Characterization of the Spoilage Microbiota of Hake Fillets Packaged Under a Modified Atmosphere (MAP) Rich in CO2 (50% CO2/50% N2) and Stored at Different Temperatures

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Raw Material, Modified Atmosphere Packaging, Storage Conditions, and Experimental Design
2.2. Sensory and Biochemical Analysis
2.3. Standard Microbiological Analysis
2.4. Bacterial DNA Extraction from Hake Fillets
2.5. qPCR Assays
2.6. High-Throughput 16S rRNA Gene Sequencing
2.7. Statistical Analysis
3. Results
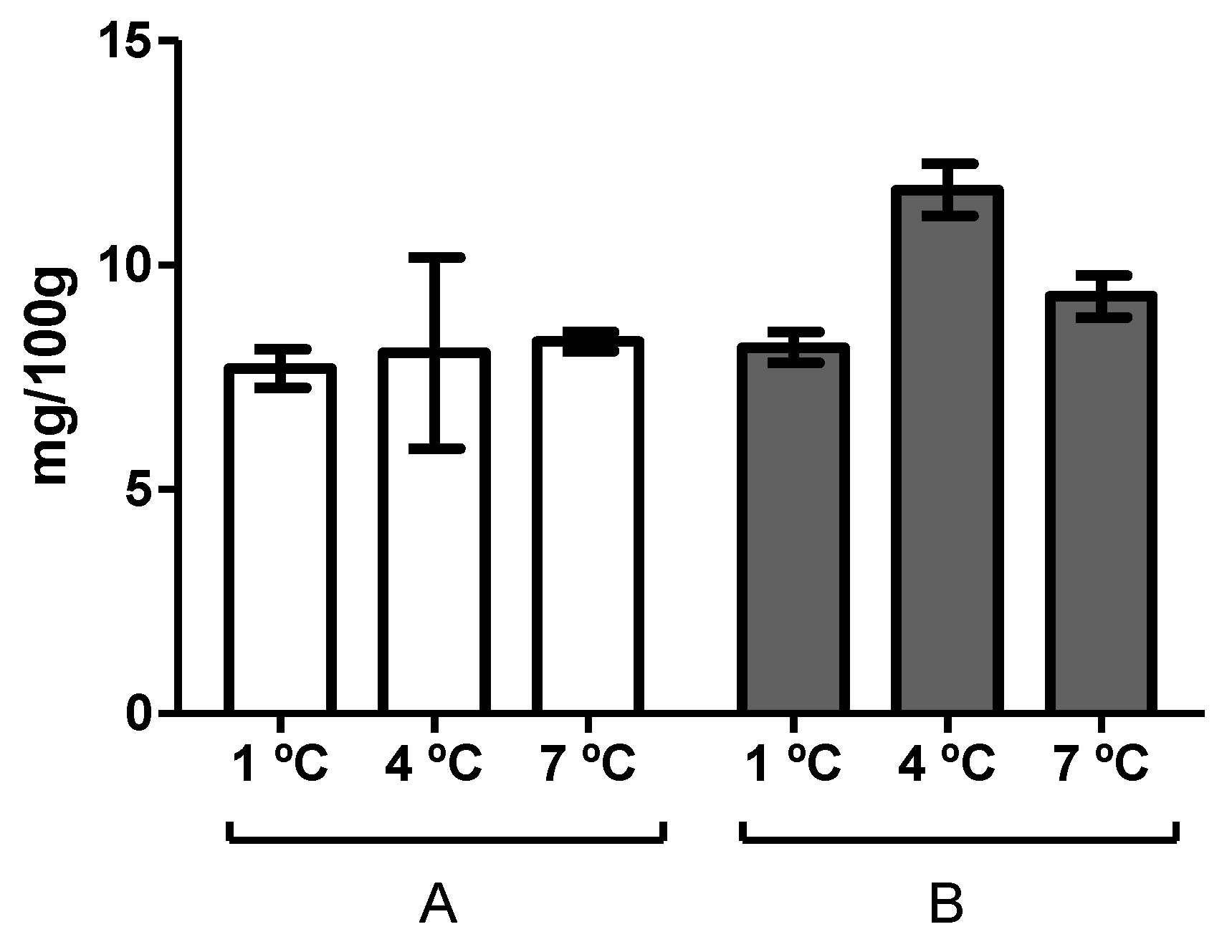
3.1. Determination of the Time of Spoilage through Sensory and Biochemical Analysis
3.2. qPCR Analysis
3.3. High-Throughput 16S rRNA Gene Sequencing
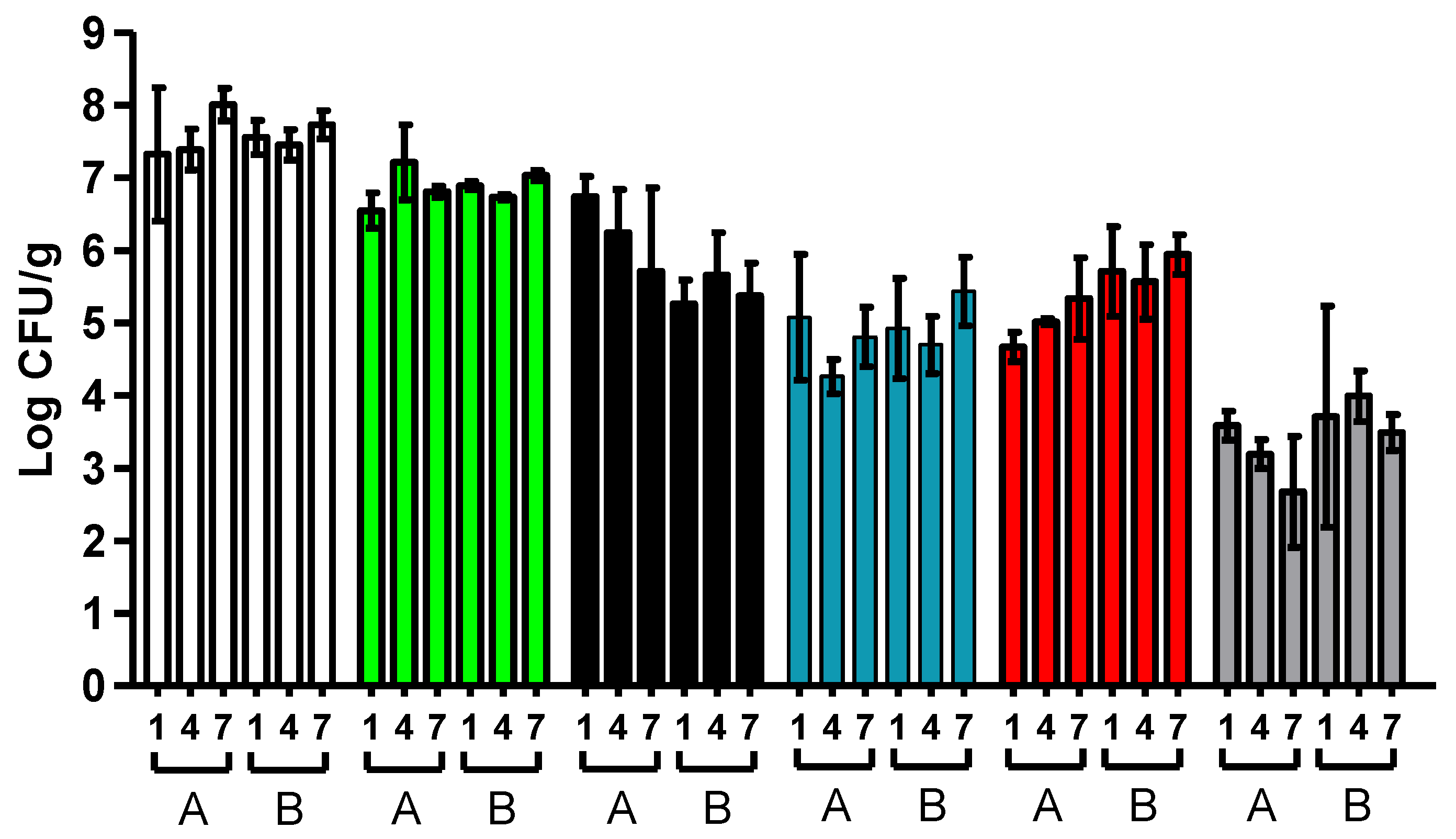
3.4. Standard Microbiological Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alheit, J.; Pitcher, T.J. Hake: Biology, Fisheries and Markets; Fish and Fisheries Series No. 15; Chapman & Hall: London, UK, 1995. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2016. Available online: http://www.fao.org/3/a-i5555e.pdf (accessed on 11 August 2019).
- Baixas-Nogueras, S.; Bover-Cid, S.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Amino Acid-Decarboxylase activity in bacteria associated with Mediterranean hake spoilage. Eur. Food Res. Technol. 2003, 217, 164–167. [Google Scholar] [CrossRef]
- MAPAMA. El Mercado de la Merluza en España. 2016. Available online: https://www.mapa.gob.es/es/pesca/temas/mercados-economia-pesquera/informemerluzaabril2016_tcm30-291641.pdf (accessed on 11 August 2019).
- Dalgaard, P. Qualitative and quantitative characterization of spoilage bacteria from packed fish. Int. J. Food Microbiol. 1995, 26, 319–333. [Google Scholar] [CrossRef]
- Koutsoumanis, K. Predictive modeling of the shelf life of fish under nonisothermal conditions. Appl. Environ. Microbiol. 2001, 67, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I. Fluorescently labelled, rRNA-Targeted oligonucleotide probes in the study of microbial ecology. Mol. Ecol. 1995, 4, 543–554. [Google Scholar] [CrossRef]
- Di Bella, J.M.; Bao, Y.; Gloor, G.B.; Burton, J.P.; Reid, G. High throughput sequencing methods and analysis for microbiome research. J. Microbiol. Methods 2013, 95, 401–414. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Burke, C.M.; Bolch, C.C.J.; Stanley, R. Seafood spoilage microbiota and associated volatile organic compounds at different storage temperatures and packaging conditions. Int. J. Food Microbiol. 2018, 280, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Parlapani, F.F.; Michailidou, S.; Anagnostopoulos, D.A.; Sakellariou, A.K.; Pasentsis, K.; Psomopoulos, F.; Argiriou, A.; Haroutounian, S.A.; Boziaris, I.S. Microbial spoilage investigation of thawed common cuttlefish (Sepia officinalis) stored at 2 °C using next generation sequencing and volatilome analysis. Food Microbiol. 2018, 76, 518–525. [Google Scholar] [CrossRef]
- Jia, S.; Huang, Z.; Lei, Y.; Zhang, L.; Li, Y.; Luo, Y. Application of Illumina-MiSeq high throughput sequencing and culture-dependent techniques for the identification of microbiota of silver carp (Hypophthalmichthys molitrix) treated by tea polyphenols. Food Microbiol. 2018, 76, 52–61. [Google Scholar] [CrossRef]
- Zotta, T.; Parente, E.; Ianniello, R.G.; De Filippis, F.; Ricciardi, A. Dynamics of bacterial communities and interaction networks in thawed fish fillets during chilled storage in air. Int. J. Food Microbiol. 2019, 293, 102–113. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Michailidou, S.; Pasentsis, K.; Argiriou, A.; Krey, G.; Boziaris, I.S. A Meta-Barcoding approach to assess and compare the storage Temperature-Dependent bacterial diversity of Gilt-Head sea bream (Sparus aurata) originating from fish farms from two geographically distinct areas of Greece. Int. J. Food Microbiol. 2018, 278, 36–43. [Google Scholar] [CrossRef]
- Kuuliala, L.; Al Hage, Y.; Ioannidis, A.-G.; Sader, M.; Kerckhof, F.-M.; Vanderroost, M.; Boon, N.; De Baets, B.; De Meulenaer, B.; Ragaert, P.; et al. Microbiological, chemical and sensory spoilage analysis of raw Atlantic cod (Gadus morhua) stored under modified atmospheres. Food Microbiol. 2018, 70, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, D. High-Throughput sequencing and metagenomics: Moving forward in the Culture-Independent analysis of food microbial ecology. Appl. Environ. Microbiol. 2013, 79, 3148–3155. [Google Scholar] [CrossRef]
- Rodrigues, P.A.; Ferrari, R.G.; Conte-Junior, C.A. Application of molecular tools to elucidate the microbiota of seafood. J. Appl. Microbiol. 2018, 124, 1347–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO 8586: 2012. Sensory Analysis—General Guidelines for the Selection, Training and Monitoring of Selected Assessors and Expert Sensory Assessors; International Organization for Standardization: Geneva, Switzerland, 2012. [Google Scholar]
- Seafish. Sensory Assessment Scoresheets for Fish and Shellfish-Torry & QIM. 2010. Available online: https://www.seafish.org/media/Publications/sensory_assessment_scoresheets_14_5_10.pdf (accessed on 11 August 2019).
- Woyewoda, A.D.; Shaw, S.J.; Ke, P.J.; Burns, B.G. Recommended Laboratory Methods for Assessment of Fish Quality; Canadian Technical Report of Fisheries and Aquatic Sciences No. 1448; Fisheries and Oceans: Ottawo, ON, Canada, 1996. [Google Scholar]
- Antunes-Rohling, A.; Artaiz, A.; Calero, S.; Halaihel, N.; Guillén, S.; Raso, J.; Álvarez, I.; Cebrián, G. Modelling microbial growth in Modified-Atmosphere-Packed hake (Merluccius merluccius) fillets stored at different temperatures. Food Res. Int. 2019, 122, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.; Danilowicz, B.S.; Meijer, W.G. The diversity of bacterial communities associated with Atlantic Cod Gadus morhua. Microb. Ecol. 2008, 55, 425–434. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation Sequencing-Based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of High-Throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-Resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Baixas-Nogueras, S.; Bover-Cid, S.; Veciana-Nogués, T.; Nunes, M.L.; Vidal-Carou, M.C. Development of a quality index method to evaluate freshness in Mediterranean hake (Merluccius merluccius). J. Food Sci. 2003, 68, 1067–1071. [Google Scholar] [CrossRef]
- Gillespie, N.C.; MacRae, I.C. The bacterial flora of some Queensland fish and its ability to cause spoilage. J. Appl. Bacteriol. 1975, 39, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Shewan, J.M.; Murray, C.K. The microbial spoilage of fish with special reference to the role of psychrophiles. In Cold Tolerant Microbes in Spoilage and the Environment; Russell, A.D., Fuller, R., Eds.; Academic Press: London, UK, 1979; pp. 36–117. [Google Scholar]
- Gram, L.; Dalgaard, P. Fish spoilage Bacteria-Problems and solutions. Curr. Opin. Biotechnol. 2002, 13, 262–266. [Google Scholar] [CrossRef]
- Gram, L.; Ravn, L.; Rasch, M.; Bruhn, J.B.; Christensen, A.B.; Givskov, M. Food spoilage—Interactions between food spoilage bacteria. Int. J. Food Microbiol. 2002, 78, 79–97. [Google Scholar] [CrossRef]
- Hilton, S.K.; Castro-Nallar, E.; Pérez-Losada, M.; Toma, I.; McCaffrey, T.A.; Hoffman, E.P.; Siegel, M.O.; Simon, G.L.; Evan Johnson, W.; Crandall, K.A. Metataxonomic and metagenomic approaches vs. Culture-Based techniques for clinical pathology. Front. Microbiol. 2016, 7, 484. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hammes, F.; De Roy, K.; Verstraete, W.; Boon, N. Past, present and future applications of flow cytometry in aquatic microbiology. Trends Biotechnol. 2010, 28, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Didelot, X.; Bowden, R.; Wilson, D.J.; Peto, T.E.; Crook, D.W. Transforming clinical microbiology with bacterial genome sequencing. Nat. Rev. Genet. 2012, 13, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLoughlin, K.S. Microarrays for pathogen detection and analysis. Brief. Funct. Genom. 2011, 10, 342–353. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2009, 11, 31–46. [Google Scholar] [CrossRef]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Mumford, R.; Jackeviciene, E.; Navalinskiene, M.; Samuitiene, M.; Boonham, N. Next-Generation sequencing and metagenomic analysis: A universal diagnostic tool in plant virology. Mol. Plant Pathol. 2009, 10, 537–545. [Google Scholar] [CrossRef]
- Dunne, W.; Westblade, L.; Ford, B. Next-Generation and Whole-Genome sequencing in the diagnostic clinical microbiology laboratory. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1719–1726. [Google Scholar] [CrossRef]
- Boziaris, I.S.; Parlapani, F.F. Chapter 3: Specific Spoilage Organisms (SSO) in Fish. In Microbiological Quality of Food: Foodborne Spoilers; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Elsevier: Amsterdam, The Netherlands; Woodhead Publishing: Cambridge, UK, 2016; pp. 61–98. [Google Scholar]
- Macé, S.; Cornet, J.; Chevalier, F.; Cardinal, M.; Pilet, M.F.; Dousset, X.; Joffraud, J.J. Characterization of the spoilage microbiota in raw salmon (Salmo salar) steaks stored under vacuum or modified atmosphere packaging combining conventional methods and PCR–TTGE. Food Microbiol. 2012, 30, 164–172. [Google Scholar] [CrossRef]
- Reynisson, E.; Lauzon, H.L.; Magnússon, H.; Jónsdóttir, R.; Ólafsdóttir, G.; Marteinsson, V.; Hreggvidsson, G. Bacterial composition and succession during storage of North-Atlantic cod (Gadus morhua) at superchilled temperatures. BMC Microbiol. 2009, 9, 250. [Google Scholar] [CrossRef] [PubMed]
- Dalgaard, P.; Mejlholm, O.; Christiansen, T.J.; Huss, H.H. Importance of Photobacterium phosphoreum in relation to spoilage of modified Atmosphere-Packed fish products. Lett. Appl. Microbiol. 1997, 24, 373–378. [Google Scholar] [CrossRef]
- Rudi, K.; Maugesten, T.; Hannevik, S.E.; Nissen, H. Explorative multivariate analyses of 16S rRNA gene data from microbial communities in Modified-Atmosphere-Packed salmon and coalfish. Appl. Environ. Microbiol. 2004, 70, 5010–5018. [Google Scholar] [CrossRef] [PubMed]
- Hovda, M.B.; Lunestad, B.T.; Sivertsvik, M.; Rosnes, J.T. Characterisation of the bacterial flora of modified atmosphere packaged farmed Atlantic cod (Gadus morhua) by PCR-DGGE of conserved 16S rRNA gene regions). Int. J. Food Microbiol. 2007, 117, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Jones, Y.; Kitts, D.D. Seafood Safety, Processing and Biotechnology; Technomic Publishing Co. Inc.: Lancaster, PA, USA, 1997. [Google Scholar]
- Bowman, J.P. The genus Psychrobacter. In The Prokaryotes—A Handbook on the Biology of Bacteria; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 920–930. [Google Scholar]
- Bjorkevoll, I.; Olsen, R.L.; Skjerdal, O.T. Origin and spoilage potential of the microbiota dominating genus Psychrobacter in sterile rehydrated Salt-Cured and dried salt-cured cod (Gadus morhua). Int. J. Food Microbiol. 2003, 84, 175–187. [Google Scholar] [CrossRef]
- Broekaert, K.; Heyndrickx, M.; Herman, L.; Devlieghere, F.; Vlaemynck, G. Seafood quality analysis: Molecular identification of dominant microbiota after ice storage on several growth media. Food Microbiol. 2011, 28, 1162–1169. [Google Scholar] [CrossRef]
- Romero, J.; Gonzalez, N.; Espejo, R.T. Marine Pseudoalteromonas sp. composes most of the bacterial population developed in oysters (Tiostrea chilensis) spoiled during storage. J. Food Sci. 2002, 67, 2300–2303. [Google Scholar] [CrossRef]
- Mejlholm, O.; Boknaes, N.; Dalgaard, P. Shelf life and safety aspects of chilled cooked and peeled shrimps (Pandalus borealis) in modified atmosphere packaging. J. Appl. Microbiol. 2005, 99, 66–76. [Google Scholar] [CrossRef]
- Prapaiwong, N.; Wallace, R.K.; Arias, C.R. Bacterial loads and microbial composition in high pressure treated oysters during storage. Int. J. Food Microbiol. 2009, 131, 145–150. [Google Scholar] [CrossRef]
- Rodriguez-Calleja, J.M.; Patterson, M.F.; Garcia-Lopez, I.; Santos, J.A.; Otero, A.; Garcia-Lopez, M.L. Incidence, radioresistance, and behavior of Psychrobacter spp. in rabbit meat. J. Food Prot. 2005, 68, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Gennari, M.; Parini, M.; Volpon, D.; Serio, M. Isolation and characterisation by conventional methods and genetic transformation of Psychrobacter and Acinetobacter from fresh and spoiled meat, milk and cheese. Int. J. Food Microbiol. 1992, 15, 61–75. [Google Scholar] [CrossRef]
- Broekaert, K.; Noseda, B.; Heyndrickx, M.; Vlaemynck, G.; Devlieghere, F. Volatile compounds associated with Psychrobacter spp. and Pseudoalteromonas spp., the dominant microbiota of brown shrimp (Crangon crangon) during aerobic storage. Int. J. Food Microbiol. 2013, 166, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Leisner, J.J.; Laursen, B.G.; Prévost, H.; Drider, D.; Dalgaard, P. Carnobacterium: Positive and negative effects in the environment and in foods. FEMS Microbiol. Rev. 2007, 31, 592–613. [Google Scholar] [CrossRef] [PubMed]
- Joffraud, J.J. Effect of bacterial interactions on the spoilage of cold-smoked salmon. Int J. Food Microbiol. 2006, 112, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Urakawa, H.; Kita-Tsukamoto, K.; Steven, S.E.; Ohwada, K.; Colwell, R.R. A proposal to transfer Vibrio marinus (Russell 1891) to a new genus Moritella gen. nov. as Moritella marina comb. nov. FEMS Microbiol. Lett. 1998, 165, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Benediktsdoóttir, E.; Heiðarsdoóttir, K.J. Growth and lysis of the fish pathogen Moritella viscosa. Lett. Appl. Microbiol. 2007, 45, 115–120. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Quality Description | Scoring Description | Points |
|---|---|---|
| Muscle firmness | Very Firm (very defined myotomes) | 0 |
| Firm (defined myotomes) | 1 | |
| Friable (moderately defined myotomes) | 2 | |
| Very friable (slightly defined myotomes) | 3 | |
| Smell | Fresh, marine, seaweed | 0 |
| Neutral, fruity | 1 | |
| Slightly sour, metallic or as preserved in the refrigerator | 2 | |
| Strong sour smelling, metallic or as preserved in refrigerator | 3 | |
| Dryness (in the cutting surface) | Low | 0 |
| Moderate | 1 | |
| High | 2 | |
| Very high yellowish, milky areas | 3 | |
| Presence of marks or bruises | None | 0 |
| Some | 1 | |
| Many | 2 | |
| Skin (Ventral Zone) | Metallic and shiny, gray tones | 0 |
| Slightly off-white | 1 | |
| Very White, brightless | 2 | |
| Elasticity of the skin (pressing with several fingers and stretching outwards) | No fingerprint after pressing | 0 |
| With fingerprint after pressing | 1 | |
| QIM SCORE | 0–14 | |
| Microbial Group | Agar | Temp | Time | Atmosphere | Plating |
|---|---|---|---|---|---|
| Aerobic Psychrotrophes | LH Agar 1 | 7 °C | 10 d | Aerobic | Spread |
| Anaerobic Psychrotrophes | LH Agar | 7 °C | 10–12 d | Anaerobic | Spread |
| Pseudomonas | GSP Agar 2 | 25 °C | 24–48 h | Aerobic | Spread |
| Shewanella | Iron Agar 3 | 25 °C | 3–4 d | Aerobic | Spread |
| Lactic Acid Bacteria | Elliker Agar 4 | 25 °C | 24–48 h | Anaerobic5 | Pour |
| Microbial Group | Forward | Reverse | Source |
|---|---|---|---|
| Photobacterium | TACTGTTGAAGTGGCGAT | TCTGCTGGGCTTTCTAAT | This work |
| Pseudomonas | AAGCTAGAGTATGGTAGAG | CACCTCAGTGTCAGTAT | This work |
| Shewanella | GTAGGGAGGAAAGGTAATA | CTTTACGCCCAGTAATTC | This work |
| Temperature (°C) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 4 | 7 | ||||||||
| Microorganism | Sample/Replicate | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 |
| Photobacterium spp. | Cq | 22.53 | 22.36 | 22.12 | 23.08 | 22.86 | 23.23 | 22.27 | 21.90 | 21.97 |
| S.D. | 0.597 | 0.487 | 0.327 | 0.433 | 0.333 | 0.321 | 0.342 | 0.299 | 0.552 | |
| Shewanella spp. | Cq | 28.46 | 30.58 | 27.21 | 29.14 | 29.75 | 28.69 | 26.51 | 28.14 | 27.95 |
| S.D. | 0.575 | 0.092 | 1.068 | 0.932 | 1.036 | 0.485 | 0.687 | 0.538 | 1.981 | |
| Pseudomonas spp. | Cq | 33.90 | 34.16 | 29.19 | 31.88 | 32.97 | 31.06 | 29.22 | 31.68 | 31.69 |
| S.D. | 0.233 | 0.010 | 0.766 | 0.555 | 1.503 | 1.615 | 1.873 | 2.140 | 2.071 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antunes-Rohling, A.; Calero, S.; Halaihel, N.; Marquina, P.; Raso, J.; Calanche, J.; Beltrán, J.A.; Álvarez, I.; Cebrián, G. Characterization of the Spoilage Microbiota of Hake Fillets Packaged Under a Modified Atmosphere (MAP) Rich in CO2 (50% CO2/50% N2) and Stored at Different Temperatures. Foods 2019, 8, 489. https://doi.org/10.3390/foods8100489
Antunes-Rohling A, Calero S, Halaihel N, Marquina P, Raso J, Calanche J, Beltrán JA, Álvarez I, Cebrián G. Characterization of the Spoilage Microbiota of Hake Fillets Packaged Under a Modified Atmosphere (MAP) Rich in CO2 (50% CO2/50% N2) and Stored at Different Temperatures. Foods. 2019; 8(10):489. https://doi.org/10.3390/foods8100489
Chicago/Turabian StyleAntunes-Rohling, Adriana, Silvia Calero, Nabil Halaihel, Pedro Marquina, Javier Raso, Juan Calanche, José Antonio Beltrán, Ignacio Álvarez, and Guillermo Cebrián. 2019. "Characterization of the Spoilage Microbiota of Hake Fillets Packaged Under a Modified Atmosphere (MAP) Rich in CO2 (50% CO2/50% N2) and Stored at Different Temperatures" Foods 8, no. 10: 489. https://doi.org/10.3390/foods8100489