In Vitro Assessment of the Impact of Industrial Processes on the Gastrointestinal Digestion of Milk Protein Matrices Using the INFOGEST Protocol

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Characteristics of the Milk Matrices
2.1.2. Estimation of Free Calcium Concentrations
2.2. Static In Vitro Gastrointestinal Digestion of the Milk Matrices
2.3. Biochemical Characterization
2.3.1. Peptide Concentration Determination
2.3.2. Quantification of Free Amino Acids
2.4. Peptide Molecular Weight Profiles
2.5. Gel Electrophoresis
2.6. Peptide Identification by HPLC-ESI-qTOF-MS/MS and Database Search
2.7. Bioinformatics Retreatment of Chromatographic and Mass Spectrometry Data
2.8. Amino Acid Occurrence
2.9. Multi Block-Based Statistical Analysis
3. Results
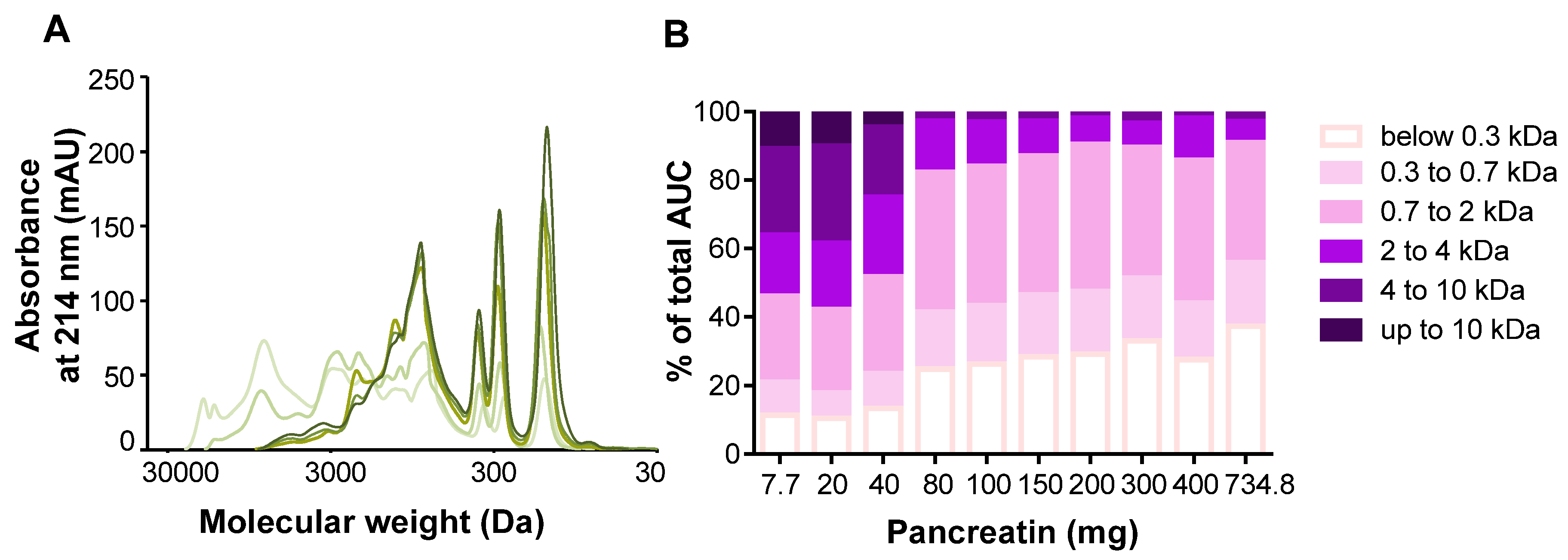
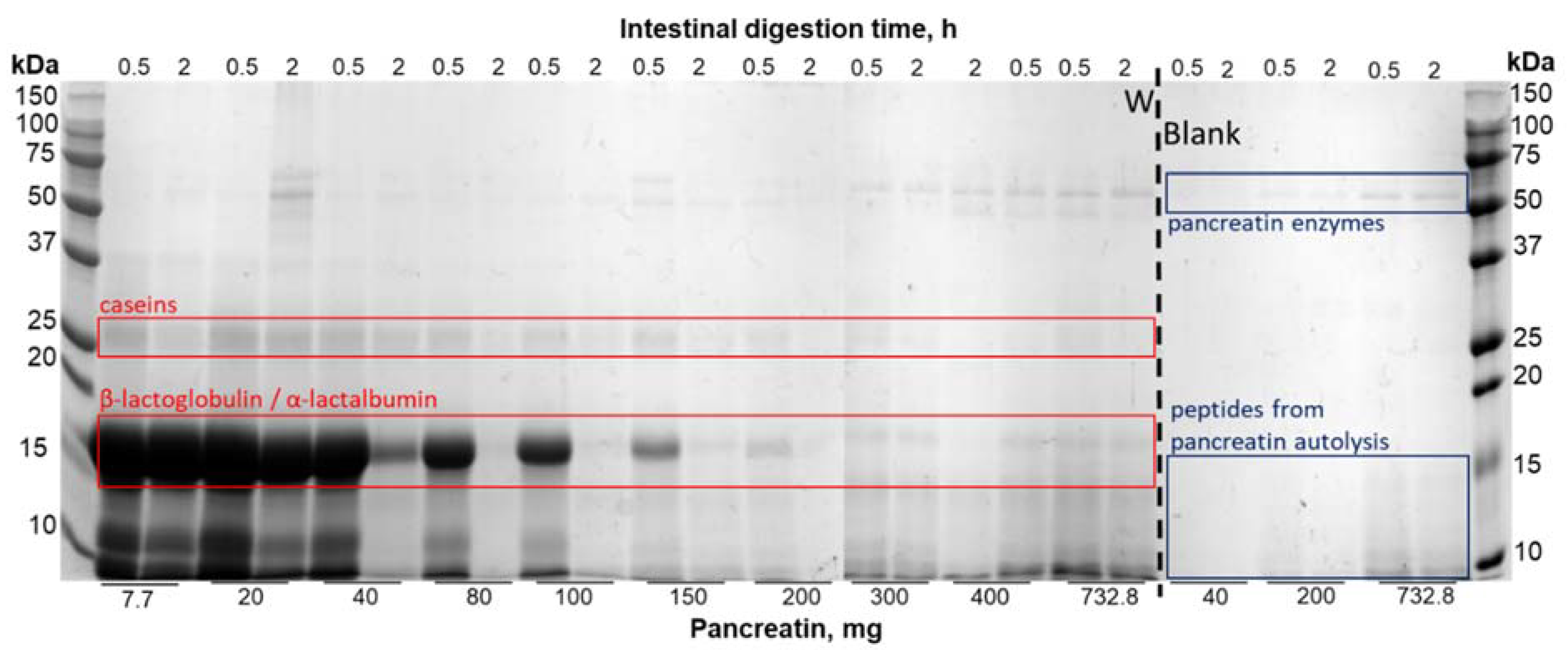
3.1. Optimization of the INFOGEST Digestion Protocol for Dairy Protein Matrices
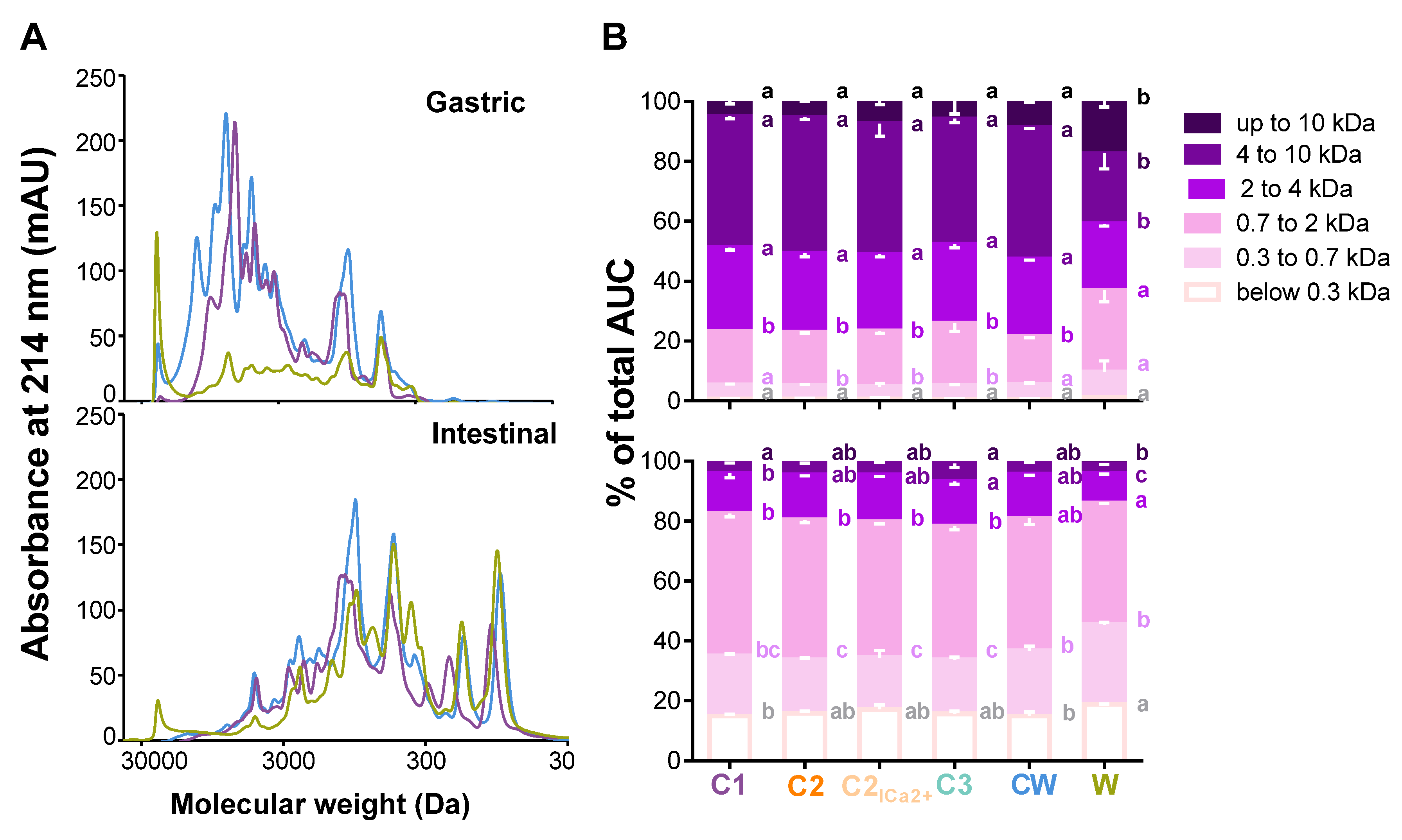
3.2. Comparison of the Gastric and Intestinal Digests of the Dairy Matrices with Size Exclusion Chromatography
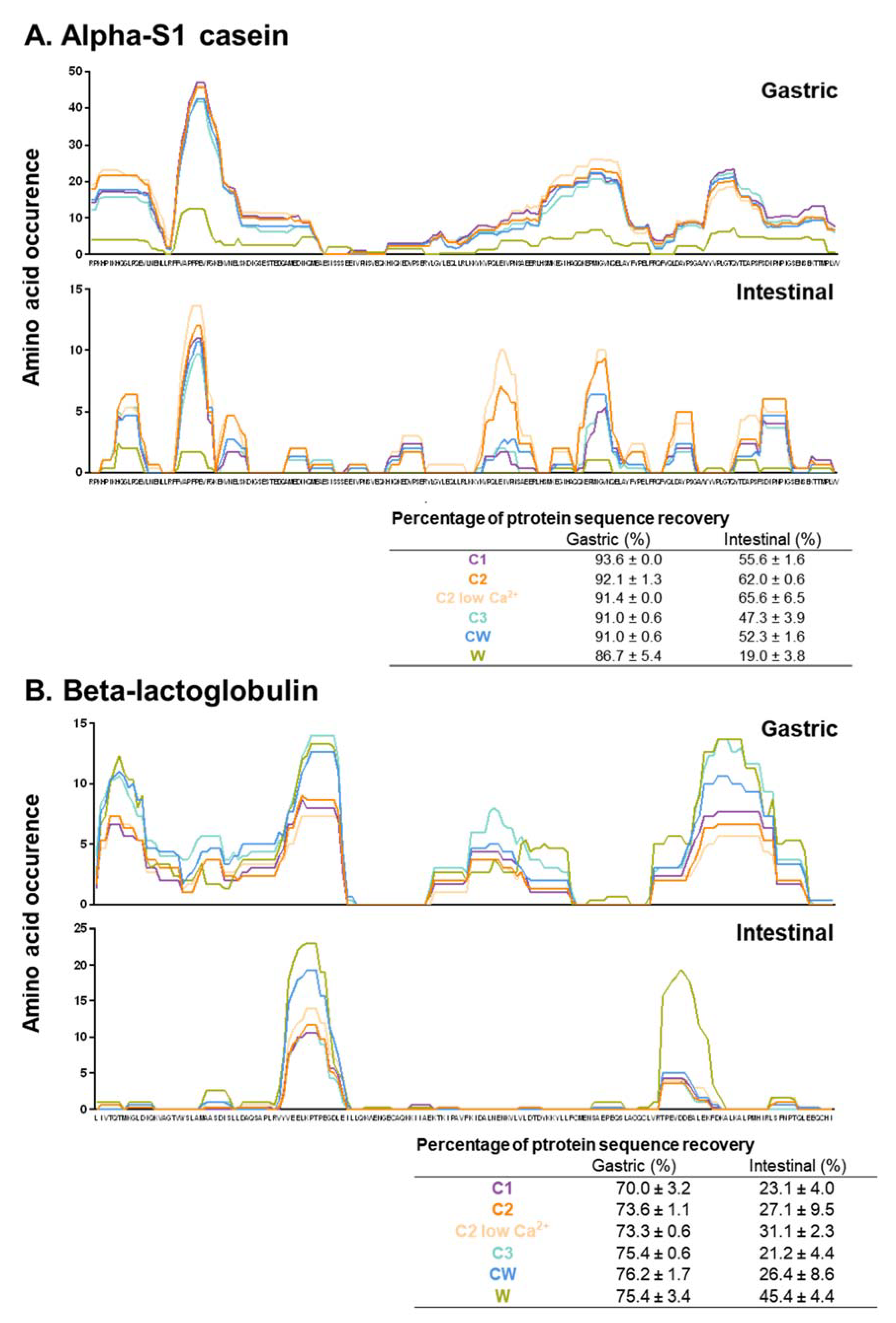
3.3. HPLC-MS/MS Analysis of the Gastric and Intestinal Peptidomes of the Dairy Matrices
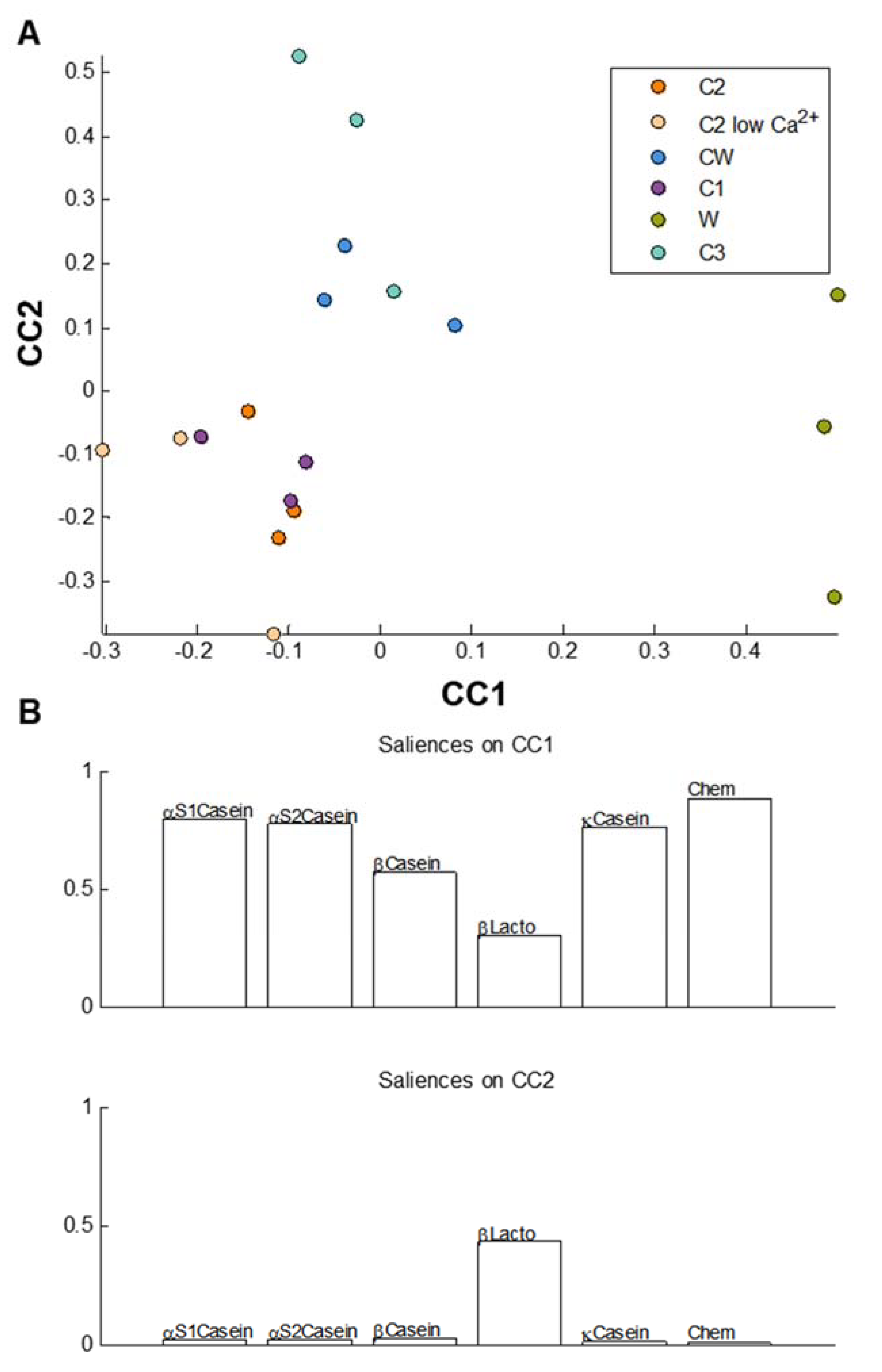
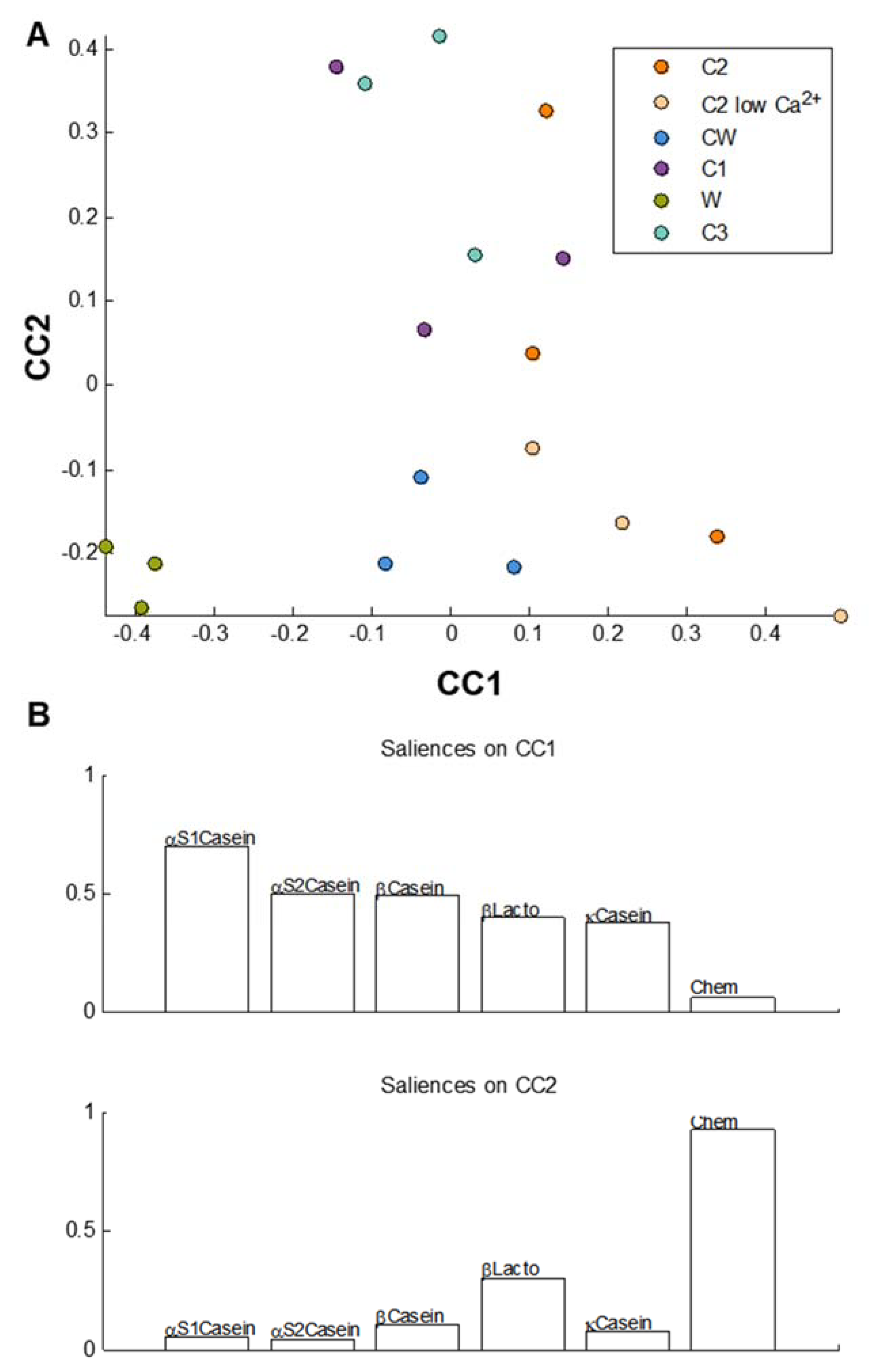
3.4. Statistical Comparisons of the Gastric and the Intestinal Digests of the Dairy Matrices
4. Discussion
4.1. Optimization of the Harmonized Protocol
4.2. Comparisons of the Molecular Weights and Resistance Zones Between the Milk Matrices
4.3. Peptide Identification During Digestion of the Matrices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fox, P.F. Milk proteins as food ingredients. Int. J. Dairy Technol. 2001, 54, 41–55. [Google Scholar] [CrossRef]
- Dalgleish, D.; Corredig, M. The Structure of the Casein Micelle of Milk and Its Changes During Processing. Annu. Rev. Food Sci. Technol. 2012, 3, 449–467. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Bos, C.; Léonil, J.; Airinei, G.; Luengo, C.; Daré, S.; Benamouzig, R.; Fouillet, H.; Fauquant, J.; Tomé, D.; et al. Compared with casein or total milk protein, digestion of milk soluble proteins is too rapid to sustain the anabolic postprandial amino acid requirement. Am. J. Clin. Nutr. 2006, 84, 1070–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahé, S.; Roos, N.; Benamouzig, R.; Davin, L.; Luengo, C.; Gagnon, L.; Gaussergès, N.; Rautureau, J.; Tomé, D. Gastrojejunal kinetics and the digestion of [15N]beta-lactoglobulin and casein in humans: The influence of the nature and quantity of the protein. Am. J. Clin. Nutr. 1996, 63, 546–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.-P.; Maubois, J.-L.; Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Lieshout, G.A.A.; Lambers, T.T.; Bragt, M.C.E.; Hettinga, K.A. How processing may affect milk protein digestion and overall physiological outcomes: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 60, 2422–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coskun, A.E.I.; Sağlam, D.; Venema, P.; Van Der Linden, E.; Scholten, E. Preparation, structure and stability of sodium caseinate and gelatin micro-particles. Food Hydrocoll. 2015, 45, 291–300. [Google Scholar] [CrossRef]
- Dalgleish, D.; Spagnuolo, P.A.; Goff, H.D. A possible structure of the casein micelle based on high-resolution field-emission scanning electron microscopy. Int. Dairy J. 2004, 14, 1025–1031. [Google Scholar] [CrossRef]
- Wang, X.; Ye, A.; Lin, Q.; Han, J.; Singh, H. Gastric digestion of milk protein ingredients: Study using an in vitro dynamic model. J. Dairy Sci. 2018, 101, 6842–6852. [Google Scholar] [CrossRef] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised staticin vitrodigestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Egger, L.; Schlegel, P.; Baumann, C.; Stoffers, H.; Guggisberg, D.; Brügger, C.; Dürr, D.; Stoll, P.; Vergères, G.; Portmann, R. Physiological comparability of the harmonized INFOGEST in vitro digestion method to in vivo pig digestion. Food Res. Int. 2017, 102, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T.; Carriere, F.; Day, L.; Deglaire, A.; Egger, L.; Freitas, D.; Golding, M.; Le Feunteun, S.; Macierzanka, A.; Ménard, O.; et al. Correlation between in vitro and in vivo data on food digestion. What can we predict with static in vitro digestion models? Crit. Rev. Food Sci. Nutr. 2017, 58, 2239–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, L.; Ménard, O.; Baumann, C.; Duerr, D.; Schlegel, P.; Stoll, P.; Vergères, G.; Dupont, D.; Portmann, R. Digestion of milk proteins: Comparing static and dynamic in vitro digestion systems with in vivo data. Food Res. Int. 2019, 118, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Bidlingmeyer, B.A.; Cohen, S.A.; Tarvin, T.L. Rapid analysis of amino acids using pre-column derivatization. J. Chromatogr. B Biomed. Sci. Appl. 1984, 336, 93–104. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nat. Cell Biol. 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Evincent, D.; Elkins, A.; Condina, M.R.; Ezernieks, V.; Rochfort, S. Quantitation and Identification of Intact Major Milk Proteins for High-Throughput LC-ESI-Q-TOF MS Analyses. PLoS ONE 2016, 11, e0163471. [Google Scholar] [CrossRef]
- Cordella, C.B.; Bertrand, D. SAISIR: A new general chemometric toolbox. Trends Anal. Chem. 2014, 54, 75–82. [Google Scholar] [CrossRef]
- Mandalari, G.; Adel-Patient, K.; Barkholt, V.; Baro, C.; Bennett, L.; Bublin, M.; Gaier, S.; Graser, G.; Ladics, G.; Mierzejewska, D.; et al. In vitro digestibility of β-casein and β-lactoglobulin under simulated human gastric and duodenal conditions: A multi-laboratory evaluation. Regul. Toxicol. Pharmacol. 2009, 55, 372–381. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, Y.-H. Whole-Cell Protein Identification Using the Concept of Unique Peptides. Genom. Proteom. Bioinform. 2010, 8, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Ménard, O.; Famelart, M.-H.; Deglaire, A.; Le Gouar, Y.; Guérin, S.; Malbert, C.-H.; Dupont, D. Gastric Emptying and Dynamic In Vitro Digestion of Drinkable Yogurts: Effect of Viscosity and Composition. Nutrients 2018, 10, 1308. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; Gumpertz, M.; Van Kempen, T. Stability of a Pancreatic Enzyme Cocktail during in Vitro Protein Digestibility Assays. J. Food Biochem. 2005, 29, 205–220. [Google Scholar] [CrossRef]
- Stewart, R.J.C.; Morton, H.; Coad, J.; Pedley, K.C. In vitro digestion for assessing micronutrient bioavailability: The importance of digestion duration. Int. J. Food Sci. Nutr. 2018, 70, 71–77. [Google Scholar] [CrossRef]
- De Cicco, M.; Mamone, G.; Di Stasio, L.; Ferranti, P.; Addeo, F.; Picariello, G. Hidden “Digestome”: Current Analytical Approaches Provide Incomplete Peptide Inventories of Food Digests. J. Agric. Food Chem. 2019, 67, 7775–7782. [Google Scholar] [CrossRef]
- Broyard, C.; Gaucheron, F. Modifications of structures and functions of caseins: A scientific and technological challenge. Dairy Sci. Technol. 2015, 95, 831–862. [Google Scholar] [CrossRef]
- Holt, C. Structure and Stability of Bovine Casein Micelles. Adv. Protein Chem. 1992, 43, 63–151. [Google Scholar] [PubMed]
- Tsioulpas, A.; Lewis, M.J.; Grandison, A.S. Effect of Minerals on Casein Micelle Stability of Cows’ Milk. J. Dairy Res. 2007, 74, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.; Brodkorb, A. The casein micelle: Historical aspects, current concepts and significance. Int. Dairy J. 2008, 18, 677–684. [Google Scholar] [CrossRef]
- Boutrou, R.; Gaudichon, C.; Dupont, D.; Jardin, J.; Airinei, G.; Marsset-Baglieri, A.; Benamouzig, R.; Tome, D.; Leonil, J. Sequential release of milk protein-derived bioactive peptides in the jejunum in healthy humans. Am. J. Clin. Nutr. 2013, 97, 1314–1323. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Native Micellar Casein (%) | Whey Proteins (%) | Calcium (%) | Total Nitrogen Content (%) | |
|---|---|---|---|---|
| C1 | 92 | 8 | 2.6 | 83 |
| C2 | 92 | 8 | 2.1 | 82.3 |
| C2 low Ca2+ | 92 | 8 | 1.6 | 85.1 |
| C3 | 92 | 8 | 2.6 | 80.1 |
| CW | 80 | 20 | 2.2 | 81 |
| W | 0 | 100 | 0.3 | 81.5 |
| Milk Matrices | Phase | MW > 10 kDa | 4 < MW < 10 kDa | 2 < MW < 4 kDa | 0.7 < MW < 2 kDa | 0.3 < MW < 0.7 kDa | MW < 3 kDa | Peptide Concentration (mg/mL−1) | Calcium Concentration (mg/mL−1) | Total Free AA Concentration (mg/100g of Product) |
|---|---|---|---|---|---|---|---|---|---|---|
| C1 | Gastric | 4.9 ± 0.7 | 43.9 ± 0.7 | 27.9 ± 0.8 | 18.0 ± 0.0 | 4.7 ± 0.4 | 0.6 ± 0.2 | 0.24 | 0.7 | <LD |
| C2 | 5.7 ± 0.5 | 45.4 ± 0.7 | 26.3 ± 1.1 | 18.0 ± 0.3 | 4.3 ± 0.6 | 0.7 ± 0.2 | 0.33 ± 0.04 | 0.6 | <LD | |
| C2 low Ca2+ | 7.6 ± 1.3 | 43.6 ± 4.2 | 25.6 ± 0.9 | 18.5 ± 0.9 | 4.3 ± 1.0 | 0.6 ± 0.5 | 0.16 | 0.4 | <LD | |
| C3 | 5.8 ± 4.1 | 41.9 ± 1.3 | 26.4 ± 1.1 | 20.9 ± 2.7 | 4.5 ± 0.4 | 0.5 ± 0.1 | 0.26 | 0.8 | <LD | |
| CW | 8.8 ± 0.4 | 43.9 ± 0.3 | 25.9 ± 0.4 | 16.0 ± 0.5 | 5.1 ± 0.4 | 0.5 ± 0.2 | 0.28 ± 0.01 | 0.5 | <LD | |
| W | 17.9 ± 2.2 | 23.5 ± 5.1 | 22.1 ± 0.6 | 27.3 ± 3.8 | 8.5 ± 3.6 | 1.2 ± 0.0 | 0.16 | 0.1 | <LD | |
| C1 | Intestinal | 0.6 ± 0.1 | 4.0 ± 0.5 | 13.4 ± 1.5 | 47.5 ± 1.1 | 20.0 ± 0.8 | 14.9 ± 0.6 | 0.33 | 0.5 | 171.2 ± 32.7 |
| C2 | 0.7 ± 0.1 | 4.3 ± 0.3 | 15.0 ± 0.3 | 46.7 ± 1.0 | 17.8 ± 0.6 | 15.9 ± 0.6 | 0.41 ± 0.02 | 0.4 | 189.0 ± 55.3 | |
| C2 low Ca2+ | 0.7 ± 0.1 | 4.5 ± 0.3 | 15.6 ± 0.6 | 45.4 ± 0.7 | 17.2 ± 2.4 | 17.2 ± 1.5 | 0.36 | 0.3 | 233.3 ± 4.5 | |
| C3 | 0.9 ± 0.2 | 6.0 ± 1.4 | 14.9 ± 0.9 | 44.6 ± 1.2 | 18.0 ± 0.9 | 15.7 ± 0.8 | 0.32 | 0.5 | 148.0 ± 27.5 | |
| CW | 0.7 ± 0.3 | 4.2 ± 0.2 | 14.6 ± 0.3 | 44.3 ± 2.1 | 21.7 ± 1.5 | 15.0 ± 1.5 | 0.34 ± 0.01 | 0.3 | 253.3 ± 15.0 | |
| W | 1.1 ± 0.1 | 3.3 ± 0.2 | 9.7 ± 0.2 | 40.6 ± 0.1 | 26.6 ± 0.7 | 18.8 ± 0.0 | 0.36 | 0.1 | 277.3 ± 4.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atallah, N.; Deracinois, B.; Boulier, A.; Baniel, A.; Jouan-Rimbaud Bouveresse, D.; Ravallec, R.; Flahaut, C.; Cudennec, B. In Vitro Assessment of the Impact of Industrial Processes on the Gastrointestinal Digestion of Milk Protein Matrices Using the INFOGEST Protocol. Foods 2020, 9, 1580. https://doi.org/10.3390/foods9111580
Atallah N, Deracinois B, Boulier A, Baniel A, Jouan-Rimbaud Bouveresse D, Ravallec R, Flahaut C, Cudennec B. In Vitro Assessment of the Impact of Industrial Processes on the Gastrointestinal Digestion of Milk Protein Matrices Using the INFOGEST Protocol. Foods. 2020; 9(11):1580. https://doi.org/10.3390/foods9111580
Chicago/Turabian StyleAtallah, Nathalie, Barbara Deracinois, Audrey Boulier, Alain Baniel, Delphine Jouan-Rimbaud Bouveresse, Rozenn Ravallec, Christophe Flahaut, and Benoit Cudennec. 2020. "In Vitro Assessment of the Impact of Industrial Processes on the Gastrointestinal Digestion of Milk Protein Matrices Using the INFOGEST Protocol" Foods 9, no. 11: 1580. https://doi.org/10.3390/foods9111580