
Genetics of Diabetic Retinopathy, a Leading Cause of Irreversible Blindness in the Industrialized World


1
Department of Ophthalmology, Eugene and Marilyn Glick Eye Institute, Indiana University, Indianapolis, IN 46202, USA
2
Department of Internal Medicine, Ascension St. Vincent Hospital, Indianapolis, IN 46260, USA
3
Clearside Biomedical, Inc., Alpharetta, GA 30005, USA
4
Retina Service, Midwest Eye Institute, Indianapolis, IN 46290, USA
*
Author to whom correspondence should be addressed.
Genes 2021, 12(8), 1200; https://doi.org/10.3390/genes12081200
Submission received: 11 January 2021
/
Revised: 29 May 2021
/
Accepted: 29 July 2021
/
Published: 31 July 2021
(This article belongs to the Section Human Genomics and Genetic Diseases)
Abstract
:Diabetic retinopathy (DR) is a chronic complication of diabetes and a leading cause of blindness in the industrialized world. Traditional risk factors, such as glycemic control and duration of diabetes, are unable to explain why some individuals remain protected while others progress to a more severe form of the disease. Differences are also observed in DR heritability as well as the response to anti-vascular endothelial growth factor (VEGF) treatment. This review discusses various aspects of genetics in DR to shed light on DR pathogenesis and treatment. First, we discuss the global burden of DR followed by a discussion on disease pathogenesis as well as the role genetics plays in the prevalence and progression of DR. Subsequently, we provide a review of studies related to DR’s genetic contribution, such as candidate gene studies, linkage studies, and genome-wide association studies (GWAS) as well as other clinical and meta-analysis studies that have identified putative candidate genes. With the advent of newer cutting-edge technologies, identifying the genetic components in DR has played an important role in understanding DR incidence, progression, and response to treatment, thereby developing newer therapeutic targets and therapies.
1. Background
Diabetic retinopathy (DR) and diabetic macular edema (DME) are the leading causes of blindness among diabetic individuals. Based on clinical characteristics, DR is divided into two broad categories: non-proliferative DR (NPDR) and proliferative DR (PDR). NPDR is an early stage of DR associated with an increase in vascular permeability and capillary occlusion. At this stage, the fundus examination exhibits pathologies such as microaneurysms, hemorrhages, and hard lipid exudates. PDR is a more advanced form of DR characterized by new and abnormal vessels that can grow into the vitreous scaffold and then shear with ensuing vitreous hemorrhage and abrupt loss of vision, especially after vitreous traction. Ultimately, this neovascularization can fibrose on the inner retinal surface and contract, leading to tractional retinal detachment. Another complication of DR is DME, which can occur at any stage and is categorized by thickening of the macula due to permeability-mediated sub-and intra-retinal accumulation of fluid. DME causes decreased visual acuity and central visual distortion (metamorphopsia) [1,2].
From 2015–2019, the prevalence for DR was 27% among patients with diabetes worldwide, including 25.2% for NPDR and 1.4% PDR, and DME was diagnosed in 4.6% of diabetic patients. The highest prevalence was reported in the Western Pacific region (36.2%), while other areas showed variation in its incidence: the Middle East and North Africa (33.8%), Europe (20.6%), and Asia (12.5%) [3]. It is estimated that DR is present in most subjects with type 1 diabetes (T1D) and nearly 60% of individuals with type 2 diabetes (T2D) after 20 years of living with diabetes. In 2010, an estimated 7.7 million Americans were diagnosed with DR, which is expected to increase to 14.6 million by 2050 [4]. Hispanic Americans are at higher risk of developing DR with a prevalence of 8%, and black and white races show about 5% prevalence. Gender also plays some role in that more males are affected by DR than females in the USA [5].
2. Pathogenesis of DR
Numerous changes occur in the retinal vasculature with DR long before fundus examination reveals any pathological changes. Long-standing diabetes leads to an early and progressive loss of retinal pericytes and endothelial cells, resulting in microaneurysms [6]. Furthermore, increased leukostasis and platelet-fibrin thrombi changes in the retinal capillaries contribute substantially to retinal ischemia and vascular obstruction. Resultant chronic hypoxia triggers angiogenic growth factors such as vascular endothelial growth factor (VEGF) that ultimately contribute to the retinal neovascularization associated with PDR [7].
Hyperglycemia and chronic inflammation predominantly drive the onset and pathogenesis of DR (Figure 1). Some studies suggest more rapid DR progression with fluctuation in glucose levels between hyper and normoglycemia in individuals with poorly controlled diabetes [8]. Various mechanisms play an integral role behind the capillary damage observed in DR, including (i) increase in polyol pathway, (ii) activation of protein kinase C, (iii) accelerated increase in non-enzymatic glycation and (iv) production of reactive oxygen species (ROS). The activation of the polyol pathway results in a loss of retinal capillary cells. An antioxidant defense system that protects the cell and tissue from oxidative stress is downregulated, specifically the enzymes superoxide dismutase (SOD) and glutathione peroxidase (GSH). An accelerated increase in advanced glycation end products (AGEs) may provoke sustained cell activation, basement membrane damage, and basement membrane thickening, further exacerbating damage to the retinal tissue.
Changes in retinal blood flow patterns also play a vital role in DR pathogenesis, such as (i) increased retinal blood flow and (ii) increased heterogeneity in its distribution [9,10]. Typically, blood flow control is balanced between endothelium-derived relaxing factors, such as nitric oxide (NO), prostacyclin, endothelium-derived hyperpolarizing factor (EDHF), and contracting factors angiotensin II, endothelin, and cyclooxygenase. The interplay of these factors plays a vital role in DR by regulating vascular tone via the stimulation or inhibition of smooth muscle cells and pericytes [11]. Notably, NO is a potent vasodilator, causing smooth muscle relaxation, and hyperglycemia results in either diminished production or quenching of NO by various mechanisms [12]. Prostacyclin (PGI2) works complementary to NO and causes relaxation of smooth muscles through cyclic adenosine monophosphate (cAMP) production, which inhibits calcium-mediated contraction.
Finally, DR is associated with increased VEGF levels in the vitreous as well as the retina. VEGF plays a vital role in neovascularization, resulting in the progression of NPDR to PDR. VEGF not only stimulates endothelial cell migration and proliferation but also acts as a survival factor for new blood vessels in diabetes [13].
3. The Genetic Basis of DR
3.1. Genetics in the Prevalence of DR
Several studies suggest variations in DR prevalence depending on racial background. Harris et al. (1998) found the prevalence of DR in patients with T2D to be 46% higher in non-Hispanic blacks and 84% higher in Mexican Americans than non-Hispanic whites [14]. Notably, the prevalence of diabetes has been found to be highest among American Indians and Alaskan Natives, followed by Hispanics and non-Hispanic whites [15]. As per the latest study involving 53,998 individuals, the prevalence of DR among American Indians and Alaskan Natives was lower than earlier statistics: NPDR (17.7%), PDR (2.3%), DME (2.3%), and sight-threatening retinopathy (STR) (4.2%) [16]. Furthermore, DR’s prevalence in Pima Indians with T2D has been found to be exceptionally high (37.8%) [17]. Similarly, a higher prevalence of DR has been reported in Navajo and Hopi Indians (40%) and Sioux Indian Tribes (45.3%) [16]. In another study involving Oklahoma Indian tribes, the prevalence of DR was found to be 20.1% [18]. These disparities in the prevalence of DR are independent of blood glucose control [19], suggesting the potential genetic basis for DR pathogenesis.
3.2. Familial Risk of DR
Interestingly, the Diabetes Control and Complications Trial (DCCT) highlighted the familial clustering of DR in the families of 372 subjects with 467 first-degree relatives with T1 and T2D. The risk of severe retinopathy was significantly higher among families of individuals with retinopathy. The correlation of severity of retinopathy was higher among parent-offspring; however, a similar familial correlation was not observed for nephropathy [20], suggesting the influence of an independent genetic risk factor.
The Find-Eye study provides further insights into the inherited factors that cause DR’s susceptibility and severity. This study involved 2368 diabetic subjects from 767 families and showed that the heritability of DR was 27%, with a polygenic heritability of PDR of 25% [21]. Hietala et al. additionally provided evidence that clustering exists for PDR in families with two or more siblings with T1D, demonstrating an increased risk of DR in siblings of probands with DR, after adjusting for glycated hemoglobin, duration of diabetes, and mean blood pressure [22]. Finally, twin studies have also reported the same degree of DR severity among twins with DM, with one early study demonstrating a concordance of 65% among T1D and 95% among T2D twins [23]. Thus, the severity of DR is apparently influenced by familial factors.
3.3. Progression of DR
DR is known to follow the classical course of disease progression from no-DR→NPDR→PDR; however, not every patient progresses through this classical paradigm. For example, some patients develop mild NPDR after 8-10 years of no DR. Of these individuals, only 50% of those with T1D develop PDR, and 20% with type 2 diabetes develop PDR. After four years of moderate to severe NPDR, the incidence of PDR is 11%, and the incidence of severe vision loss is 7.2% [9]. It is unclear why only a certain percentage of people develop PDR while the remaining are protected from DR despite a longer duration of diabetes. Previously, we identified a similar population of individuals who remained protected from DR in spite of long-standing diabetes. We performed microarray studies on circulating angiogenic cells (CACs) obtained from this population to demonstrate mapping with unique targets such as ataxia telangiectasia mutated (ATM) [24], transforming growth factor (TGFB1), plasminogen activator inhibitor (PAI-1; gene SERPINE1), endothelial nitric oxide synthase (eNOS; gene NOS3), and angiotensin-converting enzyme 2 (ACE2) [25]. Our study likely suggests that genetic mechanisms play a role in DR progression.
4. Genetic Studies
4.1. Candidate Gene Studies
Candidate gene studies are clinical and preclinical studies where a gene is identified as potentially implicated in disease pathogenesis based on the expression of already identified proteins in the disease state. A 2009 meta-analysis examined 34 genetic variants known to be associated with the pathogenesis of DR and found the aldose reductase (ALR) gene aldo-keto reductase family 1 member B, (AKR1B1) to have the highest number of polymorphisms associated with DR irrespective of ethnicity. Additional polymorphisms reported to be significantly associated with DR included NOS3, VEGFA, integrin subunit alpha 2 (ITGA2), and intercellular adhesion molecule 1(ICAM1) [26]. More recently, Sharma et al. (2019) elegantly summarized candidate gene studies associated with DR in a review highlighting 65 different genes [27]. These candidate genes are involved in a variety of cellular processes, such as glucose metabolism, inflammation, angiogenesis, and neurogenesis. These studies have overall yielded inconsistent and controversial results [28].
The majority of these studies have been performed in T2D; however, some targets were associated with T1D in candidate gene studies, such as peroxisome proliferator-activated receptor gamma (PPARG), neuropeptide Y receptor 2 (NPY2R), centrosomal protein (CEP)-135 (CEP135), and plexin domain containing 2 (PLXDC2). The following sections focus on those genes that have been found to be associated with DR based on multiple independent studies. Additionally, Table 1 summarizes the findings of these studies.
4.1.1. Aldose Reductase (ALR)
Aldose reductase has been significantly implicated in the pathogenesis of DR via the polyol pathway, by which excess intracellular glucose in retinal cells is converted to fructose [46]. The ALR enzyme catalyzes the initial, rate-limiting step of the pathway, which consists of NADPH-mediated glucose reduction into sorbitol. Sorbitol cannot diffuse freely from the cell cytoplasm; while initially it was suggested that sorbitol accumulation leads to osmotic stress [47], the amount of sorbitol measured in diabetic vessels and nerves was found to be insufficient to cause osmotic stress [48]. Furthermore, increased ALR and polyol pathway activity has also been shown to be significantly associated with pericyte degeneration, a key feature of DR, leading to abnormalities of vascular permeability and compromise of the blood-retinal barrier. In turn, breakdown of the blood-retinal barrier is thought to be a major etiology for the development of exudates and macular edema in DR [49].
The AKR1B1 gene is located on chromosome 7q33 and is expressed in retinal capillary pericytes to produce ALR. The three most reported AKR1B1 polymorphisms associated with DR include (i) rs759853 (106C/T) [50], (ii) 5′-(CA)n microsatellite (5′AKR1B1) [51], and (iii) rs9640883 [26,52]; however, studies of the association between these SNPs and DR have frequently resulted in inconsistent and contradictory findings.
Several studies of the rs759853 single nucleotide polymorphisms (SNPs) have demonstrated that the C allele is significantly associated with increased risk of DR, while the T allele may be protective [26,29,50,53,54,55,56,57,58,59]. This finding was replicated by a recent 2020 meta-analysis of 23 studies [60]. Interestingly, two recent studies of T2D in Eastern European and Arab populations demonstrated the opposite relationship, suggesting that the T allele is associated with increased risk and severity of DM, while the C allele is protective [61,62]. Finally, some studies have found no such association [63,64,65,66], including a 2018 meta-analysis [29]. It is possible that some of the variation in these findings is due to the racial heterogeneity of the studies in question; however, when stratifying by type of diabetes, the T allele of the SNP has been found to be protective against the development of DR in T1D regardless of the ethnic background [26,29]; therefore, differences in the pathogenesis of DR in T1D compared to T2D may also account for the heterogeneity of these findings.
Microsatellites are tandem repeats of 1–6 bp that are widely spread throughout the genome. The (CA)n microsatellite has also been studied in relation to DR pathogenesis, with similarly diverse findings. The three most commonly studied alleles of this SNP in DR are z, z+2, and z-2. Some studies have suggested that the z-2 allele is associated with increased ALR activity [67] and DR pathogenesis in both T2D (stronger association) and T1D (weaker association) patients [50,53,68,69]. Two large meta-analyses have confirmed these findings, and both additionally found that the z+2 allele may be significantly protective against DR development in T2D [26,70]. Notably, however, Abhary et al. (2010) found no significant association between any (CA)n allele and T2D once established risk factors were controlled for [30], suggesting that the (CA)n microsatellite may be only secondarily associated with DR through other clinical risk factors, rather than playing a primary role in its pathogenesis.
Further, the rs9640883 has also been associated with DR pathogenesis. Specifically, the C allele has been implicated in an increased risk of DR, while the T allele has been associated with decreased risk [30,57,58,61]; however, while Abhary et al. (2010) reported a significant association of rs9640883 with DR in general, they did not find this association to be significant after adjusting relevant clinical factors except for the duration of diabetes. The authors, therefore, suggest that the association of the rs9640883 allele with DR may be attributed largely to younger age at onset of diabetes [30], suggesting that this allele may be associated with the early development of diabetes as a confounding variable, rather than affecting the pathogenesis DR itself.
Other AKR1B1 SNPs that have been studied in DR include rs5053, rs918825, and rs706207; however, these were not found to be significantly associated with DR in a recent meta-analysis [71].
4.1.2. Receptor for Advanced Glycation End Products (RAGE)
A major consequence of hyperglycemia is excessive non-enzymatic glycation of proteins and other macromolecules. French chemist L.C. Maillard first reported this reaction in 1912. In the Maillard reaction reducing sugars such as glucose react with an amino group of proteins to form irreversibly modified proteins called advanced glycation end products (AGEs) [72]. AGEs accumulate both intracellularly and extracellularly and may significantly modify proteins, thereby altering function [73]. Diabetes enhances AGE accumulation in the retina’s vascular and neural components throughout life [74]. AGEs could affect the physiological function of pericytes [75] and may even lead to cell death [76]. The receptor for AGE (RAGE) was first observed on the endothelial cell surface [77] and among one of the well characterized receptors of AGEs. AGE interactions with RAGE trigger pro-inflammatory cascades, such as P21 (Ras)/MAP kinase, and AGE is associated with decreased production of prostacyclin (PGI2) and an increase in platelet aggregation and fibrin stabilization [78]. AGE binding to RAGE receptors upregulates cyclooxygenase-2 (COX), resulting in monocyte activation and vascular cell dysfunction [79], linked to oxidative stress and NF-κB activation [80,81]. Finally, AGEs have been shown to stimulate endothelial cell proliferation and angiogenesis by upregulation of VEGF and via the RhoA/Rho-associated protein kinase pathway [82,83].
Several polymorphisms in the gene encoding RAGE (AGER) have been identified as potentially associated with DR: rs1800625 (429T/C), rs1800624 (374T/A), rs2070600 (Gly82Ser), rs184003 (1704G/T), and rs3134940 (2184A/G). The rs1800624 and rs1800625 SNPs are perhaps the most widely studied. These were initially identified by Hudson et al. (2001), who found the C allele of the rs1800624 SNP to be significantly associated with DR in D2M subjects, while the A allele of the rs1800624 SNP was elevated, but not statistically significantly associated with DR. The association between the rs1800624 SNP has since been replicated in Indonesian subjects [84]; however, this relationship was not found in a 2012 meta-analysis by Yuan et al. [85].
Lindholm et al. (2006) further studied the rs1800624 SNP in Scandinavian T1D and T2D patients. The rs1800624 polymorphism was found to be related to different major histocompatibility complex, class II, DQ beta 1 (HLA-DQB1) genotypes and associated with the presence of DR in Scandinavian individuals, with a higher frequency of the A allele in T1D than T2D individuals with DR [31]. Similar findings have been more recently replicated in Pakistani individuals [86]. Furthermore, two recent meta-analyses (2016 and 2017) have provided additional evidence that the A allele of the rs1800624 polymorphism is associated with significantly increased risk of DR in both Asian and Caucasian diabetic populations [87,88], though in the case of Yu et al. 2016, this association was not sustained after T2D patients were excluded from the analysis. Conversely, two earlier meta-analyses (2010 and 2012) suggested a protective effect of the A allele in Caucasian patients with D2M [85,89]. Finally, no significant association was observed between either the rs1800624 or rs1800625 polymorphisms in a study of Malaysian patients [90] or Indian patients [32].
The rs2070600 polymorphism was also assessed in an Indian population with DR. The AGER polymorphism for rs2070600 was found to be significantly associated with DR [32,91]. This finding has been since replicated in Indonesian patients [84] and Han Chinese patients [92]. A 2002 meta-analysis has also validated the association of the Gly82Ser polymorphism with DR in a general Asian population [88]; however, this finding was only supported by three later meta-analyses (2012, 2012, and 2016) after assuming a genetic recessive model [85,88,93]. Finally, the rs184003 RAGE polymorphism has also been reported to be associated with DR in Japanese and East Asian populations [94,95].
While some of the above studies provide encouraging evidence for the association of the AGER gene with DR, other studies have not supported an association between RAGE polymorphisms and DR. As noted above, it is worth emphasizing that two 2012 meta-analyses found no significant association between rs1800624 or rs184003 and DR, despite stratification by ethnicity [85,93]. A subsequent 2016 meta-analysis moreover similarly showed no significant association between rs1800624 and DR [88]. Finally, while one study of rs184003 showed a significant association with DR in Japanese subjects as above, other studies have failed to identify a similar association [96,97]. Other SNPs such as rs3134940 have similarly not been found to be associated with DR pathogenesis [97].
4.1.3. VEGF Gene
VEGF is a potent cytokine known to play an integral role in DR pathogenesis by altering retinal vascular permeability, promoting endothelial dysfunction, and ultimately inducing retinal angiogenesis [13]. VEGF is a part of the growth factor family that includes VEGF-A, VEGF-B, VEGF-C, VEGF-D, VEGF-E, VEGF, and placental growth factor (PGF). VEGF receptor 2 (VEGFR2), also known as KDR/Flk-1, mediates vascular leakage and angiogenesis. While VEGF-A binds to VEGFR2 and VEGFR1 (Flt-1), VEGF-B and PGF binding are more specific to VEGFR1. VEGF-C and VEGF-D bind to VEGFR2 and VEGFR3 (Flt4) [98]. Recent meta-analyses have demonstrated that serum VEGF levels are significantly associated with both the presence and severity of DR in diabetic patients [86,99]. Additionally, intravitreal (IVT) injection of anti-VEGF-A is the first-line treatment for DME. Currently available anti-VEGFA medications for DME include ranibizumab, off-label bevacizumab, and aflibercept, the latter of which also binds VEGF-B and PGF [1,100].
The human VEGFA gene is present on chromosome 6 (6p21.1) encodes eight exons, while other human VEGF genes encode seven exons. Alternative splicing of the VEGF gene results in at least six transcripts containing exons 1-5 and 8. Additionally, diversity is generated via alternative splicing of exons 6 and 7. VEGF gene is highly polymorphic [101], and these polymorphisms are mainly located in the 5′ untranslated (UTR) region and in the 3′ UTR region [102]. VEGF polymorphism of rs699947, rs3025039, and rs833061 has been associated with the risk of NPDR among Asians, and rs2010963) has been associated with the risk of NPDR among Caucasians [33,103]. Moreover, rs69947, rs2010963, and rs3025039 have specifically been shown to increase VEGF mRNA and protein levels, driving the pathogenesis of DR [104]. Similarly, the DCCT/Epidemiology of Diabetes Interventions and Complications (EDIC) study discovered SNP for rs3025021 in T1D. Further, the family-based analyses demonstrated excess transmission of C at rs699947, T at rs3025021 and the C-T haplotype for both SNPs [105]. Some of the earlier studies by Abhary et al. assessed multiple tag SNPs for VEGF-A polymorphism, demonstrating a significant association with DR, including the AA genotype of rs699946 and the GG genotype of rs833068 for T1D as well as the C allele of rs3025021 and the G allele of rs10434 for T2D [106].
While there is mounting evidence of VEGFA polymorphism associations in DR, there are also regional and ethnic variations regarding VEGF, VEGFA polymorphisms, and DR. A 2019 meta-analysis of 29 previous studies found serum VEGF levels to be associated with DR in East Asian populations, but not European populations. A recent study by Khan et al. assessed SNPs rs833061, rs13207351, rs1570360, rs2010963, rs5742632, and rs6214 in NPDR, PDR, and control populations of Pakistani origin. This study could only find an association of rs13207351 with NPDR and rs5742632 with PDR [107]. Similarly, studies performed in the Caucasian Slovenian and Mexican PDR subjects did not observe an association between AA genotype of rs6921438 or rs10738760 [108,109] and rs3025021, rs3025035, rs2010963, respectively [110]; however, recent studies have also shown that rs2010963 is associated with the development and severity of DR in Egyptian [111] and Sudanese [112] patients.
Studies of the rs699947 SNP have also yielded varying results across ethnicities. Some studies have demonstrated that the rs699947 SNP is associated with DR in various ethnicities, including Korean [113], Chinese [114], Japanese [115], Pakistani [86], Australian [106], and Spanis [116] populations. However, other studies in Egyptian [117] and Chinese [118] populations have failed to find such an association, including two meta-analyses (2014 and 2015), which analyzed patients pooled from both European and East Asian population studies [104,119]. Thus, while it appears that significant regional and ethnic variations in VEGF polymorphisms and expression may play a confounding role in ethnically heterogeneous studies, the precise nature of these ethnic variations remains unclear.
4.1.4. eNOS Gene
Nitric oxide (NO) is a vasodilatory gas that plays a key role in the regulation of retinal vascular diameter [120]. Endothelial nitric oxide synthase (eNOS; gene NOS3) is the major enzyme responsible for the synthesis of nitric oxide (NO) in human vasculature [121]. eNOS also functions to maintain a protective anti-proliferative and anti-apoptotic environment within the vasculature [122,123]. Knockout eNOS−/− mice have been found to develop retinal endothelial dysfunction [124] and demonstrate accelerated development of retinopathy [125], suggesting that eNOS dysfunction may play a role in the pathogenesis of DR. Moreover, NO has been found to be significantly elevated in the vitreous of patients with proliferative diabetic retinopathy [126].
The eNOS gene is located in chromosome 7q36.1. The polymorphisms of eNOS rs869109213 (4b/a) and rs2070744 (-786T/C) have been found to be associated with DR in Tunisian [34] and Algerian [127] populations. A multi-ethnic meta-analysis by Zhao et al. (2012) also demonstrated a protective effect associated with the C allele of the rs2070744 SNP. Additionally, the 4a/4a genotype of the 27VNTR SNP has also been associated with a lower risk of developing DR [128], suggesting a protective effect of the 4a allele. This finding was replicated by Zhao et al. (2012) across a range of European and Asian ethnicities [129] and again in African patients, specifically by Qian-Qian et al. (2014) [130]. On the other hand, Cilenšek et al. (2012) found that the 4a/4a genotype is instead associated with an increased risk of PDR in Caucasian patients [131]. Lastly, multiple cross-sectional studies and meta-analyses have not found any association between the 27VNTR SNP and DR [132,133]. Most recently, Momeni et al. (2016) have suggested that the 27VNTR SNP may be associated with the development of T2D but not DR itself [134], which may explain some of these disparate findings.
4.1.5. Angiotensin-Converting Enzyme Gene
Angiotensin-I converting enzyme (ACE) is a part of the renin-angiotensin system (RAS) regulating vascular tone. ACE converts angiotensin I to angiotensin II, which in turn causes vasoconstriction. Increased ACE activity, therefore, decreases retinal blood flow and can lead to pathologic angiogenesis [135]. The RAS may be implicated in the mechanism by which VEGF induces vascular permeability [136]; therefore, several studies have suggested that ACE inhibitors and angiotensin receptor blockers may reduce the risk of DR in diabetic patients [137,138,139]. Recently, tears of patients with DR have been found to have elevated ACE and angiotensin II concentration [140], further implicating ACE expression with DR.
The ACE gene is located on the 17q23.3 chromosome. Most of the phenotypic variance in this gene’s expression is due to the intron 16 insertion/deletion (I/D) polymorphism (rs1799752 or also rs4646994) [141]. The I/D polymorphism is the most widely studied in association with DR. In their 2016 meta-analysis, Luo et al. found this polymorphism to be significantly associated with DR [35]; however, other studies have failed to find the association of I/D polymorphism with DR [142,143,144,145]. Additionally, the ACE rs4343 (2350G/A) polymorphism has also been reported to be associated with DR [36].
4.1.6. Erythropoietin Gene
While initially, erythropoietin’s (EPO) role was thought to be limited to the regulation of erythrocyte production in bone marrow, emerging studies demonstrate the pivotal role of EPO in various bodily functions, including the retina. In the early stages of DR, EPO exhibits a neuroprotective, anti-apoptotic, anti-inflammatory, anti-oxidative, and anti-permeability action by protecting retinal pigment epithelium against increased permeability. Although several preclinical and small interventional clinical studies show the potential benefit of intraperitoneal or intravitreal EPO administration in early DR [146], there is some concern regarding the use of EPO treatment in advanced stages of DR because EPO acts synergistically with VEGF, thus promoting angiogenesis. Elevated serum EPO has therefore been associated with the progression of NPDR to PDR [147] and EPO has been reported in abnormally high concentrations in the vitreous of PDR eyes [126].
Erythropoietin gene EPO is located on 7q22.1. There are mixed reports regarding EPO’s polymorphism and DR. While some studies have demonstrated an association between rs551238, rs1617640, and rs507392 and an increased risk of developing PDR [37,38,39], a recent 2020 meta-analysis could not find such an association [148], and an association was only observed between rs551238 and DR in a 2017 meta-analysis study [149].
4.2. Linkage Studies
Genetic linkage analysis is based on the hypothesis that genes residing in close proximity physically on a chromosome endure linkage during meiosis. Two genetic markers in physical proximity to each other are unlikely to be separated during the chromosomal crossover. Thus, upon identification of a chromosomal location of a particular disease phenotype, the genetic analysis further helps to decipher whether the disease is caused by a mutation in a single gene or there is an involvement of other mutations [150]. Traditionally linkage analysis studies were largely supplemented with robust genome-wide association studies (GWASs); however, with the advent of whole-genome sequencing, linkage analysis is again emerging as a critical tool for disease pathogenesis [151].
Vuori et al. performed genome-wide sib-pair linkage analysis for CACNB2, a gene for voltage-dependent L-type calcium channel subunit beta-2, to report linkage 10p12 overlapping the CACNB2 gene in PDR for T1D subjects. Furthermore, sequencing of CACNB2 revealed two coding variants, rs202152674 (R476C) and rs137886839 (S502L) [40]. In another study by Hallman et al., a genome-wide linkage scan was performed using 794 diabetes subjects from 393 Mexican American families, involving at least two diabetic siblings. The sample consisted of 567 individuals with retinopathy, including 282 affected sibling pairs. The study used 360 polymorphic markers with an average spacing of 9.4 cM to perform nonparametric linkage analysis. This study reported the highest risk of retinopathy for all families were on chromosome 3 and chromosome 12, while the risk of moderate to severe NPDR occurred on chromosomes 5, 6, and 19 [152].
4.3. Genome-Wide Association Studies
GWASs involve scanning genome for SNPs that are common in the human genome and determines how these polymorphisms are distributed across the different patient populations. GWAS serves a dual-purpose: (i) to uncover associations between individual SNPs and risk of disease between generations and (ii) to identify an individual’s risk for a particular condition [153,154]. In contrast to single-gene studies, GWAS assesses the entire genome; hence, GWAS is a non-candidate-driven approach. GWAS cannot specifically identify which genes are causal for a disease. The first successful GWAS study was published in 2002 on myocardial infarction, followed by a landmark study on age-related macular degeneration (AMD) that identified the complement factor H gene as a major risk factor for AMD [155]. As of 2020, the online repository GWAS catalog contains 4795 publications and 222481 associations, including 12 publications and five traits for DR [156].
Grassi et al. performed GWAS studies in individuals with severe DR as defined by DME or PDR in two large cohorts of T1D: (i) genetics of Kidney in Diabetes (GoKinD) and (ii) EDIC. The study included 973 cases and 1856 controls and assessed 2,543,887 SNPs to identify an intergenic SNP association, rs476141, on chromosome 1. This intergenic locus, rs476141, is situated between AKT serine threonine kinase 3 (AKT3) and zinc finger protein 238 (ZNF238). These genes play an essential role in cell survival, insulin signaling, and angiogenesis [41]. Interestingly, a similar finding for the role of insulin signaling was reported in a GWAS study performed in a Mexican population where CAMK4 (calcium/calmodulin-dependent protein kinase IV) was found to be highly associated with DR along with FMN1 (formin-1) [42]. CAMK4 increases transcriptional activity required for activating transcription factor-2 (ATF2) induced insulin gene expression [157].
Burdon et al. used an attractive study design where candidates identified in a GWAS study were subsequently validated in human retinas and animal studies. This study reported genetic variation near growth factor receptor bound-2 (GRB2) on chromosome 17q25.1 associated with sight-threatening DR. In human retinas, the GRB2 was observed to be expressed by all the retina layers, including blood vessels [43]. Furthermore, an animal model of DR showed that GRB2 is upregulated. GRB2 binds phosphorylated insulin receptor substrate 1 and activates the MAPK pathway via Ras in response to insulin. GRB2 is also involved in VEGF signaling [43], further implicating it in the pathogenesis of DR. In a larger multicentric GWAS study involving eight European and seven African cohorts, the C allele of rs142293996 in an intron of nuclear valosin-containing protein-like (NVL) was found to be associated with DR in European subjects. The NVL gene is expressed widely in the retina and encodes ATPASes associated with the cellular activities (AAA) superfamily [44]. Recently, Imamura et al. performed a GWAS study in Japanese subjects to identify novel SNP loci rs12630354 near STT3 Oligosaccharyltransferase Complex Catalytic Subunit B (STT3B) and rs140508424 in paralemmin-2 (PALM2) confers susceptibility to DR. This study also identified a novel gene EH domain containing 3 (EHD3) associated with DR susceptibility [45].
5. Conclusions and Future Perspectives
Genetics play an essential role in DR potentially explaining features, such as incidence, progression, heritability, and pathogenesis [158]. Accumulating evidence suggests that DR exhibits some level of inheritance; however, there is no monogenic association. Genetic studies show a racial variation in DR’s incidence. Heritability of DR is estimated as high as 27% and even higher (52%) for a more severe form [158] after adjusting known confounders such as blood pressure glycemic control and duration of diabetes. Putative candidate gene studies have highlighted various candidate genes associated with DR, such as AKR1B1, VEGFA, AGER, EPO, and NOS3; however, some of these gene candidates failed to replicate in populations with different ethnicities, again highlighting the role of genetics. We acknowledge that our selection of candidate genes is primarily based on the pathogenesis of DR. This review is no exhaustive summary of candidate genes for DR. For instance, a recent DRGen study identified additional candidate genes with PDR such as kruppel like factor 17 (KLF17), zinc finger protein 395 (ZNF395), myeloid cell surface antigen (CD33), pleckstrin homology domain-containing family G member 5 (PLEKHG5), and collagen type XVIII alpha 1 chain (COL18A1) [28]. Several of these candidate genes are known to have VEGF regulatory functions [28]. Future studies will likely identify the role of these genes in DR using different patient populations. Further, GWAS, a non-candidate-driven approach, may help curtail some limitations of candidate gene studies by uncovering associations between individual SNPs and DR risk. A novel approach of devising a genome wide polygenic score may help to determine the susceptibility of an individual to DR [159]. Together, precision medicine may enhance prevention and treatment strategies using transcriptomic studies. With advancing technologies and newer platforms for genetic analysis in DR, the future for treatment advances is bright.
Author Contributions
Writing—original draft preparation, A.D.B.; Writing—review and editing, A.D.B., A.S., A.B., T.A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NIH-NEI grant number R01EY02779 to Ashay Bhatwadekar.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
Thomas Ciulla has an employment relationship and equity ownership in Clearside Biomedical. This manuscript was written in his role as a Volunteer Clinical Professor at Indiana University and does not reflect any views or opinions of this corporation or its management. Ashay Bhatwadekar is an ad hoc pharmacist at PCA Pharmacy. The authors do not have any other pertinent or financial associations with any organization or entity with a financial interest in, or financial conflict with, the subject matter or materials discussed in the manuscript. The research of Ashay Bhatwadekar is supported by the NIH-NEI grant; however, the funders had no role in writing the document or in the decision to publish the results.
References
- Ciulla, T.A.; Amador, A.G.; Zinman, B. Diabetic retinopathy and diabetic macular edema: Pathophysiology, screening, and novel therapies. Diabetes Care 2003, 26, 2653–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diabetic Retinopathy. In Textbook of Diabetes; Wiley Online Library: Hoboken, NJ, USA, 2020.
- Thomas, R.; Halim, S.; Gurudas, S.; Sivaprasad, S.; Owens, D. IDF Diabetes Atlas: A review of studies utilising retinal photography on the global prevalence of diabetes related retinopathy between 2015 and 2018. Diabetes Res. Clin. Pract. 2019, 157, 107840. [Google Scholar] [CrossRef] [PubMed]
- Cdcgov Watch Out for Diabetic Retinopathy|Features|CDC. Available online: https://www.cdc.gov/features/diabetic-retinopathy/index.html (accessed on 30 June 2021).
- Diabetic Retinopathy Data and Statistics|National Eye Institute. Available online: https://www.ncbi.nlm.nih.gov/pubmed/ (accessed on 30 June 2021).
- Hammes, H.-P.; Lin, J.; Renner, O.; Shani, M.; Lundqvist, A.; Betsholtz, C.; Brownlee, M.; Deutsch, U. Pericytes and the pathogenesis of diabetic retinopathy. Diabetes 2002, 51, 3107–3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.; Xin, X.; Jee, K.; Babapoor-Farrokhran, S.; Kashiwabuchi, F.; Ma, T.; Bhutto, I.; Hassan, S.J.; Daoud, Y.; Baranano, D. VEGF secreted by hypoxic Müller cells induces MMP-2 expression and activity in endothelial cells to promote retinal neovascularization in proliferative diabetic retinopathy. Diabetes 2013, 62, 3863–3873. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Kern, T.S.; Song, B.; Stuebe, C. Mechanistic insights into pathological changes in the diabetic retina: Implications for targeting diabetic retinopathy. Am. J. Pathol. 2017, 187, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Shoshani, Y.; Harris, A.; Shoja, M.M.; Arieli, Y.; Ehrlich, R.; Primus, S.; Ciulla, T.; Cantor, A.; Wirostko, B.; Siesky, B.A. Impaired ocular blood flow regulation in patients with open-angle glaucoma and diabetes. Clin. Exp. Ophthalmol. 2012, 40, 697–705. [Google Scholar] [CrossRef]
- Ciulla, T.A.; Harris, A.; Latkany, P.; Piper, H.C.; Arend, O.; Garzozi, H.; Martin, B. Ocular perfusion abnormalities in diabetes. Acta Ophthalmol. Scand. 2002, 80, 468–477. [Google Scholar] [CrossRef]
- Curtis, T.; Gardiner, T.; Stitt, A. Microvascular lesions of diabetic retinopathy: Clues towards understanding pathogenesis? Eye 2009, 23, 1496–1508. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Boulton, M. The pathogenesis of diabetic retinopathy: Old concepts and new questions. Eye 2002, 16, 242–260. [Google Scholar] [CrossRef]
- Aiello, L.P.; Avery, R.L.; Arrigg, P.G.; Keyt, B.A.; Jampel, H.D.; Shah, S.T.; Pasquale, L.R.; Thieme, H.; Iwamoto, M.A.; Park, J.E. Vascular endothelial growth factor in ocular fluid of patients with diabetic retinopathy and other retinal disorders. N. Engl. J. Med. 1994, 331, 1480–1487. [Google Scholar] [CrossRef]
- Harris, M.I.; Klein, R.; Cowie, C.C.; Rowland, M.; Byrd-Holt, D.D. Is the risk of diabetic retinopathy greater in non-Hispanic blacks and Mexican Americans than in non-Hispanic whites with type 2 diabetes?: A US population study. Diabetes Care 1998, 21, 1230–1235. [Google Scholar] [CrossRef]
- Prevalence of Diagnosed Diabetes|Diabetes|CDC. Available online: https://www.cdc.gov/diabetes/data/statistics-report/diagnosed-diabetes.html (accessed on 30 June 2021).
- Bursell, S.-E.; Fonda, S.J.; Lewis, D.G.; Horton, M.B. Prevalence of diabetic retinopathy and diabetic macular edema in a primary care-based teleophthalmology program for American Indians and Alaskan Natives. PLoS ONE 2018, 13, e0198551. [Google Scholar]
- Nagi, D.; Pettitt, D.; Bennett, P.; Klein, R.; Knowler, W. Diabetic retinopathy assessed by fundus photography in Pima Indians with impaired glucose tolerance and NIDDM. Diabet. Med. 1997, 14, 449–456. [Google Scholar] [CrossRef]
- Lee, E.T.; Russell, D.; Morris, T.; Warn, A.; Kingsley, R.; Ogola, G. Visual impairment and eye abnormalities in Oklahoma Indians. Arch. Ophthalmol. 2005, 123, 1699–1704. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, I.B.; Brownlee, M. Beyond hemoglobin A1c—need for additional markers of risk for diabetic microvascular complications. JAMA 2010, 303, 2291–2292. [Google Scholar] [CrossRef]
- Control, D.; Group, C.T.R. Clustering of long-term complications in families with diabetes in the diabetes control and complications trial. Diabetes 1997, 46, 1829. [Google Scholar]
- Arar, N.H.; Freedman, B.I.; Adler, S.G.; Iyengar, S.K.; Chew, E.Y.; Davis, M.D.; Satko, S.G.; Bowden, D.W.; Duggirala, R.; Elston, R.C. Heritability of the severity of diabetic retinopathy: The FIND-Eye study. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3839–3845. [Google Scholar] [CrossRef]
- Hietala, K.; Forsblom, C.; Summanen, P.; Groop, P.-H. Heritability of proliferative diabetic retinopathy. Diabetes 2008, 57, 2176–2180. [Google Scholar] [CrossRef] [Green Version]
- Leslie, R.; Pyke, D. Diabetic retinopathy in identical twins. Diabetes 1982, 31, 19–21. [Google Scholar] [CrossRef]
- Bhatwadekar, A.D.; Duan, Y.; Chakravarthy, H.; Korah, M.; Caballero, S.; Busik, J.V.; Grant, M.B. Ataxia Telangiectasia Mutated Dysregulation Results in Diabetic Retinopathy. Stem Cells 2016, 34, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Jarajapu, Y.P.; Bhatwadekar, A.D.; Caballero, S.; Hazra, S.; Shenoy, V.; Medina, R.; Kent, D.; Stitt, A.W.; Thut, C.; Finney, E.M.; et al. Activation of the ACE2/angiotensin-(1-7)/Mas receptor axis enhances the reparative function of dysfunctional diabetic endothelial progenitors. Diabetes 2013, 62, 1258–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhary, S.; Hewitt, A.W.; Burdon, K.P.; Craig, J.E. A systematic meta-analysis of genetic association studies for diabetic retinopathy. Diabetes 2009, 58, 2137–2147. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Valle, M.L.; Beveridge, C.; Liu, Y.; Sharma, S. Unraveling the role of genetics in the pathogenesis of diabetic retinopathy. Eye 2019, 33, 534–541. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, A.P.; Mankad, R.N.; Marek, L.; Das, R.; Rangasamy, S.; Monickaraj, F.; Das, A. Genotypes and Phenotypes: A Search for Influential Genes in Diabetic Retinopathy. Int. J. Mol. Sci. 2020, 21, 2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Tian, Z.; Zhang, L.; Liu, R.; Guan, Q.; Jiang, J. Genetic association of AKR1B1 gene polymorphism rs759853 with diabetic retinopathy risk: A meta-analysis. Gene 2018, 676, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Abhary, S.; Burdon, K.P.; Laurie, K.J.; Thorpe, S.; Landers, J.; Goold, L.; Lake, S.; Petrovsky, N.; Craig, J.E. Aldose reductase gene polymorphisms and diabetic retinopathy susceptibility. Diabetes Care 2010, 33, 1834–1836. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, E.; Bakhtadze, E.; Sjögren, M.; Cilio, C.; Agardh, E.; Groop, L.; Agardh, C.-D. The−374 T/A polymorphism in the gene encoding RAGE is associated with diabetic nephropathy and retinopathy in type 1 diabetic patients. Diabetologia 2006, 49, 2745–2755. [Google Scholar] [CrossRef] [Green Version]
- Balasubbu, S.; Sundaresan, P.; Rajendran, A.; Ramasamy, K.; Govindarajan, G.; Perumalsamy, N.; Hejtmancik, J.F. Association analysis of nine candidate gene polymorphisms in Indian patients with Type 2 diabetic retinopathy. BMC Med. Genet. 2010, 11, 158. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Zhang, Y.; Zhang, X.; Li, X.; Liu, J. Association of VEGF Gene Polymorphisms with Susceptibility to Diabetic Retinopathy: A Systematic Review and Meta-Analysis. Horm. Metab. Res. 2020, 52, 264–279. [Google Scholar]
- Midani, F.; Ben Amor, Z.; El Afrit, M.A.; Kallel, A.; Feki, M.; Soualmia, H. The Role of Genetic Variants (rs869109213 and rs2070744) of the eNOS Gene and Bgl II in the α2 Subunit of the α2β1 Integrin Gene in Diabetic Retinopathy in a Tunisian Population, Seminars in Ophthalmology; Taylor & Francis: Oxford, UK, 2019; pp. 365–374. [Google Scholar]
- Luo, S.; Shi, C.; Wang, F.; Wu, Z. Association between the Angiotensin-Converting Enzyme (ACE) Genetic Polymorphism and Diabetic Retinopathy-A Meta-Analysis Comprising 10,168 Subjects. Int. J. Environ. Res. Public Health 2016, 13, 1142. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Pan, M.; Hu, N.; Wu, Y.Y.; Chen, H.; Zhu, J.H.; Guan, H.J.; Sang, A.M. Association of angiotensin-converting enzyme gene 2350 G/A polymorphism with diabetic retinopathy in Chinese Han population. Mol. Biol. Rep. 2013, 40, 463–468. [Google Scholar] [CrossRef]
- Fan, Y.; Fu, Y.Y.; Chen, Z.; Hu, Y.Y.; Shen, J. Gene-gene interaction of erythropoietin gene polymorphisms and diabetic retinopathy in Chinese Han. Exp. Biol. Med. 2016, 241, 1524–1530. [Google Scholar] [CrossRef] [Green Version]
- Abhary, S.; Burdon, K.P.; Casson, R.J.; Goggin, M.; Petrovsky, N.P.; Craig, J.E. Association between erythropoietin gene polymorphisms and diabetic retinopathy. Arch. Ophthalmol. 2010, 128, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Mankoč Ramuš, S.; Pungeršek, G.; Petrovič, M.G.; Petrovič, D. The GG genotype of erythropoietin rs1617640 polymorphism affects the risk of proliferative diabetic retinopathy in Slovenian subjects with Type 2 diabetes mellitus: Enemy or ally? Acta Ophthalmol. 2021. [Google Scholar] [CrossRef]
- Vuori, N.; Sandholm, N.; Kumar, A.; Hietala, K.; Syreeni, A.; Forsblom, C.; Juuti-Uusitalo, K.; Skottman, H.; Imamura, M.; Maeda, S. CACNB2 is a novel susceptibility gene for diabetic retinopathy in type 1 diabetes. Diabetes 2019, 68, 2165–2174. [Google Scholar] [CrossRef] [Green Version]
- Grassi, M.A.; Tikhomirov, A.; Ramalingam, S.; Below, J.E.; Cox, N.J.; Nicolae, D.L. Genome-wide meta-analysis for severe diabetic retinopathy. Hum. Mol. Genet. 2011, 20, 2472–2481. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.-P.; Hallman, D.M.; Gonzalez, V.H.; Klein, B.E.; Klein, R.; Hayes, M.G.; Cox, N.J.; Bell, G.I.; Hanis, C.L. Identification of diabetic retinopathy genes through a genome-wide association study among Mexican-Americans from Starr County, Texas. J. Ophthalmol. 2009, 2010, 861291. [Google Scholar] [CrossRef]
- Burdon, K.P.; Fogarty, R.D.; Shen, W.; Abhary, S.; Kaidonis, G.; Appukuttan, B.; Hewitt, A.W.; Sharma, S.; Daniell, M.; Essex, R.W. Genome-wide association study for sight-threatening diabetic retinopathy reveals association with genetic variation near the GRB2 gene. Diabetologia 2015, 58, 2288–2297. [Google Scholar] [CrossRef]
- Pollack, S.; Igo, R.P.; Jensen, R.A.; Christiansen, M.; Li, X.; Cheng, C.-Y.; Ng, M.C.; Smith, A.V.; Rossin, E.J.; Segrè, A.V. Multiethnic genome-wide association study of diabetic retinopathy using liability threshold modeling of duration of diabetes and glycemic control. Diabetes 2019, 68, 441–456. [Google Scholar] [CrossRef] [Green Version]
- Imamura, M.; Takahashi, A.; Matsunami, M.; Horikoshi, M.; Iwata, M.; Araki, S.-I.; Toyoda, M.; Susarla, G.; Ahn, J.; Park, K.H. Genome-wide association studies identify two novel loci conferring susceptibility to diabetic retinopathy in Japanese patients with type 2 diabetes. Hum. Mol. Genet. 2021, 30, 716–726. [Google Scholar] [CrossRef]
- Mathebula, S.D. Polyol pathway: A possible mechanism of diabetes complications in the eye. Afr. Vis. Eye Health 2015, 74, 5. [Google Scholar] [CrossRef]
- Dvornik, D. Aldose reductase inhibitors as pathobiochemical probes. J. Diabetes Complicat. 1992, 6, 25–34. [Google Scholar] [CrossRef]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Leto, G.; Pricci, F.; Amadio, L.; Iacobini, C.; Cordone, S.; Diaz-Horta, O.; Romeo, G.; Barsotti, P.; Rotella, C.M.; di Mario, U. Increased retinal endothelial cell monolayer permeability induced by the diabetic milieu: Role of advanced non-enzymatic glycation and polyol pathway activation. Diabetes/Metab. Res. Rev. 2001, 17, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Moczulski, D.; Scott, L.; Antonellis, A.; Rogus, J.; Rich, S.; Warram, J.; Krolewski, A. Aldose reductase gene polymorphisms and susceptibility to diabetic nephropathy in type 1 diabetes mellitus. Diabet. Med. 2000, 17, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.; Lam, K.L.; Watt, N.M.-S.; Chung, S.S.-M. An (AC) n dinucleotide repeat polymorphic marker at the 5′ end of the aldose reductase gene is associated with early-onset diabetic retinopathy in NIDDM patients. Diabetes 1995, 44, 727–732. [Google Scholar] [CrossRef]
- Li, W.; Chen, S.; Mei, Z.; Zhao, F.; Xiang, Y. Polymorphisms in sorbitol-aldose reductase (Polyol) pathway genes and their influence on risk of diabetic retinopathy among Han Chinese. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 7073. [Google Scholar] [CrossRef]
- Yang, B.; Millward, A.; Demaine, A. Functional differences between the susceptibility Z−2/C−106 and protective Z+2/T−106 promoter region polymorphisms of the aldose reductase gene may account for the association with diabetic microvascular complications. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2003, 1639, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Katakami, N.; Kaneto, H.; Takahara, M.; Matsuoka, T.; Imamura, K.; Ishibashi, F.; Kanda, T.; Kawai, K.; Osonoi, T.; Matsuhisa, M. Aldose reductase C-106T gene polymorphism is associated with diabetic retinopathy in Japanese patients with Type 2 diabetes. Diabetes Res. Clin. Pract. 2011, 92, e57–e60. [Google Scholar] [CrossRef]
- Rezaee, M.R.; Amiri, A.A.; Hashemi-Soteh, M.B.; Daneshvar, F.; Emady-Jamaly, R.; Jafari, R.; Soleimani, B.; Haghiaminjan, H. Aldose reductase C-106T gene polymorphism in Type 2 diabetics with microangiopathy in Iranian individuals. Indian J. Endocrinol. Metab. 2015, 19, 95–99. [Google Scholar]
- Kaur, N.; Vanita, V. Association of aldose reductase gene (AKR1B1) polymorphism with diabetic retinopathy. Diabetes Res. Clin. Pract. 2016, 121, 41–48. [Google Scholar] [CrossRef]
- SIu, M.; Bushuieva, O.; Ziablitsev, S.; Natrus, L. Relationship of the AKR1B1 rs759853 and rs9640883 with the development of diabetic retinopathy. J. Ophthalmol. 2017, 2, 3–7. [Google Scholar]
- Mogilevskyy, S.Y.; Panchenko, I.O.; Ziablitsev, S.V. Predicting the risk of diabetic retinopathy-associated macular edema in patients with type 2 diabetes mellitus. Oftalmol. Zhurnal 2019, 3–8. [Google Scholar] [CrossRef]
- Mrozikiewicz-Rakowska, B.; Łukawska, M.; Nehring, P.; Szymański, K.; Sobczyk-Kopcioł, A.; Krzyżewska, M.; Maroszek, P.; Płoski, R.; Czupryniak, L. Genetic predictors associated with diabetic retinopathy in patients with diabetic foot. Pol. Arch. Intern. Med. 2018, 128, 35–42. [Google Scholar]
- Lin, S.; Peng, Y.; Cao, M.; Chen, R.; Hu, J.; Pu, Z.; Cai, Z.; Mou, L. Association between Aldose Reductase Gene C (-106) T Polymorphism and Diabetic Retinopathy: A Systematic Review and Meta-Analysis. Ophthalmic Res. 2020, 63, 224–233. [Google Scholar] [CrossRef]
- Bushuyeva, O.V. New factors of diabetic retinopathy progression in Type 2 diabetes mellitus patients. East. Eur. Sci. J. Med. Sci. 2020, 1, 54–57. [Google Scholar]
- Abu-Hassan, D.; Al-Bdour, M.; Saleh, I.; Freihat, M.; El-Khateeb, M. The relationship between aldose reductase gene C106T polymorphism and the severity of retinopathy in Type 2 diabetic patients: A case–control study. J. Res. Med. Sci. 2021, 26, 2. [Google Scholar] [CrossRef]
- Deng, Y.; Yang, X.-F.; Gu, H.; Lim, A.; Ulziibat, M.; Snellingen, T.; Xu, J.; Ma, K.; Liu, N.-p. Association of C (-106) T polymorphism in aldose reductase gene with diabetic retinopathy in Chinese patients with Type 2 diabetes mellitus. Chin. Med. Sci. J. 2014, 29, 1–6. [Google Scholar] [CrossRef]
- Song, Z.-D.; Tao, Y.; Han, N.; Wu, Y.-Z. Association of the aldose reductase-106TT genotype with increased risk for diabetic retinopathy in the Chinese han population: An updated meta-analysis. Curr. Eye Res. 2016, 41, 1087–1091. [Google Scholar] [CrossRef]
- Shawki, H.A.; Elzehery, R.; Abo-hashem, E.M.; Shahin, M.; Youssef, M.M. Gene polymorphism of C106T “rs759853” is not associated with diabetic retinopathy in Egyptian patients with Type 2 diabetes mellitus. Gene Rep. 2020, 21, 100865. [Google Scholar] [CrossRef]
- Iskhakova, A.; Toropovsky, A.; Zolotarev, A.; Pavlova, O.; Komarova, M. Molecular and genetic aspects of early diagnosis of diabetic retinopathy. Int. Res. J. 2021, 3, 79–85. [Google Scholar]
- Shah, V.O.; Scavini, M.; Nikolic, J.; Sun, Y.; Vai, S.; Griffith, J.K.; Dorin, R.I.; Stidley, C.; Yacoub, M.; Vander Jagt, D.L. Z−2 microsatellite allele is linked to increased expression of the aldose reductase gene in diabetic nephropathy. J. Clin. Endocrinol. Metab. 1998, 83, 2886–2891. [Google Scholar] [CrossRef] [PubMed]
- Demaine, A.; Cross, D.; Millward, A. Polymorphisms of the aldose reductase gene and susceptibility to retinopathy in Type 1 diabetes mellitus. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4064–4068. [Google Scholar]
- Wang, Y.; Ng, M.C.; Lee, S.-C.; So, W.-Y.; Tong, P.C.; Cockram, C.S.; Critchley, J.A.; Chan, J.C. Phenotypic heterogeneity and associations of two aldose reductase gene polymorphisms with nephropathy and retinopathy in Type 2 diabetes. Diabetes Care 2003, 26, 2410–2415. [Google Scholar] [CrossRef] [Green Version]
- Mi, W.; Xia, Y.; Bian, Y. Meta-analysis of the association between aldose reductase gene (CA)n microsatellite variants and risk of diabetic retinopathy. Exp. Ther. Med. 2019, 18. [Google Scholar] [CrossRef]
- Fan, W.-Y.; Gu, H.; Yang, X.-F.; She, C.-Y.; Liu, X.-P.; Liu, N.-P. Association of candidate gene polymorphisms with diabetic retinopathy in Chinese patients with type 2 diabetes. Int. J. Ophthalmol. 2020, 13, 301. [Google Scholar] [CrossRef]
- Bucala, R.; Vlassara, H. Advanced glycosylation end products in diabetic renal and vascular disease. Am. J. Kidney Dis. 1995, 26, 875–888. [Google Scholar] [CrossRef]
- Vlassara, H.; Palace, M.R. Diabetes and advanced glycation endproducts. J. Intern. Med. 2002, 251, 87–101. [Google Scholar] [CrossRef]
- Stitt, A.W.; Li, Y.M.; Gardiner, T.A.; Bucala, R.; Archer, D.B.; Vlassara, H. Advanced glycation end products (AGEs) co-localize with AGE receptors in the retinal vasculature of diabetic and of AGE-infused rats. Am. J. Pathol 1997, 150, 523–531. [Google Scholar]
- Hughes, S.J.; Wall, N.; Scholfield, C.N.; McGeown, J.G.; Gardiner, T.A.; Stitt, A.W.; Curtis, T.M. Advanced glycation endproduct modified basement membrane attenuates endothelin-1 induced [Ca2+]i signalling and contraction in retinal microvascular pericytes. Mol. Vis. 2004, 10, 996–1004. [Google Scholar]
- Chen, B.H.; Jiang, D.Y.; Tang, L.S. Advanced glycation end-products induce apoptosis involving the signaling pathways of oxidative stress in bovine retinal pericytes. Life Sci. 2006, 79, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Neeper, M.; Schmidt, A.M.; Brett, J.; Yan, S.D.; Wang, F.; Pan, Y.C.; Elliston, K.; Stern, D.; Shaw, A. Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. J. Biol. Chem. 1992, 267, 14998–15004. [Google Scholar] [CrossRef]
- Yamagishi, S.; Fujimori, H.; Yonekura, H.; Yamamoto, Y.; Yamamoto, H. Advanced glycation endproducts inhibit prostacyclin production and induce plasminogen activator inhibitor-1 in human microvascular endothelial cells. Diabetologia 1998, 41, 1435–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, N.; Kim, Y.S.; Lanting, L.; Natarajan, R. Regulation of cyclooxygenase-2 expression in monocytes by ligation of the receptor for advanced glycation end products. J. Biol. Chem. 2003, 278, 34834–34844. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.M.; Hori, O.; Chen, J.X.; Li, J.F.; Crandall, J.; Zhang, J.; Cao, R.; Yan, S.D.; Brett, J.; Stern, D. Advanced glycation endproducts interacting with their endothelial receptor induce expression of vascular cell adhesion molecule-1 (VCAM-1) in cultured human endothelial cells and in mice. A potential mechanism for the accelerated vasculopathy of diabetes. J. Clin. Investig. 1995, 96, 1395–1403. [Google Scholar] [CrossRef] [Green Version]
- Wautier, J.L.; Schmidt, A.M. Protein glycation: A firm link to endothelial cell dysfunction. Circ. Res. 2004, 95, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Cui, Y.; Li, B.; Weng, J.; Wang, W.; Zhang, S.; Huang, X.; Guo, X.; Huang, Q. Advanced glycation end products induce immature angiogenesis in in vivo and ex vivo mouse models. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H519–H533. [Google Scholar] [CrossRef]
- Gui, F.; You, Z.; Fu, S.; Wu, H.; Zhang, Y. Endothelial Dysfunction in Diabetic Retinopathy. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef]
- Erna, R.; Sastrawan, D.; Saleh, M.I. Effect of RAGE -429T/C and Gly82Ser Gene Polymorphism in Diabetic Retinopathy in General Hospital Mohammad Hoesin Palembang. J. Res. Med. Dent. Sci. 2017, 5, 19–23. [Google Scholar]
- Yuan, D.; Yuan, D.; Liu, Q. Association of the receptor for advanced glycation end products gene polymorphisms with diabetic retinopathy in type 2 diabetes: A meta-analysis. Ophthalmologica 2012, 227, 223–232. [Google Scholar] [CrossRef]
- Qayyum, S.; Afzal, M.; Naveed, A.K. Association analysis of 374T/A (rs1800624) receptor for advanced glycation end-products (RAGE) gene polymorphism with diabetic retinopathy in Pakistani patients. Pak. J. Med. Sci. 2021, 37, 733. [Google Scholar] [CrossRef]
- Tao, D.; Mai, X.; Zhang, T.; Mei, Y. Association between the RAGE (receptor for advanced glycation end-products)-374T/A gene polymorphism and diabetic retinopathy in T2DM. Rev. Assoc. MÉDica Bras. 2017, 63, 971–977. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Yang, J.; Sui, W.; Qu, B.; Huang, P.; Chen, Y. Association of genetic variants in the receptor for advanced glycation end products gene with diabetic retinopathy: A meta-analysis. Medicine 2016, 95, e4463. [Google Scholar] [CrossRef]
- Lu, W.; Feng, B. The-374A allele of the RAGE gene as a potential protective factor for vascular complications in type 2 diabetes: A meta-analysis. Tohoku J. Exp. Med. 2010, 220, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Ng, Z.X.; Kuppusamy, U.R.; Tajunisah, I.; Fong, K.C.S.; Chua, K.H. Association analysis of− 429T/C and− 374T/A polymorphisms of receptor of advanced glycation end products (RAGE) gene in Malaysian with type 2 diabetic retinopathy. Diabetes Res. Clin. Pract. 2012, 95, 372–377. [Google Scholar] [CrossRef]
- Vanita, V. Association of RAGE (p. Gly82Ser) and MnSOD (p. Val16Ala) polymorphisms with diabetic retinopathy in T2DM patients from north India. Diabetes Res. Clin. Pract. 2014, 104, 155–162. [Google Scholar] [CrossRef]
- Jia, C.; Yong, Y.; Tianhua, X.; Zheyao, G.; Jian, Z. Association of Gly82Ser polymorphism of RAGE gene with diabetic retinopathy in Han people with type 2 diabetes of Wuxi region. Chin. J. Exp. Ophthalmol. 2016, 34, 910–914. [Google Scholar]
- Kang, P.; Tian, C.; Jia, C. Association of RAGE gene polymorphisms with type 2 diabetes mellitus, diabetic retinopathy and diabetic nephropathy. Gene 2012, 500, 1–9. [Google Scholar] [CrossRef]
- Niu, W.; Qi, Y.; Wu, Z.; Liu, Y.; Zhu, D.; Jin, W. A meta-analysis of receptor for advanced glycation end products gene: Four well-evaluated polymorphisms with diabetes mellitus. Mol. Cell. Endocrinol. 2012, 358, 9–17. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, L.; Wang, L.; Liao, Y.; Wu, Z.; Ye, F.; Xu, S.; Yi, L. Association of 1704G/T and G82S polymorphisms in the receptor for advanced glycation end products gene with diabetic retinopathy in Chinese population. J. Endocrinol. Investig. 2009, 32, 258–262. [Google Scholar] [CrossRef]
- Yoshioka, K.; Yoshida, T.; Takakura, Y.; Umekawa, T.; Kogure, A.; Toda, H.; Yoshikawa, T. Relation between polymorphisms G1704T and G82S of rage gene and diabetic retinopathy in Japanese type 2 diabetic patients. Intern. Med. 2005, 44, 417–421. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wu, Q.; Li, Y.; Fan, X.; Hao, Y.; Sun, H.; Cui, Y.; Han, L. Association of the receptor for advanced glycation end products gene polymorphisms and circulating RAGE levels with diabetic retinopathy in the Chinese population. J. Diabetes Res. 2013, 2013, 264579. [Google Scholar] [CrossRef]
- Holmes, D.I.; Zachary, I. The vascular endothelial growth factor (VEGF) family: Angiogenic factors in health and disease. Genome Biol. 2005, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Ju, H.; Sun, M.; Chen, H. Serum Vascular Endothelial Growth Factor Levels Correlate with Severity of Retinopathy in Diabetic Patients: A Systematic Review and Meta-Analysis. Dis. Markers 2019, 2019, 9401628. [Google Scholar] [CrossRef] [Green Version]
- Morello, C.M. Etiology and natural history of diabetic retinopathy: An overview. Am. J. Health Syst. Pharm. 2007, 64, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.; Soden, J.; Brenchley, P.E.; Ralph, S.; Ray, D.W. Haplotype analysis of the polymorphic human vascular endothelial growth factor gene promoter. Cancer Res. 2003, 63, 812–816. [Google Scholar]
- Khan, S.Z.; Ajmal, N.; Shaikh, R. Diabetic Retinopathy and Vascular Endothelial Growth Factor Gene Insertion/Deletion Polymorphism. Can. J. Diabetes 2020, 44, 287–291. [Google Scholar] [CrossRef]
- Hu, L.; Gong, C.; Chen, X.; Zhou, H.; Yan, J.; Hong, W. Associations between Vascular Endothelial Growth Factor Gene Polymorphisms and Different Types of Diabetic Retinopathy Susceptibility: A Systematic Review and Meta-Analysis. J. Diabetes Res. 2021, 2021, 7059139. [Google Scholar] [CrossRef]
- Xie, X.-J.; Yang, Y.-M.; Jiang, J.-K.; Lu, Y.-Q. Association between the vascular endothelial growth factor single nucleotide polymorphisms and diabetic retinopathy risk: A meta-analysis. J. Diabetes 2017, 9, 738–753. [Google Scholar] [CrossRef]
- Al-Kateb, H.; Mirea, L.; Xie, X.; Sun, L.; Liu, M.; Chen, H.; Bull, S.B.; Boright, A.P.; Paterson, A.D. Multiple variants in vascular endothelial growth factor (VEGFA) are risk factors for time to severe retinopathy in type 1 diabetes: The DCCT/EDIC genetics study. Diabetes 2007, 56, 2161–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhary, S.; Burdon, K.P.; Gupta, A.; Lake, S.; Selva, D.; Petrovsky, N.; Craig, J.E. Common sequence variation in the VEGFA gene predicts risk of diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5552–5558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Paterson, A.D.; Roshandel, D.; Raza, A.; Ajmal, M.; Waheed, N.K.; Azam, M.; Qamar, R. Association of IGF1 and VEGFA polymorphisms with diabetic retinopathy in Pakistani population. Acta Diabetol. 2020, 57, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Sajovic, J.; Cilenšek, I.; Mankoč, S.; Tajnšek, Š.; Kunej, T.; Petrovič, D.; Petrovič, M.G. Vascular endothelial growth factor (VEGF)-related polymorphisms rs10738760 and rs6921438 are not risk factors for proliferative diabetic retinopathy (PDR) in patients with type 2 diabetes mellitus (T2DM). Bosn. J. Basic Med. Sci. 2019, 19, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzić, R.; Cilenšek, I.; Pleskovič, R.Z.; Mankoč, S.; Milutinović, A. Vascular endothelial growth factor (VEGF)-related single nucleotide polymorphisms rs10738760 and rs6921438 are not associated with diabetic retinopathy (DR) in Slovenian patients with type 2 diabetes mellitus (T2DM). Bosn. J. Basic Med. Sci. 2017, 17, 328. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Salinas, R.; Garcia-Gutierrez, M.C.; Garcia-Aguirre, G.; Morales-Canton, V.; Velez-Montoya, R.; Soberon-Ventura, V.R.; Gonzalez, V.; Lechuga, R.; Garcia-Solis, P.; Garcia-Gutierrez, D.G. Evaluation of VEGF gene polymorphisms and proliferative diabetic retinopathy in Mexican population. Int. J. Ophthalmol. 2017, 10, 135. [Google Scholar]
- Amer, A.K.; Khalaf, N.A.; Aboelmakarem, S.H.; Elsobky, M.S.; Abdelrasoul, M.R.; Abdelazeem, A.A.; Noweir, S.R.; Refaat, S.; Moemen, L.A.; Mohammed, S.A.; et al. Vascular endothelial growth factor +405G/C polymorphism as a predictor of diabetic retinopathy. Bull. Natl. Res. Cent. 2020, 44, 54. [Google Scholar] [CrossRef] [Green Version]
- Hussin, H.Y.; Hussein, W.; Ibrahim, S.; Kambal, S.; Almhdi, S.; Abdrabo, A.E. Association of Vascular Endothelial Growth Factor +405G/C Polymorphism with Diabetic Retinopathy among Sudanese Patients. Am. J. Biomed. Sci. Res. 2021, 11, 437–443. [Google Scholar] [CrossRef]
- Chun, M.-Y.; Hwang, H.-S.; Cho, H.-Y.; Chun, H.-J.; Woo, J.-T.; Lee, K.-W.; Nam, M.-S.; Baik, S.-H.; Kim, Y.-S.; Park, Y. Association of vascular endothelial growth factor polymorphisms with nonproliferative and proliferative diabetic retinopathy. J. Clin. Endocrinol. Metab 2010, 95, 3547–3551. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Deng, Y.; Gu, H.; Ren, X.; Li, N.; Lim, A.; Snellingen, T.; Liu, X.; Wang, N.; Liu, N. Candidate gene association study for diabetic retinopathy in Chinese patients with type 2 diabetes. Mol. Vis. 2014, 20, 200–214. [Google Scholar]
- Nakamura, S.; Iwasaki, N.; Funatsu, H.; Kitano, S.; Iwamoto, Y. Impact of variants in the VEGF gene on progression of proliferative diabetic retinopathy. Graefes Arch. Clin. Exp. Ophthalmol. 2009, 247, 21–26. [Google Scholar] [CrossRef]
- Bleda, S.; De Haro, J.; Varela, C.; Esparza, L.; Ferruelo, A.; Acin, F. Vascular endothelial growth factor polymorphisms are involved in the late vascular complications in Type II diabetic patients. Diabetes Vasc. Dis. Res. 2011, 9, 68–74. [Google Scholar] [CrossRef]
- Abdel Fattah, R.; Eltanamly, R.; Nabih, M.; Kamal, M. Vascular endothelial growth factor gene polymorphism is not associated with diabetic retinopathy in Egyptian Patients. Middle East. Afr. J. Ophthalmol. 2016, 23, 75–78. [Google Scholar]
- Fan, X.; Wu, Q.; Li, Y.; Hao, Y.; Ning, N.; Kang, Z.; Cui, Y.; Liu, R.; Han, L. Association of polymorphisms in the vascular endothelial growth factor gene and its serum levels with diabetic retinopathy in Chinese patients with type 2 diabetes: A cross-sectional study. Chin. Med. J. 2014, 127, 651–657. [Google Scholar]
- Han, L.; Zhang, L.; Xing, W.; Zhuo, R.; Lin, X.; Hao, Y.; Wu, Q.; Zhao, J. The associations between VEGF gene polymorphisms and diabetic retinopathy susceptibility: A meta-analysis of 11 case-control studies. J. Diabetes Res. 2014, 2014, 805801. [Google Scholar] [CrossRef] [Green Version]
- Dorner, G.T.; Garhofer, G.; Kiss, B.; Polska, E.; Polak, K.; Riva, C.E.; Schmetterer, L. Nitric oxide regulates retinal vascular tone in humans. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H631–H636. [Google Scholar] [CrossRef]
- Chen, K.; Pittman, R.N.; Popel, A.S. Nitric oxide in the vasculature: Where does it come from and where does it go? A quantitative perspective. Antioxid Redox Signal 2008, 10, 1185–1198. [Google Scholar] [CrossRef] [Green Version]
- Sessa, W.C. eNOS at a glance. J. Cell Sci. 2004, 117, 2427–2429. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, E.W.; Stegeman, C.A.; Heeringa, P.; Henning, R.H.; van Goor, H. Protective role of endothelial nitric oxide synthase. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2003, 199, 8–17. [Google Scholar] [CrossRef]
- Gericke, A.; Wolff, I.; Musayeva, A.; Zadeh, J.K.; Manicam, C.; Pfeiffer, N.; Li, H.; Xia, N. Retinal arteriole reactivity in mice lacking the endothelial nitric oxide synthase (eNOS) gene. Exp. Eye Res. 2019, 181, 150–156. [Google Scholar] [CrossRef]
- Li, Q.; Verma, A.; Han, P.-Y.; Nakagawa, T.; Johnson, R.J.; Grant, M.B.; Campbell-Thompson, M.; Jarajapu, Y.P.; Lei, B.; Hauswirth, W.W. Diabetic eNOS-knockout mice develop accelerated retinopathy. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5240–5246. [Google Scholar] [CrossRef] [Green Version]
- McAuley, A.K.; Sanfilippo, P.G.; Hewitt, A.W.; Liang, H.; Lamoureux, E.; Wang, J.J.; Connell, P.P. Vitreous biomarkers in diabetic retinopathy: A systematic review and meta-analysis. J. Diabetes Complicat. 2014, 28, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Mihoubi, E.; Bouldjennet, F.; Raache, R.; Amroun, H.; Azzouz, M.; Benazouz, N.; Touil-Boukoffa, C.; Attal, N. T-786C endothelial nitric oxide gene polymorphism and type 1 diabetic retinopathy in the Algerian population. J. Fr. d’Ophtalmol. 2019, 42, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Cheema, B.S.; Sharma, R.; Bhansali, A.; Khullar, M. Endothelial nitric oxide synthase gene polymorphism and Type 2 diabetic retinopathy among Asian Indians. Acta Diabetol. 2012, 49, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Li, T.; Zheng, B.; Zheng, Z. Nitric oxide synthase 3 (NOS3) 4b/a, T-786C and G894T polymorphisms in association with diabetic retinopathy susceptibility: A meta-analysis. Ophthalmic Genet. 2012, 33, 200–207. [Google Scholar] [CrossRef]
- Qian-Qian, Y.; Yong, Y.; Jing, Z.; Dong-Hong, F.; Tian-Hua, X.; Li, Y.; Lan, L.; Jia, C.; Zhe-Yao, G. Association between a 27-bp variable number of tandem repeat polymorphism in intron 4 of the eNOS gene and risk for diabetic retinopathy Type 2 diabetes mellitus: A meta-analysis. Curr. Eye Res. 2014, 39, 1052–1058. [Google Scholar] [CrossRef] [PubMed]
- Cilenšek, I.; Mankoč, S.; Globočnik Petrovič, M.; Petrovič, D. The 4a/4a genotype of the VNTR polymorphism for endothelial nitric oxide synthase (eNOS) gene predicts risk for proliferative diabetic retinopathy in Slovenian patients (Caucasians) with type 2 diabetes mellitus. Mol. Biol. Rep. 2012, 39, 7061–7067. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.-J.; Chen, R.; Ren, H.-Z.; Guo, X.; Guo, J.; Chen, L.-M. Association between eNOS 4b/a polymorphism and the risk of diabetic retinopathy in type 2 diabetes mellitus: A meta-analysis. J. Diabetes Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- She, C.; Yang, X.; Gu, H.; Deng, Y.; Xu, J.; Ma, K.; Liu, N. The association of variable number of tandem repeats polymorphism in the endothelial nitric oxide synthase gene and diabetic retinopathy. Zhonghua Yan Ke Za Zhi 2015, 51, 338–343. [Google Scholar] [PubMed]
- Momeni, A.; Chaleshtori, M.H.; Saadatmand, S.; Kheiri, S. Correlation of Endothelial Nitric Oxide Synthase Gene Polymorphism (GG, TT and GT Genotype) with Proteinuria and Retinopathy in Type 2 Diabetic Patients. J. Clin. Diagn Res. 2016, 10, Oc32–Oc35. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, E.L.; Phipps, J.A.; Ward, M.M.; Vessey, K.A.; Wilkinson-Berka, J.L. The renin–angiotensin system in retinal health and disease: Its influence on neurons, glia and the vasculature. Prog. Retin. Eye Res. 2010, 29, 284–311. [Google Scholar] [CrossRef]
- Li, Y.; Yan, Z.; Chaudhry, K.; Kazlauskas, A. The Renin-Angiotensin-Aldosterone System (RAAS) Is One of the Effectors by Which Vascular Endothelial Growth Factor (VEGF)/Anti-VEGF Controls the Endothelial Cell Barrier. Am. J. Pathol. 2020, 190, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, F.; Zhang, Y.; Zhao, S.H.; Zhao, W.J.; Yan, S.L.; Wang, Y.G. Effects of RAS inhibitors on diabetic retinopathy: A systematic review and meta-analysis. Lancet Diabetes Endocrinol. 2015, 3, 263–274. [Google Scholar] [CrossRef]
- Sjølie, A.K.; Klein, R.; Porta, M.; Orchard, T.; Fuller, J.; Parving, H.H.; Bilous, R.; Chaturvedi, N. Effect of candesartan on progression and regression of retinopathy in type 2 diabetes (DIRECT-Protect 2): A randomised placebo-controlled trial. Lancet 2008, 372, 1385–1393. [Google Scholar] [CrossRef]
- Mauer, M.; Zinman, B.; Gardiner, R.; Suissa, S.; Sinaiko, A.; Strand, T.; Drummond, K.; Donnelly, S.; Goodyer, P.; Gubler, M.C.; et al. Renal and retinal effects of enalapril and losartan in type 1 diabetes. N. Engl. J. Med. 2009, 361, 40–51. [Google Scholar] [CrossRef]
- Neroev, V.V.; Chesnokova, N.B.; Pavlenko, T.A.; Okhotsimskaya, T.D.; Beznos, O.V.; Fadeeva, V.A.; Struchkova, S.V. Changes in the concentration of angiotensin II, angiotensin-transforming enzyme and matrix metaloproteinase-9 in tear fluid and serum in patients with diabetic retinopathy. Ophthalmology 2020, 17, 771–778. [Google Scholar]
- Rigat, B.; Hubert, C.; Alhenc-Gelas, F.; Cambien, F.; Corvol, P.; Soubrier, F. An insertion/deletion polymorphism in the angiotensin I-converting enzyme gene accounting for half the variance of serum enzyme levels. J. Clin. Investig. 1990, 86, 1343–1346. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Yang, X.F.; Gu, H.; Deng, Y.; Xu, J.; Ma, K.; Liu, N.P. Relationship of angiotensin converting enzyme gene polymorphism with diabetic retinopathy. Zhonghua Yan Ke Za Zhi 2013, 49, 52–57. [Google Scholar]
- Pirozzi, F.F.; Belini Junior, E.; Okumura, J.V.; Salvarani, M.; Bonini-Domingos, C.R.; Ruiz, M.A. The relationship between of ACE I/D and the MTHFR C677T polymorphisms in the pathophysiology of type 2 diabetes mellitus in a population of Brazilian obese patients. Arch. Endocrinol. Metab. 2018, 62, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.C.; Wang, M.; Pan, Y.H.; Zhang, X.X.; Tian, F.; Chen, Y.L.; Zhao, H.L. The relationship between ACE/AGT gene polymorphisms and the risk of diabetic retinopathy in Chinese patients with type 2 diabetes. J. Renin Angiotensin Aldosterone Syst. 2018, 19, 1470320317752955. [Google Scholar] [CrossRef] [Green Version]
- Wiwanitkit, V. Angiotensin-converting enzyme gene polymorphism is correlated to diabetic retinopathy: A meta-analysis. J. Diabetes Complicat. 2008, 22, 144–146. [Google Scholar] [CrossRef]
- Reid, G.; Lois, N. Erythropoietin in diabetic retinopathy. Vis. Res. 2017, 139, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Davidović, S.; Babić, N.; Jovanović, S.; Barišić, S.; Grković, D.; Miljković, A. Serum erythropoietin concentration and its correlation with stage of diabetic retinopathy. BMC Ophthalmol. 2019, 19, 227. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Liu, N. Meta-analysis of association between EPO gene polymorphisms and diabetic retinopathy. Ophthalmol. China 2020, 29, 454–457. [Google Scholar]
- Li, H.; Xu, H.; Li, Y.; Zhao, D.; Ma, B. Associations between erythropoietin polymorphisms and risk of diabetic microvascular complications. Oncotarget 2017, 8, 112675–112684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulst, S.M. Genetic linkage analysis. Arch. Neurol. 1999, 56, 667–672. [Google Scholar] [CrossRef]
- Ott, J.; Wang, J.; Leal, S.M. Genetic linkage analysis in the age of whole-genome sequencing. Nat. Rev. Genet. 2015, 16, 275–284. [Google Scholar] [CrossRef]
- Hallman, D.M.; Boerwinkle, E.; Gonzalez, V.H.; Klein, B.E.; Klein, R.; Hanis, C.L. A genome-wide linkage scan for diabetic retinopathy susceptibility genes in Mexican Americans with type 2 diabetes from Starr County, Texas. Diabetes 2007, 56, 1167–1173. [Google Scholar] [CrossRef] [Green Version]
- Norrgard, K. Genetic Variation and Disease: GWAS. Nat. Educ. 2008, 1, 87. [Google Scholar]
- Genome-Wide Association Studies Fact Sheet. Available online: https://www.genome.gov/about-genomics/fact-sheets/Genome-Wide-Association-Studies-Fact-Sheet (accessed on 30 June 2021).
- Haines, J.L.; Hauser, M.A.; Schmidt, S.; Scott, W.K.; Olson, L.M.; Gallins, P.; Spencer, K.L.; Kwan, S.Y.; Noureddine, M.; Gilbert, J.R. Complement factor H variant increases the risk of age-related macular degeneration. Science 2005, 308, 419–421. [Google Scholar]
- Tony Burdett, E.H.D.W.S.E.-E.B.I.N. GWAS Catalog. Available online: https://www.ebi.ac.uk/gwas/search?query=diabetic%20retinopathy (accessed on 30 June 2021).
- Ban, N.; Yamada, Y.; Someya, Y.; Ihara, Y.; Adachi, T.; Kubota, A.; Watanabe, R.; Kuroe, A.; Inada, A.; Miyawaki, K. Activating transcription factor-2 is a positive regulator in CaM kinase IV-induced human insulin gene expression. Diabetes 2000, 49, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Simó-Servat, O.; Hernández, C.; Simó, R. Genetics in diabetic retinopathy: Current concepts and new insights. Curr. Genom. 2013, 14, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Forrest, I.S.; Chaudhary, K.; Paranjpe, I.; Vy, H.M.T.; Marquez-Luna, C.; Rocheleau, G.; Saha, A.; Chan, L.; Van Vleck, T.; Loos, R.J. Genome-wide polygenic risk score for retinopathy of type 2 diabetes. Hum. Mol. Genet. 2021, 30, 952–960. [Google Scholar] [CrossRef]
Figure 1.
Schematic showing pathogenic mechanisms of diabetic retinopathy (DR).


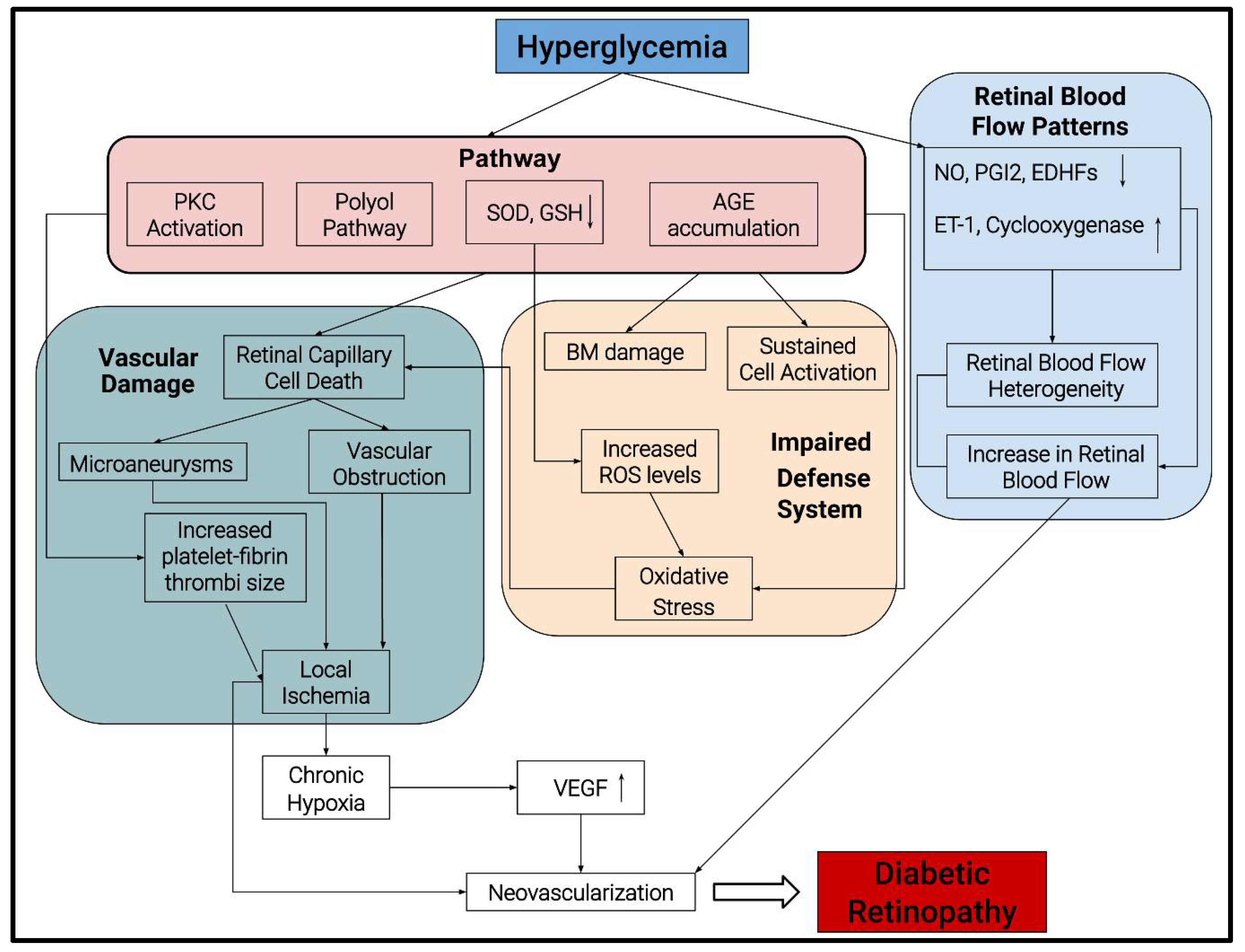{kind=link}
Table 1.
Candidate gene studies in DR.
| Candidate Gene | Gene Location | Online Mendelian Inheritance in Man (OMIM) Entry | Polymorphisms | Effect on DR | References |
|---|---|---|---|---|---|
| Aldose Reductase (AKR1B1) | 7q33 | 103880 | rs759853 | Protection from DR in T1D patients | [29] |
| Z-2 Z Z+2 | Risk for DR in T1 and T2D Protection from DR in T2D | [26] | |||
| rs9640883 | Duration of diabetes T1D and T2D | [30] | |||
| Receptor for Advanced Glycation End Product (AGER) | 6p21.32 | 600214 | rs1800624 | Risk of DR T1D | [31] |
| rs2070600 | Risk of DR in T2D | [32] | |||
| Vascular Endothelial Growth Factor (VEGFA) | 6p21.1 | 192240 | rs3025039 rs3025021, rs13207351 rs2146323 rs2010963 rs25648, rs833061 rs2010963 | Risk of DR in T2D Risk of DR in T2D Risk of DR in T2D Risk of DR in T2D Risk of PDR in T2D | [33] |
| Endothelial Nitric Oxide Synthase (NOS3) | 7q36.1 | 163729 | rs869109213 rs2070744 | Risk of DR in T2D | [34] |
| Angiotensin-I Converting Enzyme (ACE) | 17q23.3 | 106180 | rs1799752 rs4343 | Risk of DR in T2D Risk of DR in T2D | [35,36] |
| Erythropoietin (EPO) | 7q22.1 | 133170 | rs551238 rs1617640 rs507392 | Risk of DR | [37,38,39] |
| Calcium channel voltage dependent beta-2 sub unit (CACNB2) | 10p12.33-p12.31 | 600003 | rs202152674 rs137886839 | Increased risk of PDR | [40] |
| Intergenic locus in between AKT3 and ZNF238 | 1:24401312 | rs476141 | Increased risk of DR | [41] | |
| Caclium/Calmodulin-Dependent Protein Kinase IV (CAMK4) | 5q22.1 | 114080 | rs2300782 | Increased risk of DR | [42] |
| Formin 1 (FMN1) | 15q13.3 | 136535 | rs10519765 | Increased risk of DR | [42] |
| Growth factor receptor bound-2 (GRB2) | 17q25.1 | 108355 | rs9896052 | Sight threatening DR | [43] |
| Valosin -containing protein like (NVL) | 1q42.11 | 602426 | rs142293996 | Increased risk of DR | [44] |
| STT3 Oligosaccharyltransferase Complex Catalytic Subunit B (STT3B) | 3p23 | 608605 | rs12630354 | Increased risk of DR | [45] |
| Paralemmin-2 (PALM2AKAP2) | 9q31.3 | 604582 | rs140508424 | Increased risk of DR | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bhatwadekar, A.D.; Shughoury, A.; Belamkar, A.; Ciulla, T.A. Genetics of Diabetic Retinopathy, a Leading Cause of Irreversible Blindness in the Industrialized World. Genes 2021, 12, 1200. https://doi.org/10.3390/genes12081200
AMA Style
Bhatwadekar AD, Shughoury A, Belamkar A, Ciulla TA. Genetics of Diabetic Retinopathy, a Leading Cause of Irreversible Blindness in the Industrialized World. Genes. 2021; 12(8):1200. https://doi.org/10.3390/genes12081200
Chicago/Turabian StyleBhatwadekar, Ashay D., Aumer Shughoury, Ameya Belamkar, and Thomas A. Ciulla. 2021. "Genetics of Diabetic Retinopathy, a Leading Cause of Irreversible Blindness in the Industrialized World" Genes 12, no. 8: 1200. https://doi.org/10.3390/genes12081200
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.