H3K9 Promotes Under-Replication of Pericentromeric Heterochromatin in Drosophila Salivary Gland Polytene Chromosomes


 and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drosophila Larval Culturing
2.2. Salivary Gland Polytene Chromosome Immunofluorescence
2.3. Sample Preparation for Genome Sequencing
2.4. Bioinformatics
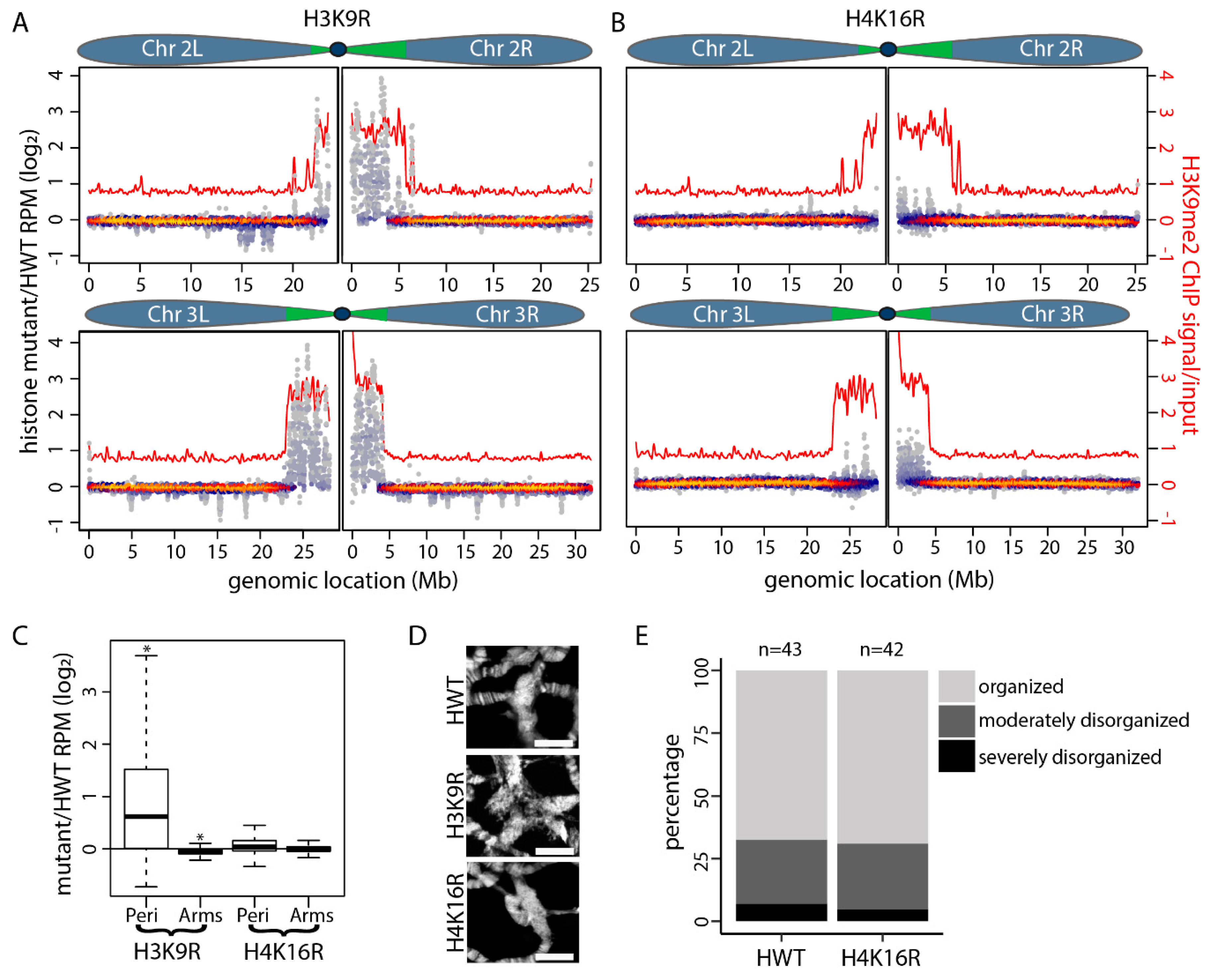
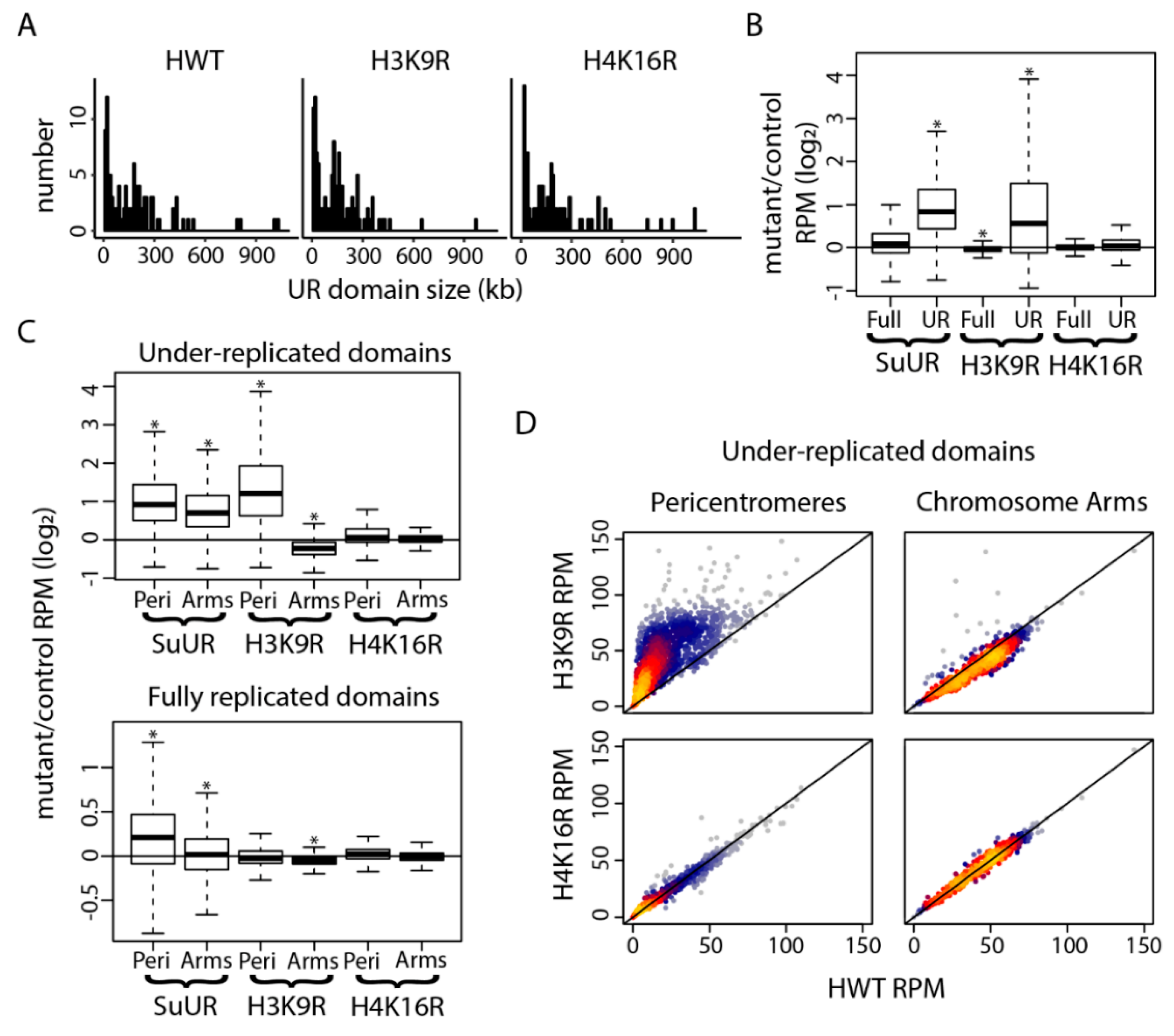
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bell, S.P. Rethinking Origin Licensing. Elife 2017, 6, e24052. [Google Scholar] [CrossRef] [PubMed]
- Zielke, N.; Edgar, B.A.; DePamphilis, M.L. Endoreplication. Cold Spring Harb. Perspect. Biol. 2013, 5, a012948. [Google Scholar] [CrossRef] [PubMed]
- Orr-Weaver, T.L. When Bigger Is Better: The Role of Polyploidy in Organogenesis. Trends Genet. 2015, 31, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Lee, O.H.; Davidson, J.M.; Duronio, R.J. Endoreplication: Polyploidy with Purpose. Genes Dev. 2009, 23, 2461–2477. [Google Scholar] [CrossRef] [PubMed]
- Fox, T.D.; Duronio, R.J. Endoreplication and Polyploidy: Insights into Development and Disease. Development 2013, 140, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Edgar, A.B.; Orr-Weaver, T.L. Endoreplication Cell Cycles: More for Less. Cell 2001, 105, 297–306. [Google Scholar] [CrossRef]
- Hua, L.B.; Orr-Weaver, T.L. DNA Replication Control During Drosophila Development: Insights into the Onset of S Phase, Replication Initiation and Fork Progression. Genetics 2017, 207, 29–47. [Google Scholar] [CrossRef]
- Lilly, A.M.; Duronio, R.J. New Insights into Cell Cycle Control from the Drosophila Endocycle. Oncogene 2005, 24, 2765–2775. [Google Scholar] [CrossRef]
- Zielke, N.; Kim, K.J.; Tran, V.; Shibutani, S.T.; Bravo, M.J.; Nagarajan, S.; van Straaten, M.; Woods, B.; von Dassow, G.; Rottig, C.; et al. Control of Drosophila Endocycles by E2f and Crl4Cdt2. Nature 2011, 480, 123–127. [Google Scholar] [CrossRef]
- Hammond, P.M.; Laird, C.D. Control of DNA Replication and Spatial Distribution of Defined DNA Sequences in Salivary Gland Cells of Drosophila melanogaster. Chromosoma 1985, 91, 279–286. [Google Scholar] [CrossRef]
- Kolesnikova, D.T.; Goncharov, F.P.; Zhimulev, I.F. Similarity in Replication Timing between Polytene and Diploid Cells Is Associated with the Organization of the Drosophila Genome. PLoS ONE 2018, 13, e0195207. [Google Scholar] [CrossRef] [PubMed]
- Spradling, A.; Orr-Weaver, T. Regulation of DNA Replication during Drosophila Development. Annu. Rev. Genet. 1987, 21, 373–403. [Google Scholar] [CrossRef] [PubMed]
- Yarosh, W.; Spradling, A.C. Incomplete Replication Generates Somatic DNA Alterations within Drosophila Polytene Salivary Gland Cells. Genes Dev. 2014, 28, 1840–1855. [Google Scholar] [CrossRef] [PubMed]
- Andreyeva, N.E.; Kolesnikova, T.D.; Belyaeva, E.S.; Glaser, R.L.; Zhimulev, I.F. Local DNA Underreplication Correlates with Accumulation of Phosphorylated H2av in the Drosophila melanogaster Polytene Chromosomes. Chromosome Res. 2008, 16, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, L.R.; Chuong, E.B.; Rivera-Mulia, J.C.; Gilbert, D.M.; Valouev, A.; Baker, J.C. Copy Number Variation Is a Fundamental Aspect of the Placental Genome. PLoS Genet. 2014, 10, e1004290. [Google Scholar] [CrossRef] [PubMed]
- Sher, N.; Bell, G.W.; Li, S.; Nordman, J.T.; Eng, T.; Eaton, M.L.; MacAlpine, D.M.; Orr-Weaver, L.T. Developmental Control of Gene Copy Number by Repression of Replication Initiation and Fork Progression. Genome Res. 2012, 22, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Eaton, L.M.; Prinz, J.A.; MacAlpine, H.K.; Tretyakov, G.; Kharchenko, P.V.; MacAlpine, D.M. Chromatin Signatures of the Drosophila Replication Program. Genome Res. 2011, 21, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Lubelsky, Y.; Prinz, J.A.; DeNapoli, L.; Li, Y.; Belsky, J.A.; MacAlpine, D.M. DNA Replication and Transcription Programs Respond to the Same Chromatin Cues. Genome Res. 2014, 24, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Bell, O.; Schwaiger, M.; Oakeley, E.J.; Lienert, F.; Beisel, C.; Stadler, M.B.; Schubeler, D. Accessibility of the Drosophila Genome Discriminates Pcg Repression, H4k16 Acetylation and Replication Timing. Nat. Struct. Mol. Biol. 2010, 17, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Pokholkova, V.G.; Koryakov, D.E.; Pindyurin, A.V.; Kozhevnikova, E.N.; Belyakin, S.N.; Andreyenkov, O.V.; Belyaeva, E.S.; Zhimulev, I.F. Tethering of Suur and Hp1 Proteins Results in Delayed Replication of Euchromatic Regions in Drosophila melanogaster Polytene Chromosomes. Chromosoma 2015, 124, 209–220. [Google Scholar] [CrossRef]
- Armstrong, L.R.; Penke, T.J.R.; Strahl, B.D.; Matera, A.G.; McKay, D.J.; MacAlpine, D.M.; Duronio, R.J. Chromatin Conformation and Transcriptional Activity Are Permissive Regulators of DNA Replication Initiation in Drosophila. Genome Res. 2018, 28, 1688–1700. [Google Scholar] [CrossRef]
- MacAlpine, M.D.; Rodriguez, H.K.; Bell, S.P. Coordination of Replication and Transcription Along a Drosophila Chromosome. Genes Dev. 2004, 18, 3094–3105. [Google Scholar] [CrossRef]
- Prioleau, M.; MacAlpine, D.M. DNA Replication Origins—Where Do We Begin? Genes Dev. 2016, 30, 1683–1697. [Google Scholar] [CrossRef] [PubMed]
- Das, P.S.; Borrman, T.; Liu, V.W.T.; Yang, S.C.-H.; Bechhoefer, J.; Rhind, N. Replication Timing Is Regulated by the Number of Mcms Loaded at Origins. Genome Res. 2015, 25, 1886–1892. [Google Scholar] [CrossRef] [PubMed]
- Rhind, N.; Yang, S.C.-H.; Bechhoefer, J. Reconciling Stochastic Origin Firing with Defined Replication Timing. Chromosome Res. 2010, 18, 35–43. [Google Scholar] [CrossRef]
- Miotto, B.; Ji, Z.; Struhl, K. Selectivity of Orc Binding Sites and the Relation to Replication Timing, Fragile Sites and Deletions in Cancers. Proc. Natl. Acad. Sci. USA 2016, 113, E4810–E4819. [Google Scholar] [CrossRef] [PubMed]
- Hua, L.B.; Bell, G.W.; Kashevsky, H.; von Stetina, J.R.; Orr-Weaver, T.L. Dynamic Changes in Orc Localization and Replication Fork Progression During Tissue Differentiation. BMC Genom. 2018, 19, 623. [Google Scholar] [CrossRef]
- Kolesnikova, D.T.; Posukh, O.V.; Andreyeva, E.N.; Bebyakina, D.S.; Ivankin, A.V.; Zhimulev, I.F. Drosophila Suur Protein Associates with Pcna and Binds Chromatin in a Cell Cycle-Dependent Manner. Chromosoma 2013, 122, 55–66. [Google Scholar] [CrossRef]
- Munden, A.; Rong, Z.; Sun, A.; Gangula, R.; Mallal, S.; Nordman, J.T. Rif1 Inhibits Replication Fork Progression and Controls DNA Copy Number in Drosophila. Elife 2018, 7, e39140. [Google Scholar] [CrossRef] [PubMed]
- Canzio, D.; Chang, E.Y.; Shankar, S.; Kuchenbecker, K.M.; Simon, M.D.; Madhani, H.D.; Narlikar, G.J.; Al-Sady, B. Chromodomain-Mediated Oligomerization of Hp1 Suggests a Nucleosome-Bridging Mechanism for Heterochromatin Assembly. Mol. Cell 2011, 41, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.A.; Elnatan, D.; Keenen, M.M.; Trnka, M.J.; Johnston, J.B.; Burlingame, A.L.; Agard, D.A.; Redding, S.; Narlikar, G.J. Liquid Droplet Formation by Hp1α Suggests a Role for Phase Separation in Heterochromatin. Nature 2017, 547, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Strom, R.A.; Emelyanov, A.V.; Mir, M.; Fyodorov, D.V.; Darzacq, X.; Karpen, G.H. Phase Separation Drives Heterochromatin Domain Formation. Nature 2017, 547, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Makunin, V.I.; Volkova, E.I.; Belyaeva, E.S.; Nabirochkina, E.N.; Pirrotta, V.; Zhimulev, I.F. The Drosophila Suppressor of Underreplication Protein Binds to Late-Replicating Regions of Polytene Chromosomes. Genetics 2002, 160, 1023–1034. [Google Scholar] [PubMed]
- Belyaeva, S.E.; Zhimulev, I.F.; Volkova, E.I.; Alekseyenko, A.A.; Moshkin, Y.M.; Koryakov, D.E. Su(Ur)Es: A Gene Suppressing DNA Underreplication in Intercalary and Pericentric Heterochromatin of Drosophila melanogaster Polytene Chromosomes. Proc. Natl. Acad. Sci. USA 1998, 95, 7532–7537. [Google Scholar] [CrossRef] [PubMed]
- Nordman, T.J.; Kozhevnikova, E.N.; Verrijzer, C.P.; Pindyurin, A.V.; Andreyeva, E.N.; Shloma, V.V.; Zhimulev, I.F.; Orr-Weaver, T.L. DNA Copy-Number Control through Inhibition of Replication Fork Progression. Cell Rep. 2014, 9, 841–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pindyurin, V.A.; Boldyreva, L.V.; Shloma, V.V.; Kolesnikova, T.D.; Pokholkova, G.V.; Andreyeva, E.N.; Kozhevnikova, E.N.; Ivanoschuk, I.G.; Zarutskaya, E.A.; Demakov, S.A.; et al. Interaction between the Drosophila Heterochromatin Proteins Suur and Hp1. J. Cell Sci. 2008, 121, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Andreyeva, N.E.; Bernardo, T.J.; Kolesnikova, T.D.; Lu, X.; Yarinich, L.A.; Bartholdy, B.A.; Guo, X.; Posukh, O.V.; Healton, S.; Willcockson, M.A.; et al. Regulatory Functions and Chromatin Loading Dynamics of Linker Histone H1 During Endoreplication in Drosophila. Genes Dev. 2017, 31, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Lucia, D.F.; Ni, J.; Vaillant, C.; Sun, F. Hp1 Modulates the Transcription of Cell-Cycle Regulators in Drosophila melanogaster. Nucleic Acids Res. 2005, 33, 2852–2858. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.J.; Karpen, G.H. Heterochromatin Genome Stability Requires Regulators of Histone H3 K9 Methylation. PLoS Genet. 2009, 5, e1000435. [Google Scholar] [CrossRef]
- Li, X.; Corsa, C.A.S.; Pan, P.W.; Wu, L.; Ferguson, D.; Yu, X.; Min, J.; Dou, Y. Mof and H4 K16 Acetylation Play Important Roles in DNA Damage Repair by Modulating Recruitment of DNA Damage Repair Protein Mdc1. Mol. Cell. Biol. 2010, 30, 5335–5347. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, D.; Liu, Y.; Chen, S.; Sun, F. Histone Acetyltransferase Hmof Promotes S Phase Entry and Tumorigenesis in Lung Cancer. Cell. Signal. 2013, 25, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.R.; Reinberg, D. Is There a Code Embedded in Proteins That Is Based on Post-Translational Modifications? Nat. Rev. Mol. Cell Biol. 2008, 9, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Berger, S.L. The Emerging Field of Dynamic Lysine Methylation of Non-Histone Proteins. Curr. Opin. Genet. Dev. 2008, 18, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Glozak, A.M.; Sengupta, N.; Zhang, X.; Seto, E. Acetylation and Deacetylation of Non-Histone Proteins. Gene 2005, 363, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Günesdogan, U.; Jäckle, H.; Herzig, A. A Genetic System to Asses in Vivo the Functions of Histones and Histone Modifications in Higher Eukaryotes. EMBO Rep. 2010, 11, 772–776. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.D.; Klusza, S.; Penke, T.J.R.; Meers, M.P.; Curry, K.P.; McDaniel, S.L.; Malek, P.Y.; Cooper, S.W.; Tatomer, D.C.; Lieb, J.D.; et al. Interrogating the Function of Metazoan Histones Using Engineered Gene Clusters. Dev. Cell 2015, 32, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.L.; Duronio, R.J. Cell Cycle Analysis in H4K16R Diploid Wing Imaginal Disc Cells. unpublished.
- Penke, R.T.J.; McKay, D.J.; Strahl, B.D.; Matera, A.G.; Duronio, R.J. Direct Interrogation of the Role of H3k9 in Metazoan Heterochromatin Function. Genes Dev. 2016, 30, 1866–1880. [Google Scholar] [CrossRef]
- Penke, R.T.J.; McKay, D.J.; Strahl, B.D.; Matera, A.G.; Duronio, R.J. Functional Redundancy of Variant and Canonical Histone H3 Lysine 9 Modification in Drosophila. Genet. 2018, 208, 229–244. [Google Scholar] [CrossRef]
- Quinlan, R.A.; Hall, I.M. Bedtools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Abyzov, A.; Urban, A.E.; Snyder, M.; Gerstein, M. Cnvnator: An Approach to Discover, Genotype and Characterize Typical and Atypical Cnvs from Family and Population Genome Sequencing. Genome Res. 2011, 21, 974–984. [Google Scholar] [CrossRef]
- Riddle, C.N.; Minoda, A.; Kharchenko, P.V.; Alekseyenko, A.A.; Schwartz, Y.B.; Tolstorukov, M.Y.; Gorchakov, A.A.; Jaffe, J.D.; Kennedy, C.; Linder-Basso, D.; et al. Plasticity in Patterns of Histone Modifications and Chromosomal Proteins in Drosophila Heterochromatin. Genome Res. 2011, 21, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, A.R.; Carlson, J.W.; Wan, K.H.; Park, S.; Mendez, I.; Galle, S.E.; Booth, B.W.; Pfeiffer, B.D.; George, R.A.; Svirskas, R.; et al. The Release 6 Reference Sequence of the Drosophila melanogaster Genome. Genome Res. 2015, 25, 445–458. [Google Scholar] [CrossRef]
- Slade, D. Maneuvers on Pcna Rings During DNA Replication and Repair. Genes 2018, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.G.; Kedziora, K.M.; Limas, J.C.; Cook, J.G.; Purvis, J.E. Accurate Delineation of Cell Cycle Phase Transitions in Living Cells with Pip-Fucci. Cell Cycle 2018, 17, 2496–2516. [Google Scholar] [CrossRef] [PubMed]
- Kharchenko, V.P.; Aleksyenko, A.A.; Schwartz, Y.B.; Minoda, A.; Riddle, N.C.; Ernst, J.; Sobo, P.J.; Larschan, E.; Gorchakov, A.A.; Gu, T.; et al. Comprehensive Analysis of the Chromatin Landscape in Drosophila. Nature 2011, 471, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Copur, O.; Gorchakov, A.; Finkl, K.; Kuroda, M.I.; Muller, J. Sex-Specific Phenotypes of Histone H4 Point Mutants Establish Dosage Compensation as the Critical Function of H4k16 Acetylation in Drosophila. Proc. Natl. Acad. Sci. USA 2018, 115, 13336–13341. [Google Scholar] [CrossRef] [PubMed]
- Seller, A.C.; O’Farrell, P.H. Rif1 Prolongs the Embryonic S Phase at the Drosophila Mid-Blastula Transition. PLoS Biol. 2018, 16, e2005687. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armstrong, R.L.; Penke, T.J.R.; Chao, S.K.; Gentile, G.M.; Strahl, B.D.; Matera, A.G.; McKay, D.J.; Duronio, R.J. H3K9 Promotes Under-Replication of Pericentromeric Heterochromatin in Drosophila Salivary Gland Polytene Chromosomes. Genes 2019, 10, 93. https://doi.org/10.3390/genes10020093
Armstrong RL, Penke TJR, Chao SK, Gentile GM, Strahl BD, Matera AG, McKay DJ, Duronio RJ. H3K9 Promotes Under-Replication of Pericentromeric Heterochromatin in Drosophila Salivary Gland Polytene Chromosomes. Genes. 2019; 10(2):93. https://doi.org/10.3390/genes10020093
Chicago/Turabian StyleArmstrong, Robin L., Taylor J. R. Penke, Samuel K. Chao, Gabrielle M. Gentile, Brian D. Strahl, A. Gregory Matera, Daniel J. McKay, and Robert J. Duronio. 2019. "H3K9 Promotes Under-Replication of Pericentromeric Heterochromatin in Drosophila Salivary Gland Polytene Chromosomes" Genes 10, no. 2: 93. https://doi.org/10.3390/genes10020093