fruit fly [30]. In D. magna, the transcription of dmrt11E and dmrt99B is higher in ovaries than in the testes [31]. In M. rosenbergii, the transcription of both dmrt11E and dmrt99B is found in spermatogonia and spermatozoa during spermatogenesis [15]. In D. magna, the dmrt93B is detected only in the testes, and not in the ovaries [31]. Based on polypeptide phylogenetic analysis, A. franciscana DSX-1, DSX-2 and DSX-6 are very close to DMRT93B, DMRT99B and DMRT11E, respectively, thus they are hypothesized to play a similar role in development of gonadal tissue in Artemia.
In insects, sex-specific splicing generates sex-specific transcripts from the sex-specific dsx gene controlling sexually-dimorphic traits. The disruption of dsx gene function produces either male-specific sexually-dimorphic defects or intersexual phenotypes [10]. Sex-specific dsx splicing in B. mori depends on the presence of MASC protein in the embryo to promote the production of the male-specific splicing variants [32]. Recently, a masc gene has been characterized in A. franciscana. Masc RNAi during embryonic development of sexual A. franciscana could slightly change the ratio of females/males [9]. This suggests that A. franciscana probably also has a similar sex determining mechanism in which the splicing of dsx plays an important role in sex determination. In this study, the RACE method has not been applied on A. franciscana dsx-3 and dsx-5, thus potential variant transcripts of these two genes remain unknown. However, three diferent transcripts of Af.dsx-4 locus were isolated in A. franciscana suggesting that a specific splicing mechanism of dsx may exist in A. franciscana. In crustaceans, up to now, evidence for alternative splicing of the dsx gene could only be found in D. magna, involved in environmental sex determination. Doublesex-1 of D. magna is identified to directly determine the gender via the splicing at 5′end of mRNA to produce doublesex-1α or doublesex-1β transcript. Therefore, they have different 5′UTR sequences in their structural transcript, which leads to the control of the translation of key regulator protein of the male phenotype [22,33]. In A. franciscana, the Af.dsxM isoform has a different 5′UTR sequence in comparison to the Af.dsxF and Af.dsx4 isoforms, thus this may influence the efficient translation to corresponding protein. In comparison to the AF.DSX-F primary polypeptide sequence, AF.DSX-M has a longer N-terminal sequence, this difference could affect the formation of the dimer structure that is needed for its function [34].
Moreover, the transcription of Af.dsxM could only be found in male animals, and Af.dsxF was only present in female animals (Figure 5). This reinforces the hypothesis that sex-specific transcripts of Af.dsx-4 are related to sex determination in A. franciscana. Interestingly, Af.dsx4 was found present in both sexes (Figure 1B) and since Af.dsxF and Af.dsx4 probably encode for the same Af.DSX-F protein, females are predicted to have only Af.DSX-F homodimer. In contrast, the male has both Af.dsxM and Af.dsx4, therefore their Af.DSX-F and Af.DSX-M proteins probably are present together, resulting in three different types of protein dimer (Af.DSX-F homodimer, Af.DSX-M homodimer and Af.DSX-F/Af.DSX-M heterodimer). In B. mori, the complication of cis- and trans-spliced transcripts of dsx has also been reported to generate seventeen alternatively spliced forms and eleven putative trans-spliced variants. However, all transcripts encode for only four female-specific, two male-specific DSX proteins and one DSX protein common to males and females [35,36]. Therefore, the alternative splicing of Af.dsx-4 locus may need to be investigated further to unravel its precise role in sex determination.
5. Conclusions
Six Dsx/Dmrt gene family loci were identified on different scaffolds, and their transcripts were sequenced from a cDNA sample of A. franciscana in this study. The expression profiles showed that these genes are all expressed in both sexes except the Af.dsx-4 locus. We determined that the Af.dsx-4 locus potentially transcribes three different isoforms, including Af.dsxM, Af.dsxF and Af.dsx4, probably due to a splicing event. Af.dsxF and Af.dsx4 transcripts are found only in female animals, whereas Af.dsxM and Af.dsx4 transcripts are detected in male animals. These three transcripts of Af.dsx-4 locus putatively encode Af.DSX-F proteins and Af.DSX-M protein, which is characterized by large differences in amino acid sequence at the N-terminal and minor changes at the C-terminal. Herein, Af.dsx-4 is a suitable candidate for further investigations into sex determination in A. franciscana.
Author Contributions
D.N.V.: Methodology, Investigation, Validation, Writing—Original Draft; O.C.: Supervision, Review & Editing; S.D.V.: Resources, Review & Editing; G.S. and P.B.: Review & Editing, Project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Vietnam International Education Development.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Vos, S.; Bossier, P.; Van Stappen, G.; Vercauteren, I.; Sorgeloos, P.; Vuylsteke, M. A First AFLP-Based Genetic Linkage Map for Brine Shrimp Artemia franciscana and Its Application in Mapping the Sex Locus. PLoS ONE 2013, 8, e57585. [Google Scholar] [CrossRef] [Green Version]
- Bowen, S.T. The Genetics of Artemia salina. V. Crossing over between the X and Y Chromosomes. Genetics 1965, 52, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Parnes, S.; Khalaila, I.; Hulata, G.; Sagi, A. Sex Determination in Crayfish: Are Intersex Cherax quadricarinatus (Decapoda, Parastacidae) Genetically Females? Genet. Res. 2003, 82, 107–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staelens, J.; Rombaut, D.; Vercauteren, I.; Argue, B.; Benzie, J.; Vuylsteke, M. High-Density Linkage Maps and Sex-Linked Markers for the Black Tiger Shrimp (Penaeus monodon). Genetics 2008, 179, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Ventura, T.; Sagi, A. The Insulin-like Androgenic Gland Hormone in Crustaceans: From a Single Gene Silencing to a Wide Array of Sexual Manipulation-Based Biotechnologies. Biotechnol. Adv. 2012, 30, 1543–1550. [Google Scholar] [CrossRef]
- Cui, Z.; Hui, M.; Liu, Y.; Song, C.; Li, X.; Li, Y.; Liu, L.; Shi, G.; Wang, S.; Li, F.; et al. High-Density Linkage Mapping Aided by Transcriptomics Documents ZW Sex Determination System in the Chinese Mitten Crab Eriocheir sinensis. Heredity 2015, 115, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Huylmans, A.K.; Toups, M.A.; MacOn, A.; Gammerdinger, W.J.; Vicoso, B. Sex-Biased Gene Expression and Dosage Compensation on the Artemia Franciscana Z-Chromosome. Genome Biol Evol 2019, 11, 1033–1044. [Google Scholar] [CrossRef] [Green Version]
- Jo, E.; Lee, S.J.; Choi, E.; Kim, J.; Lee, J.H.; Park, H. Sex-Biased Gene Expression and Isoform Profile of Brine Shrimp Artemia Franciscana by Transcriptome Analysis. Animals 2021, 11, 2630. [Google Scholar] [CrossRef]
- Li, D.-R.; Ye, H.-L.; Yang, J.-S.; Yang, F.; Wang, M.-R.; de Vos, S.; Vuylsteke, M.; Sorgeloos, P.; van Stappen, G.; Bossier, P.; et al. Identification and Characterization of a Masculinizer (Masc) Gene Involved in Sex Differentiation in Artemia. Gene 2017, 614, 56–64. [Google Scholar] [CrossRef]
- Xu, J.; Zhan, S.; Chen, S.; Zeng, B.; Li, Z.; James, A.A.; Tan, A.; Huang, Y. Sexually Dimorphic Traits in the Silkworm, Bombyx Mori, Are Regulated by Doublesex. Insect Biochem Mol. Biol 2017, 80, 42–51. [Google Scholar] [CrossRef]
- Mawaribuchi, S.; Ito, Y.; Ito, M. Independent Evolution for Sex Determination and Differentiation in the DMRT Family in Animals. Biol. Open 2019, 8, bio041962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burtis, K.C.; Baker, B.S. Drosophila Doublesex Gene Controls Somatic Sexual Differentiation by Producing Alterna-tively Spliced MRNAs Encoding Related Sex-Specific Polypeptides. Cell 1989, 56, 997–1010. [Google Scholar] [CrossRef]
- Verhulst, E.C.; van de Zande, L. Double Nexus—Doublesex Is the Connecting Element in Sex Determination. Brief. Funct. Genom. 2015, 14, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Yoon, B.-H.; Kim, W.-J.; Kim, D.-W.; Hurwood, D.A.; Lyons, R.E.; Salin, K.R.; Kim, H.-S.; Baek, I.; Chand, V.; et al. Optimizing Hybrid de Novo Transcriptome Assembly and Extending Genomic Resources for Giant Freshwater Prawns (Macrobrachium Rosenbergii): The Identification of Genes and Markers Associated with Re-production. Int. J. Mol. Sci. 2016, 17, 690. [Google Scholar] [CrossRef]
- Yu, Y.-Q.; Ma, W.-M.; Zeng, Q.-G.; Qian, Y.-Q.; Yang, J.-S.; Yang, W.-J. Molecular Cloning and Sexually Di-morphic Expression of Two Dmrt Genes in the Giant Freshwater Prawn, Macrobrachium Rosenbergii. Agric. Res. 2014, 3, 181–191. [Google Scholar] [CrossRef]
- Amterat Abu Abayed, F.; Manor, R.; Aflalo, E.D.; Sagi, A. Screening for Dmrt Genes from Embryo to Mature Macro-brachium Rosenbergii Prawns. Gen. Comp. Endocrinol. 2019, 282, 113205. [Google Scholar] [CrossRef]
- Zhong, P.; Zhou, T.; Zhang, Y.; Chen, Y.; Yi, J.; Lin, W.; Guo, Z.; Xu, A.; Yang, S.; Chan, S.; et al. Potential Involvement of a DMRT Family Member (Mr-Dsx) in the Regulation of Sexual Differentiation and Moulting in the Giant River Prawn Macrobrachium Rosenbergii. Aquac. Res. 2019, 50, 3037–3049. [Google Scholar] [CrossRef]
- Ma, K.; Qiu, G.; Feng, J.; Li, J. Transcriptome Analysis of the Oriental River Prawn, Macrobrachium Nipponense Using 454 Pyrosequencing for Discovery of Genes and Markers. PLoS ONE 2012, 7, e39727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, E.F.; Qiu, G.F. A Novel Dmrt Gene Is Specifically Expressed in the Testis of Chinese Mitten Crab, Eriocheir Sinensis. Dev. Genes. Evol. 2010, 220, 151–159. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Yu, K.; Xiang, J. Identification and Characterization of a Doublesex Gene Which Regulates the Expression of Insulin-like Androgenic Gland Hormone in Fenneropenaeus Chinensis. Gene 2018, 649, 1–7. [Google Scholar] [CrossRef]
- Peng, J.; Wei, P.; Zhang, B.; Zhao, Y.; Zeng, D.; Chen, X.; Li, M.; Chen, X. Gonadal Transcriptomic Analysis and Dif-ferentially Expressed Genes in the Testis and Ovary of the Pacific White Shrimp (Litopenaeus Vannamei). BMC Genom. 2015, 16, 1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental Sex Determination in the Branchiopod Crustacean Daphnia Magna: Deep Consevation of a Doublesex Gene in the Sex-Determining Pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, J.C.; Fitzgibbon, Q.P.; Smith, G.; Elizur, A.; Ventura, T. Y-Linked IDmrt1 Paralogue (IDMY) in the Eastern Spiny Lobster, Sagmariasus Verreauxi: The First Invertebrate Sex-Linked Dmrt. Dev. Biol. 2017, 430, 337–345. [Google Scholar] [CrossRef]
- De Vos, S.; van Stappen, G.; Sorgeloos, P.; Vuylsteke, M.; Rombauts, S.; Bossier, P. Identification of Salt Stress Response Genes Using the Artemia Transcriptome. Aquaculture 2019, 500, 305–314. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhyloge-ny.Fr: New Generation Phylogenetic Services for Non-Specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, W.; Zarkower, D. Similarity of DNA Binding and Transcriptional Regulation by Caenorhabditis Elegans MAB-3 and Drosophila Melanogaster DSX Suggests Conservation of Sex Determining Mechanisms. Development 1999, 126, 873–881. [Google Scholar] [CrossRef]
- Bayrer, J.R.; Zhang, W.; Weiss, M.A. Dimerization of Doublesex Is Mediated by a Cryptic Ubiquitin-Associated Domain Fold: Implications for Sex-Specific Gene Regulation. J. Biol. Chem. 2005, 280, 32989–32996. [Google Scholar] [CrossRef] [Green Version]
- An, W.; Cho, S.; Ishii, H.; Wensink, P.C. Sex-Specific and Non-Sex-Specific Oligomerization Domains in Both of the Doublesex Transcription Factors from Drosophila Melanogaster. Mol. Cell. Biol. 1996, 16, 3106–3111. [Google Scholar] [CrossRef] [Green Version]
- De Vos, S. Genomic Tools and Sex Determination in the Extremophile Brine Shrimp Artemia franciscana. Ph.D. Thesis, Faculty of Bioscience Engineering, Ghent University, Ghent, Belgium, 2014. [Google Scholar]
- Kato, Y.; Kobayashi, K.; Oda, S.; Colbourn, J.K.; Tatarazako, N.; Watanabe, H.; Iguchi, T. Molecular Cloning and Sexually Dimorphic Expression of DM-Domain Genes in Daphnia Magna. Genomics 2008, 91, 94–101. [Google Scholar] [CrossRef]
- Kiuchi, T.; Koga, H.; Kawamoto, M.; Shoji, K.; Sakai, H.; Arai, Y. A Single Female-Specific PiRNA Is the Primary Determiner of Sex in the Silkworm. Nature 2014, 509, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Nong, Q.D.; Matsuura, T.; Kato, Y.; Watanabe, H. Two Doublesex1 Mutants Revealed a Tunable Gene Network Underlying Intersexuality in Daphnia magna. PLoS ONE 2020, 15, e0238256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, B.; Singh, R.; Narendra, U.; Zhu, L.; Weiss, M.A. Regulation of Sexual Dimorphism: Mutational and Chemogenetic Analysis of the Doublesex DM Domain. Mol. Cell. Biol. 2006, 26, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Xu, H.; Guo, H.; O’Brochta, D.A.; Wang, F.; Ma, S.; Zhang, L.; Zha, X.; Zhao, P.; Xia, Q. New Insights into the Genomic Organization and Splicing of the Doublesex Gene, a Terminal Regulator of Sexual Differ-entiation in the Silkworm Bombyx Mori. PLoS ONE 2013, 8, e79703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Xu, H.; Wang, F.; Ma, S.; Zha, X.; Guo, H.; Zhao, P.; Xia, Q. Novel Female-Specific Trans-Spliced and Al-ternative Splice Forms of Dsx in the Silkworm Bombyx Mori. Biochem. Biophys. Res. Commun. 2013, 431, 630–635. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The evidence of transcription of Dsx/Dmrt genes from the cDNA samples of the pooled adult females and pooled adult males in A. franciscana. (A): The presence of dsx-1, dsx-2, dsx-3, dsx-5 and dsx-6 transcripts by semi-quantitative PCR (tubulin was used as the internal control). (B): The alternative transcript of A. franciscana Af.dsx-4, including dsx-4αA, dsx-4αB and dsx-4βC, confirmed the presence by PCR from the cDNA of the pooled adult animals.
Figure 1.
The evidence of transcription of Dsx/Dmrt genes from the cDNA samples of the pooled adult females and pooled adult males in A. franciscana. (A): The presence of dsx-1, dsx-2, dsx-3, dsx-5 and dsx-6 transcripts by semi-quantitative PCR (tubulin was used as the internal control). (B): The alternative transcript of A. franciscana Af.dsx-4, including dsx-4αA, dsx-4αB and dsx-4βC, confirmed the presence by PCR from the cDNA of the pooled adult animals.

Figure 2.
The alternative splicing model of the Af.dsx-4 locus in A. franciscana. E1 to E6 stand for six exons on the DNA scaffold containing the Af.dsx-4 locus. The red bar indicates the location of the DM domain. The black bar is the open reading frame of Af.dsx-4, starting at ATG and stopping at TAG or TAA. The dsx-4αA (Af.dsxF) is female-specific splicing, while the dsx-4βC (Af.dsxM) is male-specific splicing. The dsx-4αB (Af.dsx4) is present in males and females.
Figure 2.
The alternative splicing model of the Af.dsx-4 locus in A. franciscana. E1 to E6 stand for six exons on the DNA scaffold containing the Af.dsx-4 locus. The red bar indicates the location of the DM domain. The black bar is the open reading frame of Af.dsx-4, starting at ATG and stopping at TAG or TAA. The dsx-4αA (Af.dsxF) is female-specific splicing, while the dsx-4βC (Af.dsxM) is male-specific splicing. The dsx-4αB (Af.dsx4) is present in males and females.

Figure 3.
The alignment of primary polypeptide A. franciscana Af.DSX-F and Af.DSX-M. The box indicates the DM domain sequence; the asterisks indicate the conserved amino acids.
Figure 3.
The alignment of primary polypeptide A. franciscana Af.DSX-F and Af.DSX-M. The box indicates the DM domain sequence; the asterisks indicate the conserved amino acids.

Figure 4.
The DSX/DMRT protein phylogenetic tree of Artemia franciscana and other arthropods. The phylogeny tree was generated using NGPhylogeny (a free web service) [26].
Figure 4.
The DSX/DMRT protein phylogenetic tree of Artemia franciscana and other arthropods. The phylogeny tree was generated using NGPhylogeny (a free web service) [26].

Figure 5.
The presence of sex-specific transcript encoded by Af.dsx-4 locus in A. franciscana. The sex-specific transcripts of Af.dsxF and Af.dsxF as determined by PCR from the cDNA samples of 20 juvenile animals individually. The amplification PCR products of Af.dsxM transcript could only be seen in the male individuals (lanes 1 to 10), while Af.dsxF transcript could only be found in female individuals (lanes 11 to 20). M: DNA marker.
Figure 5.
The presence of sex-specific transcript encoded by Af.dsx-4 locus in A. franciscana. The sex-specific transcripts of Af.dsxF and Af.dsxF as determined by PCR from the cDNA samples of 20 juvenile animals individually. The amplification PCR products of Af.dsxM transcript could only be seen in the male individuals (lanes 1 to 10), while Af.dsxF transcript could only be found in female individuals (lanes 11 to 20). M: DNA marker.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
PCR primers used to confirm the Dsx/Dmrt transcripts of A. franciscana.
| Genes | Gene-Specific Primers (5′-3′) | Size in bp |
|---|---|---|
| Doublesex-1 (Dmrt93B) | For: ATCCTTTGGGACAATGGATGTTACACTC Rev: CTTTCATAGTGTTGTCAAAACGCGC | 903 |
| Doublesex-2 (Dmrt99B) | For: TGGGAGAGAGACAGGAATGTAAATACATAC Rev: GCAGCTCTGAGTGATACCAAAAAATTAAG | 1459 |
| Doublesex-3 | For: ATATGAAGTGAGCAATGGTGATACGC Rev CGAATTCGTTTAATCACACATTTAAGGCCAC | 1169 |
| Af.dsx-4 | Af.dsxF-For: TCTTCAACCATGAAAACAAACAGGCATCTACC Af.dsxF-Rev-A: ATGTTTTCTGGTGTCTTCTCTGTCACAAC | 920 |
| Af.dsx4-For: TCTTCAACCATGAAAACAAACAGGCATCTACC Af.dsx4-Rev-B: AGAAAGAGATGGTGGAAATTGC | 1010 | |
| Af.dsxM-For: AGTTCGGTTTGTGGTTCCTCACGG Af.dsxM-Rev-C: CAGGCCAATTTTTTGAATATTATCTAGAAGC | 1124 | |
| Doublesex-5 | For: CTACTATCTTACACCTAAGTGCGCCTGAG Rev: GCAACGATTGAAGAGAAATGGGAAAGGATC | 918 |
| Doublesex-6 (Dmrt11E) | For: GATTAAACGAACTAAGGCCAGAGAGA Rev: CACAAAGAACGAAAATAAGAAACTAACACAC | 966 |
| Tubulin | For: GCAGTGGTCTACAAGGTTTC | 605 |
| Rev: TGCATTGACGTCTTTTGGTACGACATCTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Viet, D.N.; Christiaens, O.; De Vos, S.; Smagghe, G.; Bossier, P. The Sex-Specific Splicing of Doublesex in Brine Shrimp Artemia franciscana. Genes 2022, 13, 1997. https://doi.org/10.3390/genes13111997
AMA Style
Viet DN, Christiaens O, De Vos S, Smagghe G, Bossier P. The Sex-Specific Splicing of Doublesex in Brine Shrimp Artemia franciscana. Genes. 2022; 13(11):1997. https://doi.org/10.3390/genes13111997
Chicago/Turabian StyleViet, Dung Nguyen, Olivier Christiaens, Stephanie De Vos, Guy Smagghe, and Peter Bossier. 2022. "The Sex-Specific Splicing of Doublesex in Brine Shrimp Artemia franciscana" Genes 13, no. 11: 1997. https://doi.org/10.3390/genes13111997
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.