
A Cold-Shock Protein from the South Pole-Dwelling Soil Bacterium Arthrobacter sp. Confers Cold Tolerance to Rice

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Determination of Relative Copy Numbers via TaqMan Real-Time Polymerase Chain Reaction (PCR)
2.3. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR Analysis
2.4. Cold Treatment of Rice Plants
2.5. Chlorophyll Measurements
2.6. RNA Sequencing (RNA-seq) and Data Processing
3. Results
3.1. Expression of 35S:ArCspA Transgenic Rice
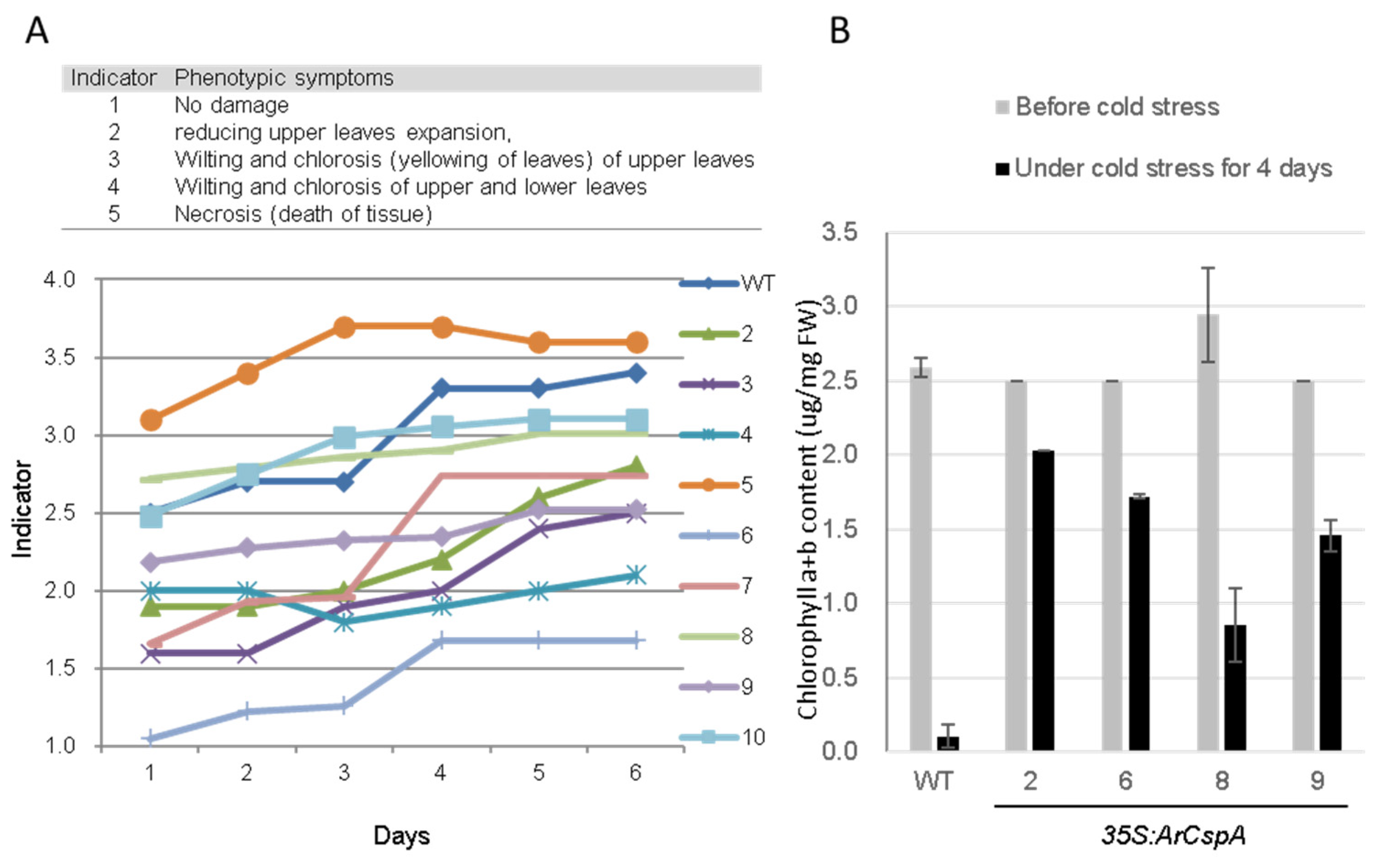
3.2. Effects of ArCspA Overexpression under Cold-Stress Conditions
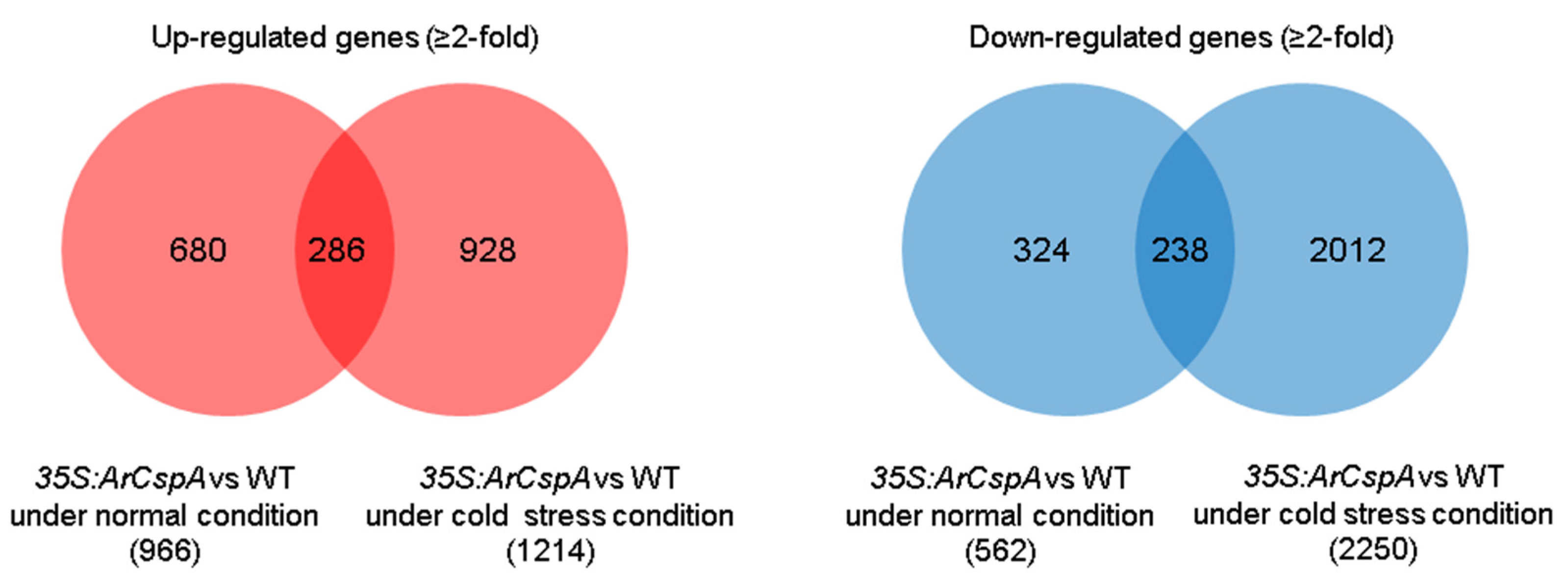
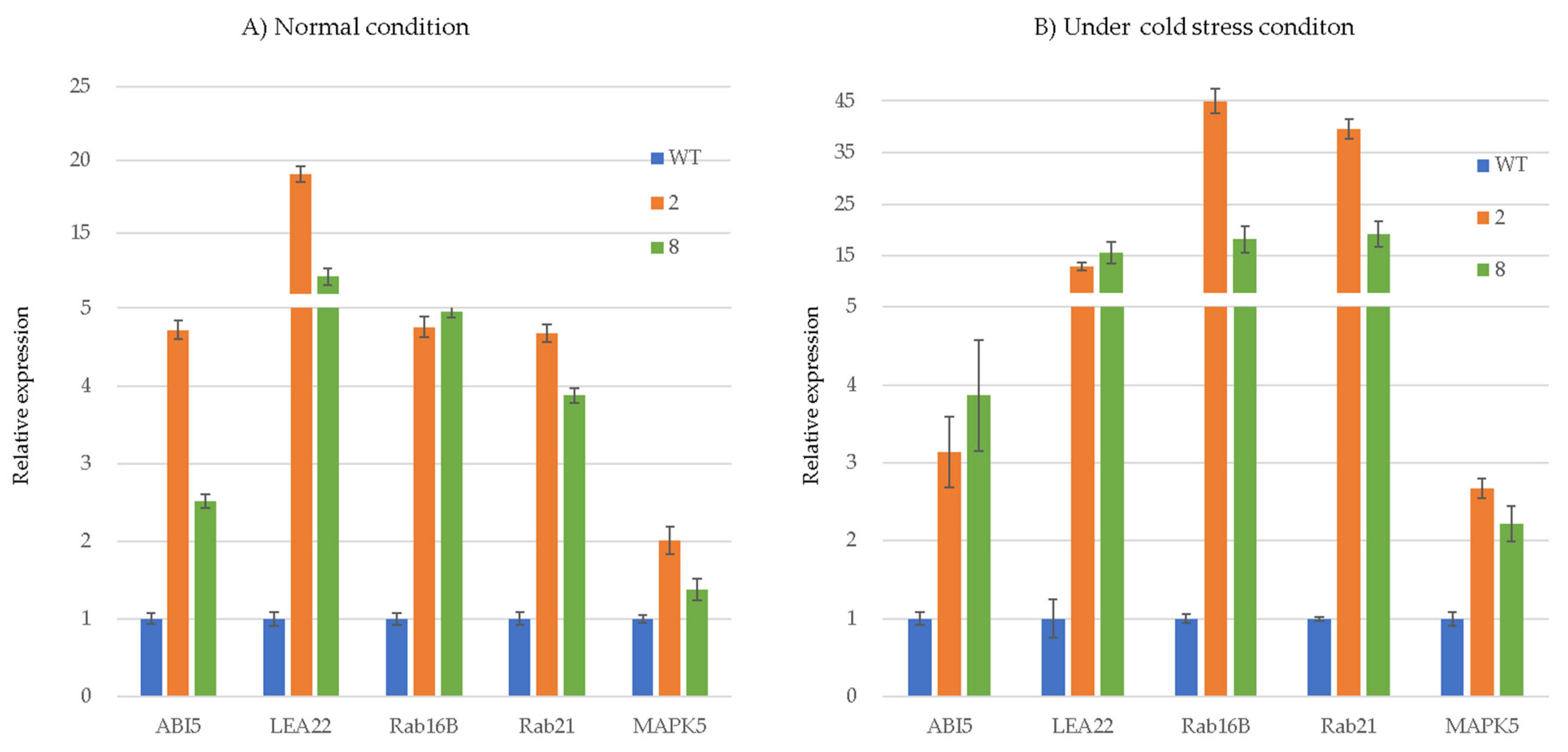
3.3. Transcriptome Analysis of 35S:ArCspA Transgenic Plants under Normal and Cold-Stress Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef]
- Molina, J.; Sikora, M.; Garud, N.; Flowers, J.M.; Rubinstein, S.; Reynolds, A.; Huang, P.; Jackson, S.; Schaal, B.A.; Bustamante, C.D.; et al. Molecular evidence for a single evolutionary origin of domesticated rice. Proc. Natl. Acad. Sci. USA 2011, 108, 8351–8356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahamed, A.; Murai-Hatano, M.; Ishikawa-Sakurai, J.; Hayashi, H.; Kawamura, Y.; Uemura, M. Cold stress-induced acclimation in rice is mediated by root-specific aquaporins. Plant Cell Physiol. 2012, 53, 1445–1456. [Google Scholar] [CrossRef]
- Pareek, A.; Khurana, A.K.; Sharma, A.; Kumar, R. An overview of signaling regulons during cold stress tolerance in plants. Curr. Genom. 2017, 18, 498–511. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajashekar, C.B.; Zhou, H.E.; Zhang, Y.; Li, W.; Wang, X. Suppression of phospholipase Dalpha1 induces freezing tolerance in Arabidopsis: Response of cold-responsive genes and osmolyte accumulation. J. Plant Physiol. 2006, 163, 916–926. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Renhu, N.; Naito, M.; Nakamura, A.; Shiba, H.; Yamamoto, T.; Suzaki, T.; Iida, H.; Miura, K. Ca(2+)-permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca(2+) increase and cold tolerance in Arabidopsis. Sci. Rep. 2018, 8, 550. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Dai, X.; Xu, Y.; Luo, W.; Zheng, X.; Zeng, D.; Pan, Y.; Lin, X.; Liu, H.; Zhang, D. COLD1 confers chilling tolerance in rice. Cell 2015, 160, 1209–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laohavisit, A.; Davies, J.M. Annexins. In Coding and Decoding of Calcium Signals in Plants; Springer: Berlin/Heidelberg, Germany, 2011; pp. 111–128. [Google Scholar]
- Morgan, R.O.; Martin-Almedina, S.; Garcia, M.; Jhoncon-Kooyip, J.; Fernandez, M.P. Deciphering function and mechanism of calcium-binding proteins from their evolutionary imprints. Biochim. Biophys. Acta 2006, 1763, 1238–1249. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Ding, Y.; Shi, Y.; Zhang, X.; Gong, Z.; Yang, S. The cbfs triple mutants reveal the essential functions of CBF s in cold acclimation and allow the definition of CBF regulons in Arabidopsis. New Phytol. 2016, 212, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Kidokoro, S.; Yoneda, K.; Takasaki, H.; Takahashi, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Different cold-signaling pathways function in the responses to rapid and gradual decreases in temperature. Plant Cell 2017, 29, 760–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought-and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar] [PubMed] [Green Version]
- Artus, N.N.; Uemura, M.; Steponkus, P.L.; Gilmour, S.J.; Lin, C.; Thomashow, M.F. Constitutive expression of the cold-regulated Arabidopsis thaliana COR15a gene affects both chloroplast and protoplast freezing tolerance. Proc. Natl. Acad. Sci. USA 1996, 93, 13404–13409. [Google Scholar] [CrossRef] [Green Version]
- Stockinger, E.J.; Gilmour, S.J.; Thomashow, M.F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 1997, 94, 1035–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.G.; Inouye, M. The cold-shock response—A hot topic. Mol. Microbiol. 1994, 11, 811–818. [Google Scholar] [CrossRef]
- Ng, H.; Ingraham, J.L.; Marr, A.G. Damage and derepression in Escherichia coli resulting from growth at low temperatures. J. Bacteriol. 1962, 84, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, J.A.; Mertens, P.R. Cold shock proteins: From cellular mechanisms to pathophysiology and disease. Cell Commun. Signal. 2018, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Brandi, A.; Pietroni, P.; Gualerzi, C.O.; Pon, C.L. Post-transcriptional regulation of CspA expression in Escherichia coli. Mol. Microbiol 1996, 19, 231–240. [Google Scholar] [CrossRef]
- Horn, G.; Hofweber, R.; Kremer, W.; Kalbitzer, H.R. Structure and function of bacterial cold shock proteins. Cell. Mol. Life Sci. 2007, 64, 1457–1470. [Google Scholar] [CrossRef]
- Phadtare, S.; Severinov, K. RNA remodeling and gene regulation by cold shock proteins. RNA Biol. 2010, 7, 788–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.-F.; Xu, Z.-S.; Guo, J.-K.; Wang, Y.-X.; Abernathy, B.; Fu, J.-D.; Chen, X.; Zhou, Y.-B.; Chen, M.; Ye, X.-G. Improved drought tolerance in wheat plants overexpressing a synthetic bacterial cold shock protein gene SeCspA. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-H.; Sasaki, K.; Imai, R. Cold shock domain protein 3 regulates freezing tolerance in Arabidopsis thaliana. J. Biol. Chem. 2009, 284, 23454–23460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusaro, A.F.; Bocca, S.N.; Ramos, R.L.B.; Barrôco, R.M.; Magioli, C.; Jorge, V.C.; Coutinho, T.C.; Rangel-Lima, C.M.; De Rycke, R.; Inzé, D. AtGRP2, a cold-induced nucleo-cytoplasmic RNA-binding protein, has a role in flower and seed development. Planta 2007, 225, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Nakaminami, K.; Karlson, D.T.; Imai, R. Functional conservation of cold shock domains in bacteria and higher plants. Proc. Natl. Acad. Sci. USA 2006, 103, 10122–10127. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Li, Q.H.; Deng, F.; Yu, Z.W.; Luo, X.Z.; Sun, J.L. Synovial fluid concentrations of cold-inducible RNA-binding protein are associated with severity in knee osteoarthritis. Clin. Chim. Acta 2017, 464, 44–49. [Google Scholar] [CrossRef]
- Lee, S.-K.; Park, S.-H.; Lee, J.-W.; Lim, H.-M.; Jung, S.-Y.; Park, I.-C.; Park, S.-C. A putative cold shock protein-encoding gene isolated from Arthrobacter sp. A2-5 confers cold stress tolerance in yeast and plants. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 775–782. [Google Scholar] [CrossRef]
- Lim, H.; Hwang, H.-J.; Kim, A.-R.; Cho, M.-H.; Ji, H.; Kim, C.-K.; Ji, S.U.; Cho, J.-I.; Park, S.-C.; Lee, G.-S. A simple, rapid and systematic method for the developed GM rice analysis. Plant Biotechnol. Rep. 2016, 10, 25–33. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, J.; Lee, T.-H.; Jun, K.M.; Kim, T.H.; Kim, Y.-H.; Park, H.-M.; Jeon, J.-S.; An, G.; Yoon, U.-H. FSTVAL: A new web tool to validate bulk flanking sequence tags. Plant Methods 2012, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-L.; Lee, K.; Cho, W.; Park, D.; Lee, I.H.; Lim, H. Genetic Diversity and Physiological Response to Drought Stress of Chamaecyparis obtuse from Six Geographical Locations. Plant Breed Biotechnol. 2021, 9, 112–123. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Kishor Pb, K. Bacterial Cold Shock Proteins - the Molecular Chaperones for Multiple Stress Tolerance. Adv. Biotechnol. Microbiol. 2019, 12. [Google Scholar] [CrossRef]
- Janská, A.; Maršík, P.; Zelenková, S.; Ovesná, J. Cold stress and acclimation–what is important for metabolic adjustment? Plant Biol. 2010, 12, 395–405. [Google Scholar] [CrossRef]
- Seo, M.; Koshiba, T. Complex regulation of ABA biosynthesis in plants. Trends Plant Sci. 2002, 7, 41–48. [Google Scholar] [CrossRef]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Mundy, J.; Chua, N.-H. Abscisic acid and water-stress induce the expression of a novel rice gene. EMBO J. 1988, 7, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Mundy, J.; Chua, N.-H. Four tightly linked rab genes are differentially expressed in rice. Plant Mol. Biol. 1990, 14, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C phosphatase activity function as abscisic acid sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.; Rodrigues, A.; Saez, A.; Rubio, S.; Antoni, R.; Dupeux, F.; Park, S.Y.; Márquez, J.A.; Cutler, S.R.; Rodriguez, P.L. Modulation of drought resistance by the abscisic acid receptor PYL5 through inhibition of clade A PP2Cs. Plant J. 2009, 60, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hwang, H.; Hong, J.-W.; Lee, Y.-N.; Ahn, I.P.; Yoon, I.S.; Yoo, S.-D.; Lee, S.; Lee, S.C.; Kim, B.-G. A rice orthologue of the ABA receptor, OsPYL/RCAR5, is a positive regulator of the ABA signal transduction pathway in seed germination and early seedling growth. J. Exp. Bot. 2012, 63, 1013–1024. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.; Guan, Y.; Ren, H.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 66, 675–683. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.-N.; Xue, L.-J.; Zou, M.-J.; Liu, J.-Y.; Chen, F.; Xue, H.-W. Rice ABI5-Like1 regulates abscisic acid and auxin responses by affecting the expression of ABRE-containing genes. J. Plant Physiol. 2011, 156, 1397–1409. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Chen, J.; Wang, Q.; Yang, Y. Direct phosphorylation and activation of a mitogen-activated protein kinase by a calcium-dependent protein kinase in rice. Plant Cell 2014, 26, 3077–3089. [Google Scholar] [CrossRef] [Green Version]
- Xie, G.; Kato, H.; Imai, R. Biochemical identification of the OsMKK6–OsMPK3 signalling pathway for chilling stress tolerance in rice. Biochem. J. 2012, 443, 95–102. [Google Scholar] [CrossRef]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998, 280, 104–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Lee, C.M.; Doherty, C.J.; Gilmour, S.J.; Kim, Y.; Thomashow, M.F. Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network. Plant J. 2015, 82, 193–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.T.; Zarka, D.G.; Van Buskirk, H.A.; Fowler, S.G.; Thomashow, M.F. Roles of the CBF2 and ZAT12 transcription factors in configuring the low temperature transcriptome of Arabidopsis. Plant J. 2005, 41, 195–211. [Google Scholar] [CrossRef]
- Cui, L.H.; Byun, M.Y.; Oh, H.G.; Kim, S.J.; Lee, J.; Park, H.; Lee, H.; Kim, W.T. Poaceae Type II galactinol synthase 2 from antarctic flowering plant Deschampsia antarctica and rice improves cold and drought tolerance by accumulation of raffinose family oligosaccharides in transgenic rice plants. Plant Cell Physiol. 2020, 61, 88–104. [Google Scholar] [CrossRef]
- Fang, J.-C.; Tsai, Y.-C.; Chou, W.-L.; Liu, H.-Y.; Chang, C.-C.; Wu, S.-J.; Lu, C.-A. A CCR4-associated factor 1, OsCAF1B, confers tolerance of low-temperature stress to rice seedlings. Plant Mol. Biol. 2021, 105, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Chaikam, V.; Karlson, D.T. Comparison of structure, function and regulation of plant cold shock domain proteins to bacterial and animal cold shock domain proteins. BMB Rep. 2010, 43, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Teana, A.; Brandi, A.; Falconi, M.; Spurio, R.; Pon, C.L.; Gualerzi, C. Identification of a cold shock transcriptional enhancer of the Escherichia coli gene encoding nucleoid protein H-NS. Proc. Natl. Acad. Sci. USA 1991, 88, 10907–10911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landsman, D. RNP-1, an RNA-binding motif is conserved in the DNA-binding cold shock domain. Nucleic Acids Res. 1992, 20, 2861–2864. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Go Function | No. Genes | No. Genes | p-Value | |
|---|---|---|---|---|
| (Reflist) | (Up- or Downregulated) | |||
| Upregulated genes in 35S:ArCspA/WT under normal conditions | ||||
| Response to stress | GO:0006950 | 2115 | 113 | 1.47 × 10−17 |
| Response to cold | GO:0009409 | 92 | 12 | 2.31 × 10−6 |
| Response to heat | GO:0009408 | 102 | 13 | 1.12 × 10−6 |
| Response to hormone | GO:0009725 | 589 | 31 | 1.48 × 10−5 |
| Cellular nitrogen compound metabolic process | GO:0034641 | 3075 | 38 | 1.36 × 10−4 |
| Cellular component organization or biogenesis | GO:0071840 | 2335 | 24 | 3.62 × 10−5 |
| Detoxification | GO:0098754 | 269 | 16 | 4.56 × 10−4 |
| Gene expression | GO:0010467 | 1708 | 16 | 1.60 × 10−4 |
| Upregulated genes in 35S:ArCspA/WT under cold-stress conditions | ||||
| Gene expression | GO:0010467 | 1708 | 18 | 2.66 × 10−6 |
| Downregulated genes in 35S:ArCspA/WT under cold-stress conditions | ||||
| Cell cycle | GO:0007049 | 410 | 45 | 7.46 × 10−6 |
| Cell wall | GO:0005618 | 326 | 31 | 1.74 × 10−3 |
| Chloroplast | GO:0009507 | 1414 | 30 | 4.68 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.Y.; Kim, J.S.; Cho, W.; Jun, K.M.; Du, X.; Kim, K.D.; Kim, Y.-K.; Lee, G.-S. A Cold-Shock Protein from the South Pole-Dwelling Soil Bacterium Arthrobacter sp. Confers Cold Tolerance to Rice. Genes 2021, 12, 1589. https://doi.org/10.3390/genes12101589
Kim SY, Kim JS, Cho W, Jun KM, Du X, Kim KD, Kim Y-K, Lee G-S. A Cold-Shock Protein from the South Pole-Dwelling Soil Bacterium Arthrobacter sp. Confers Cold Tolerance to Rice. Genes. 2021; 12(10):1589. https://doi.org/10.3390/genes12101589
Chicago/Turabian StyleKim, So Young, Joung Sug Kim, Woosuk Cho, Kyong Mi Jun, Xiaoxuan Du, Kyung Do Kim, Yeon-Ki Kim, and Gang-Seob Lee. 2021. "A Cold-Shock Protein from the South Pole-Dwelling Soil Bacterium Arthrobacter sp. Confers Cold Tolerance to Rice" Genes 12, no. 10: 1589. https://doi.org/10.3390/genes12101589