Cytogenetic Analysis of the Asian Box Turtles of the Genus Cuora (Testudines, Geoemydidae)

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Taxonomic Verification
2.3. Chromosomes Preparation, Staining, and C-Banding
2.4. Giemsa Staining and Karyogram Construction
2.5. Fluorescence In Situ Hybridization (FISH)
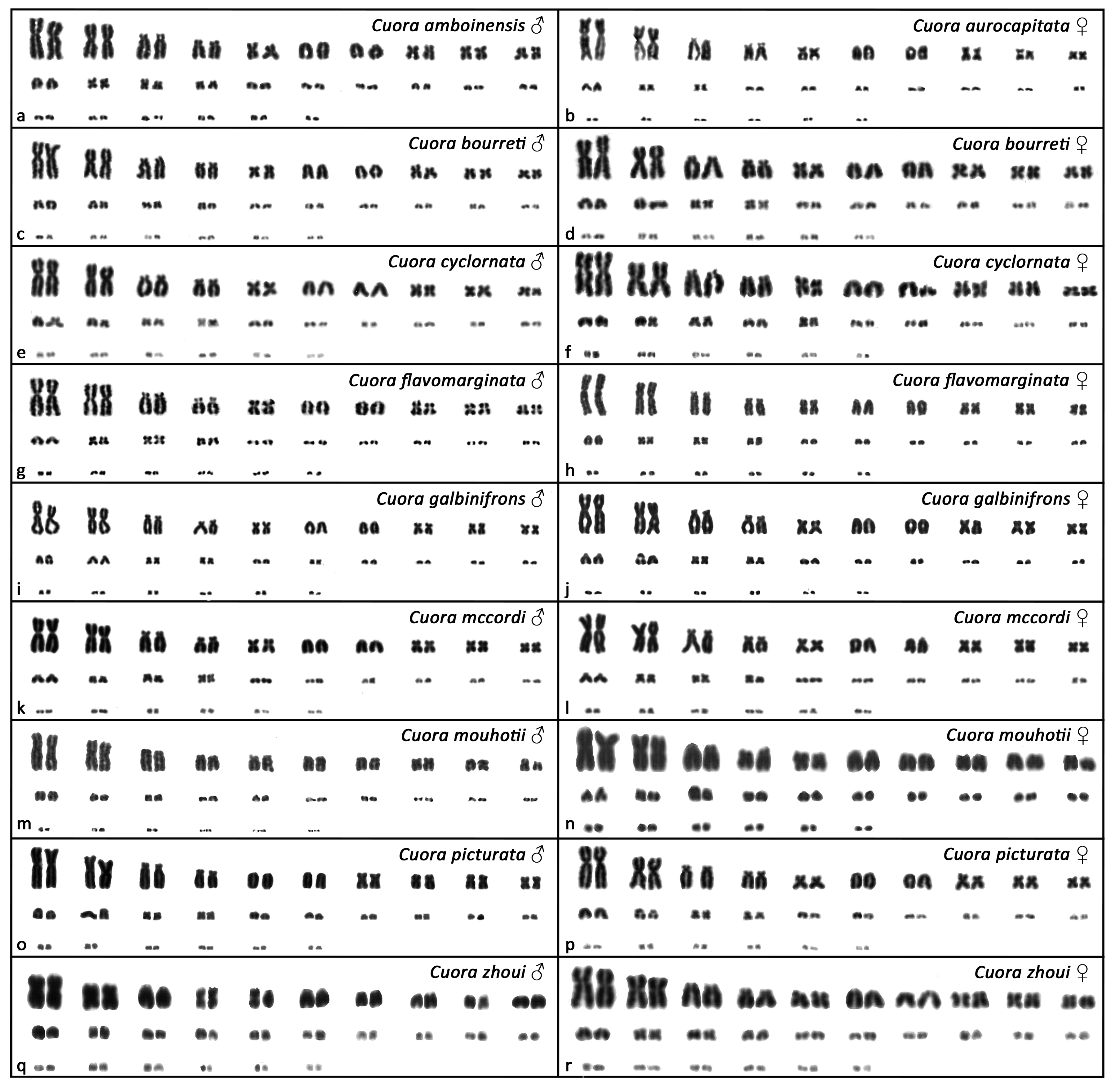
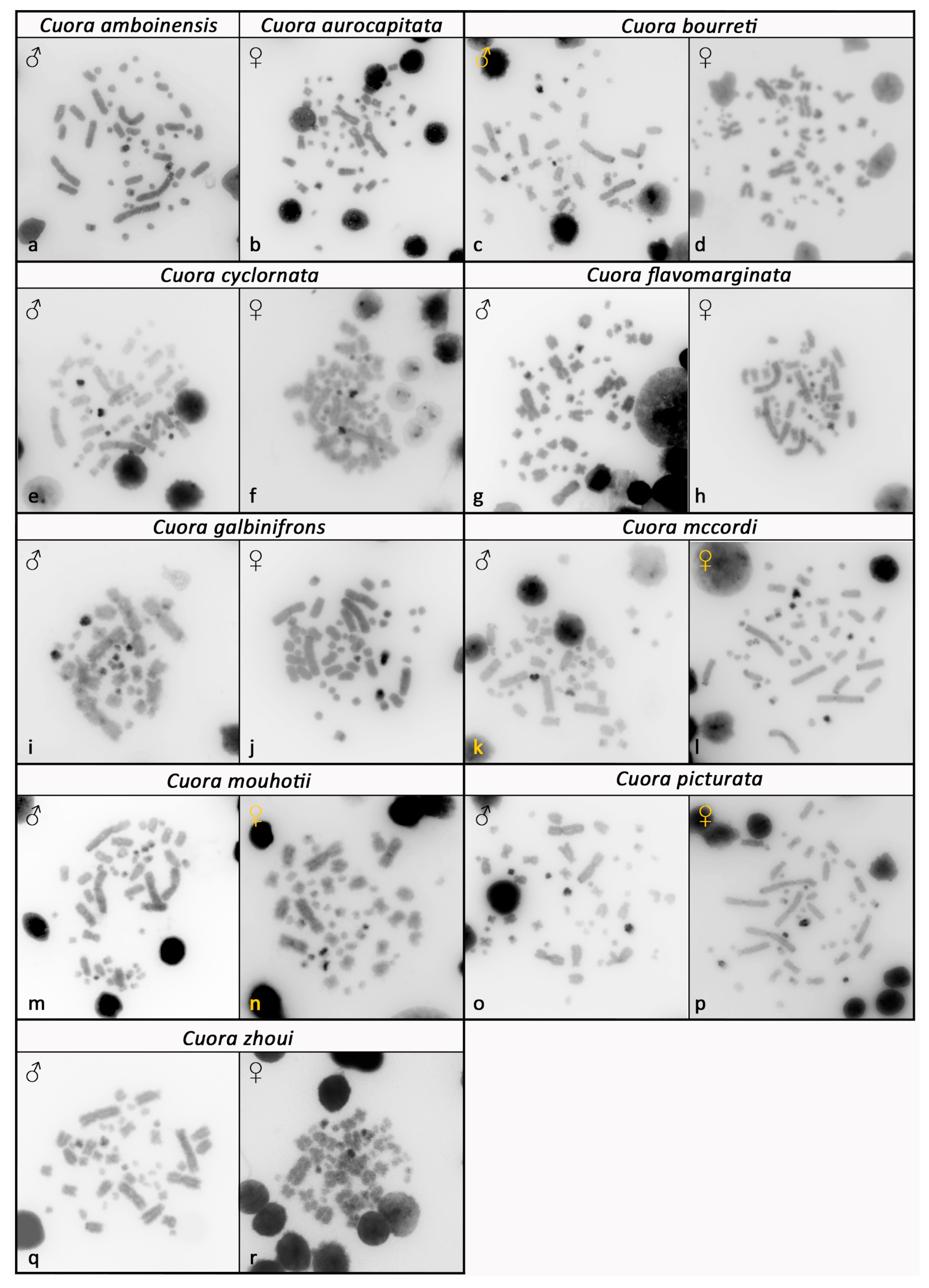
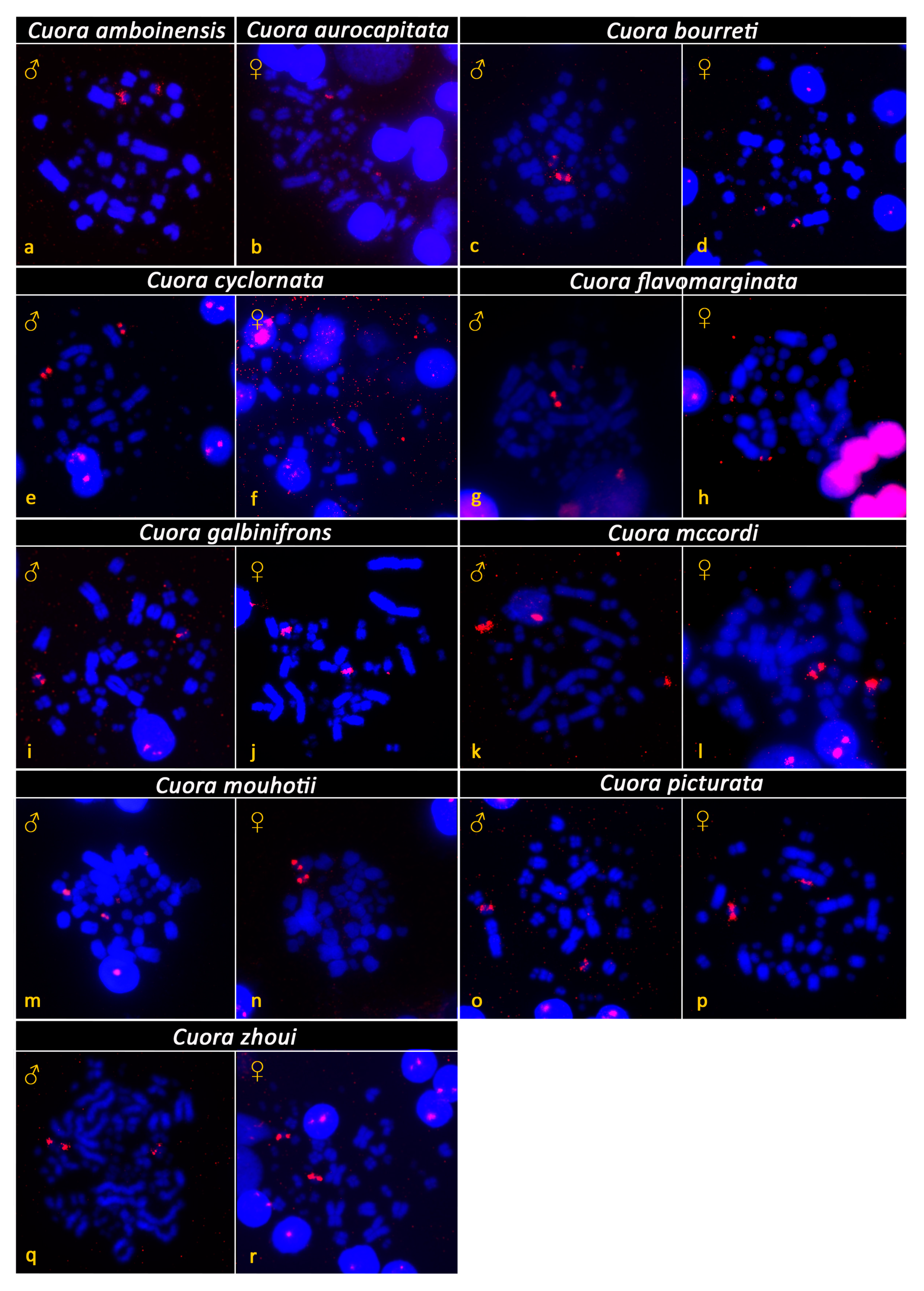
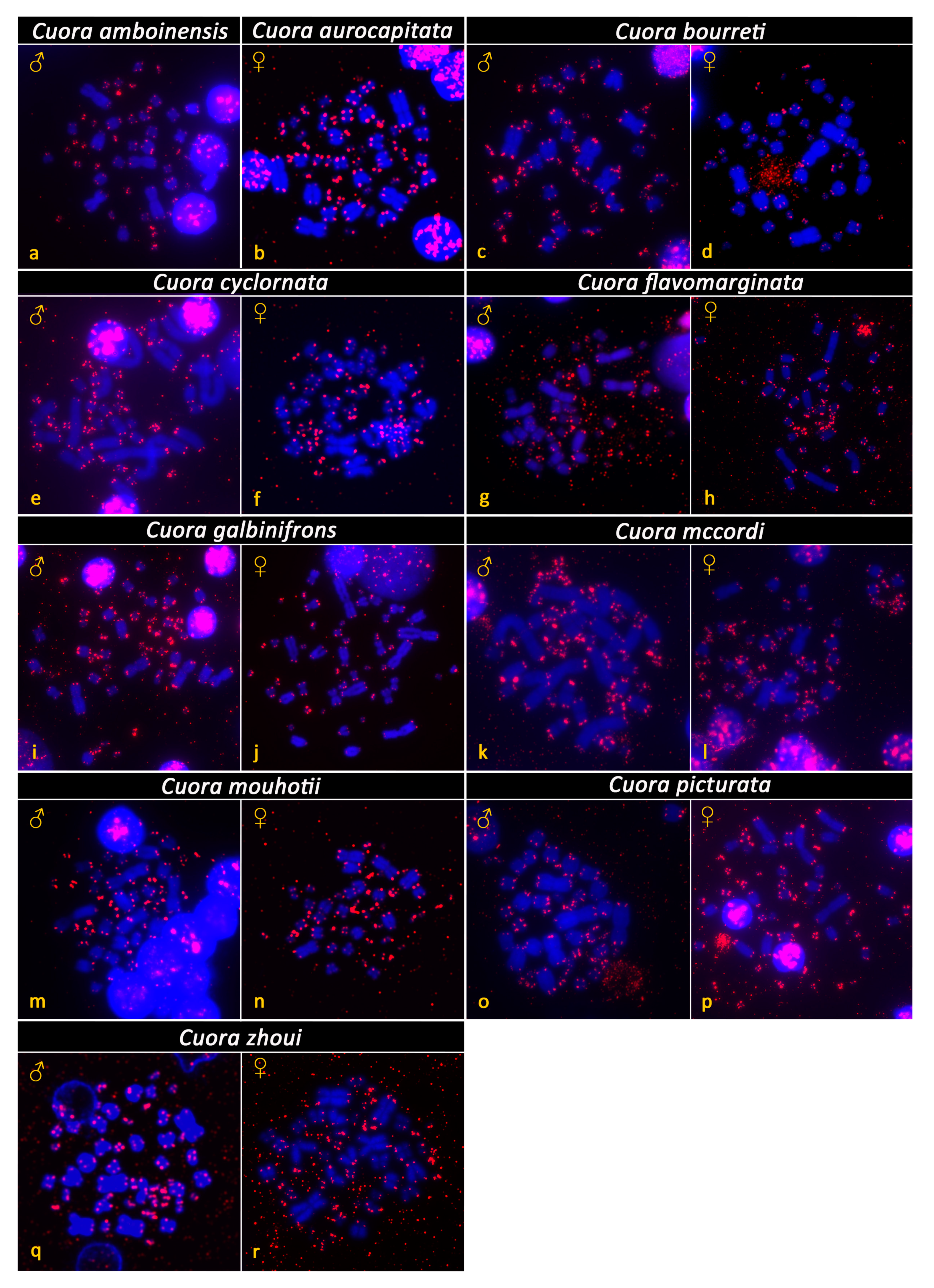
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montiel, E.E.; Badenhorst, D.; Lee, L.S.; Literman, R.; Trifonov, V.; Valenzuela, N. Cytogenetic insights into the evolution of chromosomes and sex determination reveal striking homology of turtle sex chromosomes to amphibian autosomes. Cytogenet. Genome Res. 2016, 148, 292–304. [Google Scholar] [CrossRef]
- Clemente, L.; Mazzoleni, S.; Pensabene Bellavia, E.; Augstenová, B.; Auer, M.; Praschag, P.; Protiva, T.; Velenský, P.; Wagner, P.; Fritz, U.; et al. Interstitial telomeric repeats are rare in turtles. Genes 2020, 11, 657. [Google Scholar] [CrossRef] [PubMed]
- Olmo, E.; Signorino, G.G. Chromorep: A Reptile Chromosomes Database. Available online: http://chromorep.univpm.it (accessed on 14 April 2020).
- Rhodin, A.G.J.; Stanford, C.B.; Van Dijk, P.P.; Eisemberg, C.; Luiselli, L.; Mittermeier, R.A.; Hudson, R.; Horne, B.D.; Goode, E.V.; Kuchling, G.; et al. Global conservation status of turtles and tortoises (order Testudines). Chelonian Conserv. Biol. 2018, 17, 135–161. [Google Scholar] [CrossRef]
- Farkas, B.; Ziegler, T.; Pham, C.T.; Ong, A.V.; Fritz, U. A new species of Pelodiscus from northeastern Indochina (Testudines, Trionychidae). ZooKeys 2019, 824, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Loc-Barragán, J.A.; Reyes-Velasco, J.; Woolrich-Pina, G.A.; Grunwald, C.I.; de Anaya, M.V.; Rangel-Mendoza, J.A.; López-Luna, M.A. A new species of mud turtle of genus Kinosternon (Testudines: Kinosternidae) from the Pacific coastal plain of northwestern Mexico. Zootaxa 2020, 3885, 509–529. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Ramírez, M.; Caballero, S.; Morales-Betancourt, M.A.; Lasso, C.A.; Amaya, L.; Martínez, J.G.; das Neves Silva Viana, M.; Vogt, R.C.; Farias, I.P.; Hrbek, T.; et al. Genomic analyses reveal two species of the matamata (Testudines: Chelidae: Chelus spp.) and clarify their phylogeography. Mol. Phylogenet. Evol. 2020, 148, 106823. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Freed, P.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 13 April 2020).
- Ventura, K.; Moreira, C.N.; Moretti, R.; Yonenaga-Yassuda, Y.; Rodrigues, M.T. The lowest diploid number in Testudines: Banding patterns, telomeric and 45s rDNA FISH in Peltocephalus dumerilianus, 2n = 26 and FN = 52 (Pleurodira, Podocnemididae). Genet. Mol. Biol. 2014, 37, 61–63. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, M.G.; Bastos, C.E.M.C.; Nagamachi, C.Y.; Pieczarka, J.C.; Vicari, M.R.; Noronha, R.C.R. Physical mapping of repetitive DNA suggests 2n reduction in Amazon turtles Podocnemis (Testudines: Podocnemididae). PLoS ONE 2018, 13, e0197536. [Google Scholar] [CrossRef] [Green Version]
- Stock, A.D.; Burnham, D.B.; Hsu, T.C. Giemsa banding of meiotic chromosomes with description of a procedure for cytological preparations from solid tissues. Cytogenetics 1972, 11, 534–539. [Google Scholar] [CrossRef]
- Killebrew, F.C. Mitotic chromosomes of turtles. IV. The Emydidae. Tex. J. Sci. 1977, 29, 245–253. [Google Scholar]
- Bista, B.; Valenzuela, N. Turtle Insights into the evolution of the reptilian karyotype and the genomic architecture of sex determination. Genes 2020, 11, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, N.; Adams, D.C. Chromosome number and sex determination coevolve in turtles. Evolution 2011, 65, 1808–1813. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Turtles of the genera Geoemyda and Pangshura (Testudines: Geoemydidae) lack differentiated sex chromosomes: The end of a 40-year error cascade for Pangshura. PeerJ 2019, 7, e6241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson Pokorná, M.; Kratochvíl, L. What was the ancestral sex-determining mechanism in amniote vertebrates? Biol. Rev. 2016, 91, 1–12. [Google Scholar] [CrossRef]
- Straková, B.; Rovatsos, M.; Kubička, L.; Kratochvíl, L. Evolution of sex determination in amniotes: Did stress and sequential hermaphroditism produce environmental determination? BioEssays 2020, 42, e2000050. [Google Scholar] [CrossRef]
- Badenhorst, D.; Hillier, L.W.; Literman, R.; Montiel, E.E.; Radhakrishnan, S.; Shen, Y.; Minx, P.; Janes, D.E.; Warren, W.C.; Edwards, S.V.; et al. Physical mapping and refinement of the painted turtle genome (Chrysemys picta) inform amniote genome evolution and challenge turtle-bird chromosomal conservation. Genome Biol. Evol. 2015, 7, 2038–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bull, J.J.; Moon, R.G.; Legler, J.M. Male heterogamety in kinosternid turtles (genus Staurotypus). Cytogenet. Genome Res. 1974, 13, 419–425. [Google Scholar] [CrossRef]
- Ezaz, T.; Valenzuela, N.; Grützner, F.; Miura, I.; Georges, A.; Burke, R.L.; Graves, J.A. An XX/XY sex microchromosome system in a freshwater turtle, Chelodina longicollis (Testudines: Chelidae) with genetic sex determination. Chromosome Res. 2006, 14, 139–150. [Google Scholar] [CrossRef]
- Martinez, P.A.; Ezaz, T.; Valenzuela, N.; Georges, A.; Marshall Graves, J.A. An XX/XY heteromorphic sex chromosome system in the Australian chelid turtle Emydura macquarii: A new piece in the puzzle of sex chromosome evolution in turtles. Chromosome Res. 2008, 16, 815–825. [Google Scholar] [CrossRef]
- Kawagoshi, T.; Nishida, C.; Matsuda, Y. The origin and differentiation process of X and Y chromosomes of the black marsh turtle (Siebenrockiella crassicollis, Geoemydidae, Testudines). Chromosome Res. 2012, 20, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Literman, R.; Radhakrishnan, S.; Tamplin, J.; Burke, R.; Dresser, C.; Valenzuela, N. Development of sexing primers in Glyptemys insculpta and Apalone spinifera turtles uncovers an XX/XY sex-determining system in the critically-endangered bog turtle Glyptemys muhlenbergii. Conserv. Genet. Res. 2017, 9, 651–658. [Google Scholar] [CrossRef]
- Rovatsos, M.; Praschag, P.; Fritz, U.; Kratochvil, L. Stable Cretaceous sex chromosomes enable molecular sexing in softshell turtles (Testudines: Trionychidae). Sci. Rep. 2017, 7, 42150. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lin, J.W.; Tseng, S.P.; Chen, T.S.; Lin, S.M. Human disturbance as a possible cause of genetic introgression from exotic into native Mauremys turtles. Anim. Conserv. 2019, 22, 556–567. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Sex is determined by XX/XY sex chromosomes in Australasian side-necked turtles (Testudines: Chelidae). Sci. Rep. 2020, 10, 4276. [Google Scholar] [CrossRef] [Green Version]
- Viana, P.F.; Feldberg, E.; Cioffi, M.B.; de Carvalho, V.T.; Menezes, S.; Vogt, R.C.; Liehr, T.; Ezaz, T. The Amazonian red side-necked turtle Rhinemys rufipes (Spix, 1824) (Testudines, Chelidae) has a GSD sex-determining mechanism with an ancient XY sex microchromosome System. Cells 2020, 9, 2088. [Google Scholar] [CrossRef]
- Sharma, G.P.; Kaur, P.; Nakhasi, U. Female heterogamety in the Indian cryptodiran chelonian, Kachuga smithi Gray. In Dr. B.S. Chauhan Commemoration Volume; Tiwari, K.K., Srivastava, C.B., Eds.; Zoological Society of India: Bhubaneshwar, Orissa, India, 1975; pp. 359–368. [Google Scholar]
- Rhodin, A.G.J.; Iverson, J.B.; Bour, R.; Fritz, U.; Georges, A.; Shaffer, H.B.; Van Dijk, P.P.; TTWG—Turtle Taxonomy Working Group. Turtles of the world: Annotated checklist and atlas of taxonomy, synonymy, distribution and conservation status (8th edition). Chelonian Res. Monogr. 2017, 7, 1–202. [Google Scholar]
- Colston, T.J.; Kulkarni, P.; Jetz, W.; Pyron, R.A. Phylogenetic and spatial distribution of evolutionary diversification, isolation, and threat in turtles and crocodilians (non-avian archosauromorphs). BMC Evol. Biol. 2020, 20, 81. [Google Scholar] [CrossRef]
- Bickham, J.W. A cytosystematic study of turtles in the genera Clemmys, Mauremys and Sacalia. Herpetologica 1975, 31, 198–204. [Google Scholar]
- Bickham, J.; Carr, J. Taxonomy and phylogeny of the higher categories of Cryptodiran turtles based on a cladistic analysis of chromosomal data. Copeia 1983, 4, 918–932. [Google Scholar] [CrossRef] [Green Version]
- Carr, J.L.; Bickham, J.W. Phylogenetic implications of karyotype variation in the Batagurinae (Testudines: Emydidae). Genetica 1986, 70, 89–106. [Google Scholar] [CrossRef]
- Guo, C.; Nie, L.; Wang, M. A cytogenetic study of four species of turtle from China. Acta Genet. Sin. 1995, 22, 40–45. [Google Scholar] [PubMed]
- Siripiyasing, P.; Tanomtong, A.; Jumrusthanasan, S.; Patawang, I.; Phimphan, S.; Sanoamuang, L.O. First cytogenetic study of Malayan snail-eating turtle, Malayemys macrocephala (Testudines, Geoemydidae) in Thailand. Cytologia 2013, 78, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, M.G.; Souza, L.F.; Vicari, M.R.; de Bastos, C.; de Sousa, J.V.; Nagamachi, C.Y.; Pieczarka, J.C.; Martins, C.; Noronha, R. Molecular cytogenetics characterization of Rhinoclemmys punctularia (Testudines, Geoemydidae) and description of a Gypsy-H3 association in its genome. Gene 2020, 738, 144477. [Google Scholar] [CrossRef]
- Yadollahvand, R.; Kami, H.G.; Kalbassi, M.R. Cytogenetic characterisation of the Caspian Pond Turtle, Mauremys caspica in Golestan and Mazandaran provinces, Iran (Reptilia: Testudines). Zool. Middle East 2013, 59, 214–219. [Google Scholar] [CrossRef]
- Ewert, M.A.; Etchberger, C.R.; Nelson, C.E. Turtle sex-determining modes and TSD patterns, and some TSD pattern correlates. In Temperature-Dependent Sex Determination in Vertebrates; Valenzuela, N., Lance, V.A., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2004; pp. 21–32. [Google Scholar]
- Farrell, R. Temperature sex determination (TSD) project for the yellow-margined box turtle, Cuora flavomarginata. In TSA Turtle Survival Alliance; 2007; pp. 1–46. Available online: https://turtlesurvival.org (accessed on 17 December 2020).
- Stanford, C.B.; Iverson, J.B.; Rhodin, A.G.; van Dijk, P.P.; Mittermeier, R.A.; Kuchling, G.; Berry, K.H.; Bertolero, A.; Bjorndal, K.A.; Blanck, T.E.; et al. Turtles and tortoises are in trouble. Curr. Biol. 2020, 30, R721–R735. [Google Scholar] [CrossRef] [PubMed]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 17 December 2020).
- Gorman, G.C. The chromosomes of the Reptilia, a cytotaxonomic interpretation. In Cytotaxonomy and Vertebrate Evolution; Chiarelli, A.B., Capanna, E., Eds.; Acad. Press: London, UK; New York, NY, USA, 1973; pp. 347–424. [Google Scholar]
- Burbrink, F.T.; Lawson, R.; Slowinski, J.B. Mitochondrial DNA phylogeography of the polytypic North American ratsnake (Elaphe obsoleta): A critique of the subspecies concept. Evolution 2000, 54, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- FinchTV v1.4.0. Available online: https://digitalworldbiology.com/FinchTV (accessed on 25 December 2020).
- Geneious Prime 2020. Available online: https://www.geneious.com (accessed on 25 December 2020).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Endow, S.A. Polytenization of the ribosomal genes on the X and Y chromosomes of Drosophila melanogaster. Genetics 1982, 100, 375–385. [Google Scholar]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef] [Green Version]
- Rovatsos, M.T.; Marchal, J.A.; Romero-Fernández, I.; Fernández, F.J.; Giagia-Athanosopoulou, E.B.; Sánchez, A. Rapid, independent, and extensive amplification of telomeric repeats in pericentromeric regions in karyotypes of arvicoline rodents. Chromosome Res. 2011, 19, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, H.B.; Minx, P.; Warren, D.E.; Shedlock, A.M.; Thomson, R.C.; Valenzuela, N.; Abramyan, J.; Amemiya, C.T.; Badenhorst, D.; Biggar, K.K.; et al. The western painted turtle genome, a model for the evolution of extreme physiological adaptations in a slowly evolving lineage. Genome Biol. 2013, 14, R28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tollis, M.; DeNardo, D.F.; Cornelius, J.A.; Dolby, G.A.; Edwards, T.; Henen, B.T.; Karl, A.E.; Murphy, R.W.; Kusumi, K. The Agassiz’s desert tortoise genome provides a resource for the conservation of a threatened species. PLoS ONE 2017, 12, e0177708. [Google Scholar] [CrossRef] [PubMed]
- Brian Simison, W.; Parham, J.F.; Papenfuss, T.J.; Lam, A.W.; Henderson, J.B. An annotated chromosome-level reference genome of the Red-Eared Slider Turtle (Trachemys scripta elegans). Genome Biol. Evol. 2020, 12, 456–462. [Google Scholar] [CrossRef]
- Bickham, J.W.; Baker, R.J. Chromosome homology and evolution of emydid turtles. Chromosoma 1976, 54, 201–219. [Google Scholar] [CrossRef] [PubMed]
- De Smet, W.H.O. The chromosomes of 11 species of Chelonia (Reptilia). Acta Zool. Pathol. Antverp. 1978, 70, 15–34. [Google Scholar]
- Haiduk, M.W.; Bickham, J.W. Chromosomal homologies and evolution of testudinoid turtles with emphasis on the systematic placement of Platysternon. Copeia 1982, 1982, 60–66. [Google Scholar] [CrossRef]
- Scardino, R.; Mazzoleni, S.; Rovatsos, M.; Vecchioni, L.; Dumas, F. Molecular cytogenetic characterization of the Sicilian endemic pond turtle Emys trinacris and the yellow-bellied slider Trachemys scripta scripta (Testudines, Emydidae). Genes 2020, 11, 702. [Google Scholar] [CrossRef]
- Nanda, I.; Schrama, D.; Feichtinger, W.; Haaf, T.; Schartl, M.; Schmid, M. Distribution of telomeric (TTAGGG)(n) sequences in avian chromosomes. Chromosoma 2002, 111, 215–227. [Google Scholar] [CrossRef]
- Rovatsos, M.; Kratochvíl, L.; Altmanová, M.; Johnson Pokorná, M. Interstitial telomeric motifs in squamate reptiles: When the exceptions outnumber the rule. PLoS ONE 2015, 10, e0134985. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, E.H.; Habermann, F.A.; Lacerda, O.; Sbalqueiro, I.J.; Wienberg, J.; Müller, S. Chromosome reshuffling in birds of prey: The karyotype of the world’s largest eagle (Harpy eagle, Harpia harpyja) compared to that of the chicken (Gallus gallus). Chromosoma 2005, 114, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Galgon, F.; Fritz, U. Captive bred hybrids between Chinemys reevesii (Gray, 1831) and Cuora amboinensis kamaroma Rummler & Fritz, 1991 (Testudines: Geoemydidae). Herpetozoa 2002, 15, 137–148. [Google Scholar]
- Wink, M.; Guicking, D.; Fritz, U. Molecular evidence for hybrid origin of Mauremys iversoni Pritchard et McCord 1991, and Mauremys pritchardi McCord, 1997. Zool. Abh. Mus. Tierkd. Dresd. 2001, 51, 41–49. [Google Scholar]
- Buskirk, J.R.; Parham, J.F.; Feldman, C.R. On the hybridization between two distantly related Asian turtles (Testudines: Sacalia x Mauremys). Salamandra 2005, 41, 21–26. [Google Scholar]
- Schilde, M.; Barth, D.; Fritz, U. An Ocadia sinensis x Cyclemys shanensis hybrid. Asian Herpetol. Res. 2004, 10, 120–125. [Google Scholar]
- Stuart, B.L.; Parham, J.F. Recent hybrid origin of three rare Chinese turtles. Conserv. Genet. 2007, 8, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Wang, L.; Nie, L.; Huang, Z.; Jiang, Y.; Jing, W.; Liu, L. Interspecific hybridization between Mauremys reevesii and Mauremys sinensis: Evidence from morphology and DNA sequence data. Afr. J. Biotechnol. 2011, 10, 6716–6724. [Google Scholar]
- Suzuki, D.; Yabe, T.; Hikida, T. Hybridization between Mauremys japonica and Mauremys reevesii inferred by nuclear and mitochondrial DNA analyses. J. Herpetol. 2013, 48, 445–454. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ♂ | ♀ |
|---|---|---|
| Cuora amboinensis | 1 | - |
| Cuora aurocapitata | - | 3 |
| Cuora bourreti | 1 | 1 |
| Cuora cyclornata | 1 | 1 |
| Cuora flavomarginata | 1 | 1 |
| Cuora galbinifrons | 2 | 2 |
| Cuora mccordi | 1 | 1 |
| Cuora mouhotii | 2 | 3 |
| Cuora picturata | 1 | 1 |
| Cuora zhoui | 2 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clemente, L.; Mazzoleni, S.; Pensabene, E.; Protiva, T.; Wagner, P.; Fritz, U.; Kratochvíl, L.; Rovatsos, M. Cytogenetic Analysis of the Asian Box Turtles of the Genus Cuora (Testudines, Geoemydidae). Genes 2021, 12, 156. https://doi.org/10.3390/genes12020156
Clemente L, Mazzoleni S, Pensabene E, Protiva T, Wagner P, Fritz U, Kratochvíl L, Rovatsos M. Cytogenetic Analysis of the Asian Box Turtles of the Genus Cuora (Testudines, Geoemydidae). Genes. 2021; 12(2):156. https://doi.org/10.3390/genes12020156
Chicago/Turabian StyleClemente, Lorenzo, Sofia Mazzoleni, Eleonora Pensabene, Tomáš Protiva, Philipp Wagner, Uwe Fritz, Lukáš Kratochvíl, and Michail Rovatsos. 2021. "Cytogenetic Analysis of the Asian Box Turtles of the Genus Cuora (Testudines, Geoemydidae)" Genes 12, no. 2: 156. https://doi.org/10.3390/genes12020156