Common Features of the Pericentromere and Nucleolus


Department of Biology, University of North Carolina at Chapel Hill, Chapel Hill, NC 27599-3280, USA
*
Author to whom correspondence should be addressed.
Genes 2019, 10(12), 1029; https://doi.org/10.3390/genes10121029
Submission received: 25 November 2019
/
Revised: 5 December 2019
/
Accepted: 7 December 2019
/
Published: 10 December 2019
(This article belongs to the Special Issue Chromosome Segregation Defects in the Origin of Genomic Instability)
{kind=link}
Abstract
:Both the pericentromere and the nucleolus have unique characteristics that distinguish them amongst the rest of genome. Looping of pericentromeric DNA, due to structural maintenance of chromosome (SMC) proteins condensin and cohesin, drives its ability to maintain tension during metaphase. Similar loops are formed via condensin and cohesin in nucleolar ribosomal DNA (rDNA). Condensin and cohesin are also concentrated in transfer RNA (tRNA) genes, genes which may be located within the pericentromere as well as tethered to the nucleolus. Replication fork stalling, as well as downstream consequences such as genomic recombination, are characteristic of both the pericentromere and rDNA. Furthermore, emerging evidence suggests that the pericentromere may function as a liquid–liquid phase separated domain, similar to the nucleolus. We therefore propose that the pericentromere and nucleolus, in part due to their enrichment of SMC proteins and others, contain similar domains that drive important cellular activities such as segregation, stability, and repair.
1. Introduction
During cell division, the centromere functions as an essential genetic locus for ensuring faithful chromosome segregation. Microtubules connect to each centromere on sister chromatids via a proteinaceous complex called the kinetochore. Eukaryotic centromeres range in complexity from simple point centromeres in budding yeast to regional centromeres in fission yeast, plants, and mammals [1,2]. In budding yeast, the pericentromere is defined as the cohesin- and condensin-enriched region spanning 30–50 kb on either side of the conserved centromeric sequence [3,4,5]. However, in regional centromeres, which are enriched in heterochromatic alpha satellite repeats, there are multiple sites of microtubule attachment. Despite these differences, the interkinetochore distance is conserved among eukaryotes [6], suggesting an important conservation of centromere mechanics.
The nucleolus, as the site of ribosome biogenesis, arises from the compartmentalization of ribosomal DNA (rDNA). In budding yeast, rDNA is localized to chromosome XII, consisting of 150 tandem repeats of ~ 9 kb each. rDNA repeats in higher eukaryotes such as human are more dispersed, with repeats on five different chromosomes, but nonetheless they have similar properties regarding rDNA structure [7].
The pericentromere and nucleolus have a number of similarities between their domains. These include condensin/cohesin localization and proteins that regulate the formation of DNA loops. Transfer RNA (tRNA) genes, enriched in condensin and cohesin, are also located within the pericentromere region and tethered to the nucleolus. Replication fork stalling occurs across both centromeres and rDNA repeats, along with a propensity for control of genomic recombination. Lastly, while the nucleolus is a well-identified liquid phase separated region, emerging evidence suggests that the pericentromere may have similar properties. Here, we discuss in detail these commonalities between the pericentromere and the nucleolus.
2. Common Features in Both the Pericentromere and the Nucleolus
2.1. DNA Loops Are Enriched in the Pericentromere and Nucleolus
While DNA looping was first observed in 1906 in salamander eggs [8], DNA loops have since been thoroughly documented in a number of different species [9]. These loops can potentially regulate many functions within the cell, including transcription, recombination, and replication [6,10]. Though DNA loops are present in various locations throughout the genome, both the pericentromere and the nucleolus are regions of high loop density that are controlled by unique protein interactors.
In the pericentromere, highly looped DNA has been found in budding yeast (Saccharomyces cerevisiae) [11,12] as well as in multicellular organisms such as Xenopus laevis [13] and chicken cells [14]. In budding yeast, centromeres from the 16 chromosomes cluster together into a disc approximately 50 nm by 250 nm, which connects to the microtubule plus-ends (for review, see [6]). From here, the pericentric region consists of protruding intramolecular centromere loops, or C-loops, that are formed by loss of sister chromatid cohesion, with radial sub-loops forming off of each C-loop [11,12,15,16] (Figure 1A). The structural maintenance of chromosome (SMC) proteins condensin and cohesin have both been implicated in forming and maintaining these pericentromeric loops in budding yeast [11,12,17,18]. It has been proposed that these loops play an important role in maintaining the mechanics of the pericentromere [15,16], and may provide a mechanism for chromatin condensation [19,20]. Furthermore, the loops generated between the repeat sequences found in higher eukaryotic centromeres may also facilitate recombination [21].
The rDNA present within the nucleolus also features characteristic looping behavior (Figure 1B). In budding yeast, fluorescence in situ hybridization (FISH) staining of the entire rDNA indicates a loop-like structure in nucleoli [22]. Similar to the pericentromere, SMC proteins cohesin and condensin have both been implicated in rDNA loop formation. Cohesin mutations result in reduced looping of the rDNA genes for 35S and 5S in budding yeast [23], which may affect their transcription [24]. Live cell imaging of rDNA in condensin mutants further implicates condensin in the dynamics of loop formation, with time-lapsed imaged mutants displaying a delay compared to wild-type in loop formation [25]. Condensin-mediated loop extrusion of rDNA is further supported by globally generated Hi-C contact maps [26]. Both nucleolar transcription factor 1 (UBF), a mammalian protein containing high mobility group (HMG) dox domains, and its yeast homolog, high mobility protein 1 (Hmo1), bind preferentially to actively transcribed rDNA genes [27,28] and are enriched in the nucleolus [29]. Electron spectroscopic imaging in Xenopus suggests that UBF dimers bend approximately 150 bp rDNA into a loop formation [30,31]. While Hmo1 has yet to specifically be identified in the looping feature of rDNA, the reliance on the HMG box for the looping activity of UBF suggests that Hmo1 may have a similar role [32]. Also in Xenopus, immunostaining indicates that Pol III sites (which bind to nucleolar tRNA genes) are localized to DNA loops [33], suggesting an additional nucleolar DNA looping site.
2.2. SMC Proteins in the Pericentromere and Nucleolus Display Common DNA Regulatory Roles
Structural maintenance of chromosome (SMC) protein complexes, such as condensin and cohesin, are essential to regulation of chromosome function and structure. Condensin and cohesin are enriched both in the pericentromere [3,42,50,51] and in the nucleolus [52,53] in a variety of different organisms, suggesting an important conservation of function. These proteins play an important role in cohesion between sister chromatids, chromosome segregation, DNA replication, DNA damage repair, and DNA loop formation. In eukaryotes, SMC proteins form heterodimers, creating a V-shaped molecule with a variable conformation [54]. While there are two different identified condensin complexes (condensin I and II), only condensin I has been found in fungi such as budding and fission yeast [20].
The SMC heterodimer in condensin I/II consists of Smc2/Smc4. Non-SMC subunits in condensin I includes chromosome associated protein H (CAP-H; a member of the kleisin protein family), and HEAT repeat (which consists of Huntingtin elongation factor 3 [EF3], protein phosphatase 2A [PP2A], and the yeast kinase, target of rapamycin 1 [TOR1])-containing chromosome associated protein D2 (CAP-D2) and chromosome associated protein G (CAP-G), whereas condensin II contains CAP-H2, CAP-D3, and CAP-G2 [20]. As an SMC protein complex, condensin has a number of different roles in regard to DNA regulation. As its name suggest, condensin is essential for chromosome condensation, or heterochromatin formation [52,55]. More recently, studies have found that condensin is involved in extruding pericentric DNA loops [12,17,18] as well as cross-linking DNA in trans [56]. Live imaging of yeast condensin along double-tethered λ -DNA also showed that condensin mediates loop extrusion [34]. The localization of condensin is further regulated by proteins such as the histone deacetylase Sir2. Sir2 contributes to the axial position of condensin in the pericentromere, in which condensin is located proximal to the yeast mitotic spindle. In yeast lacking Sir2, condensin becomes more radially displaced, distal to the spindle and perpendicular to the spindle axis, and can appear as a bilobed distribution similar to pericentric cohesin [17]. Condensin also has a role in positive supercoiling of DNA in both Xenopus [57] and budding yeast [58], which may promote proper segregation as positively supercoiled DNA is more resistant to pulling forces [6,59]. Meanwhile, in rDNA, FISH studies in budding yeast indicate that condensin is necessary for rDNA looping [43,44,45,46]. DNA replication fork blocking protein Fob1, a known rDNA binding protein, is responsible for loading condensin onto rDNA repeats in yeast [47].
Cohesin contains a Smc1/Smc3 heterodimer, as well as two other subunits: Scc1 (also referred to as Mcd1 or Rad21) and Scc3 (also known as SA) [54]. Cohesin can form a ring-like molecule [60,61] among many other possible configurations [62,63,64]. While first identified for its prominent role in sister chromatid cohesion [65], the exact mechanism of the cohesion ability of cohesin remains debated. Possible models include the ring model, in which a single cohesin molecule embraces both sister chromatids, the handcuff model, in which two cohesin rings on either sister chromatid bind together, and the bracelet model, in which a cohesin oligomer wraps around the sister chromatids [66]. Chromatin immunoprecipitation (ChIP) assays on yeast strains with mutated alleles of cohesin subunit Mcd1 also revealed that cohesin preferentially binds the pericentromere versus the chromosome arms [67]. In vivo studies using yeast indicate that during metaphase, cohesin is radially displaced from the pericentric DNA [11,12], which is dependent on its ability to passively diffuse along the chromosome [18], and suggests that it plays a role in linking the C-loops generated by condensin. However, other studies suggest a cohesin-mediated loop extrusion model, particularly during interphase [62,68]. In mammalian cells, ChIP-seq in combination with Hi-C indicates that cohesin is localized to topological associated domains (TADs), an indicator of loop formations [69], and further studies suggest cohesin may directly regulate these loops [70,71,72,73]. Outside of the pericentromere, cohesin is recruited to both tRNA genes and rDNA sites in a Sir2-dependent mechanism [48,74]. Cohesin is further involved in the cohesion of rDNA sister chromatids in budding yeast [65,75], and controls mitotic rDNA organization [76]. Mutations of cohesin are associated with both disorganization of the nucleolus and reduced looping of rDNA [23], concurrent with less rRNA production and subsequent protein translation [77], suggesting a particular role for cohesin at this locus.
Condensin and cohesin feature small ubiquitin-like modifier (SUMO) sites, or sumoylation sites [78], a reversible modification which affects their distribution in both the pericentromere and at rDNA sites. Deletion of Ulp2 (also referred to as Smt4), an isopeptidase that removes SUMO from proteins, causes a decrease in pericentric condensin clustering [79], as well as a decrease in condensin localization to rDNA sites [80] in budding yeast. This result is consistent with decrease in tension at the pericentromere [79] and lack of sister chromatid cohesion [81], suggesting a role for sumoylation in these activities of pericentric condensin.
DNA topoisomerase II (Top2) has distinct interactions with condensin and cohesin in both the pericentromere and rDNA in a variety of eukaryotic species [82,83,84]. In the pericentromere, Top2 has an essential role in regulating mitotic chromosome structure and tension [81,85]. Depletion of condensin in Drosophila disrupts Top2 centromeric localization [86], suggesting that condensin plays a role in regulating Top2 localization. A similar dependence for condensin-mediated Top2 is observed at the rDNA locus in budding yeast, with both reduced binding of Top2 in the absence of condensin as well as a lack of restoration of segregation defects with Top2 overexpression in the condensin mutants [83]. Sumoylation sites are present in Top2 [81], potentially influencing its activity. Deletion of either Top2 or isopeptidase Ulp2/Smt4 in budding yeast causes a decrease in compaction at the pericentromere during metaphase [79] as well as increased pericentromere stretching [85]. These data suggest that Top2 activity, coordinated by sumoylation, regulates pericentromere dynamics. In yeast lacking cohesin, in which biorientation of sister kinetochores is lost during mitosis, depletion of Top2 actually restores biorientation, indicating that linkage between sister chromatids is a balance between cohesion and catenation [87]. Similar findings have also been observed in DT40 chicken cells [88]. A combination of ChIP-seq and Hi-C analysis indicates that human Top2 associates with cohesin subunits and rDNA-binding proteins [84], confirming a potential role of Top2 at these locations.
In mammalian cells, a zinc finger protein called CCCTC-binding factor (CTCF) is involved in forming chromatin loops [89,90] and associates with cohesin [91,92]. CTCF can function as an insulator, preventing interaction between active and inactive chromatin and blocking enhancer activity [93]. While CTCF has not been identified in lower eukaryotic species such as Saccharomyces cerevisiae, Schizosaccharomyces pombe, and Caenorhabditis elegans [94], transformation studies suggest that CTCF has a similar insulating function in yeast [95]. Furthermore, members of the Ctf19/COMA complex, which regulates pericentromeric cohesin enrichment [51,96,97], play a vital role pericentromere loop formation [98], perhaps similar to mechanism by which CTCF mediates cohesin at the base of loops [99]. This suggests that the Ctf19/COMA complex could function as the yeast equivalent of CTCF. ChIP-seq analysis also indicates that CTCF associates with Pol III sites (tRNA genes) [100], which as discussed below, are tethered to the nucleolus and localized in the pericentromere. CTCF further regulates rDNA in human cells [101,102], and localizes to the nucleolus in mammalian cells and Drosophila [103,104]. Knockdown of condensin increases CTCF binding to rDNA, suggesting a role for condensin in negatively regulating CTCF [105]. These data therefore suggest a common role for CTCF, possibly via association with SMC proteins, in regulating both pericentric and nucleolar DNA.
2.3. tRNA Genes Are Localized to Both the Pericentromere and the Nucleolus
Transfer ribonucleic acid (tRNA) genes, also referred to as tDNA, are short sequences located throughout the genome and are bound by transcription factor RNA polymerase III (Pol III) [106]. These genes, of which there are 274 in yeast and approximately 450 in humans, are dispersed throughout the genome. However, nucleotide sequencing in fission yeast has shown that some tRNA genes are localized in the pericentromere in fission yeast [107], and FISH studies have identified pericentric tRNA genes in both fission and budding yeast [39,40,41]. Tfc1, a subunit of the Pol III transcription factor complex, was also found to be localized in the pericentric region in budding yeast [35]. Some tRNA genes are also located in the periphery of the nucleolus in fission and budding yeast [40,41,108], and early processing of tRNAs has previously been found to occur in the nucleolus in yeast [109]. It has further been shown that the tethering of tRNA genes to the nucleolus is influenced by proximity to centromeres, with closer proximity of Pol III-transcribed genes to centromeres associated with less nucleoli association [110]. This suggests an additional level of regulation of tRNA gene localization that is influenced by Rabl configuration-like organization, consistent with chromosome arms extending away from centromeres towards the nucleolus [111,112].
tRNA genes are sites of enriched cohesin [42] and condensin [5,41]. Interestingly, condensin mutations result in loss of nucleolar clustering of tRNA genes, suggesting that condensin plays a role in tRNA localization [41]. Deletion of tRNA genes on chromosome III in budding yeast disrupts not only condensin localization to tDNA sites, but also affects centromere–centromere interaction [112], suggesting effects of tRNA genes on chromosome structure. Cohesin regulates tRNA activity, with mutations that cause human cohesinopathies resulting in defects in tRNA gene-mediated silencing when expressed in budding yeast [113]. The histone deacetylase Sir2, which as previously mentioned affects axial condensin localization at the pericentromere [17], is similarly responsible for enrichment of condensin and cohesin at tRNA sites [74].
Cbf5, a small nucleolar ribonucleoprotein, further regulates tRNA distribution. Mutations in Cbf5 disrupts both the nucleolar localization of pre-tRNAs as well as alleviating tRNA gene-mediated silencing [49]. Cbf5 also regulates pericentric condensin; in budding yeast, Cbf5 mutants have decreased condensin enrichment at the pericentromere [35], suggesting that condensin regulation may underlie the effects of Cbf5 on nucleolar tRNA localization.
Certain DNA elements called chromatin barriers or insulators, which are present in multiple eukaryotic species, play a role in structurally defining functionally distinct chromatin regions [114]. In particular, tRNA genes have been shown to separate heterochromeric DNA from unsilenced regions. In fission yeast, a centromeric tRNA gene was found to play a role in defining centromeric heterochromatin and normal meiotic segregation [115], and is dependent on Pol III activity [116]. In human cells, tRNA genes have a similar function, with multimerized tDNAs increasing enhancer blocking [117]. Similarly, tRNA-mediated gene silencing is dependent on its nucleolar localization in budding yeast [40]. In addition, tRNA genes represent sites of replication fork pausing in budding and fission yeast, possibly due to their high rates of transcription [118,119]. Furthermore, in both specific tRNA genes and centromeres, there is a dependency on replisome progression complex member Tof1, but not Mrc1, for replication fork pausing [120]. The function of tRNA as a chromatin barrier in multiple eukaryotic species suggests a distinct conservation of this mechanism.
2.4. Replication Fork Stalling in Pericentromere and rDNA
Both centromeres and rDNA repeats in the nucleolus are characterized by blockades that disrupt fork progression. While highly repetitive DNA sequences are thought to promote replication fork pausing [121,122,123], even the ~125 bp non-repetitive point centromeres in yeast feature fork stalling [120,124]. A complex of S-phase checkpoint proteins (Tof1, Mrc1, Csm3) that are responsible for slowing DNA synthesis in the presence of DNA damage localize specifically to replication forks in budding yeast [125]. Tof1 in particular is required for fork pausing at yeast centromeres [120]. Deletion of Csm3, which interacts directly with Tof1 [126], is important for establishment of fork pausing [127]. DNA helicase Rrm3 promotes replication fork progression at multiple genomic sites, including tRNA genes, rDNA, and centromeres [36,128]. Csm3 may facilitate this fork pausing by blocking Rrm3 helicase-induced progression through replication forks [129], suggesting a common mechanism of stalled forks in both the nucleolus and the pericentromere.
Chl4, Iml3, and Mcm21, proteins that are members of the Ctf19 complex in yeast (analogous to the constitutive centromere associated network, or CCAN, in mammals), are involved in kinetochore assembly at the centromere [130]. Interestingly, Chl4, Iml3, and Mcm21 are all required for pericentric cohesin enrichment [96,97]. Loss of either Iml3 or Chl4 causes a decrease of pericentric cohesin, which is counteracted by slowing replication with hydroxyurea treatment [96]. The cohesin-loading function of these proteins could therefore be instrumental to ensuring that pericentric cohesin is in place prior to the replication fork, ensuring proper cohesion of the resulting sister chromatids. A yeast model using a conditional dicentric strain, which allows the study of de novo kinetochore assembly, demonstrated that Chl4/Iml3/Mcm21 mutants all suppress dicentric breakage [131]. The lack of breakage that is normally induced in a dicentric strain is consistent with a lack of de novo kinetochore assembly in these mutants. Furthermore, de novo kinetochore assembly in Chl4/Iml3 mutants is rescued by pausing replication using hydroxyurea [98]. By slowing the replication process, this may allow additional time for these mutants to resume proper kinetochore assembly.
At the rDNA locus, stalling at replication forks has been well-established in both yeast and mammals [132]. In a single 9 kb rDNA repeat in budding yeast, the 35S gene is transcribed by Pol I, followed by the 5S gene that is transcribed by Pol III in the opposing direction. A replication fork barrier (RFB) is located at the 3’ end of the 35S gene, allowing replication to occur through 35S but blocking replication in the opposing direction [133]. At the human rDNA locus, however, the replication fork barrier functions in a uniquely bi-directional manner, blocking replication from occurring in both directions at this junction [134]. In yeast, a protein called Fob1 is required for RFB activity at the rDNA locus [135,136]. Fob1 co-localizes with rDNA-binding protein Hmo1 [29] and also condensin [47], proteins that as discussed previously may be involved in DNA looping. Similarly, atomic force microscopy imaging indicates that the RFB sequence may actually wrap around Fob1 in a nucleosome-like fashion [136].
Top2, which as previously mentioned associates with SMC proteins, plays an important role regarding replication termination both in the pericentromere as well as in rDNA repeats. Termination regions (TERs) are located at the point of two converging replication forks, contain fork pausing elements [137], and are crucial for terminating replication [138]. In budding yeast, TERs are particularly concentrated near centromeres, and mutation of Top2 causes double-stranded breaks and recombination at these sites [138]. Top2 mutants result in repair checkpoint activation that is counteracted by a Top2-Fob1 double mutant, implying that Top2 mediates proper replication termination in rDNA as well [139].
DNA ligase 4 (Dnl4), a protein involved in non-homologous end-joining DNA repair [140], may play a role at the replication fork barrier in rDNA sites. Dnl4 interacts with replication fork-associated Sgs1 to prevent fork breakage-mediated events in rDNA [37]. While deletions of Dnl4 in budding yeast may influence segregation events at the pericentromere [38], it remains unclear if Dnl4 may have a similar role specifically in pericentric fork pausing.
2.5. Recombination Control in the Pericentromere and rDNA
The similarities between the pericentric region and nucleolar rDNA suggests that these two regions may also have similar features in regard to control of recombination and resulting genomic instability. Highly repetitive regions of the genome, such as those in regional centromeres of mammals and rDNA in eukaryotes, are susceptible to mitotic recombination. While recombination during meiosis tends be greatly repressed near the centromere in a variety of species [141], in mitosis, recombination occurs at budding yeast pericentromeres in the form of gene conversion [142], though this may occur at reduced levels closer to the centromere [143]. In mammals, however, which feature regional centromeres with high numbers of repeat sequences, mitotic recombination is a common occurrence. Using chromosome-oriented FISH to specifically target centromere repeats in mouse cells, it has been shown that there are extremely high mitotic recombination events at centromeres compared to the rest of the genome [144]. As mentioned previously, it has also been proposed that the highly repetitive sequences found in higher eukaryotes promotes recombination, which further drives the loop formation that is necessary for proper centromere function [21].
Replication fork stalling further facilitates recombination events [145]. Double-stranded breaks sometimes occur as a result of stalled replication forks [146,147], which are commonly repaired by either homologous recombination in yeast, or non-homologous end-joining in mammals. The rDNA-binding protein Fob1, which is essential for RFB formation, is particularly important for facilitating these recombination events [148,149]. As demonstrated in mutant yeast strains with lower copy numbers of rDNA, this activity of Fob1 is also dependent on the rate of transcription, with higher transcription correlating to more recombination even in Fob1 mutants [150]. Recombination hot-spot (HOT1), a DNA element that increases levels of inter- and intrachromosomal homologous recombination between repeats [151], is highly prevalent surrounding rDNA repeats. Analysis of HOT1 mutants indicated that only the set containing mutated Fob1 had defects in homologous recombination, suggesting that Fob1 is an important mediator at these locations [135]. Kobayashi et al. further demonstrated that Fob1 mediates the expansion/contraction of rDNA repeats [149]. Whether fork stalling might influence recombination at the pericentric region, however, has yet to be determined.
Sir2, which as previously mentioned affects localization of condensin/cohesin at the pericentromere and rDNA sites, appears to negatively influence recombination in rDNA repeats. Sir2 mutants have a reliance on recombination genes (Rad50 and Rad52) that are dispensable in wild-type strains [152], suggesting a unique recombination pathway controlled by Sir2. Deletion of Sir2 also increases the number of rDNA repeats, and decreases rDNA-associated cohesin [48], suggesting that Sir2 may mediate or regulate levels of recombination by enhancing sister chromatid cohesion. Interestingly, in the pericentromere of fission yeast, cohesin prevents double-stranded breaks and the resulting recombination events from occurring during meiosis [153]. These data suggest a potential role for Sir2 and cohesin in controlling recombination events in both the pericentromere and rDNA.
Top2, as a mediator of replication fork progression and sister chromatid decatenation, may regulate recombination events at the pericentromere and at rDNA sites. Top2 mutants have enhanced recombination events at TERs, regions that are concentrated near centromeres [138], and enhanced recombination is observed at rDNA sites in Top2 mutants [154], suggesting that Top2 may suppress recombination at these regions. Conversely, the association between Top2 and cohesin/CTCF [84] at potential loop anchor points may actually promote rearrangement events. In mammalian cells, double-stranded breaks induced by the anti-cancer drug etoposide corresponds with sites occupied by CTCF and Top2 [155,156]. Meanwhile, double-stranded breaks are decreased in Top2 mutants [155]. These data suggest that Top2 may have unique effects on genomic stability, particularly through its interactions with proteins such as cohesin.
2.6. Phase Separation in the Nucleolus and Pericentromere
Phase separation as a mechanism for defining discrete departments within the cell has been a rapidly growing field within cell biology. The nucleolus, which consists of liquid–liquid separated phases, also known as biomolecular condensates [157], has been the target of many such studies. Liquid–liquid phase separation (LLPS) has been identified in the nucleolus of Xenopus [158,159] as well as C. elegans [160]. Photobleaching experiments in budding yeast further suggest an organized network of the nucleolus, with distinct segregation of nucleolar proteins following mitosis [161]. Membraneless organelles of not only the nucleolus but also those such as stress granules and nuclear speckles are characterized by RNA-protein interactions [162], suggesting that these interactions may promote phase separation. Supporting this notion, mutation of the RNA recognition motif of nucleolar protein NPM1 prevents the formation of liquid-like droplets [159,163]. LLPS is thought to be crucial in defining the organization of nucleoli [159], facilitating the role of nucleolar sub compartmentalization in RNA processing and ribosome biogenesis [164]. In human cell lines, the liquid-like state of the nucleolus is further involved in quality control of misfolded proteins [165].
In addition to LLPS, polymer–polymer phase separation (PPPS) may also play a role in compartmentalizing not only the nucleolus but also in defining chromosome territories. Polymer models suggest that entropic forces generated by chromatin polymers constrain chromosome territories [166,167,168,169]. Furthermore, computational modeling of nucleolar structure suggests nucleolar phase separation may be driven by polymer crowding, even without assuming the presence DNA binding factors [170]. Bead-spring models of chromatin dynamics further suggest that nucleolar PPPS is formed by chromosomal cross-linking and DNA loop formation [171]. In addition, this notion that DNA loops drive PPPS in the nucleolus suggests that a similar mechanism may regulate PPPS at the pericentromere, which as previously discussed is characterized by the formation of DNA loops.
While the pericentromere is less well-studied compared to the nucleolus in regard to potential LLPS properties, there is emerging evidence that supports such phase separation. The chromosomal passenger complex (CPC), which includes the kinase Aurora B as well as other subunits such as INCENP and borealin, regulates tension between sister kinetochores, ensuring proper segregation of sister chromatids [130,172]. In vitro experiments indicate that the non-kinase subunits of the CPC form liquid-like droplets at physiological centromere concentrations, and experiments using HeLa cells further suggest that phase separation induced by CPC component borealin mediates its location to the inner centromere [173]. The authors found that alpha satellite RNA was associated with the liquid-like droplets, suggesting a potential role of CEN RNA in formation of this liquid–liquid phase, similar to nucleolar RNA/protein interactions facilitating LLPS [162]. Interestingly, Aurora B mediates pericentric enrichment of condensin in HeLa cells [174], suggesting a potential interplay for other pericentric proteins in mediating phase separation. Furthermore, interphasic centromeres can localize to the nucleolus in both Drosophila and human cells [175,176,177]. In human cells, the CPC component INCENP associates with centromeric alpha satellite RNA, targeting the interphasic nucleolar localization of centromeres [178]. Whether this activity may be linked to a potential role of CPC components in liquid–liquid phase separation has yet to be determined, however.
Repetitive heterochromatin, such as the alpha satellite repeats present in regional centromeres, has been shown to promote liquid–liquid phase separation. In particular, heterochromatin protein 1α (HP1α), a protein localized to heterochromatin in mitotic centromeres in higher eukaryotes [179], is responsible for liquid-like droplet formation in Drosophila and mammalian cells [180,181]. While HP1 has not been identified in budding yeast, the histone deacetylase Sir2 may have a similar role in regulating heterochromatin [182]. It has further been suggested that phase separation of these repetitive DNA sequences may drive chromatin organization and folding [183], suggesting a potential mechanism at centromeres and in the nucleolus.
3. Conclusions
As two seemingly discrete regions, the pericentromere and nucleolus have a number of commonalities in regard to their features and regulatory mechanisms. The similar chromatin structure of both the pericentromere and rDNA, regulated in part by SMC proteins, may impart similar features such as fork pausing, tRNA tethering, regulation of genomic instability, and phase separation. Mechanisms underlying such activities in one region may therefore be used to guide studies of similar activities in the other region.
Author Contributions
Conceptualization, K.B.; writing—original draft preparation, C.J.L.; writing—review and editing, C.J.L. and K.B.; visualization, C.J.L.; supervision, K.B.; funding acquisition, K.B.
Funding
This research was funded by the National Institutes of Health (NIH) R37-GM32238.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Verdaasdonk, J.S.; Bloom, K. Centromeres: Unique chromatin structures that drive chromosome segregation. Nat. Rev. Mol. Cell Biol. 2011, 12, 320–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleveland, D.W.; Mao, Y.; Sullivan, K.F. Centromeres and kinetochores: From epigenetics to mitotic checkpoint signaling. Cell 2003, 112, 407–421. [Google Scholar] [CrossRef] [Green Version]
- Blat, Y.; Kleckner, N. Cohesins bind to preferential sites along yeast chromosome III, with differential regulation along arms versus the centric region. Cell 1999, 98, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Megee, P.C.; Mistrot, C.; Guacci, V.; Koshland, D. The centromeric sister chromatid cohesion site directs Mcd1p binding to adjacent sequences. Mol. Cell 1999, 4, 445–450. [Google Scholar] [CrossRef]
- D’Ambrosio, C.; Schmidt, C.K.; Katou, Y.; Kelly, G.; Itoh, T.; Shirahige, K.; Uhlmann, F. Identification of cis-acting sites for condensin loading onto budding yeast chromosomes. Genes Dev. 2008, 22, 2215–2227. [Google Scholar] [CrossRef] [Green Version]
- Lawrimore, J.; Bloom, K. The regulation of chromosome segregation via centromere loops. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 352–370. [Google Scholar] [CrossRef]
- Kobayashi, T. Ribosomal RNA gene repeats, their stability and cellular senescence. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2014, 90, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Hertwig, O. Lehrbuch der Entwicklungsgeschichte des Menschen und der Wirbeltiere (Textbook of developmental History of Humans and Vertebrates); Nabu Press: Charleston, SC, USA, 1906; p. 548. [Google Scholar]
- Hofmann, A.; Heermann, D.W. The role of loops on the order of eukaryotes and prokaryotes. FEBS Lett. 2015, 589, 2958–2965. [Google Scholar] [CrossRef] [Green Version]
- Filipski, J.; Mucha, M. Structure, function and DNA composition of Saccharomyces cerevisiae chromatin loops. Gene 2002, 300, 63–68. [Google Scholar] [CrossRef]
- Yeh, E.; Haase, J.; Paliulis, L.V.; Joglekar, A.; Bond, L.; Bouck, D.; Salmon, E.D.; Bloom, K.S. Pericentric chromatin is organized into an intramolecular loop in mitosis. Curr. Biol. 2008, 18, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Stephens, A.D.; Haase, J.; Vicci, L.; Taylor, R.M., 2nd; Bloom, K. Cohesin, condensin, and the intramolecular centromere loop together generate the mitotic chromatin spring. J. Cell Biol. 2011, 193, 1167–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aze, A.; Sannino, V.; Soffientini, P.; Bachi, A.; Costanzo, V. Centromeric DNA replication reconstitution reveals DNA loops and ATR checkpoint suppression. Nat. Cell Biol. 2016, 18, 684–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, S.A.; Vagnarelli, P.; Dong, Y.; Hori, T.; McEwen, B.F.; Fukagawa, T.; Flors, C.; Earnshaw, W.C. A super-resolution map of the vertebrate kinetochore. Proc. Natl. Acad. Sci. USA 2010, 107, 10484–10489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrimore, J.; Vasquez, P.A.; Falvo, M.R.; Taylor, R.M., 2nd; Vicci, L.; Yeh, E.; Forest, M.G.; Bloom, K. DNA loops generate intracentromere tension in mitosis. J. Cell Biol. 2015, 210, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Lawrimore, J.; Aicher, J.K.; Hahn, P.; Fulp, A.; Kompa, B.; Vicci, L.; Falvo, M.; Taylor, R.M., 2nd; Bloom, K. ChromoShake: A chromosome dynamics simulator reveals that chromatin loops stiffen centromeric chromatin. Mol. Biol. Cell 2016, 27, 153–166. [Google Scholar] [CrossRef]
- Stephens, A.D.; Quammen, C.W.; Chang, B.; Haase, J.; Taylor, R.M., 2nd; Bloom, K. The spatial segregation of pericentric cohesin and condensin in the mitotic spindle. Mol. Biol. Cell 2013, 24, 3909–3919. [Google Scholar] [CrossRef]
- Lawrimore, J.; Doshi, A.; Friedman, B.; Yeh, E.; Bloom, K. Geometric partitioning of cohesin and condensin is a consequence of chromatin loops. Mol. Biol. Cell 2018, 29, 2737–2750. [Google Scholar] [CrossRef]
- Vas, A.C.; Andrews, C.A.; Kirkland Matesky, K.; Clarke, D.J. In vivo analysis of chromosome condensation in Saccharomyces cerevisiae. Mol. Biol. Cell 2007, 18, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T. Condensins: Universal organizers of chromosomes with diverse functions. Genes Dev. 2012, 26, 1659–1678. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, R.J.; Humphrey, T.C. A role for recombination in centromere function. Trends Genet. 2010, 26, 209–213. [Google Scholar] [CrossRef]
- Guacci, V.; Hogan, E.; Koshland, D. Chromosome condensation and sister chromatid pairing in budding yeast. J. Cell Biol. 1994, 125, 517–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, B.; Bose, T.; Lee, K.K.; Wang, F.; Lu, S.; Ross, R.T.; Zhang, Y.; French, S.L.; Beyer, A.L.; Slaughter, B.D.; et al. Cohesion promotes nucleolar structure and function. Mol. Biol. Cell 2014, 25, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Mayan, M.; Aragon, L. Cis-interactions between non-coding ribosomal spacers dependent on RNAP-II separate RNAP-I and RNAP-III transcription domains. Cell Cycle 2010, 9, 4328–4337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.; Acar, M. Live-Cell Imaging of Chromatin Condensation Dynamics by CRISPR. iScience 2018, 4, 216–235. [Google Scholar] [CrossRef] [PubMed]
- Lazar-Stefanita, L.; Scolari, V.F.; Mercy, G.; Muller, H.; Guerin, T.M.; Thierry, A.; Mozziconacci, J.; Koszul, R. Cohesins and condensins orchestrate the 4D dynamics of yeast chromosomes during the cell cycle. EMBO J. 2017, 36, 2684–2697. [Google Scholar] [CrossRef] [PubMed]
- Merz, K.; Hondele, M.; Goetze, H.; Gmelch, K.; Stoeckl, U.; Griesenbeck, J. Actively transcribed rRNA genes in S. cerevisiae are organized in a specialized chromatin associated with the high-mobility group protein Hmo1 and are largely devoid of histone molecules. Genes Dev. 2008, 22, 1190–1204. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, A.C.; Sullivan, G.J.; McStay, B. UBF binding in vivo is not restricted to regulatory sequences within the vertebrate ribosomal DNA repeat. Mol. Cell Biol. 2002, 22, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Gadal, O.; Labarre, S.; Boschiero, C.; Thuriaux, P. Hmo1, an HMG-box protein, belongs to the yeast ribosomal DNA transcription system. EMBO J. 2002, 21, 5498–5507. [Google Scholar] [CrossRef] [Green Version]
- Stefanovsky, V.Y.; Pelletier, G.; Bazett-Jones, D.P.; Crane-Robinson, C.; Moss, T. DNA looping in the RNA polymerase I enhancesome is the result of non-cooperative in-phase bending by two UBF molecules. Nucleic Acids Res. 2001, 29, 3241–3247. [Google Scholar] [CrossRef] [Green Version]
- Stefanovsky, V.Y.; Bazett-Jones, D.P.; Pelletier, G.; Moss, T. The DNA supercoiling architecture induced by the transcription factor xUBF requires three of its five HMG-boxes. Nucleic Acids Res. 1996, 24, 3208–3215. [Google Scholar] [CrossRef] [Green Version]
- Putnam, C.D.; Copenhaver, G.P.; Denton, M.L.; Pikaard, C.S. The RNA polymerase I transactivator upstream binding factor requires its dimerization domain and high-mobility-group (HMG) box 1 to bend, wrap, and positively supercoil enhancer DNA. Mol. Cell Biol. 1994, 14, 6476–6488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, C.; Wang, Z.; Roeder, R.G.; Gall, J.G. RNA polymerase III in Cajal bodies and lampbrush chromosomes of the Xenopus oocyte nucleus. Mol. Biol. Cell 2002, 13, 3466–3476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganji, M.; Shaltiel, I.A.; Bisht, S.; Kim, E.; Kalichava, A.; Haering, C.H.; Dekker, C. Real-time imaging of DNA loop extrusion by condensin. Science 2018, 360, 102–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snider, C.E.; Stephens, A.D.; Kirkland, J.G.; Hamdani, O.; Kamakaka, R.T.; Bloom, K. Dyskerin, tRNA genes, and condensin tether pericentric chromatin to the spindle axis in mitosis. J. Cell Biol. 2014, 207, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Azvolinsky, A.; Dunaway, S.; Torres, J.Z.; Bessler, J.B.; Zakian, V.A. The S. cerevisiae Rrm3p DNA helicase moves with the replication fork and affects replication of all yeast chromosomes. Genes Dev. 2006, 20, 3104–3116. [Google Scholar] [CrossRef] [Green Version]
- Fritsch, O.; Burkhalter, M.D.; Kais, S.; Sogo, J.M.; Schar, P. DNA ligase 4 stabilizes the ribosomal DNA array upon fork collapse at the replication fork barrier. DNA Repair (Amst) 2010, 9, 879–888. [Google Scholar] [CrossRef]
- Hanlon, S.L.; Li, J.J. Re-replication of a centromere induces chromosomal instability and aneuploidy. PLoS Genet. 2015, 11, e1005039. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, O.; Tanaka, A.; Tanizawa, H.; Grewal, S.I.; Noma, K. Centromeric localization of dispersed Pol III genes in fission yeast. Mol. Biol. Cell 2010, 21, 254–265. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.; Haeusler, R.A.; Good, P.D.; Engelke, D.R. Nucleolar clustering of dispersed tRNA genes. Science 2003, 302, 1399–1401. [Google Scholar] [CrossRef] [Green Version]
- Haeusler, R.A.; Pratt-Hyatt, M.; Good, P.D.; Gipson, T.A.; Engelke, D.R. Clustering of yeast tRNA genes is mediated by specific association of condensin with tRNA gene transcription complexes. Genes Dev. 2008, 22, 2204–2214. [Google Scholar] [CrossRef] [Green Version]
- Glynn, E.F.; Megee, P.C.; Yu, H.G.; Mistrot, C.; Unal, E.; Koshland, D.E.; DeRisi, J.L.; Gerton, J.L. Genome-wide mapping of the cohesin complex in the yeast Saccharomyces cerevisiae. PLoS Biol. 2004, 2, E259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavoie, B.D.; Tuffo, K.M.; Oh, S.; Koshland, D.; Holm, C. Mitotic chromosome condensation requires Brn1p, the yeast homologue of Barren. Mol. Biol. Cell 2000, 11, 1293–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavoie, B.D.; Hogan, E.; Koshland, D. In vivo requirements for rDNA chromosome condensation reveal two cell-cycle-regulated pathways for mitotic chromosome folding. Genes Dev. 2004, 18, 76–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavoie, B.D.; Hogan, E.; Koshland, D. In vivo dissection of the chromosome condensation machinery: Reversibility of condensation distinguishes contributions of condensin and cohesin. J. Cell Biol. 2002, 156, 805–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matos-Perdomo, E.; Machin, F. The ribosomal DNA metaphase loop of Saccharomyces cerevisiae gets condensed upon heat stress in a Cdc14-independent TORC1-dependent manner. Cell Cycle 2018, 17, 200–215. [Google Scholar] [CrossRef] [Green Version]
- Johzuka, K.; Terasawa, M.; Ogawa, H.; Ogawa, T.; Horiuchi, T. Condensin loaded onto the replication fork barrier site in the rRNA gene repeats during S phase in a FOB1-dependent fashion to prevent contraction of a long repetitive array in Saccharomyces cerevisiae. Mol. Cell Biol. 2006, 26, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Horiuchi, T.; Tongaonkar, P.; Vu, L.; Nomura, M. SIR2 regulates recombination between different rDNA repeats, but not recombination within individual rRNA genes in yeast. Cell 2004, 117, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Kendall, A.; Hull, M.W.; Bertrand, E.; Good, P.D.; Singer, R.H.; Engelke, D.R. A CBF5 mutation that disrupts nucleolar localization of early tRNA biosynthesis in yeast also suppresses tRNA gene-mediated transcriptional silencing. Proc. Natl. Acad. Sci. USA 2000, 97, 13108–13113. [Google Scholar] [CrossRef] [Green Version]
- Fukagawa, T.; Nogami, M.; Yoshikawa, M.; Ikeno, M.; Okazaki, T.; Takami, Y.; Nakayama, T.; Oshimura, M. Dicer is essential for formation of the heterochromatin structure in vertebrate cells. Nat. Cell Biol. 2004, 6, 784–791. [Google Scholar] [CrossRef]
- Eckert, C.A.; Gravdahl, D.J.; Megee, P.C. The enhancement of pericentromeric cohesin association by conserved kinetochore components promotes high-fidelity chromosome segregation and is sensitive to microtubule-based tension. Genes Dev. 2007, 21, 278–291. [Google Scholar] [CrossRef] [Green Version]
- Freeman, L.; Aragon-Alcaide, L.; Strunnikov, A. The condensin complex governs chromosome condensation and mitotic transmission of rDNA. J. Cell Biol. 2000, 149, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, N.; Nakamura, T.; Kokubu, A.; Ebe, M.; Nagao, K.; Yanagida, M. Dissection of the essential steps for condensin accumulation at kinetochores and rDNAs during fission yeast mitosis. J. Cell Biol. 2008, 180, 1115–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, T. At the heart of the chromosome: SMC proteins in action. Nat. Rev. Mol. Cell Biol. 2006, 7, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Strunnikov, A.V.; Hogan, E.; Koshland, D. SMC2, a Saccharomyces cerevisiae gene essential for chromosome segregation and condensation, defines a subgroup within the SMC family. Genes Dev. 1995, 9, 587–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terakawa, T.; Bisht, S.; Eeftens, J.M.; Dekker, C.; Haering, C.H.; Greene, E.C. The condensin complex is a mechanochemical motor that translocates along DNA. Science 2017, 358, 672–676. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Rybenkov, V.V.; Crisona, N.J.; Hirano, T.; Cozzarelli, N.R. 13S condensin actively reconfigures DNA by introducing global positive writhe: Implications for chromosome condensation. Cell 1999, 98, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Baxter, J.; Sen, N.; Martinez, V.L.; De Carandini, M.E.; Schvartzman, J.B.; Diffley, J.F.; Aragon, L. Positive supercoiling of mitotic DNA drives decatenation by topoisomerase II in eukaryotes. Science 2011, 331, 1328–1332. [Google Scholar] [CrossRef]
- Strick, T.R.; Bensimon, D.; Croquette, V. Micro-mechanical measurement of the torsional modulus of DNA. Genetica 1999, 106, 57–62. [Google Scholar] [CrossRef]
- Haering, C.H.; Lowe, J.; Hochwagen, A.; Nasmyth, K. Molecular architecture of SMC proteins and the yeast cohesin complex. Mol. Cell 2002, 9, 773–788. [Google Scholar] [CrossRef]
- Gruber, S.; Haering, C.H.; Nasmyth, K. Chromosomal cohesin forms a ring. Cell 2003, 112, 765–777. [Google Scholar] [CrossRef] [Green Version]
- Skibbens, R.V. Condensins and cohesins—One of these things is not like the other! J. Cell Sci. 2019, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmann, F.; Lee, B.G.; Than, T.; Sinn, L.; O’Reilly, F.J.; Yatskevich, S.; Rappsilber, J.; Hu, B.; Nasmyth, K.; Lowe, J. A folded conformation of MukBEF and cohesin. Nat. Struct. Mol. Biol. 2019, 26, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stigler, J.; Camdere, G.O.; Koshland, D.E.; Greene, E.C. Single-Molecule Imaging Reveals a Collapsed Conformational State for DNA-Bound Cohesin. Cell Rep. 2016, 15, 988–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaelis, C.; Ciosk, R.; Nasmyth, K. Cohesins: Chromosomal proteins that prevent premature separation of sister chromatids. Cell 1997, 91, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Nasmyth, K.; Haering, C.H. Cohesin: Its roles and mechanisms. Annu. Rev. Genet. 2009, 43, 525–558. [Google Scholar] [CrossRef] [Green Version]
- Heidinger-Pauli, J.M.; Mert, O.; Davenport, C.; Guacci, V.; Koshland, D. Systematic reduction of cohesin differentially affects chromosome segregation, condensation, and DNA repair. Curr. Biol. 2010, 20, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Davidson, I.F.; Bauer, B.; Goetz, D.; Tang, W.; Wutz, G.; Peters, J.M. DNA loop extrusion by human cohesin. Science 2019. [Google Scholar] [CrossRef]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef] [Green Version]
- Gassler, J.; Brandão, H.B.; Imakaev, M.; Flyamer, I.M.; Ladstätter, S.; Bickmore, W.A.; Peters, J.M.; Mirny, L.A.; Tachibana, K. A mechanism of cohesin-dependent loop extrusion organizes zygotic genome architecture. EMBO J. 2017, 36, 3600–3618. [Google Scholar] [CrossRef]
- Rao, S.S.P.; Huang, S.C.; Glenn St Hilaire, B.; Engreitz, J.M.; Perez, E.M.; Kieffer-Kwon, K.R.; Sanborn, A.L.; Johnstone, S.E.; Bascom, G.D.; Bochkov, I.D.; et al. Cohesin Loss Eliminates All Loop Domains. Cell 2017, 171, 305–320.e24. [Google Scholar] [CrossRef] [Green Version]
- Nuebler, J.; Fudenberg, G.; Imakaev, M.; Abdennur, N.; Mirny, L.A. Chromatin organization by an interplay of loop extrusion and compartmental segregation. Proc. Natl. Acad. Sci. USA 2018, 115, E6697–E6706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wutz, G.; Varnai, C.; Nagasaka, K.; Cisneros, D.A.; Stocsits, R.R.; Tang, W.; Schoenfelder, S.; Jessberger, G.; Muhar, M.; Hossain, M.J.; et al. Topologically associating domains and chromatin loops depend on cohesin and are regulated by CTCF, WAPL, and PDS5 proteins. EMBO J. 2017, 36, 3573–3599. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Valsakumar, V.; Poorey, K.; Bekiranov, S.; Smith, J.S. Genome-wide analysis of functional sirtuin chromatin targets in yeast. Genome Biol. 2013, 14, R48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guacci, V.; Koshland, D.; Strunnikov, A. A direct link between sister chromatid cohesion and chromosome condensation revealed through the analysis of MCD1 in S. cerevisiae. Cell 1997, 91, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Dauban, L.; Kamgoue, A.; Wang, R.; Leger-Silvestre, I.; Beckouet, F.; Cantaloube, S.; Gadal, O. Quantification of the dynamic behaviour of ribosomal DNA genes and nucleolus during yeast Saccharomyces cerevisiae cell cycle. J. Struct. Biol. 2019, 208, 152–164. [Google Scholar] [CrossRef]
- Bose, T.; Lee, K.K.; Lu, S.; Xu, B.; Harris, B.; Slaughter, B.; Unruh, J.; Garrett, A.; McDowell, W.; Box, A.; et al. Cohesin proteins promote ribosomal RNA production and protein translation in yeast and human cells. PLoS Genet. 2012, 8, e1002749. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Yong-Gonzalez, V.; Kikuchi, Y.; Strunnikov, A. SIZ1/SIZ2 control of chromosome transmission fidelity is mediated by the sumoylation of topoisomerase II. Genetics 2006, 172, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Stephens, A.D.; Snider, C.E.; Bloom, K. The SUMO deconjugating peptidase Smt4 contributes to the mechanism required for transition from sister chromatid arm cohesion to sister chromatid pericentromere separation. Cell Cycle 2015, 14, 2206–2218. [Google Scholar] [CrossRef] [Green Version]
- Strunnikov, A.V.; Aravind, L.; Koonin, E.V. Saccharomyces cerevisiae SMT4 encodes an evolutionarily conserved protease with a role in chromosome condensation regulation. Genetics 2001, 158, 95–107. [Google Scholar]
- Bachant, J.; Alcasabas, A.; Blat, Y.; Kleckner, N.; Elledge, S.J. The SUMO-1 isopeptidase Smt4 is linked to centromeric cohesion through SUMO-1 modification of DNA topoisomerase II. Mol. Cell 2002, 9, 1169–1182. [Google Scholar] [CrossRef]
- Porter, A.C.; Farr, C.J. Topoisomerase II: Untangling its contribution at the centromere. Chromosome Res. Int. J. Mol. Supramol. Evol. Asp. Chromosome Biol. 2004, 12, 569–583. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, C.; Kelly, G.; Shirahige, K.; Uhlmann, F. Condensin-dependent rDNA decatenation introduces a temporal pattern to chromosome segregation. Curr. Biol. 2008, 18, 1084–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uuskula-Reimand, L.; Hou, H.; Samavarchi-Tehrani, P.; Rudan, M.V.; Liang, M.; Medina-Rivera, A.; Mohammed, H.; Schmidt, D.; Schwalie, P.; Young, E.J.; et al. Topoisomerase II beta interacts with cohesin and CTCF at topological domain borders. Genome Biol. 2016, 17, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warsi, T.H.; Navarro, M.S.; Bachant, J. DNA topoisomerase II is a determinant of the tensile properties of yeast centromeric chromatin and the tension checkpoint. Mol. Biol. Cell 2008, 19, 4421–4433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, P.A.; Queiroz-Machado, J.; Sunkel, C.E. Condensin-dependent localisation of topoisomerase II to an axial chromosomal structure is required for sister chromatid resolution during mitosis. J. Cell Sci. 2003, 116, 4763–4776. [Google Scholar] [CrossRef] [Green Version]
- Dewar, H.; Tanaka, K.; Nasmyth, K.; Tanaka, T.U. Tension between two kinetochores suffices for their bi-orientation on the mitotic spindle. Nature 2004, 428, 93–97. [Google Scholar] [CrossRef]
- Vagnarelli, P.; Morrison, C.; Dodson, H.; Sonoda, E.; Takeda, S.; Earnshaw, W.C. Analysis of Scc1-deficient cells defines a key metaphase role of vertebrate cohesin in linking sister kinetochores. EMBO Rep. 2004, 5, 167–171. [Google Scholar] [CrossRef] [Green Version]
- De Wit, E.; Vos, E.S.; Holwerda, S.J.; Valdes-Quezada, C.; Verstegen, M.J.; Teunissen, H.; Splinter, E.; Wijchers, P.J.; Krijger, P.H.; de Laat, W. CTCF Binding Polarity Determines Chromatin Looping. Mol. Cell 2015, 60, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Splinter, E.; Heath, H.; Kooren, J.; Palstra, R.J.; Klous, P.; Grosveld, F.; Galjart, N.; de Laat, W. CTCF mediates long-range chromatin looping and local histone modification in the beta-globin locus. Genes Dev. 2006, 20, 2349–2354. [Google Scholar] [CrossRef] [Green Version]
- Parelho, V.; Hadjur, S.; Spivakov, M.; Leleu, M.; Sauer, S.; Gregson, H.C.; Jarmuz, A.; Canzonetta, C.; Webster, Z.; Nesterova, T.; et al. Cohesins functionally associate with CTCF on mammalian chromosome arms. Cell 2008, 132, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Wendt, K.S.; Yoshida, K.; Itoh, T.; Bando, M.; Koch, B.; Schirghuber, E.; Tsutsumi, S.; Nagae, G.; Ishihara, K.; Mishiro, T.; et al. Cohesin mediates transcriptional insulation by CCCTC-binding factor. Nature 2008, 451, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Herold, M.; Bartkuhn, M.; Renkawitz, R. CTCF: Insights into insulator function during development. Development 2012, 139, 1045–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heger, P.; Marin, B.; Schierenberg, E. Loss of the insulator protein CTCF during nematode evolution. BMC Mol. Biol. 2009, 10, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Defossez, P.A.; Gilson, E. The vertebrate protein CTCF functions as an insulator in Saccharomyces cerevisiae. Nucleic Acids Res. 2002, 30, 5136–5141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernius, J.; Marston, A.L. Establishment of cohesion at the pericentromere by the Ctf19 kinetochore subcomplex and the replication fork-associated factor, Csm3. PLoS Genet. 2009, 5, e1000629. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.M.; Waples, W.G.; Lavoie, B.D.; Biggins, S. Pericentromeric sister chromatid cohesion promotes kinetochore biorientation. Mol. Biol. Cell 2009, 20, 3818–3827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, D.M.; Bennett, M.; Friedman, B.; Lawrimore, J.; Yeh, E.; Bloom, K. Fork pausing allows centromere DNA loop formation and kinetochore assembly. Proc. Natl. Acad. Sci. USA 2018, 115, 11784–11789. [Google Scholar] [CrossRef] [Green Version]
- Fudenberg, G.; Imakaev, M.; Lu, C.; Goloborodko, A.; Abdennur, N.; Mirny, L.A. Formation of Chromosomal Domains by Loop Extrusion. Cell Rep. 2016, 15, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Oler, A.J.; Alla, R.K.; Roberts, D.N.; Wong, A.; Hollenhorst, P.C.; Chandler, K.J.; Cassiday, P.A.; Nelson, C.A.; Hagedorn, C.H.; Graves, B.J.; et al. Human RNA polymerase III transcriptomes and relationships to Pol II promoter chromatin and enhancer-binding factors. Nat. Struct. Mol. Biol. 2010, 17, 620–628. [Google Scholar] [CrossRef]
- Zentner, G.E.; Saiakhova, A.; Manaenkov, P.; Adams, M.D.; Scacheri, P.C. Integrative genomic analysis of human ribosomal DNA. Nucleic Acids Res. 2011, 39, 4949–4960. [Google Scholar] [CrossRef] [Green Version]
- Van de Nobelen, S.; Rosa-Garrido, M.; Leers, J.; Heath, H.; Soochit, W.; Joosen, L.; Jonkers, I.; Demmers, J.; van der Reijden, M.; Torrano, V.; et al. CTCF regulates the local epigenetic state of ribosomal DNA repeats. Epigenetics Chromatin 2010, 3, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrano, V.; Navascues, J.; Docquier, F.; Zhang, R.; Burke, L.J.; Chernukhin, I.; Farrar, D.; Leon, J.; Berciano, M.T.; Renkawitz, R.; et al. Targeting of CTCF to the nucleolus inhibits nucleolar transcription through a poly(ADP-ribosyl)ation-dependent mechanism. J. Cell Sci. 2006, 119, 1746–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero, P.A.; Maggert, K.A. The CCCTC-binding factor (CTCF) of Drosophila contributes to the regulation of the ribosomal DNA and nucleolar stability. PLoS ONE 2011, 6, e16401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Jia, J.; Wu, C.; Yao, M.; Li, M.; Jin, J.; Jiang, C.; Cai, Y.; Pei, D.; Pan, G.; et al. Ribosomal RNA gene transcription mediated by the master genome regulator protein CCCTC-binding factor (CTCF) is negatively regulated by the condensin complex. J. Biol. Chem. 2013, 288, 26067–26077. [Google Scholar] [CrossRef] [Green Version]
- Dieci, G.; Fiorino, G.; Castelnuovo, M.; Teichmann, M.; Pagano, A. The expanding RNA polymerase III transcriptome. Trends Genet 2007, 23, 614–622. [Google Scholar] [CrossRef]
- Kuhn, R.M.; Clarke, L.; Carbon, J. Clustered tRNA genes in Schizosaccharomyces pombe centromeric DNA sequence repeats. Proc. Natl. Acad. Sci. USA 1991, 88, 1306–1310. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Haeusler, R.A.; Good, P.D.; Thompson, M.; Nagar, S.; Engelke, D.R. Silencing near tRNA genes requires nucleolar localization. J. Biol. Chem. 2005, 280, 8637–8639. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, E.; Houser-Scott, F.; Kendall, A.; Singer, R.H.; Engelke, D.R. Nucleolar localization of early tRNA processing. Genes Dev. 1998, 12, 2463–2468. [Google Scholar] [CrossRef] [Green Version]
- Belagal, P.; Normand, C.; Shukla, A.; Wang, R.; Leger-Silvestre, I.; Dez, C.; Bhargava, P.; Gadal, O. Decoding the principles underlying the frequency of association with nucleoli for RNA polymerase III-transcribed genes in budding yeast. Mol. Biol. Cell 2016, 27, 3164–3177. [Google Scholar] [CrossRef] [Green Version]
- Jin, Q.W.; Fuchs, J.; Loidl, J. Centromere clustering is a major determinant of yeast interphase nuclear organization. J. Cell Sci. 2000, 113, 1903–1912. [Google Scholar]
- Hamdani, O.; Dhillon, N.; Hsieh, T.S.; Fujita, T.; Ocampo, J.; Kirkland, J.G.; Lawrimore, J.; Kobayashi, T.J.; Friedman, B.; Fulton, D.; et al. tRNA Genes Affect Chromosome Structure and Function via Local Effects. Mol. Cell Biol. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gard, S.; Light, W.; Xiong, B.; Bose, T.; McNairn, A.J.; Harris, B.; Fleharty, B.; Seidel, C.; Brickner, J.H.; Gerton, J.L. Cohesinopathy mutations disrupt the subnuclear organization of chromatin. J. Cell Biol. 2009, 187, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.L.; Elgin, S.C. Putting boundaries on silence. Cell 1999, 99, 459–462. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.C.; Merrett, S.L.; Willard, H.F. A heterochromatin barrier partitions the fission yeast centromere into discrete chromatin domains. Curr. Biol. 2006, 16, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, K.C.; White, C.V.; Willard, H.F. An RNA polymerase III-dependent heterochromatin barrier at fission yeast centromere 1. PLoS ONE 2007, 2, e1099. [Google Scholar] [CrossRef]
- Raab, J.R.; Chiu, J.; Zhu, J.; Katzman, S.; Kurukuti, S.; Wade, P.A.; Haussler, D.; Kamakaka, R.T. Human tRNA genes function as chromatin insulators. EMBO J. 2012, 31, 330–350. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, A.M.; Newlon, C.S. DNA replication fork pause sites dependent on transcription. Science 1996, 272, 1030–1033. [Google Scholar] [CrossRef]
- Pryce, D.W.; Ramayah, S.; Jaendling, A.; McFarlane, R.J. Recombination at DNA replication fork barriers is not universal and is differentially regulated by Swi1. Proc. Natl. Acad. Sci. USA 2009, 106, 4770–4775. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, B.; Calzada, A.; Labib, K. Mrc1 and Tof1 regulate DNA replication forks in different ways during normal S phase. Mol. Biol. Cell 2007, 18, 3894–3902. [Google Scholar] [CrossRef] [Green Version]
- Voineagu, I.; Narayanan, V.; Lobachev, K.S.; Mirkin, S.M. Replication stalling at unstable inverted repeats: Interplay between DNA hairpins and fork stabilizing proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 9936–9941. [Google Scholar] [CrossRef] [Green Version]
- Voineagu, I.; Surka, C.F.; Shishkin, A.A.; Krasilnikova, M.M.; Mirkin, S.M. Replisome stalling and stabilization at CGG repeats, which are responsible for chromosomal fragility. Nat. Struct. Mol. Biol. 2009, 16, 226–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, K.; Lambert, S.; Baldacci, G.; Murray, J.M.; Carr, A.M. Nearby inverted repeats fuse to generate acentric and dicentric palindromic chromosomes by a replication template exchange mechanism. Genes Dev. 2009, 23, 2876–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenfeder, S.A.; Newlon, C.S. Replication forks pause at yeast centromeres. Mol. Cell Biol. 1992, 12, 4056–4066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katou, Y.; Kanoh, Y.; Bando, M.; Noguchi, H.; Tanaka, H.; Ashikari, T.; Sugimoto, K.; Shirahige, K. S-phase checkpoint proteins Tof1 and Mrc1 form a stable replication-pausing complex. Nature 2003, 424, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.L.; Pot, I.; Chang, M.; Xu, H.; Aneliunas, V.; Kwok, T.; Newitt, R.; Aebersold, R.; Boone, C.; Brown, G.W.; et al. Identification of protein complexes required for efficient sister chromatid cohesion. Mol. Biol. Cell 2004, 15, 1736–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calzada, A.; Hodgson, B.; Kanemaki, M.; Bueno, A.; Labib, K. Molecular anatomy and regulation of a stable replisome at a paused eukaryotic DNA replication fork. Genes Dev. 2005, 19, 1905–1919. [Google Scholar] [CrossRef] [Green Version]
- Ivessa, A.S.; Zhou, J.Q.; Schulz, V.P.; Monson, E.K.; Zakian, V.A. Saccharomyces Rrm3p, a 5’ to 3’ DNA helicase that promotes replication fork progression through telomeric and subtelomeric DNA. Genes Dev. 2002, 16, 1383–1396. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, B.K.; Bairwa, N.K.; Bastia, D. The Tof1p-Csm3p protein complex counteracts the Rrm3p helicase to control replication termination of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2006, 103, 897–902. [Google Scholar] [CrossRef] [Green Version]
- Biggins, S. The composition, functions, and regulation of the budding yeast kinetochore. Genetics 2013, 194, 817–846. [Google Scholar] [CrossRef] [Green Version]
- Mythreye, K.; Bloom, K.S. Differential kinetochore protein requirements for establishment versus propagation of centromere activity in Saccharomyces cerevisiae. J. Cell Biol. 2003, 160, 833–843. [Google Scholar] [CrossRef] [Green Version]
- Mirkin, E.V.; Mirkin, S.M. Replication fork stalling at natural impediments. Microbiol. Mol. Biol. Rev. 2007, 71, 13–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, B.J.; Fangman, W.L. A replication fork barrier at the 3’ end of yeast ribosomal RNA genes. Cell 1988, 55, 637–643. [Google Scholar] [CrossRef]
- Little, R.D.; Platt, T.H.; Schildkraut, C.L. Initiation and termination of DNA replication in human rRNA genes. Mol. Cell Biol. 1993, 13, 6600–6613. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Horiuchi, T. A yeast gene product, Fob1 protein, required for both replication fork blocking and recombinational hotspot activities. Genes Cells 1996, 1, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T. The replication fork barrier site forms a unique structure with Fob1p and inhibits the replication fork. Mol. Cell Biol. 2003, 23, 9178–9188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenfeder, S.A.; Newlon, C.S. A replication map of a 61-kb circular derivative of Saccharomyces cerevisiae chromosome III. Mol. Biol. Cell 1992, 3, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Fachinetti, D.; Bermejo, R.; Cocito, A.; Minardi, S.; Katou, Y.; Kanoh, Y.; Shirahige, K.; Azvolinsky, A.; Zakian, V.A.; Foiani, M. Replication termination at eukaryotic chromosomes is mediated by Top2 and occurs at genomic loci containing pausing elements. Mol. Cell 2010, 39, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Mundbjerg, K.; Jorgensen, S.W.; Fredsoe, J.; Nielsen, I.; Pedersen, J.M.; Bentsen, I.B.; Lisby, M.; Bjergbaek, L.; Andersen, A.H. Top2 and Sgs1-Top3 Act Redundantly to Ensure rDNA Replication Termination. PLoS Genet. 2015, 11, e1005697. [Google Scholar] [CrossRef] [Green Version]
- Wilson, T.E.; Grawunder, U.; Lieber, M.R. Yeast DNA ligase IV mediates non-homologous DNA end joining. Nature 1997, 388, 495–498. [Google Scholar] [CrossRef]
- Termolino, P.; Cremona, G.; Consiglio, M.F.; Conicella, C. Insights into epigenetic landscape of recombination-free regions. Chromosoma 2016, 125, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Liebman, S.W.; Symington, L.S.; Petes, T.D. Mitotic recombination within the centromere of a yeast chromosome. Science 1988, 241, 1074–1077. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.S.; Greenwell, P.W.; Dominska, M.; Gawel, M.; Hamilton, M.; Petes, T.D. A fine-structure map of spontaneous mitotic crossovers in the yeast Saccharomyces cerevisiae. PLoS Genet. 2009, 5, e1000410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaco, I.; Canela, A.; Vera, E.; Blasco, M.A. Centromere mitotic recombination in mammalian cells. J. Cell Biol. 2008, 181, 885–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothstein, R.; Michel, B.; Gangloff, S. Replication fork pausing and recombination or “gimme a break”. Genes Dev. 2000, 14, 1–10. [Google Scholar]
- Mehta, A.; Haber, J.E. Sources of DNA double-strand breaks and models of recombinational DNA repair. Cold Spring Harb. Perspect. Biol. 2014, 6, a016428. [Google Scholar] [CrossRef] [Green Version]
- Syeda, A.H.; Hawkins, M.; McGlynn, P. Recombination and replication. Cold Spring Harb. Perspect. Biol. 2014, 6, a016550. [Google Scholar] [CrossRef] [Green Version]
- Johzuka, K.; Horiuchi, T. Replication fork block protein, Fob1, acts as an rDNA region specific recombinator in S. cerevisiae. Genes Cells 2002, 7, 99–113. [Google Scholar] [CrossRef]
- Kobayashi, T.; Heck, D.J.; Nomura, M.; Horiuchi, T. Expansion and contraction of ribosomal DNA repeats in Saccharomyces cerevisiae: Requirement of replication fork blocking (Fob1) protein and the role of RNA polymerase I. Genes Dev. 1998, 12, 3821–3830. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, Y.; Horiuchi, T.; Kobayashi, T. Transcription-dependent recombination and the role of fork collision in yeast rDNA. Genes Dev. 2003, 17, 1497–1506. [Google Scholar] [CrossRef] [Green Version]
- Keil, R.L.; Roeder, G.S. Cis-acting, recombination-stimulating activity in a fragment of the ribosomal DNA of S. cerevisiae. Cell 1984, 39, 377–386. [Google Scholar] [CrossRef]
- Gottlieb, S.; Esposito, R.E. A new role for a yeast transcriptional silencer gene, SIR2, in regulation of recombination in ribosomal DNA. Cell 1989, 56, 771–776. [Google Scholar] [CrossRef]
- Nambiar, M.; Smith, G.R. Pericentromere-Specific Cohesin Complex Prevents Meiotic Pericentric DNA Double-Strand Breaks and Lethal Crossovers. Mol. Cell 2018, 71, 540–553.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christman, M.F.; Dietrich, F.S.; Fink, G.R. Mitotic recombination in the rDNA of S. cerevisiae is suppressed by the combined action of DNA topoisomerases I and II. Cell 1988, 55, 413–425. [Google Scholar] [CrossRef]
- Canela, A.; Maman, Y.; Jung, S.; Wong, N.; Callen, E.; Day, A.; Kieffer-Kwon, K.R.; Pekowska, A.; Zhang, H.; Rao, S.S.P.; et al. Genome Organization Drives Chromosome Fragility. Cell 2017, 170, 507–521.e18. [Google Scholar] [CrossRef] [Green Version]
- Gothe, H.J.; Bouwman, B.A.M.; Gusmao, E.G.; Piccinno, R.; Petrosino, G.; Sayols, S.; Drechsel, O.; Minneker, V.; Josipovic, N.; Mizi, A.; et al. Spatial Chromosome Folding and Active Transcription Drive DNA Fragility and Formation of Oncogenic MLL Translocations. Mol. Cell 2019, 75, 267–283.e12. [Google Scholar] [CrossRef]
- Correll, C.C.; Bartek, J.; Dundr, M. The Nucleolus: A Multiphase Condensate Balancing Ribosome Synthesis and Translational Capacity in Health, Aging and Ribosomopathies. Cells 2019, 8, 869. [Google Scholar] [CrossRef] [Green Version]
- Brangwynne, C.P.; Mitchison, T.J.; Hyman, A.A. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 4334–4339. [Google Scholar] [CrossRef] [Green Version]
- Feric, M.; Vaidya, N.; Harmon, T.S.; Mitrea, D.M.; Zhu, L.; Richardson, T.M.; Kriwacki, R.W.; Pappu, R.V.; Brangwynne, C.P. Coexisting Liquid Phases Underlie Nucleolar Subcompartments. Cell 2016, 165, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Weber, S.C.; Brangwynne, C.P. Inverse size scaling of the nucleolus by a concentration-dependent phase transition. Curr. Biol. 2015, 25, 641–646. [Google Scholar] [CrossRef] [Green Version]
- Girke, P.; Seufert, W. Compositional reorganization of the nucleolus in budding yeast mitosis. Mol. Biol. Cell 2019, 30, 591–606. [Google Scholar] [CrossRef]
- Sawyer, I.A.; Sturgill, D.; Dundr, M. Membraneless nuclear organelles and the search for phases within phases. Wiley Interdiscip. Rev. RNA 2019, 10, e1514. [Google Scholar] [CrossRef] [PubMed]
- Mitrea, D.M.; Cika, J.A.; Guy, C.S.; Ban, D.; Banerjee, P.R.; Stanley, C.B.; Nourse, A.; Deniz, A.A.; Kriwacki, R.W. Nucleophosmin integrates within the nucleolus via multi-modal interactions with proteins displaying R-rich linear motifs and rRNA. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, F.M.; van Koningsbruggen, S.; Navascues, J.; Lamond, A.I. The multifunctional nucleolus. Nat. Rev. Mol. Cell Biol. 2007, 8, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Frottin, F.; Schueder, F.; Tiwary, S.; Gupta, R.; Korner, R.; Schlichthaerle, T.; Cox, J.; Jungmann, R.; Hartl, F.U.; Hipp, M.S. The nucleolus functions as a phase-separated protein quality control compartment. Science 2019, 365, 342–347. [Google Scholar] [CrossRef]
- Verdaasdonk, J.S.; Vasquez, P.A.; Barry, R.M.; Barry, T.; Goodwin, S.; Forest, M.G.; Bloom, K. Centromere tethering confines chromosome domains. Mol. Cell 2013, 52, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Tjong, H.; Gong, K.; Chen, L.; Alber, F. Physical tethering and volume exclusion determine higher-order genome organization in budding yeast. Genome Res. 2012, 22, 1295–1305. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Mozziconacci, J.; Bancaud, A.; Gadal, O. Principles of chromatin organization in yeast: Relevance of polymer models to describe nuclear organization and dynamics. Curr. Opin. Cell Biol. 2015, 34, 54–60. [Google Scholar] [CrossRef]
- Vasquez, P.A.; Hult, C.; Adalsteinsson, D.; Lawrimore, J.; Forest, M.G.; Bloom, K. Entropy gives rise to topologically associating domains. Nucleic Acids Res. 2016, 44, 5540–5549. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.; Marie-Nelly, H.; Herbert, S.; Carrivain, P.; Blanc, H.; Koszul, R.; Fabre, E.; Zimmer, C. A predictive computational model of the dynamic 3D interphase yeast nucleus. Curr. Biol. 2012, 22, 1881–1890. [Google Scholar] [CrossRef] [Green Version]
- Hult, C.; Adalsteinsson, D.; Vasquez, P.A.; Lawrimore, J.; Bennett, M.; York, A.; Cook, D.; Yeh, E.; Forest, M.G.; Bloom, K. Enrichment of dynamic chromosomal crosslinks drive phase separation of the nucleolus. Nucleic Acids Res. 2017, 45, 11159–11173. [Google Scholar] [CrossRef] [Green Version]
- Lampson, M.A.; Cheeseman, I.M. Sensing centromere tension: Aurora B and the regulation of kinetochore function. Trends Cell Biol. 2011, 21, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, P.; Palomba, F.; Niedzialkowska, E.; Digman, M.A.; Gratton, E.; Stukenberg, P.T. The inner centromere is a biomolecular condensate scaffolded by the chromosomal passenger complex. Nat. Cell Biol. 2019, 21, 1127–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, T.; Fang, Y.; Spector, D.L.; Hirano, T. Spatial and temporal regulation of Condensins I and II in mitotic chromosome assembly in human cells. Mol. Biol. Cell 2004, 15, 3296–3308. [Google Scholar] [CrossRef] [PubMed]
- Ochs, R.L.; Press, R.I. Centromere autoantigens are associated with the nucleolus. Exp. Cell Res. 1992, 200, 339–350. [Google Scholar] [CrossRef]
- Carvalho, C.; Pereira, H.M.; Ferreira, J.; Pina, C.; Mendonca, D.; Rosa, A.C.; Carmo-Fonseca, M. Chromosomal G-dark bands determine the spatial organization of centromeric heterochromatin in the nucleus. Mol. Biol. Cell 2001, 12, 3563–3572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padeken, J.; Heun, P. Centromeres in nuclear architecture. Cell Cycle 2013, 12, 3455–3456. [Google Scholar] [CrossRef] [Green Version]
- Wong, L.H.; Brettingham-Moore, K.H.; Chan, L.; Quach, J.M.; Anderson, M.A.; Northrop, E.L.; Hannan, R.; Saffery, R.; Shaw, M.L.; Williams, E.; et al. Centromere RNA is a key component for the assembly of nucleoproteins at the nucleolus and centromere. Genome Res. 2007, 17, 1146–1160. [Google Scholar] [CrossRef] [Green Version]
- Yi, Q.; Chen, Q.; Liang, C.; Yan, H.; Zhang, Z.; Xiang, X.; Zhang, M.; Qi, F.; Zhou, L.; Wang, F. HP1 links centromeric heterochromatin to centromere cohesion in mammals. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef]
- Strom, A.R.; Emelyanov, A.V.; Mir, M.; Fyodorov, D.V.; Darzacq, X.; Karpen, G.H. Phase separation drives heterochromatin domain formation. Nature 2017, 547, 241. [Google Scholar] [CrossRef]
- Larson, A.G.; Elnatan, D.; Keenen, M.M.; Trnka, M.J.; Johnston, J.B.; Burlingame, A.L.; Agard, D.A.; Redding, S.; Narlikar, G.J. Liquid droplet formation by HP1alpha suggests a role for phase separation in heterochromatin. Nature 2017, 547, 236. [Google Scholar] [CrossRef] [Green Version]
- Hickman, M.A.; Froyd, C.A.; Rusche, L.N. Reinventing heterochromatin in budding yeasts: Sir2 and the origin recognition complex take center stage. Eukaryot. Cell 2011, 10, 1183–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.J. Potential Role of Phase Separation of Repetitive DNA in Chromosomal Organization. Genes (Basel) 2017, 8, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
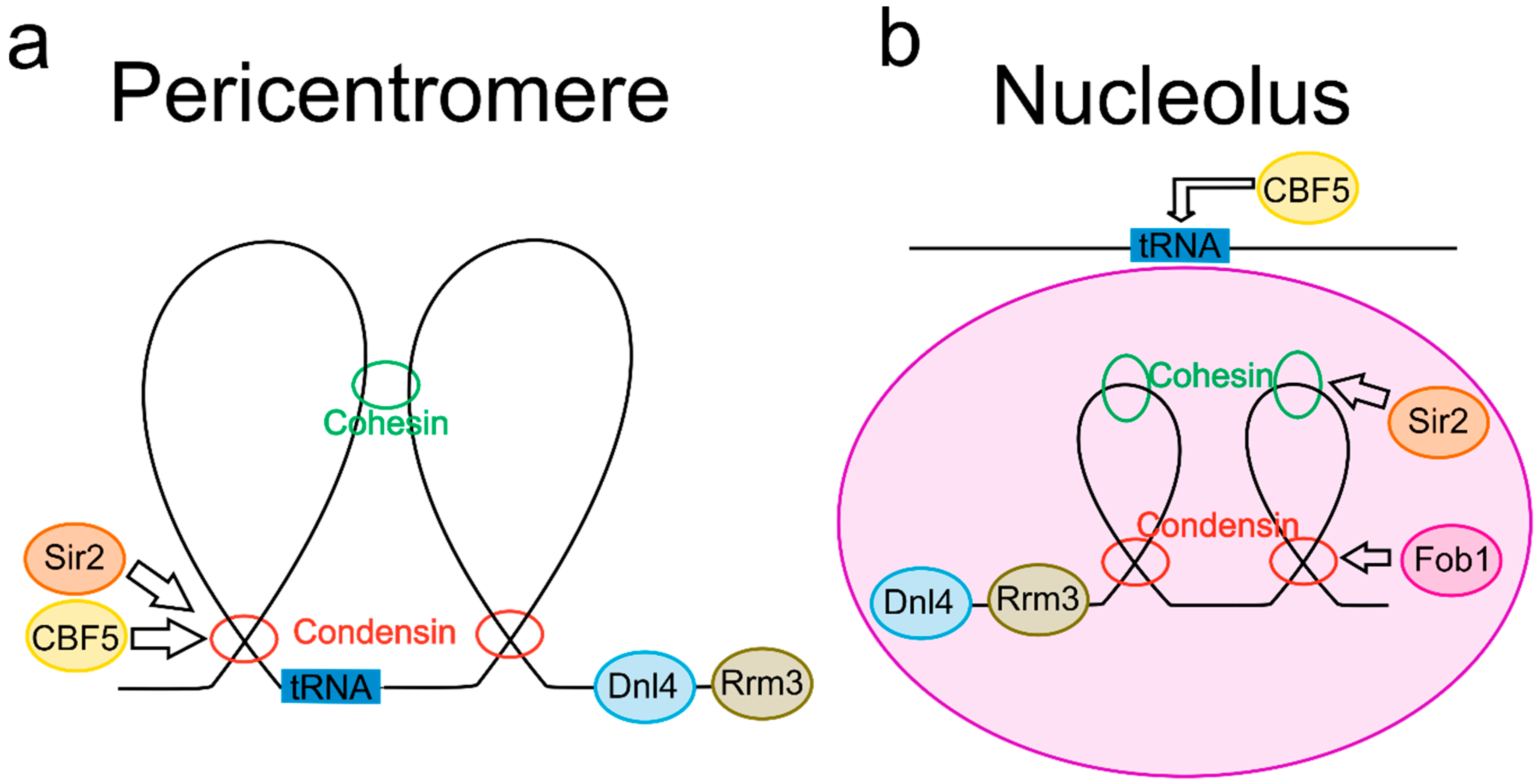
Figure 1.
DNA loops in the pericentromere and nucleolus. (a) Pericentromere loop schematic. Condensin extrudes DNA in the pericentromere [12,17,18,34] while cohesin radially links nearby loops [11,12]. Pericentric condensin enrichment is controlled by both Cbf5, a small nucleolar ribonucleoprotein, [35] and the histone deacetylase Sir2 [17]. DNA helicase Rrm3 regulates replication fork stalling at the pericentromere [36], and DNA ligase 4 (Dnl4) regulates segregation with a potential role in pericentric fork stalling as well [37,38]. tRNA genes are located in the pericentromere [39,40,41], and are associated with both condensin and cohesin [5,41,42]. (b) Nucleolus loop schematic. Condensin [43,44,45,46] and cohesin [23] both regulate loop formation in rDNA. DNA replication fork blocking protein (Fob1) regulates enrichment of condensin in rDNA [47], whereas Sir2 regulates cohesin rDNA localization [48]. Dnl4 [37] and Rrm3 [36] both control fork stalling at rDNA repeats. tRNA genes are tethered to the nucleolus in a Cbf5-dependent manner [49].
Figure 1.
DNA loops in the pericentromere and nucleolus. (a) Pericentromere loop schematic. Condensin extrudes DNA in the pericentromere [12,17,18,34] while cohesin radially links nearby loops [11,12]. Pericentric condensin enrichment is controlled by both Cbf5, a small nucleolar ribonucleoprotein, [35] and the histone deacetylase Sir2 [17]. DNA helicase Rrm3 regulates replication fork stalling at the pericentromere [36], and DNA ligase 4 (Dnl4) regulates segregation with a potential role in pericentric fork stalling as well [37,38]. tRNA genes are located in the pericentromere [39,40,41], and are associated with both condensin and cohesin [5,41,42]. (b) Nucleolus loop schematic. Condensin [43,44,45,46] and cohesin [23] both regulate loop formation in rDNA. DNA replication fork blocking protein (Fob1) regulates enrichment of condensin in rDNA [47], whereas Sir2 regulates cohesin rDNA localization [48]. Dnl4 [37] and Rrm3 [36] both control fork stalling at rDNA repeats. tRNA genes are tethered to the nucleolus in a Cbf5-dependent manner [49].

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lawrimore, C.J.; Bloom, K. Common Features of the Pericentromere and Nucleolus. Genes 2019, 10, 1029. https://doi.org/10.3390/genes10121029
AMA Style
Lawrimore CJ, Bloom K. Common Features of the Pericentromere and Nucleolus. Genes. 2019; 10(12):1029. https://doi.org/10.3390/genes10121029
Chicago/Turabian StyleLawrimore, Colleen J., and Kerry Bloom. 2019. "Common Features of the Pericentromere and Nucleolus" Genes 10, no. 12: 1029. https://doi.org/10.3390/genes10121029
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.