
A Computational Approach for Molecular Characterization of Covaxin (BBV152) and Its Ingredients for Assessing Its Efficacy against COVID-19



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Analysis
2.2. Investigation of S Protein-ACE2 Interaction in the Presence of 2-PE and Imidazoquinolinone
2.3. Docking Analysis between ACE2, HSA, and S Protein with 2-PE and Imidazoquinolinone
2.4. Drug-Likeness Analysis of 2-PE and Imidazoquinolinone
3. Results and Discussion
3.1. Physicochemical Properties and Structural Analysis of S Protein, ACE2, and HSA
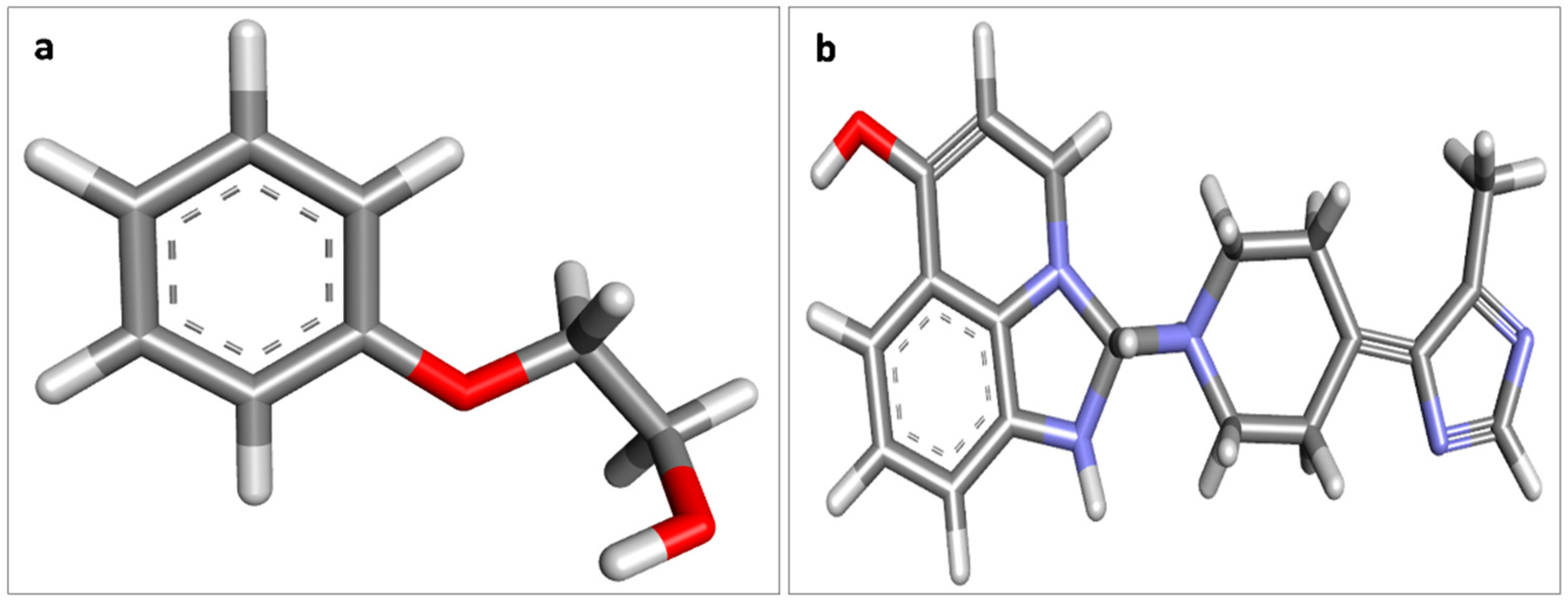
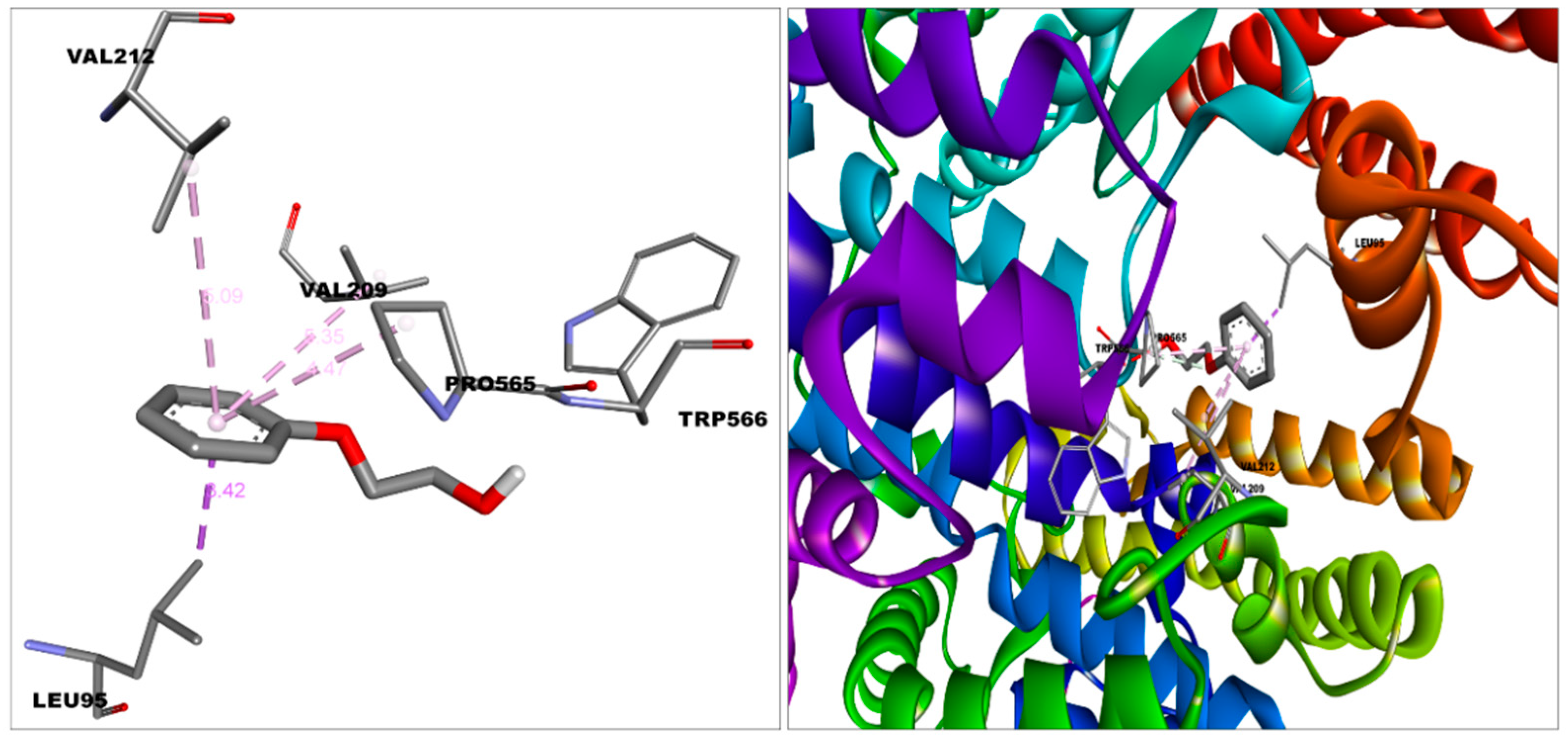
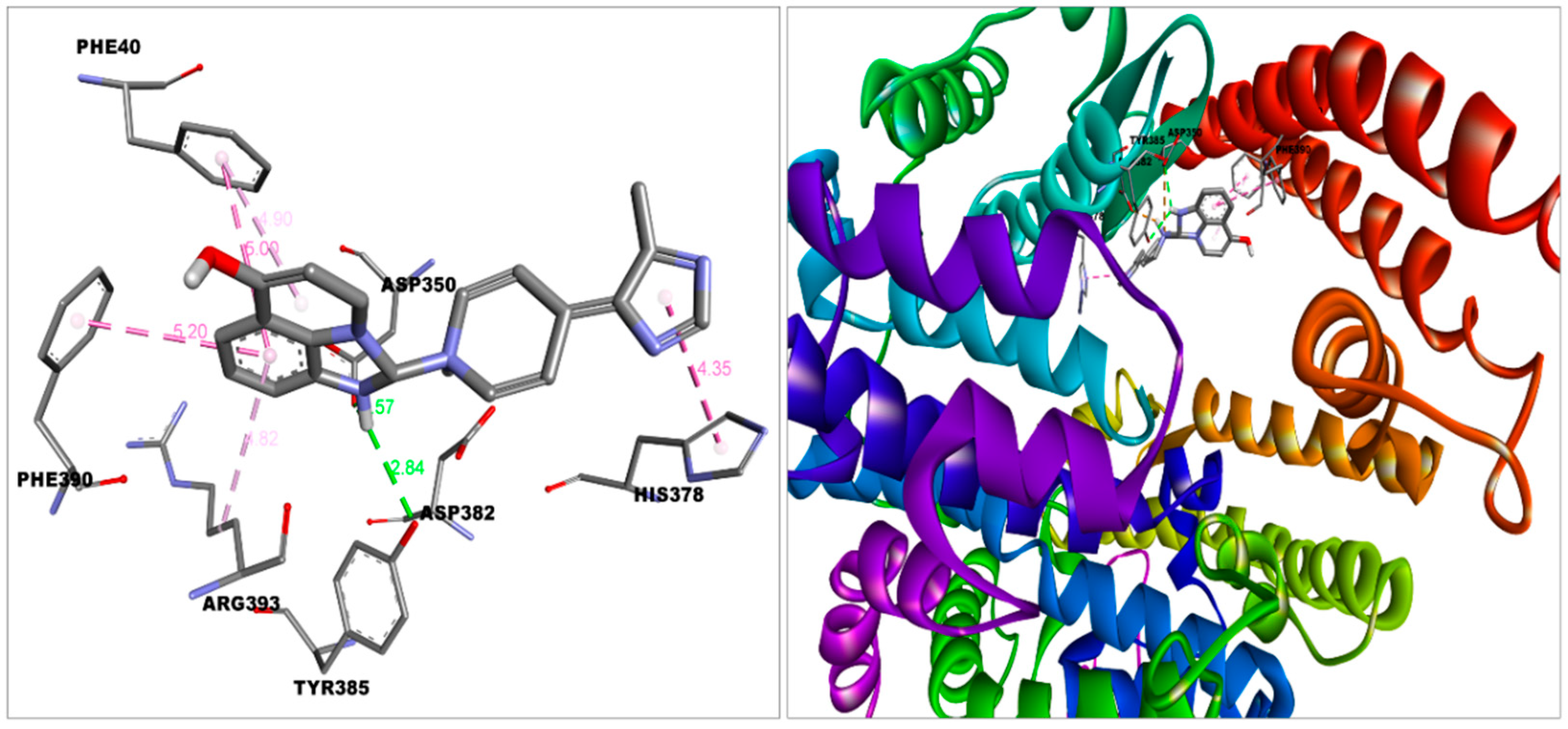
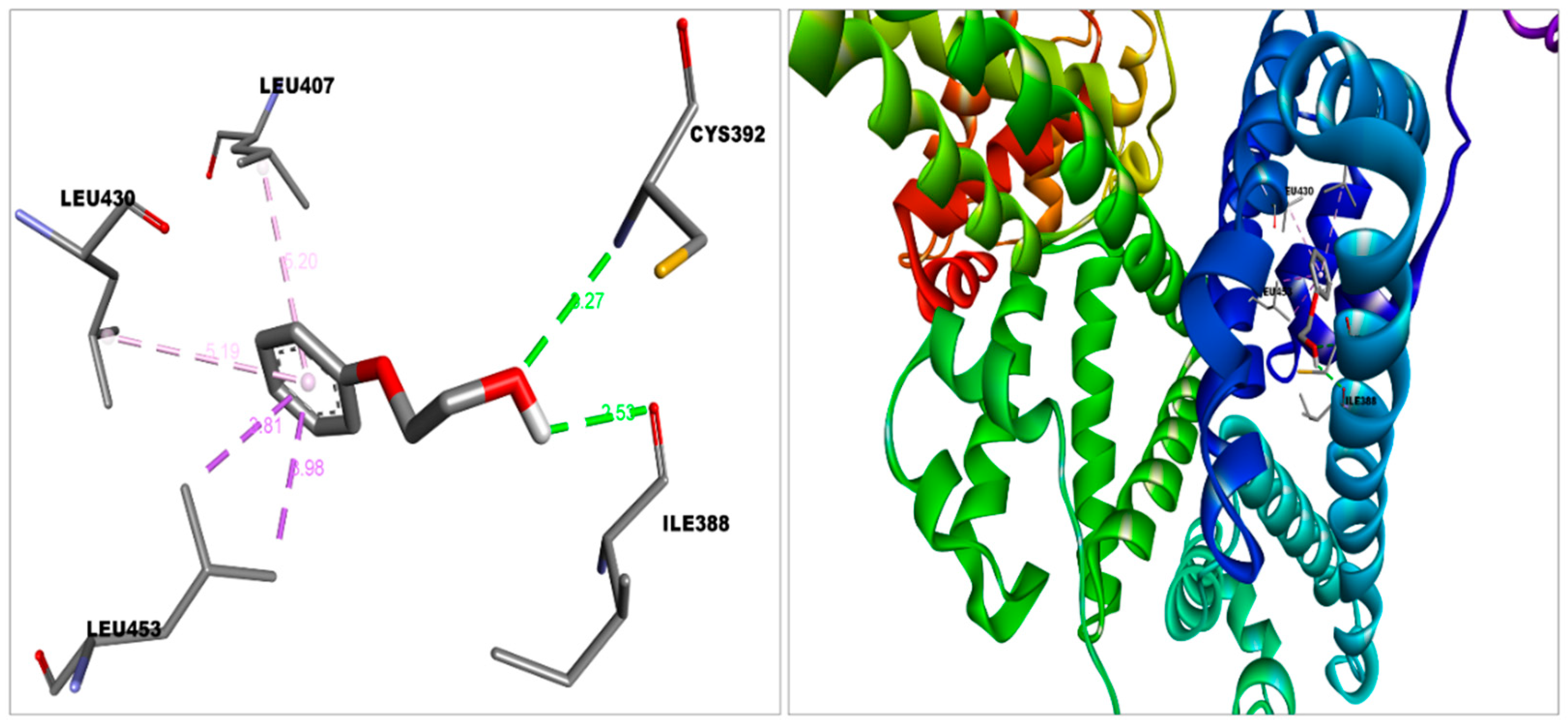
3.2. Molecular Interactions between Covaxin Adjuvant with S Protein, ACE2, and HSA
3.3. Protein–Protein Interaction Study in the Presence of Covaxin Adjuvants with S Protein, ACE2, and HSA
3.4. Drug Likeliness Analysis of Covaxin Ingredients
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seitz, B.M.; Aktipis, A.; Buss, D.M.; Alcock, J.; Bloom, P.; Gelfand, M.; Haselton, M.G. The pandemic exposes human nature: 10 evolutionary insights. Proc. Natl. Acad. Sci. USA 2020, 117, 27767–27776. [Google Scholar] [CrossRef] [PubMed]
- Talukder, A.; Kalita, C.; Neog, N.; Goswami, C.; Sarma, M.K.; Hazarika, I. A comparative analysis on the safety and efficacy of Covaxin versus other vaccines against COVID-19: A review. Z. Für Nat. C 2022, 77, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Darbar, S.; Agarwal, S.; Saha, S. COVID-19 Vaccine: COVAXIN®-India’s First Indigenous Effective Weapon to Fight against Coronavirus (A Review). Parana J. Sci. Educ. 2021, 7, 1–9. [Google Scholar]
- Ganneru, B.; Jogdand, H.; Daram, V.K.; Das, D.; Molugu, N.R.; Prasad, S.D.; Kannappa, S.V.; Ella, K.M.; Ravikrishnan, R.; Vadrevu, K.M. Th1 skewed immune response of whole virion inactivated SARS CoV 2 vaccine and its safety evaluation. iScience 2021, 24, 102298. [Google Scholar] [CrossRef] [PubMed]
- Ganapathi, L.; Van Haren, S.; Dowling, D.J.; Bergelson, I.; Shukla, N.M.; Malladi, S.S.; Balakrishna, R.; Tanji, H.; Ohto, U.; Shimizu, T.; et al. The Imidazoquinoline Toll-Like Receptor-7/8 Agonist Hybrid-2 Potently Induces Cytokine Production by Human Newborn and Adult Leukocytes. PLoS ONE 2015, 10, e0134640. [Google Scholar] [CrossRef]
- Gupta, R.K.; Siber, G.R. Adjuvants for human vaccines—Current status, problems and future prospects. Vaccine 1995, 13, 1263–1276. [Google Scholar] [CrossRef]
- Veve, M.P. Side Effects of Drugs Annual. Vaccines 2018, 42, 383–413. [Google Scholar] [CrossRef]
- Vidor, E. Poliovirus vaccine-inactivated. Vaccines 2013, 573–597. [Google Scholar] [CrossRef]
- Miller, S.M.; Cybulski, V.; Whitacre, M.; Bess, L.S.; Livesay, M.T.; Walsh, L.; Burkhart, D.; Bazin, G.; Evans, J.T. Novel Lipidated Imidazoquinoline TLR7/8 Adjuvants Elicit Influenza-Specific Th1 Immune Responses and Protect Against Heterologous H3N2 Influenza Challenge in Mice. Front. Immunol. 2020, 11, 406. [Google Scholar] [CrossRef]
- Dréno, B.; Zuberbier, T.; Gelmetti, C.; Gontijo, G.; Marinovich, M. Safety review of phenoxyethanol when used as a preservative in cosmetics. J. Eur. Acad. Derm. Venereol. 2019, 33, 15–24. [Google Scholar] [CrossRef]
- Hua, D.; Xu, P. Recent advances in biotechnological production of 2-phenyl ethanol. Biotechnol. Adv. 2011, 29, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.S.; Fatemi, R.; Winlow, W. SARS-CoV-2 Bound Human Serum Albumin and Systemic Septic Shock. Front. Cardiovasc. Med. 2020, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Guido, T.C.; Louro, S.R.; Anteneodo, C. Hydration of hydrophobic biological porphyrins. J. Chem. Physics. 2011, 134, 02B608. [Google Scholar] [CrossRef]
- Guido, T.R.; Louro, S.R.; Pascutti, P.G.; Anteneodo, C. Solvation of anionic water—Soluble porphyrins: A computational study. Int. J. Quantum. Chem. 2010, 110, 2094–2100. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Qadeer, A.A.; McKay, M.R. Preliminary identification of potential vaccine targets for the COVID-19 coronavirus (SARS-CoV-2) based on SARS-CoV immunological studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef]
- Gu, W.; Zhou, T.; Ma, J.; Sun, X.; Lu, Z. Analysis of synonymous codon usage in SARS Coronavirus and other Nidovirales. Virus Res. 2004, 101, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Bosch, B.J.; Van der Zee, R.; De Haan, C.A.; Rottier, P.J. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef]
- Jena, A.B.; Kanungo, N.; Nayak, V.; Chainy, G.B.N.; Dandapat, J. Catechin and curcumin interact with S protein of SARS-CoV2 and ACE2 of human cell membrane: Insights from computational studies. Sci. Rep. 2021, 11, 2043. [Google Scholar] [CrossRef]
- Mallick, R.; Duttaroy, A.K. Origin and Structural Biology of Novel Coronavirus (SARS-CoV-2). Adv. Exp. Med. Biol. 2021, 1352, 1–13. [Google Scholar] [CrossRef]
- Walls, A.C.; Tortorici, M.A.; Frenz, B.; Snijder, J.; Li, W.; Rey, F.A.; DiMaio, F.; Bosch, B.J.; Veesler, D. Glycan shield and epitope masking of a coronavirus spike protein observed by cryo-electron microscopy. Nat. Struct. Mol. Biol. 2016, 23, 899–905. [Google Scholar] [CrossRef]
- Millet, J.K.; Whittaker, G.R. Host cell proteases: Critical determinants of coronavirus tropism and pathogenesis. Virus Res. 2015, 202, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. Proteom. Protoc. Handb. 2005, 571–607. [Google Scholar]
- Geourjon, C.; Deleage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein–protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef]
- Pi, M.; Kapoor, K.; Ye, R.; Nishimoto, S.K.; Smith, J.C.; Baudry, J.; Quarles, L.D. Evidence for Osteocalcin Binding and Activation of GPRC6A in β-Cells. Endocrinology 2016, 157, 1866–1880. [Google Scholar] [CrossRef]
- Jena, A.B.; Kanungo, N.; Chainy, G.B.N.; Devaraji, V.; Dandapat, J. 8-Hydroxydihydrosanguinarine (8-HDS), a pyridone containing analogue of sanguinarine, can be a potential inhibitor of S protein and M protease of SARS CoV2: Insights from computational studies. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Dassault Systemes BIOVIA. BIOVIA Discovery Studio—BIOVIA—Dassault Systèmes®, 2017. Available online: https://www.3ds.com/products-services/biovia/products/molecular-modeling-simulation/biovia-discovery-studio/(accessed on 29 September 2021).
- Pulendran, B.; Arunachalam, P.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef]
- Jena, A.B.; Samal, R.R.; Kumari, K.; Pradhan, J.; Chainy, G.B.; Subudhi, U.; Pal, S.; Dandapat, J. The benzene metabolite p-benzoquinone inhibits the catalytic activity of bovine liver catalase: A biophysical study. Int. J. Biol. Macromol. 2021, 167, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Oprea, T.I.; Bauman, J.E.; Bologa, C.G.; Buranda, T.; Chagaev, A.; Edwards, B.S.; Jarvik, J.W.; Gresham, H.D.; Haynes, M.K.; Hjelle, B.; et al. Drug repurposing from an academic perspective. Drug Discov. Today Ther. Strat. 2011, 8, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Wrap, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Lee, P.; Wu, X. Modifications of human serum albumin and their binding effect. Curr. Pharm. Des. 2015, 21, 1862–1865. [Google Scholar] [CrossRef] [PubMed]
- Sprinzak, E.; Sattath, S.; Margalit, H. How reliable are experimental protein-protein interaction data? J. Mol. Biol. 2003, 327, 919–923. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A BOILED-Egg To Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. Chem. Med. Chem. 2016, 11, 1117–1121. [Google Scholar] [CrossRef]
- Lounkine, E.; Keiser, M.J.; Whitebread, S.; Mikhailov, D.; Hamon, J.; Jenkins, J.L.; Lavan, P.; Weber, E.; Doak, A.K.; Côté, S.; et al. Large-scale prediction and testing of drug activity on side-effect targets. Nature 2012, 486, 361–367. [Google Scholar] [CrossRef]
- Keiser, M.J.; Setola, V.; Irwin, J.J.; Laggner, C.; Abbas, A.I.; Hufeisen, S.J.; Jensen, N.H.; Kuijer, M.B.; Matos, R.C.; Tran, T.B.; et al. Predicting new molecular targets for known drugs. Nature 2009, 462, 175–181. [Google Scholar] [CrossRef]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A web server for target prediction of bioactive small molecules. Nucleic Acids. Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef]
- Kola, I.; Landis, J. Can the pharmaceutical industry reduce attrition rates? Nat. Rev. Drug Discov. 2004, 3, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Braun, P.; Gingras, A.C. History of protein-protein interactions: From egg-white to complex networks. Proteomics 2012, 12, 1478–1498. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.S.; Srinivas, K.; Sujini, G.N.; Kumar, G.N. Protein-protein interaction detection: Methods and analysis. Int. J. Proteom. 2014, 147648. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein–Ligand | Binding Affinity (Kcal/mol) | Types of Interaction | Interacting AA Name; AA No |
|---|---|---|---|
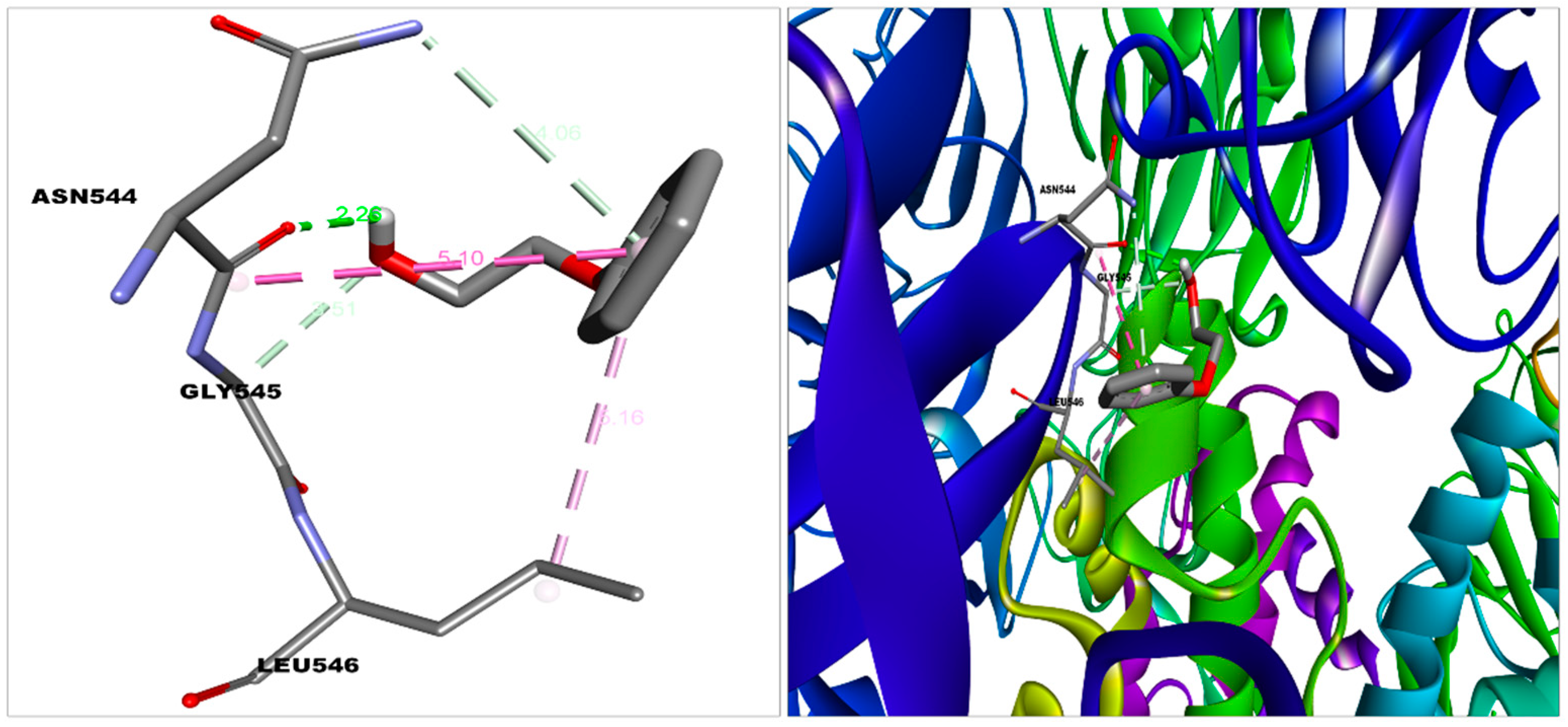
| S protein—2-PE | −5.2 | Van der Waal | Gln564, Phe565, Val576, Phe543, Leu517, Cys391, Ala522, Leu518, Pro521 |
| Carbon hydrogen bond, Pi donor hydrogen bond | Gly545 | ||
| Conventional hydrogen bond | Asn544 | ||
| Pi-alkyl | Leu546 | ||
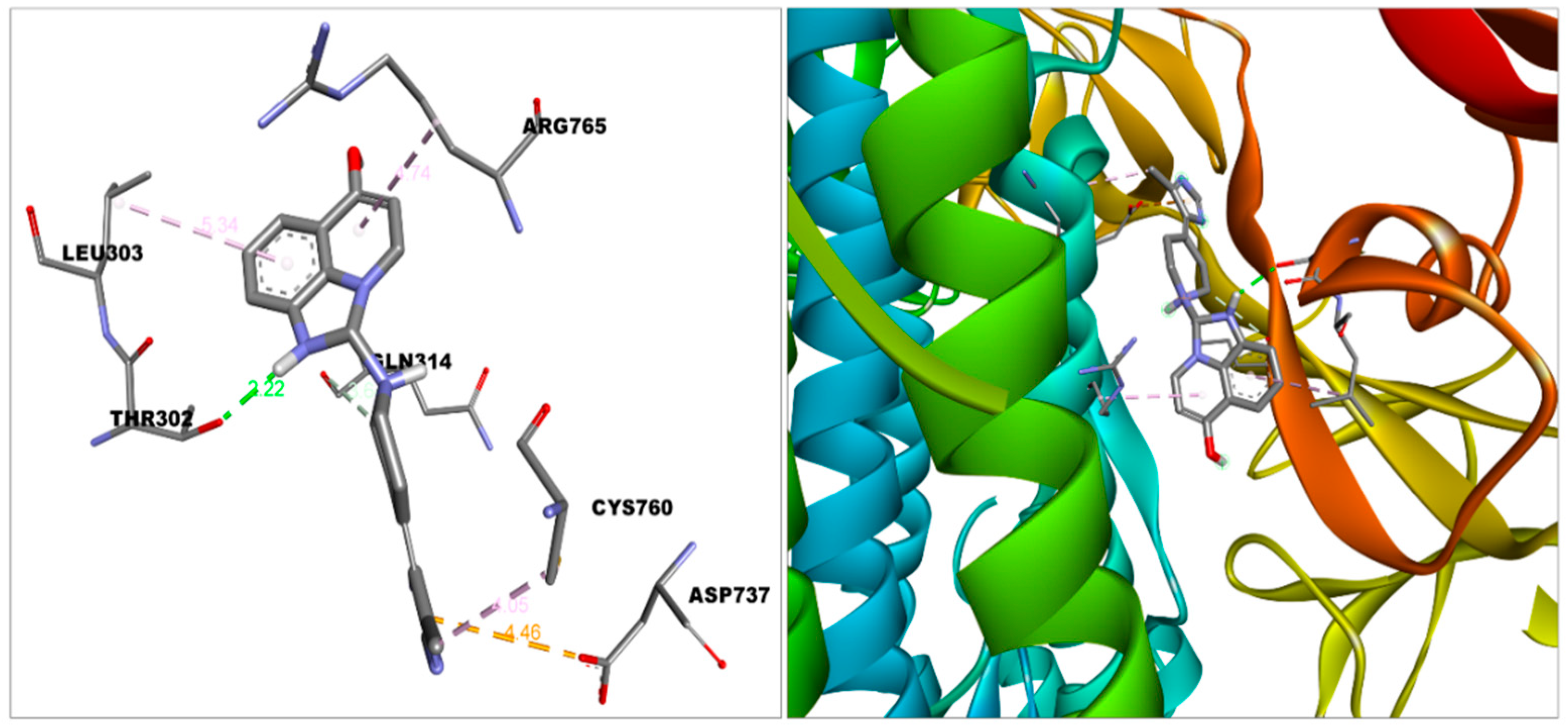
| S protein—2-Imidazoquinolinone | −8.5 | Van der Waal | Asn317, Ser316, Thr315, Thr761, Thr302, Tyr313, Thr768, Gln314, Asn764, Thr739 |
| Conventional hydrogen bond | Thr302 | ||
| Pi-Anion | Asp-737 | ||
| Pi-alkyl | Cys760, Leu303 | ||
| Alkyl | Arg765 | ||
| ACE2—2-PE | −5.3 | Van der Waal | Leu91, Asn210, Lys562, Ala396, Glu564 |
| Carbon hydrogen bond | Pro565 | ||
| Pi-alkyl | Val212,Val209 | ||
| Pi-sigma | Leu95 | ||
| Unfavourable donor donor | Trp566 | ||
| ACE2—Imidazoquinolinone | −8.5 | Van der Waal | Ala348, Thr347, Glu402, His401, Trp69 |
| Conventional hydrogen bond | Tyr385 | ||
| Pi-alkyl | Arg393 | ||
| Pi-Pi stacked | Phe390, Phe40 | ||
| Pi-Pi T-shaped | His378 | ||
| Salt bridge | Asp350, Asp382 | ||
| HSA—2-PE | −5.3 (Domain-IIIA) | Van der Waal | Asn391, Ala449, Leu387, Val433, Phe403, Tyr411 |
| Conventional hydrogen bond | Cys392, Ile388 | ||
| Pi-sigma | Leu453 | ||
| Pi-alkyl | Leu430, Leu407 | ||
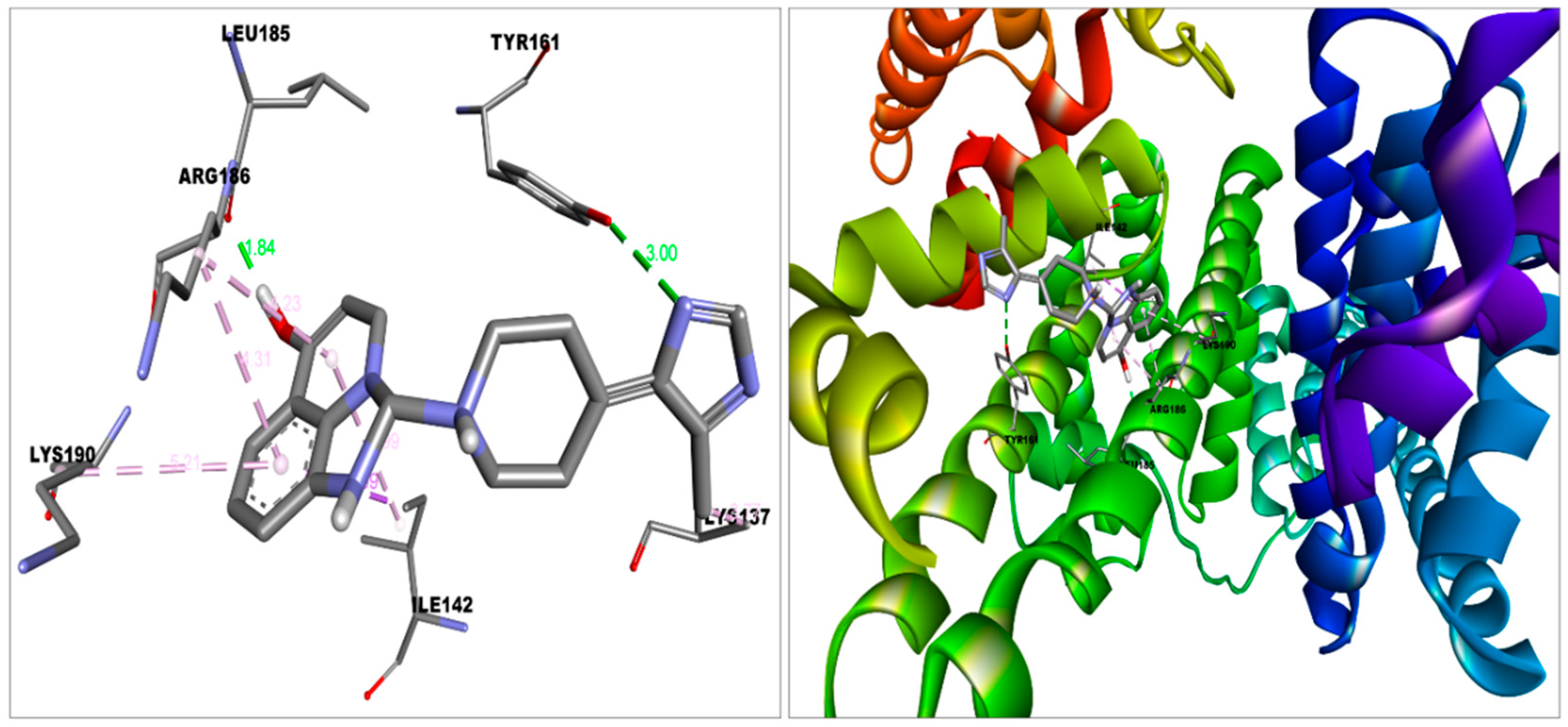
| HAS—Imidazoquinolinone | −9.1 (Domain-IB) | Van der Waal | Met123, Phe134, Glu141, Tyr138, Phe157, Gly189, His146, Leu115 |
| Conventional hydrogen bond | Tyr161, Leu185 | ||
| Pi-sigma | Ile142 | ||
| Pi-alkyl | Arg186, Lys190 | ||
| Alkyl | Lys137 |
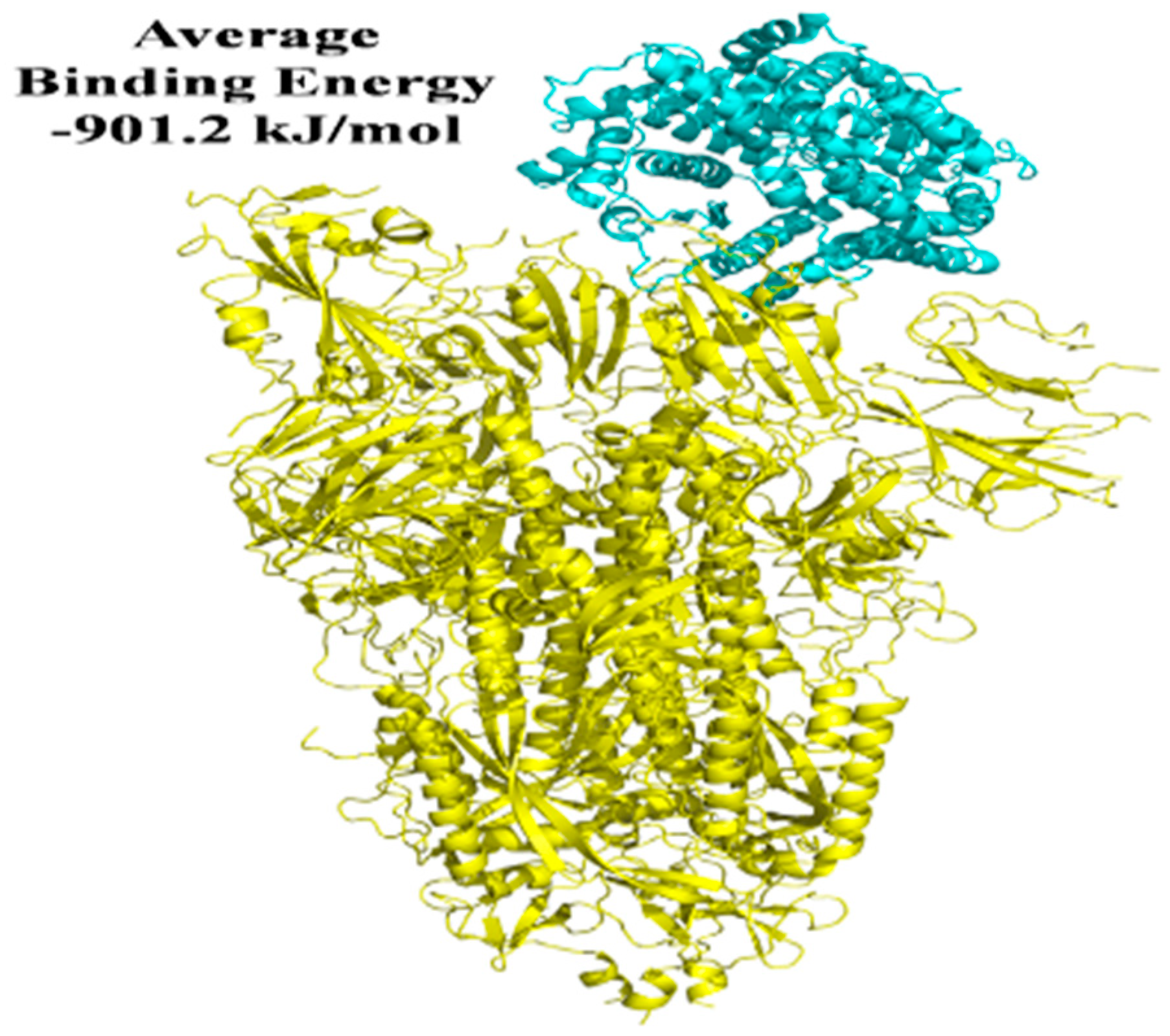
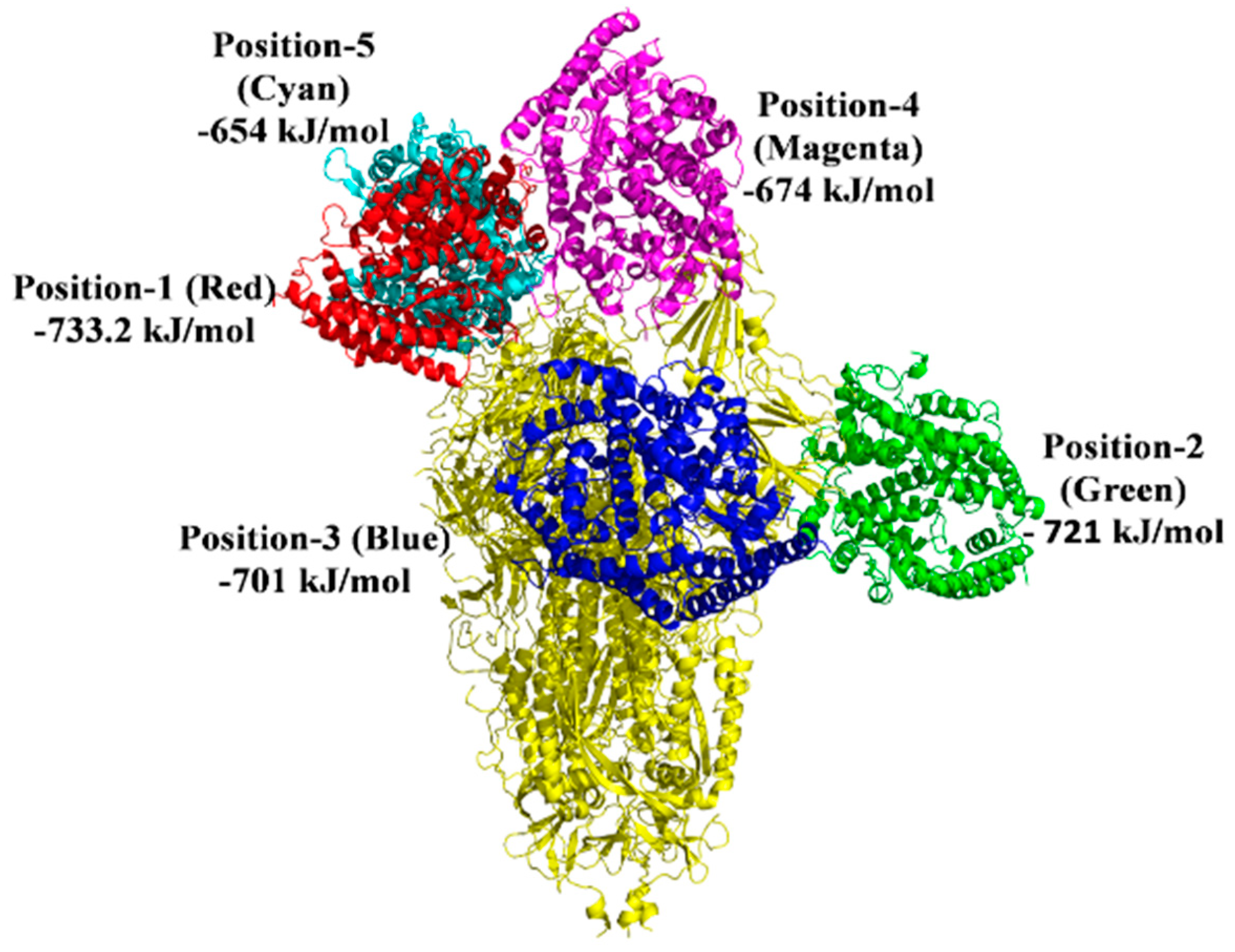
| Macromolecule | Binding Positions | Lowest Energy (kJ/mol) | Average Lowest Energy (kJ/mol) |
|---|---|---|---|
| S protein-ACE2 | 1 | −928.9 | −901.2 |
| S protein-ACE2 | 2 | −923 | |
| S protein-ACE2 | 3 | −902.4 | |
| S protein-ACE2 | 4 | −853.3 | |
| S protein-ACE2 | 5 | −898.4 | |
| S protein with 2-PE-ACE2 | 1 | −733.2 | −696.64 |
| S protein with 2-PE-ACE2 | 2 | −721 | |
| S protein with 2-PE-ACE2 | 3 | −701 | |
| S protein with 2-PE-ACE2 | 4 | −674 | |
| S protein with 2-PE-ACE2 | 5 | −654 | |
| S protein with imidazoquinolinone-ACE2 | 1 | −630.9 | −589.46 |
| S protein with imidazoquinolinone-ACE2 | 2 | −605.6 | |
| S protein with imidazoquinolinone-ACE2 | 3 | −567.8 | |
| S protein with imidazoquinolinone-ACE2 | 4 | −544.1 | |
| S protein with imidazoquinolinone-ACE2 | 5 | −598.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jena, A.B.; Duttaroy, A.K. A Computational Approach for Molecular Characterization of Covaxin (BBV152) and Its Ingredients for Assessing Its Efficacy against COVID-19. Future Pharmacol. 2022, 2, 306-319. https://doi.org/10.3390/futurepharmacol2030021
Jena AB, Duttaroy AK. A Computational Approach for Molecular Characterization of Covaxin (BBV152) and Its Ingredients for Assessing Its Efficacy against COVID-19. Future Pharmacology. 2022; 2(3):306-319. https://doi.org/10.3390/futurepharmacol2030021
Chicago/Turabian StyleJena, Atala B., and Asim K. Duttaroy. 2022. "A Computational Approach for Molecular Characterization of Covaxin (BBV152) and Its Ingredients for Assessing Its Efficacy against COVID-19" Future Pharmacology 2, no. 3: 306-319. https://doi.org/10.3390/futurepharmacol2030021