
Effect of miR-302b MicroRNA Inhibition on Chicken Primordial Germ Cell Proliferation and Apoptosis Rate

 , ,
, ,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Animal Care
2.2. Isolation, Establishment, and Maintenance of PGC Lines
2.3. Cell Counting Using Arthur Fluorescence Cell Counter
2.4. Apoptosis Rate Measurement Using Arthur Fluorescence Cell Counter
2.5. Immunostaining of PGCs
2.6. RNA Isolation
2.7. CDNA Writing, qRT-PCR
2.8. Inhibition gga-miR-302b-5P and gga-miR-302b-3P Using MicroRNA Inhibitors
2.9. Statistical Analysis
3. Results
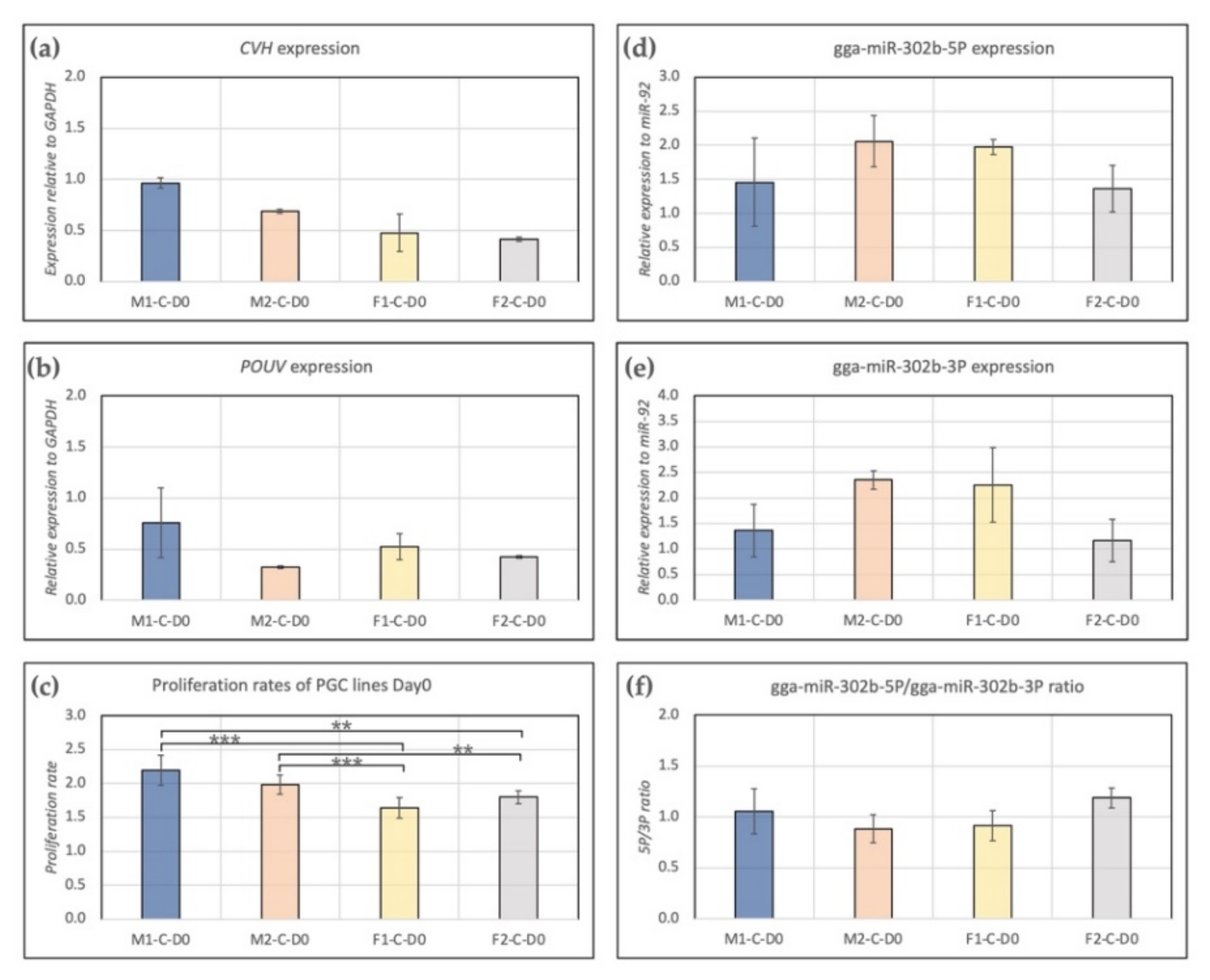
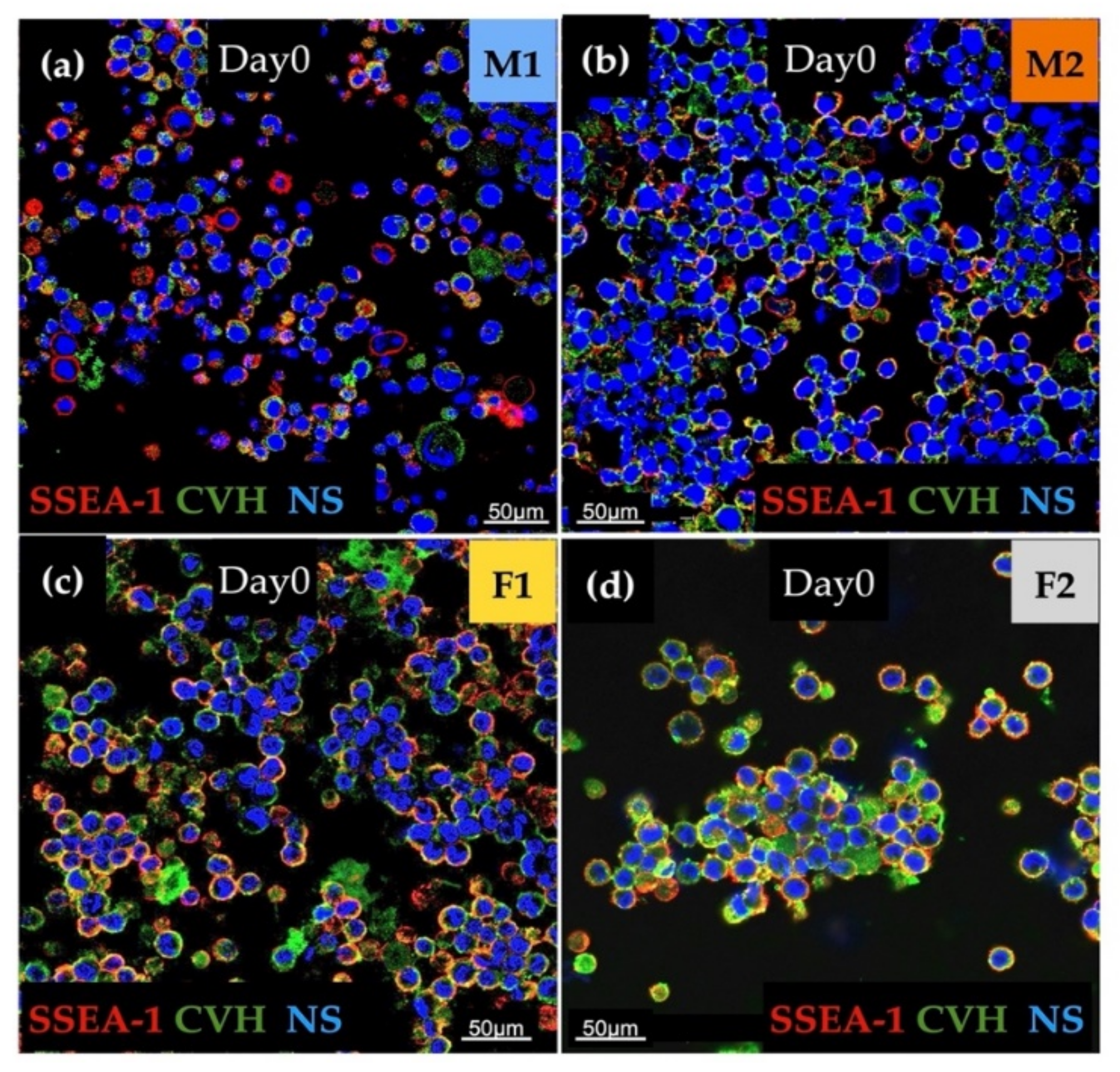
3.1. Characterization of Stem Cell- and Germ Cell-Specific Marker Expression Profile of PG Cell Lines at Day 0
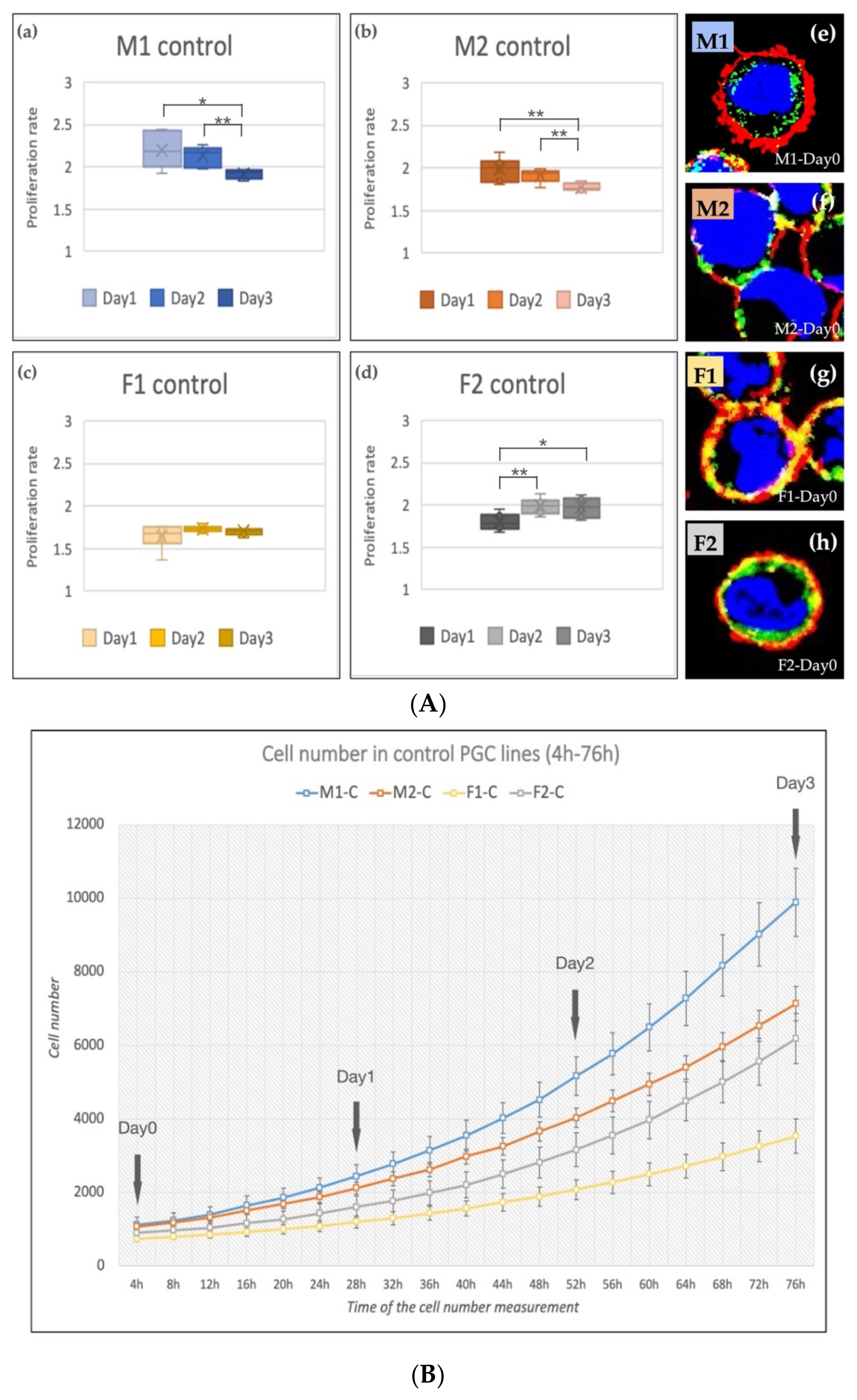
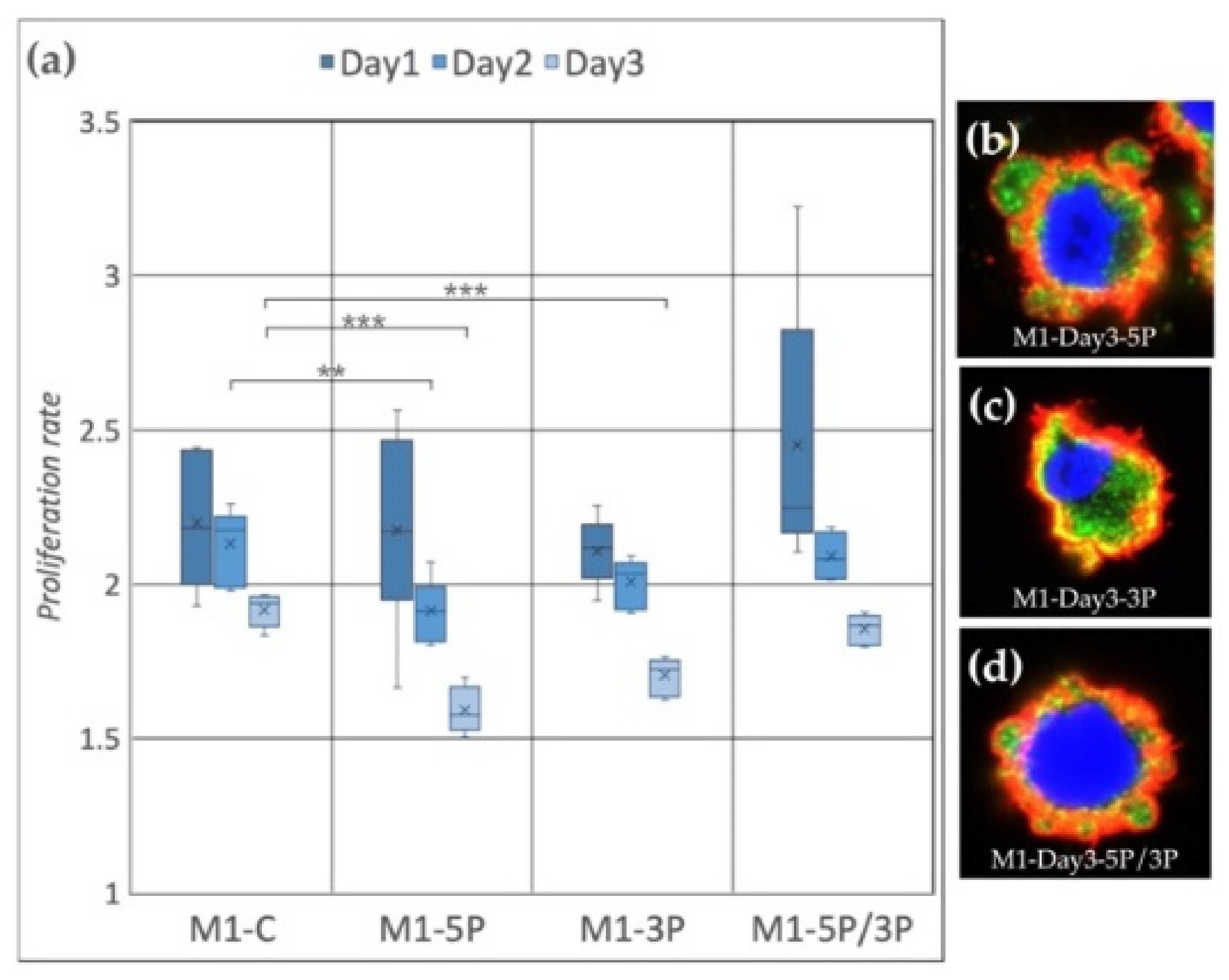
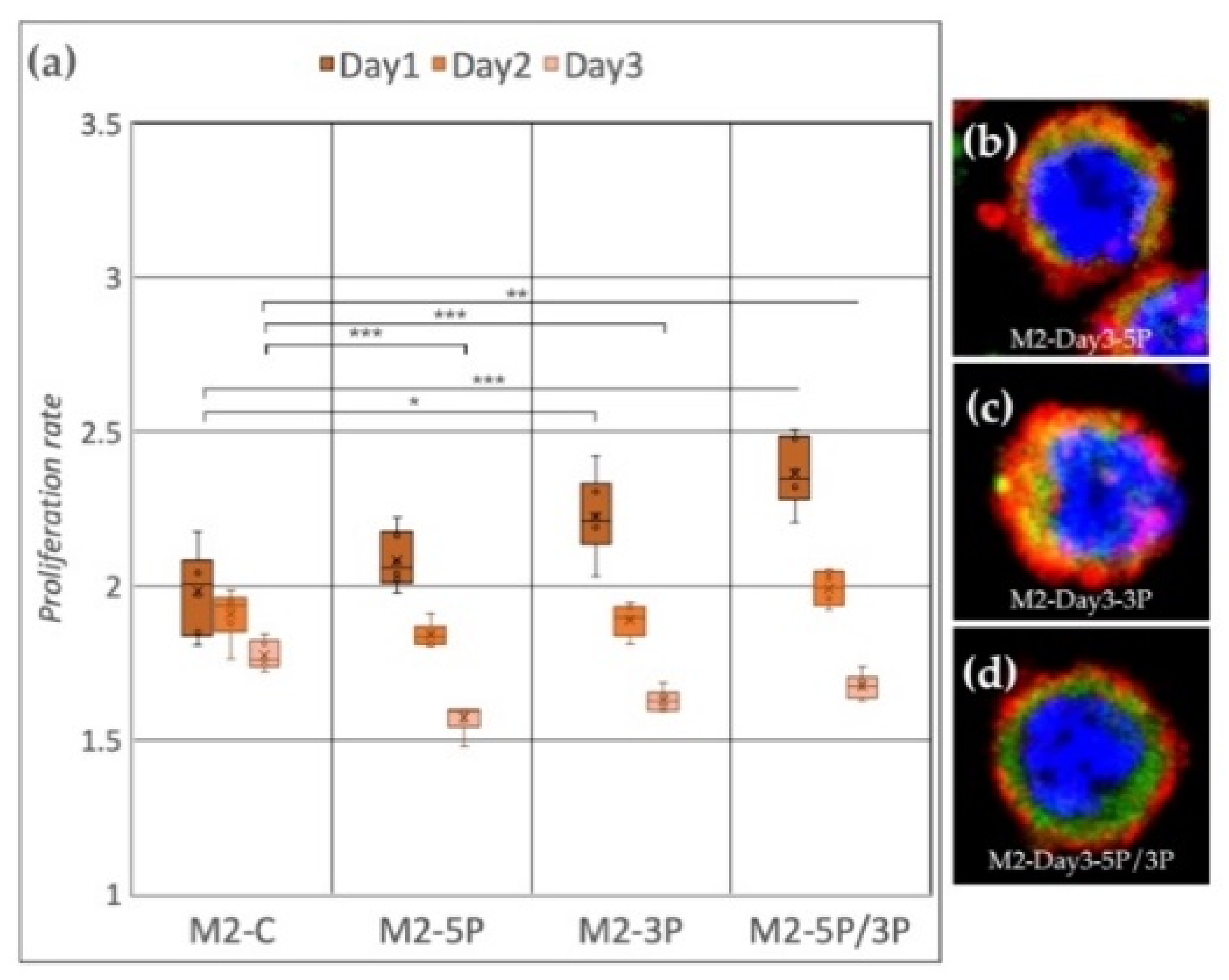
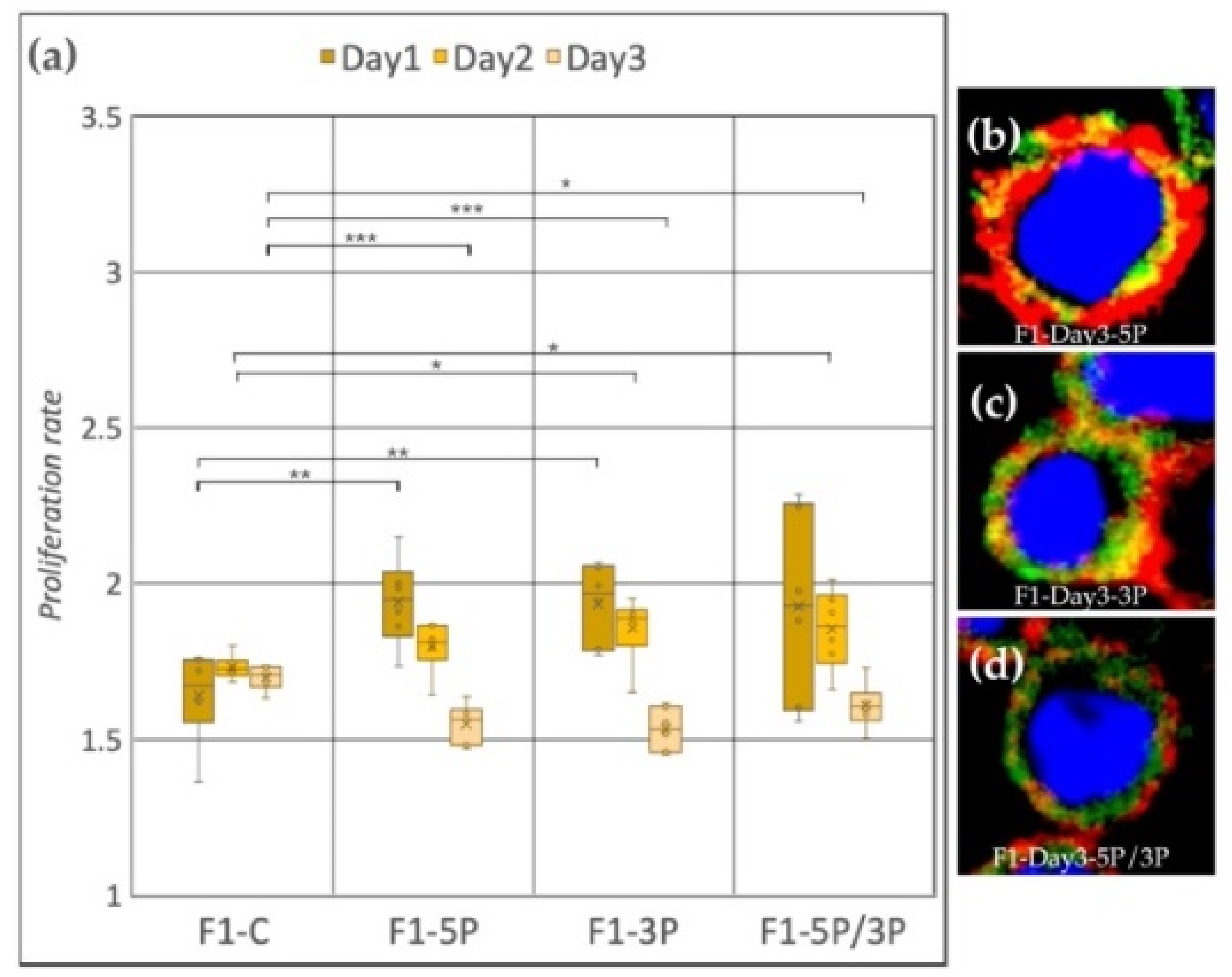
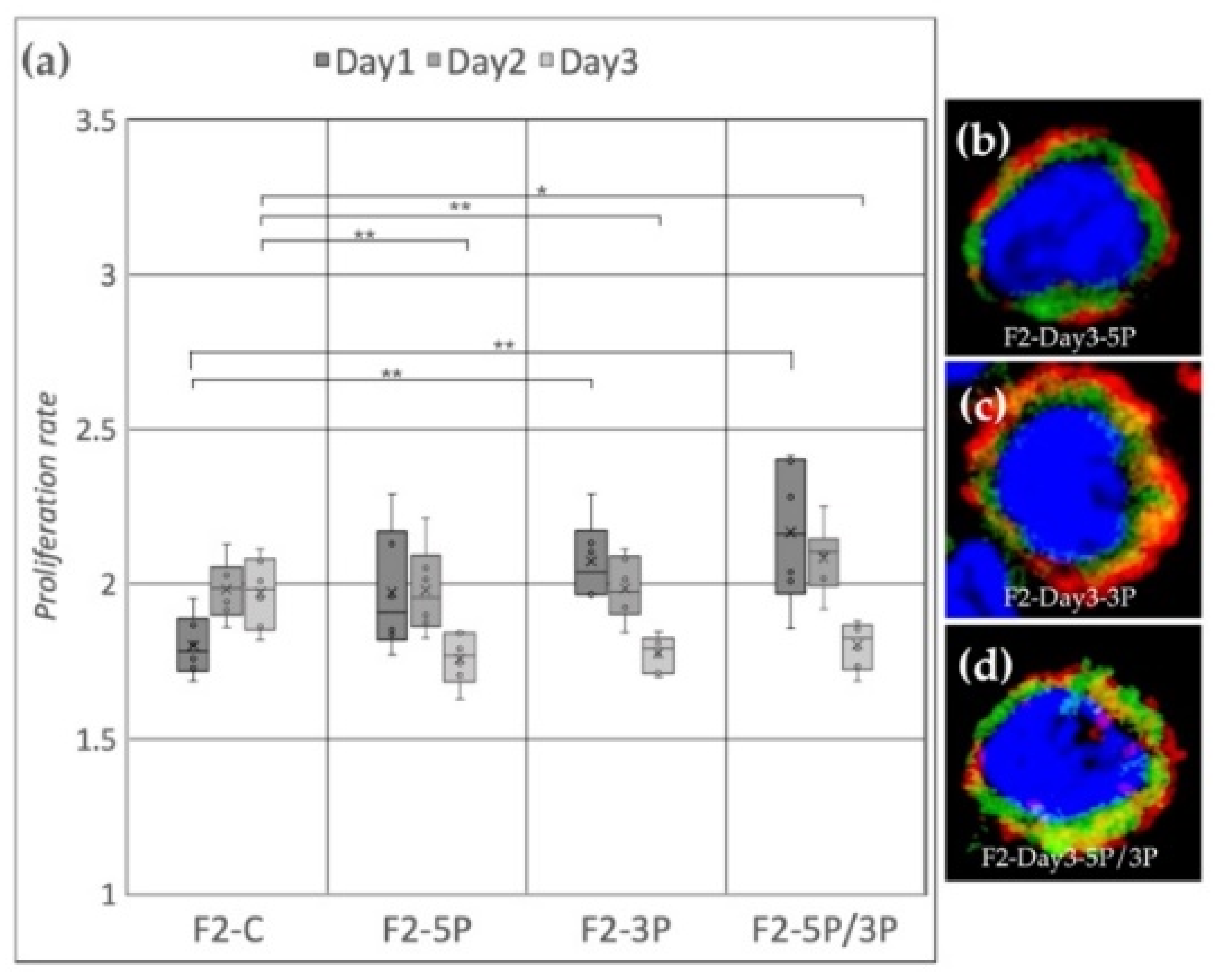
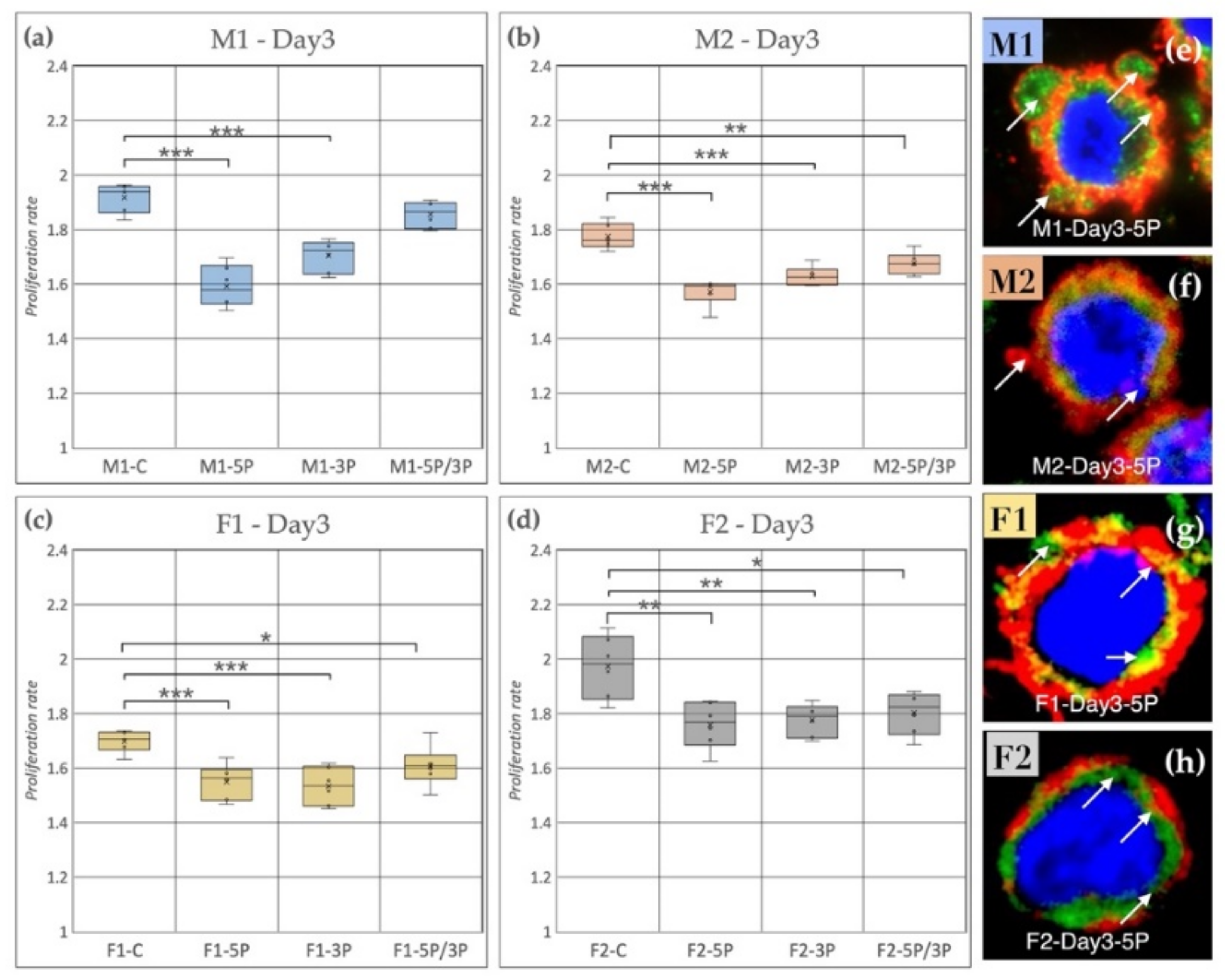
3.2. Effect of Anti-gga-miR-302b-5P, -3P and 5P/3P Inhibition on the Proliferation Rate of PGC Lines
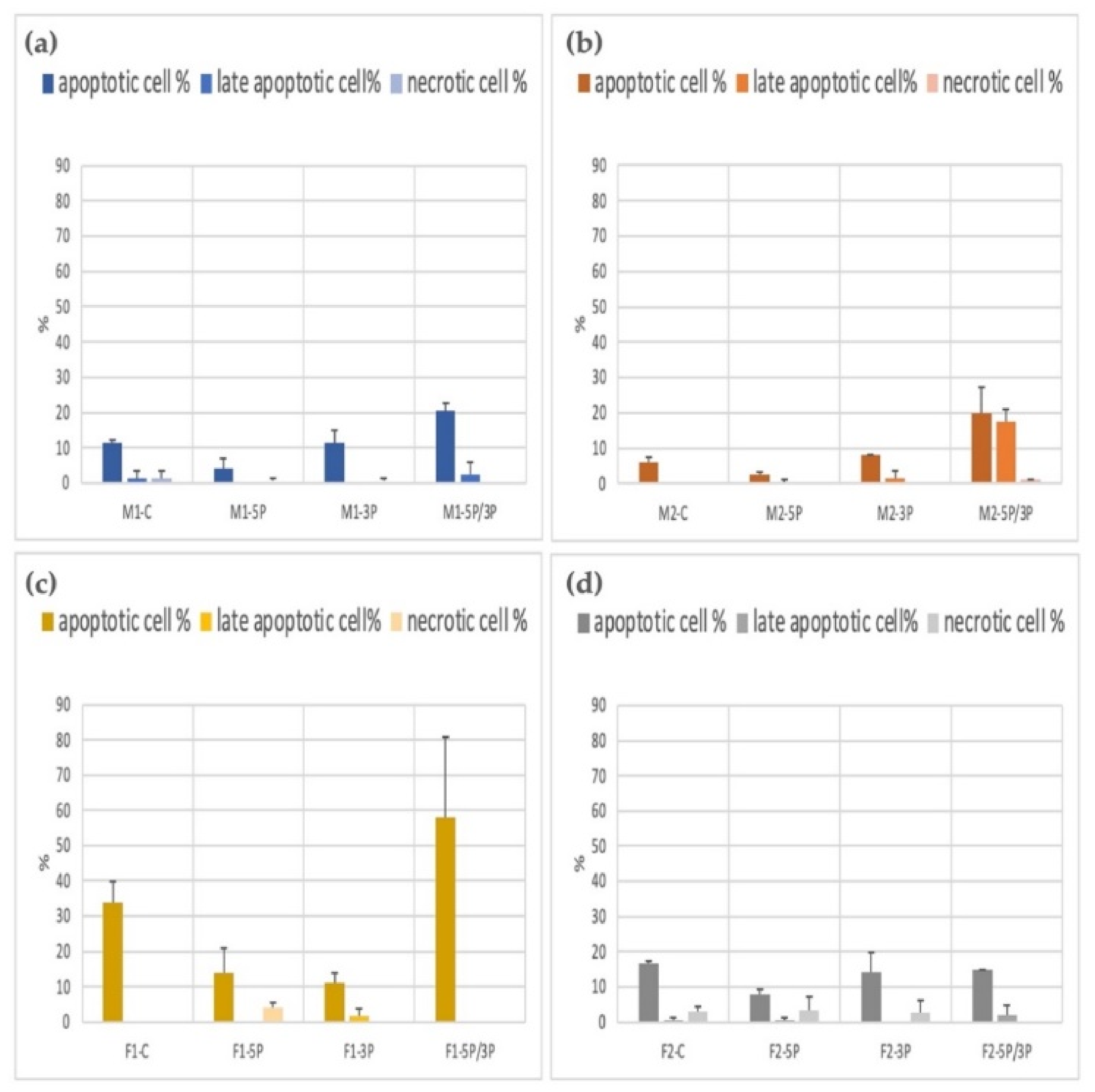
3.3. Determination of the Apoptotic, Late Apoptotic, and Necrotic Cell Ratio of PGC Lines
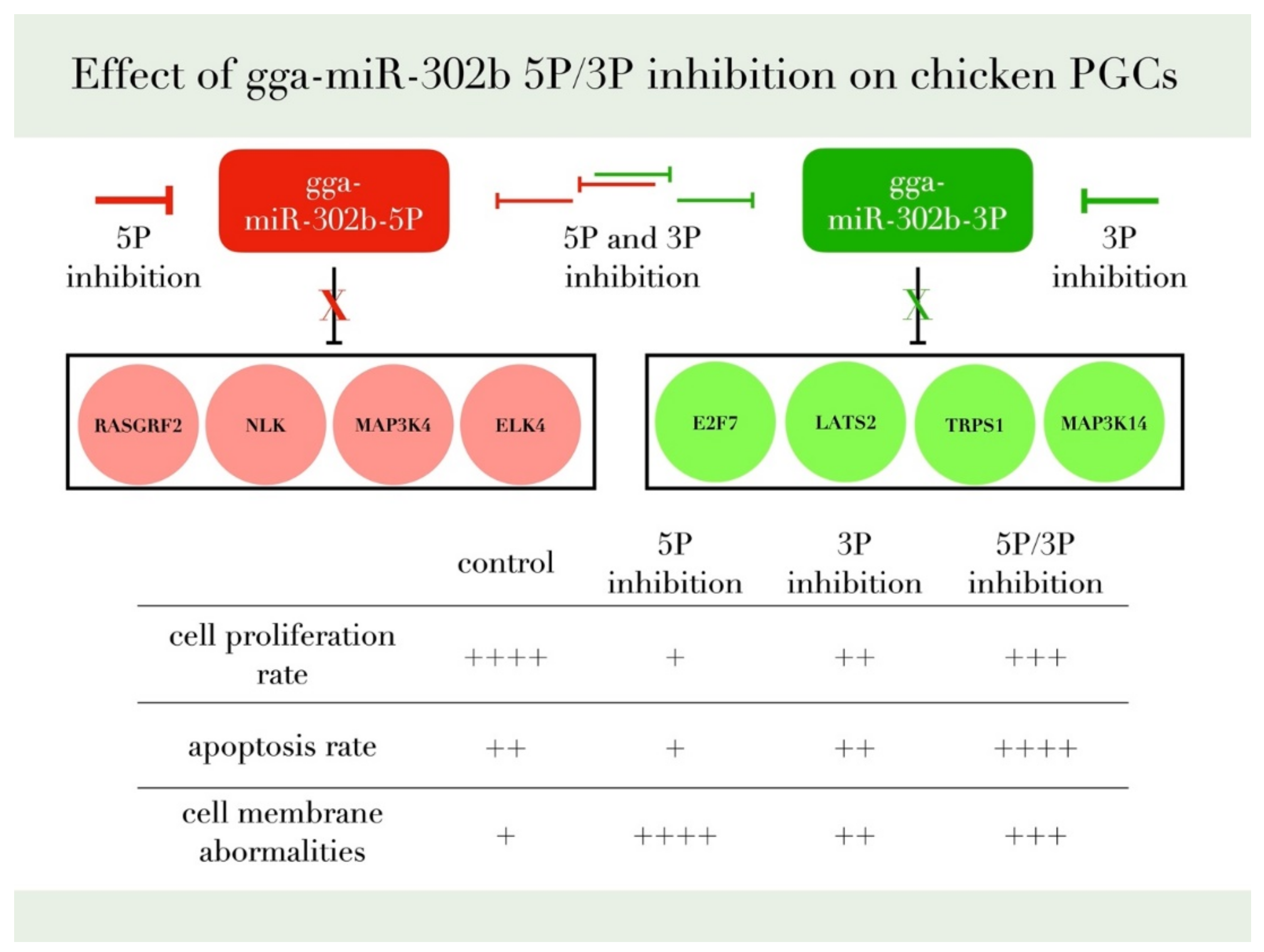
4. Discussion

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reik, W.; Surani, M.A. Germline and Pluripotent Stem Cells. Cold Spring Harb. Perspect. Biol. 2015, 7, a019422. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.J.; Kaufman, M.H. Establishment in Culture of Pluripotential Cells from Mouse Embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Kagami, H.; Tagami, T. Development, Differentiation and Manipulation of Chicken Germ Cells. Dev. Growth Differ. 2013, 55, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Nandi, S.; Whyte, J.; Taylor, L.; Sherman, A.; Nair, V.; Kaiser, P.; McGrew, M.J. Cryopreservation of Specialized Chicken Lines Using Cultured Primordial Germ Cells. Poult. Sci. 2016, 95, 1905–1911. [Google Scholar] [CrossRef] [PubMed]
- Whyte, J.; Glover, J.D.; Woodcock, M.; Brzeszczynska, J.; Taylor, L.; Sherman, A.; Kaiser, P.; McGrew, M.J. FGF, Insulin, and SMAD Signaling Cooperate for Avian Primordial Germ Cell Self-Renewal. Stem Cell Rep. 2015, 5, 1171–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradi, S.; Braun, T.; Baharvand, H. MiR-302b-3p Promotes Self-Renewal Properties in Leukemia Inhibitory Factor-Withdrawn Embryonic Stem Cells. Cell J. 2018, 20, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Rad, S.M.A.H.; Bavarsad, M.S.; Arefian, E.; Jaseb, K.; Shahjahani, M.; Saki, N. The Role of MicroRNAs in Stemness of Cancer Stem Cells. Oncol. Rev. 2013, 7, 8. [Google Scholar] [CrossRef]
- Maraghechi, P.; Hiripi, L.; Tóth, G.; Bontovics, B.; Bosze, Z.; Gócza, E. Discovery of Pluripotency-Associated MicroRNAs in Rabbit Preimplantation Embryos and Embryonic Stem-like Cells. Reproduction 2013, 145, 421–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Long, B.; Han, W.; Yuan, S.; Wang, K. MicroRNAs: Important Regulators of Stem Cells. Stem Cell Res. Ther. 2017, 8, 110. [Google Scholar] [CrossRef]
- Houbaviy, H.B.; Murray, M.F.; Sharp, P.A. Embryonic Stem Cell-Specific MicroRNAs. Dev. Cell 2003, 5, 351–358. [Google Scholar] [CrossRef]
- Rengaraj, D.; Lee, B.R.; Lee, S.I.; Seo, H.W.; Han, J.Y. Expression Patterns and Mirna Regulation of DNA Methyltransferases in Chicken Primordial Germ Cells. PLoS ONE 2011, 6, e19524. [Google Scholar] [CrossRef]
- Rengaraj, D.; Park, T.S.; Lee, S.I.; Lee, B.R.; Han, B.K.; Song, G.; Han, J.Y. Regulation of Glucose Phosphate Isomerase by the 3′UTR-Specific MiRNAs MiR-302b and MiR-17-5p in Chicken Primordial Germ Cells. Biol. Reprod. 2013, 89, 33. [Google Scholar] [CrossRef]
- Houbaviy, H.B.; Dennis, L.; Jaenisch, R.; Sharp, P.A. Characterization of a Highly Variable Eutherian MicroRNA Gene Characterization of a Highly Variable Eutherian MicroRNA Gene. RNA 2005, 11, 1245–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many Roads to Maturity: MicroRNA Biogenesis Pathways and Their Regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef]
- Choo, K.B.; Soon, Y.L.; Nguyen, P.N.N.; Hiew, M.S.Y.; Huang, C.J. MicroRNA-5p and -3p Co-Expression and Cross-Targeting in Colon Cancer Cells. J. Biomed. Sci. 2014, 21, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemzadeh, M.R. Role of Micro RNAs in Stem Cells, Cardiac Differentiation and Cardiovascular Diseases. Gene Rep. 2017, 8, 11–16. [Google Scholar] [CrossRef]
- Darnell, D.K.; Kaur, S.; Stanislaw, S.; Konieczka, J.K.; Yatskievych, T.A.; Antin, P.B. MicroRNA Expression during Chick Embryo Development. Dev. Dyn. 2006, 235, 3156–3165. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nature reviews. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Zhan, J.; Jiao, D.; Wang, Y.; Song, J.; Wu, J.; Wu, L.; Chen, Q.; Ma, S. Integrated MicroRNA and Gene Expression Profiling Reveals the Crucial MiRNAs in Curcumin Anti-Lung Cancer Cell Invasion. Thorac. Cancer 2017, 8, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Jiao, D.M.; Yan, L.; Wang, L.S.; Hu, H.Z.; Tang, X.L.; Chen, J.; Wang, J.; Li, Y.; Chen, Q.Y. Exploration of Inhibitory Mechanisms of Curcumin in Lung Cancer Metastasis Using a MiRNA-Transcription Factor-Target Gene Network. PLoS ONE 2017, 12, e0172470. [Google Scholar]
- Picon-Ruiz, M.; Pan, C.; Drews-Elger, K.; Jang, K.; Besser, A.H.; Zhao, D.; Morata-Tarifa, C.; Kim, M.; Ince, T.A.; Azzam, D.J.; et al. Interactions between Adipocytes and Breast Cancer Cells Stimulate Cytokine Production and Drive Src/Sox2/MiR-302b-Mediated Malignant Progression. Cancer Res. 2016, 76, 491–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodayari, N.; Mohammed, K.A.; Lee, H.; Kaye, F.; Nasreen, N. MicroRNA-302b Targets Mcl-1 and Inhibits Cell Proliferation and Induces Apoptosis in Malignant Pleural Mesothelioma Cells. Am. J. Cancer Res. 2016, 6, 1996. [Google Scholar] [PubMed]
- Lu, X.X.; Cao, L.Y.; Chen, X.; Xiao, J.; Zou, Y.; Chen, Q. PTEN Inhibits Cell Proliferation, Promotes Cell Apoptosis, and Induces Cell Cycle Arrest via Downregulating the PI3K/AKT/HTERT Pathway in Lung Adenocarcinoma A549 Cells. BioMed Res. Int. 2016, 2016, 2476842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lázár, B.; Molnár, M.; Sztán, N.; Végi, B.; Drobnyák, Á.; Tóth, R.; Tokodyné Szabadi, N.; McGrew, M.J.; Gócza, E.; Patakiné Várkonyi, E. Successful Cryopreservation and Regeneration of a Partridge Colored Hungarian Native Chicken Breed Using Primordial Germ Cells. Poult. Sci. 2021, 100, 101207. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A Series of Normal Stages in the Development of the Chick Embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef] [PubMed]
- Lázár, B.; Anand, M.; Tóth, R.; Várkonyi, E.P.; Liptói, K.; Gócza, E. Comparison of the MicroRNA Expression Profiles of Male and Female Avian Primordial Germ Cell Lines. Stem Cells Int. 2018, 2018, 1780679. [Google Scholar] [CrossRef] [Green Version]
- Cai, D.; He, K.; Chang, S.S.; Tong, D.; Huang, C. MicroRNA-302b Enhances the Sensitivity of Hepatocellular Carcinoma Cell Lines to 5-FU via Targeting Mcl-1 and DPYD. Int. J. Mol. Sci. 2015, 16, 23668–23682. [Google Scholar] [CrossRef] [Green Version]
- West, J.A.; Viswanathan, S.R.; Yabuuchi, A.; Cunniff, K.; Takeuchi, A.; Park, I.H.; Sero, J.E.; Zhu, H.; Perez-Atayde, A.; Frazier, A.L.; et al. A Role for Lin28 in Primordial Germ-Cell Development and Germ-Cell Malignancy. Nature 2009, 460, 909–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Chuva de Sousa Lopes, S.M.; Kaneda, M.; Tang, F.; Hajkova, P.; Lao, K.; O’Carroll, D.; Das, P.P.; Tarakhovsky, A.; Miska, E.A.; et al. MicroRNA Biogenesis Is Required for Mouse Primordial Germ Cell Development and Spermatogenesis. PLoS ONE 2008, 3, e1738. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.I.; Lee, B.R.; Hwang, Y.S.; Lee, H.C.; Rengaraj, D.; Song, G.; Park, T.S.; Han, J.Y. MicroRNA-Mediated Posttranscriptional Regulation Is Required for Maintaining Undifferentiated Properties of Blastoderm and Primordial Germ Cells in Chickens. Proc. Natl. Acad. Sci. USA 2011, 108, 10426–10431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.-Y.; Cai, K.-T.; Ma, J.; Chen, G.; Dang, Y.-W.; Lu, H.-P.; Pan, S.-L. Evaluation of MiR-302b-5p Expression and Molecular Mechanism in Hepatocellular Carcinoma: Findings Based on RT-qPCR and in Silico Analysis. Pathol. Res. Pract. 2019, 215, 152424. [Google Scholar] [CrossRef]
- Lin, S.L.; Chang, D.C.; Ying, S.Y.; Leu, D.; Wu, D.T.S. MicroRNA MiR-302 Inhibits the Tumorigenecity of Human Pluripotent Stem Cells by Coordinate Suppression of the CDK2 and CDK4/6 Cell Cycle Pathways. Cancer Res. 2010, 70, 9473–9482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cataldo, A.; Cheung, D.G.; Balsari, A.; Tagliabue, E.; Coppola, V.; Iorio, M.; Palmieri, D.; Croce, C.M. MiR-302b Enhances Breast Cancer Cell Sensitivity to Cisplatin by Regulating E2F1 and the Cellular DNA Damage Response. Oncotarget 2016, 7, 786–797. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, W.-Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.-F. Signaling Pathway of MAPK/ERK in Cell Proliferation, Differentiation, Migration, Senescence and Apoptosis. J. Recept. Signal Transduct. 2015, 35, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Li, D.; Huang, Y.; Cui, X.; Liao, S.; Wang, J.; Liu, F.; Li, C.; Gao, M.; Chen, J.; et al. HSF4 Promotes G1/S Arrest in Human Lens Epithelial Cells by Stabilizing P53. Biochim. Biophys. Acta-Mol. Cell Res. 2015, 1853, 1808–1817. [Google Scholar] [CrossRef] [Green Version]
- Rengaraj, D.; Lee, B.R.; Choi, J.W.; Lee, S.I.; Seo, H.W.; Kim, T.H.; Choi, H.J.; Song, G.; Han, J.Y. Gene Pathways and Cell Cycle-Related Genes in Cultured Avian Primordial Germ Cells. Poult. Sci. 2012, 91, 3167–3177. [Google Scholar] [CrossRef]
- Kang, K.S.; Lee, H.C.; Kim, H.J.; Lee, H.G.; Kim, Y.M.; Lee, H.J.; Park, Y.H.; Yang, S.Y.; Rengaraj, D.; Park, T.S.; et al. Spatial and Temporal Action of Chicken Primordial Germ Cells during Initial Migration. Reproduction 2015, 149, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer Card, D.A.; Hebbar, P.B.; Li, L.; Trotter, K.W.; Komatsu, Y.; Mishina, Y.; Archer, T.K. Oct4/Sox2-Regulated MiR-302 Targets Cyclin D1 in Human Embryonic Stem Cells. Mol. Cell. Biol. 2008, 28, 6426–6438. [Google Scholar] [CrossRef] [Green Version]
- Orkin, S.H.; Hochedlinger, K. Chromatin Connections to Pluripotency and Cellular Reprogramming. Cell 2011, 145, 835–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Fink, S.L.; Cookson, B.T. Apoptosis, Pyroptosis, and Necrosis: Mechanistic Description of Dead and Dying Eukaryotic Cells. Infect. Immun. 2005, 73, 1907–1916. [Google Scholar] [CrossRef] [Green Version]
- Srihawong, T.; Kuwana, T.; Siripattarapravat, K.; Tirawattanawanich, C. Chicken Primordial Germ Cell Motility in Response to Stem Cell Factor Sensing. Int. J. Dev. Biol. 2015, 59, 453–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Gan, L.; Si, J.; Zhang, J.; Liu, Z.; Zhao, J.; Gou, Z.; Zhang, H. Role of MiR-302/367 Cluster in Human Physiology and Pathophysiology. Acta Biochim. Biophys. Sin. 2020, 52, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Zhao, Z.; Wang, Z.; Li, Q.; Wang, X.; Wang, W.; Song, T.; Huang, C. MicroRNA-302b-3p Suppresses Cell Proliferation Through AKT Pathway by Targeting IGF-1R in Human Gastric Cancer. Cell. Physiol. Biochem. 2017, 42, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; He, Y.; Mcleod, H.L.; Xie, Y.; Xiao, D.; Hu, H.; Chen, P. MiR-302b Inhibits Tumorigenesis by Targeting EphA2 via Wnt/β-Catenin/EMT Signaling Cascade in Gastric Cancer. BMC Cancer 2017, 17, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Full Name (Organism) | NCBI Number | Primers | Length of the Product (bp) | |
|---|---|---|---|---|---|
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase (Gallus gallus) | NM_204305.1 | FW | GACGTGCAGCAGGAACACTA | 112 |
| RV | CTTGGACTTTGCCAGAGAGG | ||||
| POUV | POU domain class 5 transcription factor 3 (Pou5f3) (Gallus gallus) | NM_001110178.1 | FW | GAGGCAGAGAACACGGACAA | 109 |
| RV | TTCCCTTCACGTTGGTCTCG | ||||
| CVH | DEAD-box helicase 4 (DDX4) (Gallus gallus) | NM_204708.1 | FW | GAACCTACCATCCACCAGCA | 113 |
| RV | ATGCTACCGAAGTTGCCACA | ||||
| Name | Gene | Accession Number | Assay ID | Sequence |
|---|---|---|---|---|
| miR-92 | hsa-miR-92 | MI0000719 | 000430 | UAUUGCACUUGUCCCGGCCUG |
| gga-miR-302b-3P | hsa-miR-302b | MI0000772 | 000531 | UAAGUGCUUCCAUGUUUUAGUAG |
| gga-miR-302b-5P | gga-miR-302b* | MI0003700 | 008131_mat | ACUUUAACAUGGAGGUGCUUUCU |
| Name | Gene | Catalog Number | Assay ID | Type of Inhibitor |
|---|---|---|---|---|
| anti-miR-302b-3p | hsa-miR-302b-3P | AM17000 | AM10081 | anti-miRTM-miRNA inhibitor |
| anti-miR-302b-5p | gga-miR-302b-5P | 4464084 | MH11349 | mirVanaTM miRNA Inhibitor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lázár, B.; Szabadi, N.T.; Anand, M.; Tóth, R.; Ecker, A.; Urbán, M.; Aponte, M.T.S.; Stepanova, G.; Hegyi, Z.; Homolya, L.; et al. Effect of miR-302b MicroRNA Inhibition on Chicken Primordial Germ Cell Proliferation and Apoptosis Rate. Genes 2022, 13, 82. https://doi.org/10.3390/genes13010082
Lázár B, Szabadi NT, Anand M, Tóth R, Ecker A, Urbán M, Aponte MTS, Stepanova G, Hegyi Z, Homolya L, et al. Effect of miR-302b MicroRNA Inhibition on Chicken Primordial Germ Cell Proliferation and Apoptosis Rate. Genes. 2022; 13(1):82. https://doi.org/10.3390/genes13010082
Chicago/Turabian StyleLázár, Bence, Nikolett Tokodyné Szabadi, Mahek Anand, Roland Tóth, András Ecker, Martin Urbán, Maria Teresa Salinas Aponte, Ganna Stepanova, Zoltán Hegyi, László Homolya, and et al. 2022. "Effect of miR-302b MicroRNA Inhibition on Chicken Primordial Germ Cell Proliferation and Apoptosis Rate" Genes 13, no. 1: 82. https://doi.org/10.3390/genes13010082