Exploring the Use of a Modified High-Temperature, Short-Time Continuous Heat Exchanger with Extended Holding Time (HTST-EHT) for Thermal Inactivation of Trypsin Following Selective Enzymatic Hydrolysis of the β-Lactoglobulin Fraction in Whey Protein Isolate

Abstract
:1. Introduction
2. Material and Methods
2.1. Targeted Enzymatic Hydrolysis
2.2. Degree of Hydrolysis (%DH)
2.3. Termination of Enzymatic Hydrolysis
2.3.1. Acid Inactivation and Determination of Residual Enzymatic Activity Effect
2.3.2. Water Bath Heating
2.3.3. Pilot-Scale Heat Exchanger Heating
2.4. Protein Profile Analysis
2.4.1. High-Performance Liquid Chromatography (HPLC)
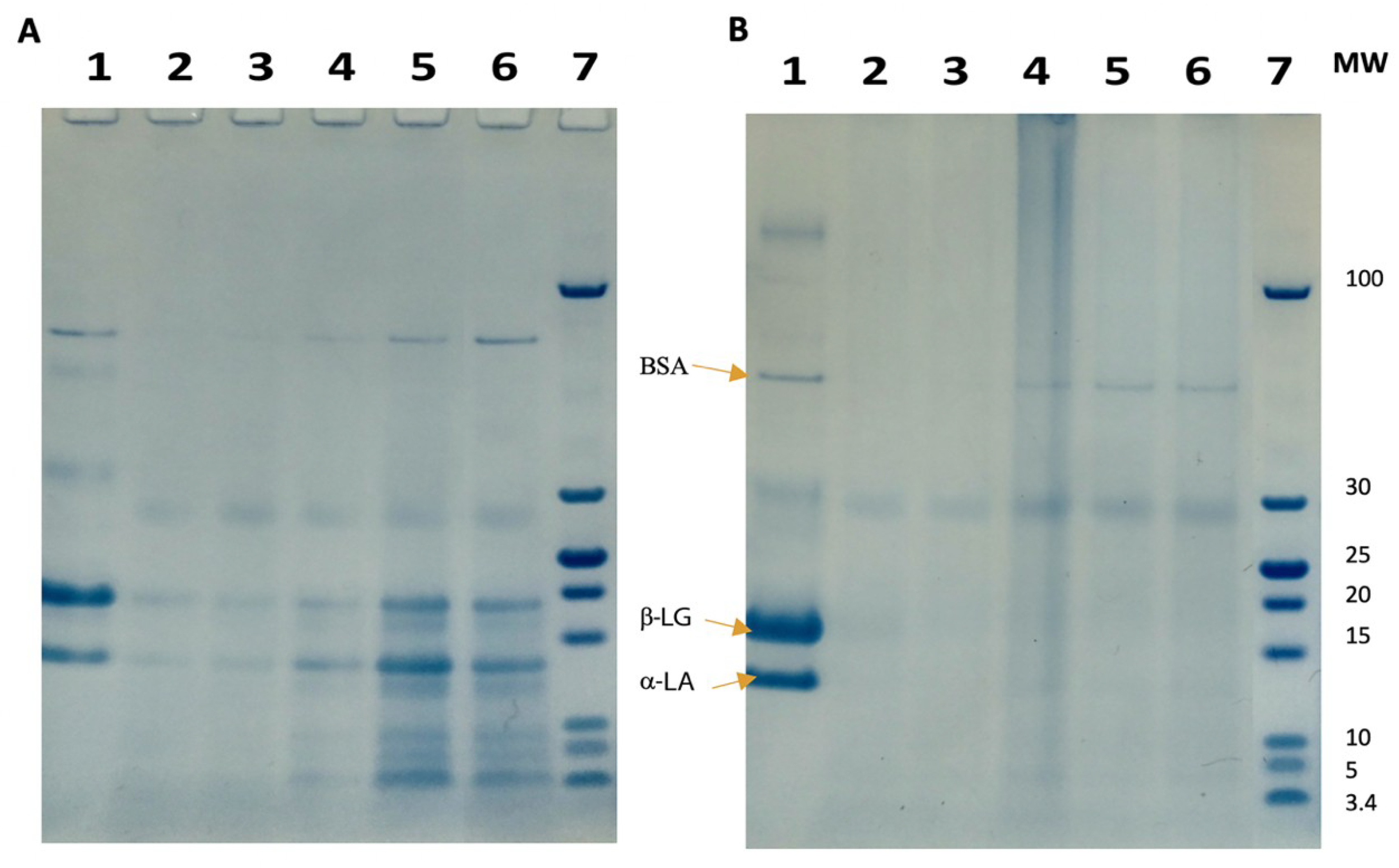
2.4.2. Electrophoresis
3. Results and Discussion
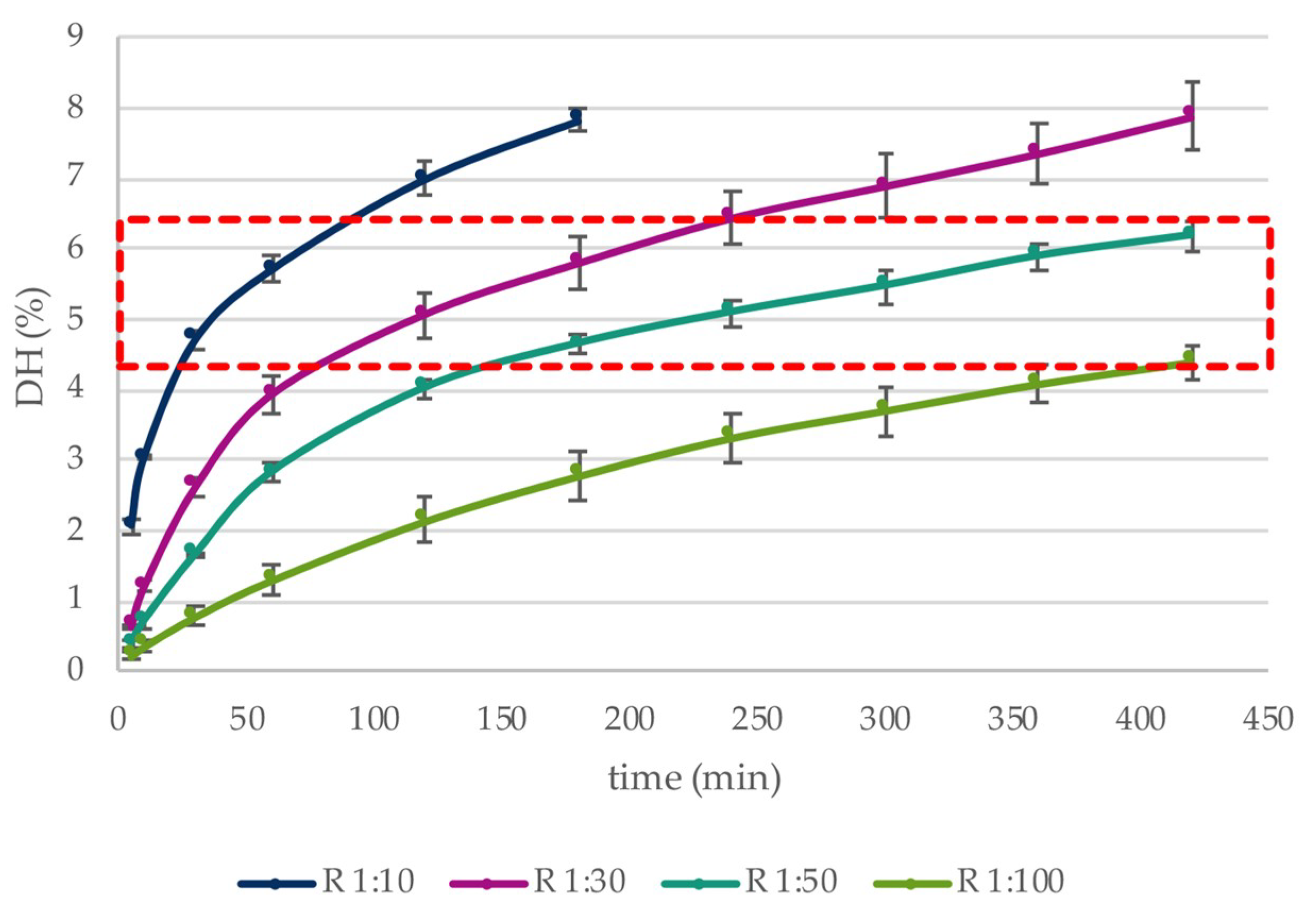
3.1. Enzymatic Hydrolysis
3.2. Inactivation Assays
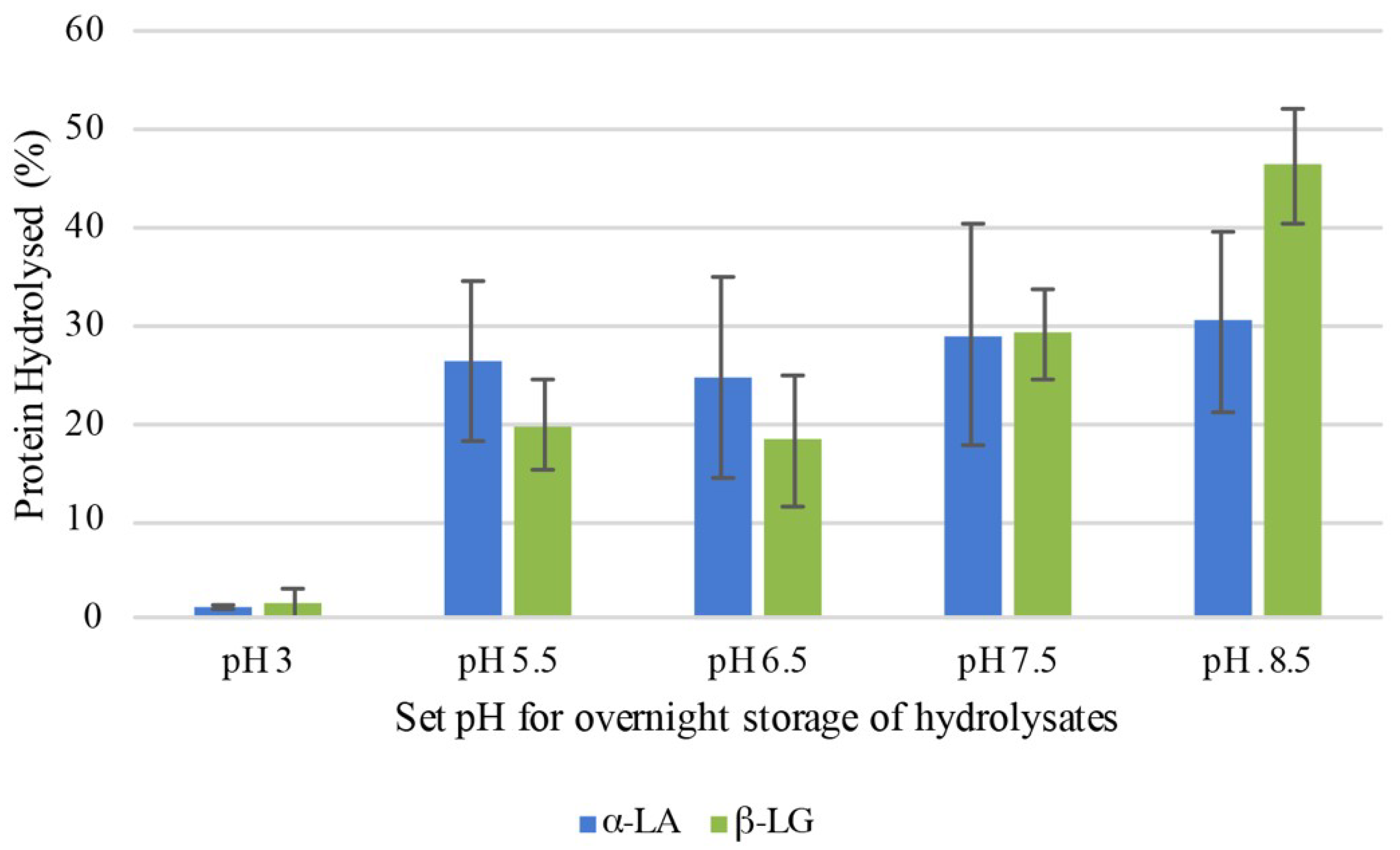
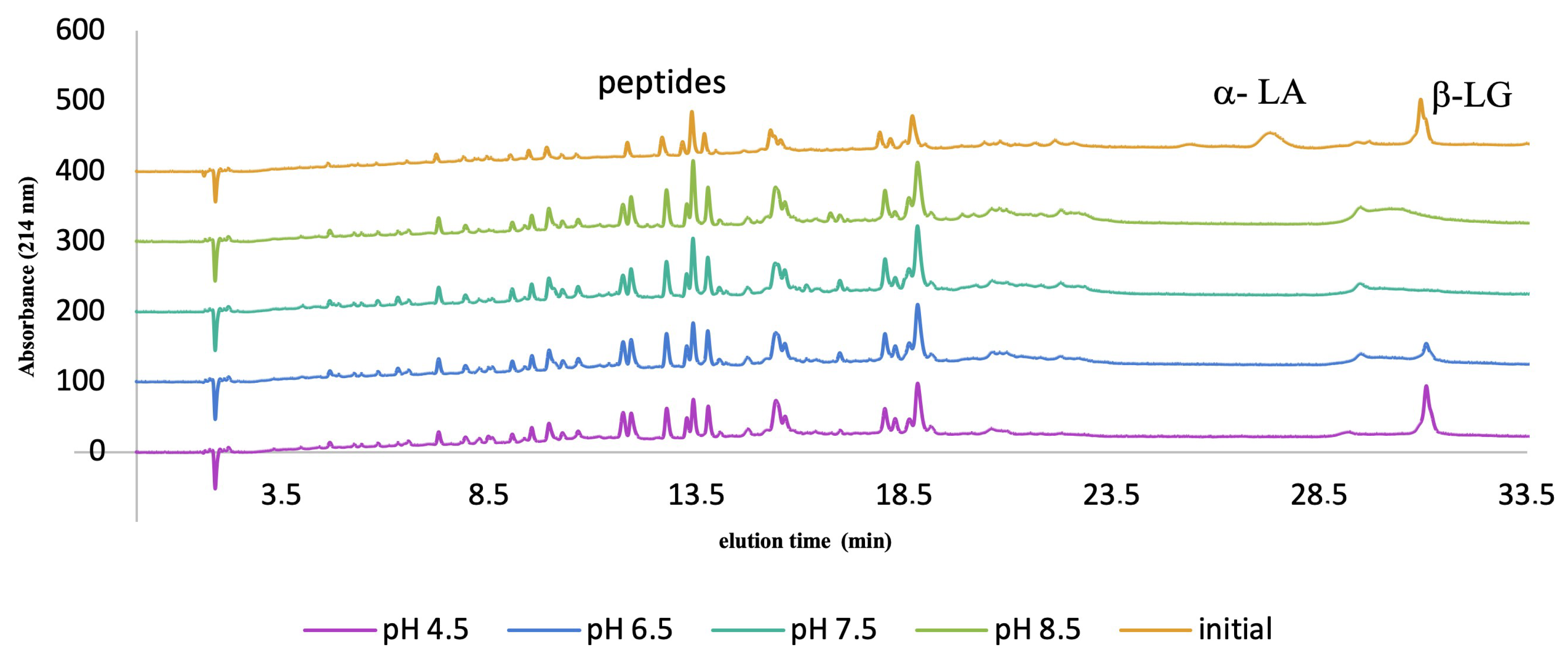
3.2.1. Acid Inactivation
3.2.2. Heat Inactivation: Water Bath
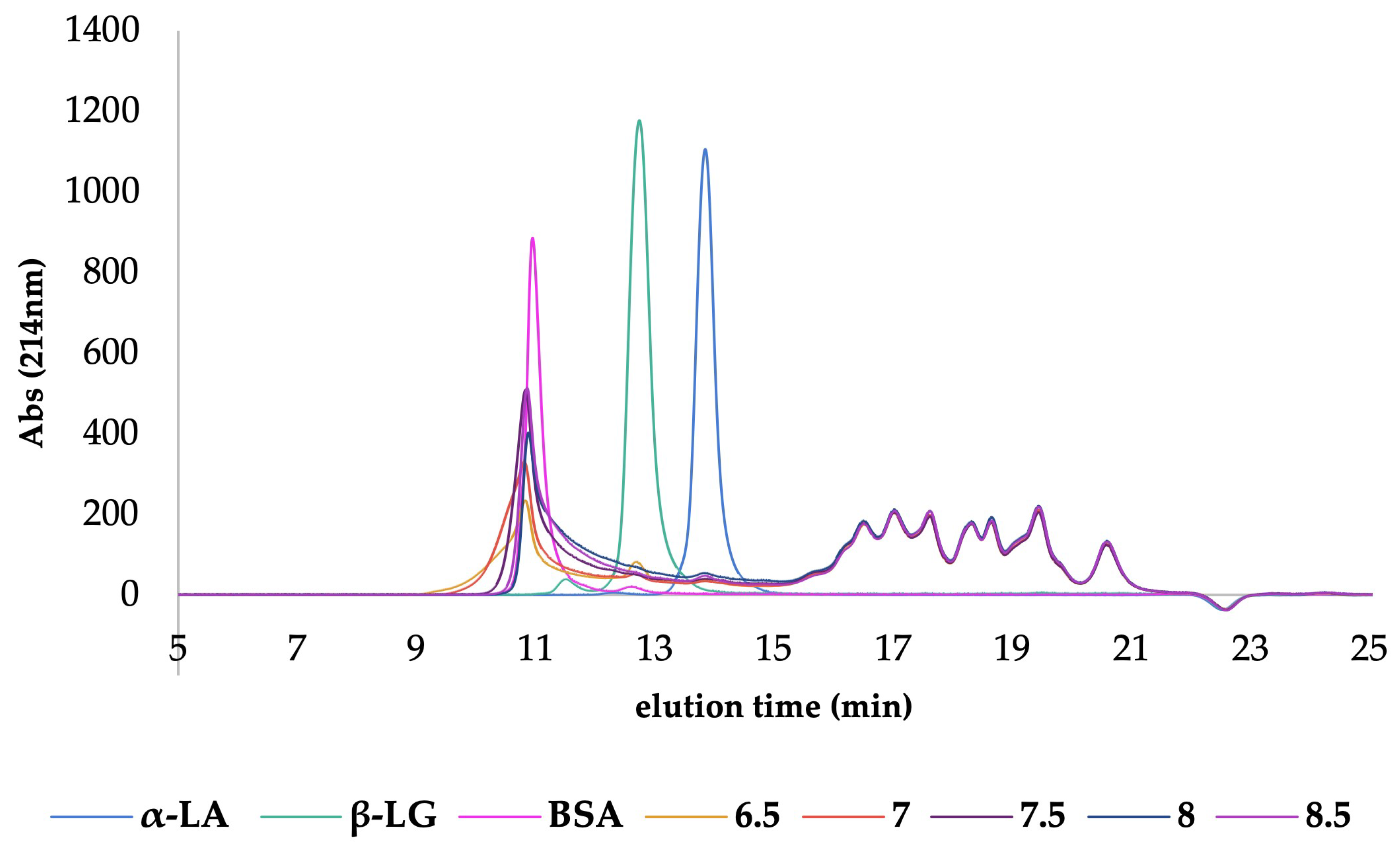
3.2.3. Heat Inactivation: Heat Exchanger
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lahl, W.J.; Braun, S.D. Enzymatic production of protein hydrolysates for food use. Food Technol. 1994, 48, 68–71. [Google Scholar]
- Woiciechowski, A.L.; Nitsche, S.; Pandey, A.; Soccol, C.R. Acid and enzymatic hydrolysis to recover reducing sugars from cassava bagasse: An economic study. Braz. Arch. Biol. Technol. 2002, 45, 393–400. [Google Scholar] [CrossRef]
- Gulati, S.L.; Gaur, A.C. Enzymatic hydrolysis of cellulose from agricultural and industrial wastes. Biol. Wastes 1988, 25, 309–313. [Google Scholar] [CrossRef]
- Guellil, A.; Boualam, M.; Quiquampoix, H.; Ginestet, P.; Audic, J.M.; Block, J.C. Hydrolysis of wastewater colloidal organic matter by extracellular enzymes extracted from activated sludge flocs. Water Sci. Technol. 2001, 43, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.C.; Song, I.S. Enzymatic hydrolysis of foodwaste and methane production using UASB bioreactor. Int. J. Gr. Energy 2011, 8, 361–371. [Google Scholar] [CrossRef]
- Mitidieri, S.; Souza Martinelli, A.H.; Schrank, A.; Vainstein, M.H. Enzymatic detergent formulation containing amylase from Aspergillus niger: A comparative study with commercial detergent formulations. Bioresour. Technol. 2006, 97, 1217–1224. [Google Scholar] [CrossRef]
- McCarthy, A.; O’Callaghan, Y.; O’Brien, N. Protein hydrolysates from agricultural crops—Bioactivity and potential for functional food development. Agriculture 2013, 3, 112–130. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymatic hydrolysis of proteins for increased solubility. J. Agric. Food Chem. 1976, 24, 1090–1093. [Google Scholar] [CrossRef]
- Kong, X.; Zhou, H.; Qian, H. Enzymatic hydrolysis of wheat gluten by proteases and properties of the resulting hydrolysates. Food Chem. 2007, 102, 759–763. [Google Scholar] [CrossRef]
- Banach, J.C.; Lin, Z.; Lamsal, B.P. Enzymatic modification of milk protein concentrate and characterization of resulting functional properties. LWT-Food Sci. Technol. 2013, 54, 397–403. [Google Scholar] [CrossRef]
- Selamassakul, O.; Laohakunjit, N.; Kerdchoechuen, O.; Ratanakhanokchai, K. A novel multi-biofunctional protein from brown rice hydrolysed by endo/endo-exoproteases. Food Funct. 2016, 7, 2635–2644. [Google Scholar] [CrossRef]
- Purschke, B.; Meinlschmidt, P.; Horn, C.; Rieder, O.; Jäger, H. Improvement of techno-functional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur. Food Res. Technol. 2018, 244, 999–1013. [Google Scholar] [CrossRef]
- Ryan, G.; Nongonierma, A.B.; O’Regan, J.; FitzGerald, R.J. Functional properties of bovine milk protein isolate and associated enzymatic hydrolysates. Int. Dairy J. 2018, 81, 113–121. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Franco, C.; Zhang, W. Functions, applications and production of protein hydrolysates from fish processing co-products (FPCP). Food Res. Int. 2013, 50, 289–297. [Google Scholar] [CrossRef]
- Baraldi, I.J.; Giordano, R.L.C.; Zangirolami, T.C. Enzymatic hydrolysis as an environmentally friendly process compared to thermal hydrolysis for instant coffee production. Braz. J. Chem. Eng. 2016, 33, 763–771. [Google Scholar] [CrossRef]
- Clemente, A. Enzymatic protein hydrolysates in human nutrition. Trends Food Sci. Technol. 2001, 11, 254–262. [Google Scholar] [CrossRef]
- Fiocchi, A.; Restani, P.; Bernardini, R.; Lucarelli, S.; Lombardi, G.; Magazzù, G.; Marseglia, G.L.; Pittschieler, K.; Tripodi, S.; Troncone, R.; et al. A hydrolysed rice-based formula is tolerated by children with cow’s milk allergy: A multi-centre study. Clin. Exp. Allergy 2006, 36, 311–316. [Google Scholar] [CrossRef]
- Gauthier, S.F.; Pouliot, Y.; Saint-Sauveur, D. Immunomodulatory peptides obtained by the enzymatic hydrolysis of whey proteins. Int. Dairy J. 2006, 16, 1315–1323. [Google Scholar] [CrossRef]
- Sinha, R.; Radha, C.; Prakash, J.; Kaul, P. Whey protein hydrolysate: Functional properties, nutritional quality and utilization in beverage formulation. Food Chem. 2007, 101, 1484–1491. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.N.S.; Lee, H.Y.; Jo, C.; Suh, J.W.; Ahn, D.U. Enzymatic hydrolysis of ovomucoid and the functional properties of its hydrolysates. Poult. Sci. 2015, 94, 2280–2287. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef]
- Dullius, A.; Goettert, M.I.; de Souza, C.F.V. Whey protein hydrolysates as a source of bioactive peptides for functional foods—Biotechnological facilitation of industrial scale-up. J. Funct. Foods 2018, 42, 58–74. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; O Keeffe, M.B.; Fitz Geralde, R.J. Milk protein hydrolysates and bioactive peptides. In Advanced Dairy Chemistry: Volume 1B: Proteins: Applied Aspects: Fourth Edition; McSweeney, P.L.H., O’Mahony, J.A., Eds.; Springer: New York, NY, USA, 2016; pp. 417–482. [Google Scholar]
- Tsabouri, S.; Douros, K.; Priftis, K.N. Cow’s milk allergenicity. Endocr. Metab. Immune Disord. Drug Targets 2014, 14, 16–26. [Google Scholar] [CrossRef]
- Severin, S.; Xia, W.S. Enzymatic hydrolysis of whey proteins by two different proteases and their effect on the functional properties of resulting protein hydrolysates. J. Food Biochem. 2006, 30, 77–97. [Google Scholar] [CrossRef]
- Walmsley, S.J.; Rudnick, P.A.; Liang, Y.; Dong, Q.; Stein, S.E.; Nesvizhskii, A.I. Comprehensive analysis of protein digestion using six trypsins reveals the origin of trypsin as a significant source of variability in proteomics. J. Proteome Res. 2013, 12, 5666–5680. [Google Scholar] [CrossRef]
- Butré, C.I.; Sforza, S.; Gruppen, H.; Wierenga, P.A. Determination of the influence of substrate concentration on enzyme selectivity using whey protein isolate and Bacillus licheniformis protease. J. Agric. Food Chem. 2014, 62, 10230–10239. [Google Scholar] [CrossRef]
- Deng, Y.; Gruppen, H.; Wierenga, P.A. Comparison of protein hydrolysis catalyzed by bovine, porcine, and human trypsins. J. Agric. Food Chem. 2018, 66, 4219–4232. [Google Scholar] [CrossRef]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmentier, M. Influence of hydrolysis degree on the functional properties of salmon byproducts hydrolysates. J. Food Sci. 2004, 69, C615–C622. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Barre, C.; FitzGerald, R.J. Enzymatic generation of whey protein hydrolysates under pH-controlled and non pH-controlled conditions: Impact on physicochemical and bioactive properties. Food Chem. 2016, 199, 246–251. [Google Scholar] [CrossRef]
- Chae, H.J.; In, M.J.; Kim, M.H. Process development for the enzymatic hydrolysis of food protein: Effects of pre-treatment and post-treatments on degree of hydrolysis and other product characteristics. Biotechnol. Bioprocess Eng. 1998, 3, 35–39. [Google Scholar] [CrossRef]
- O’Loughlin, I.B.; Murray, B.A.; Kelly, P.M.; Fitzgerald, R.J.; Brodkorb, A. Enzymatic hydrolysis of heat-induced aggregates of whey protein isolate. J. Agric. Food Chem. 2012, 60, 4895–4904. [Google Scholar] [CrossRef]
- Guan, H.; Diao, X.; Jiang, F.; Han, J.; Kong, B. The enzymatic hydrolysis of soy protein isolate by Corolase PP under high hydrostatic pressure and its effect on bioactivity and characteristics of hydrolysates. Food Chem. 2018, 245, 89–96. [Google Scholar] [CrossRef]
- Alting, A.C.; Meijer, R.J.G.M.; van Beresteijn, E.C.H. Selective hydrolysis of milk proteins to facilitate the elimination of the ABBOS epitope of bovine serum albumin and other immunoreactive epitopes. J. Food Prot. 2016, 61, 1007–1012. [Google Scholar] [CrossRef]
- Martinez, K.D.; Baeza, R.I.; Millán, F.; Pilosof, A.M.R. Effect of limited hydrolysis of sunflower protein on the interactions with polysaccharides in foams. Food Hydrocoll. 2005, 19, 361–369. [Google Scholar] [CrossRef]
- Avramenko, N.A.; Low, N.H.; Nickerson, M.T. The effects of limited enzymatic hydrolysis on the physicochemical and emulsifying properties of a lentil protein isolate. Food Res. Int. 2013, 51, 162–169. [Google Scholar] [CrossRef]
- Cucu, T.; Platteau, C.; Taverniers, I.; Devreese, B.; De Loose, M.; De Meulenaer, B. Effect of partial hydrolysis on the hazelnut and soybean protein detectability by ELISA. Food Control 2013, 30, 497–503. [Google Scholar] [CrossRef]
- Nieto-Nieto, T.V.; Wang, Y.X.; Ozimek, L.; Chen, L. Effects of partial hydrolysis on structure and gelling properties of oat globular proteins. Food Res. Int. 2014, 55, 418–425. [Google Scholar] [CrossRef]
- Xu, X.; Liu, W.; Liu, C.; Luo, L.; Chen, J.; Luo, S.; McClements, D.J.; Wu, L. Effect of limited enzymatic hydrolysis on structure and emulsifying properties of rice glutelin. Food Hydrocoll. 2016, 61, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Butré, C.I.; Sforza, S.; Wierenga, P.A.; Gruppen, H. Determination of the influence of the pH of hydrolysis on enzyme selectivity of Bacillus licheniformis protease towards whey protein isolate. Int. Dairy J. 2015, 44, 44–53. [Google Scholar] [CrossRef]
- Permyakov, E.A.; Berliner, L.J. α-Lactalbumin: Structure and function. FEBS Lett. 2000, 473, 269–274. [Google Scholar] [CrossRef]
- Layman, D.K.; Lönnerdal, B.; Fernstrom, J.D. Applications for α-lactalbumin in human nutrition. Nutr. Rev. 2018, 76, 444–460. [Google Scholar] [CrossRef]
- Cheison, S.C.; Leeb, E.; Toro-Sierra, J.; Kulozik, U. Influence of hydrolysis temperature and pH on the selective hydrolysis of whey proteins by trypsin and potential recovery of native alpha-lactalbumin. Int. Dairy J. 2011, 21, 166–171. [Google Scholar] [CrossRef]
- Chelulei Cheison, S.; Brand, J.; Leeb, E.; Kulozik, U. Analysis of the effect of temperature changes combined with different alkaline pH on the β-lactoglobulin trypsin hydrolysis pattern using MALDI-TOF-MS/MS. J. Agric. Food Chem. 2011, 59, 1572–1581. [Google Scholar] [CrossRef]
- Lisak, K.; Toro-Sierra, J.; Kulozik, U.; Bozanic, R.; Cheison, S.C. Chymotrypsin selectively digests β-lactoglobulin in whey protein isolate away from enzyme optimal conditions: Potential for native α-lactalbumin purification. J. Dairy Res. 2013, 80, 14–20. [Google Scholar] [CrossRef]
- Wijayanti, H.B.; Bansal, N.; Deeth, H.C. Stability of whey proteins during thermal processing: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1235–1251. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Lardeux, C.; FitzGerald, R.J. Impact of enzyme inactivation conditions during the generation of whey protein hydrolysates on their physicochemical and bioactive properties. Int. J. Food Sci. Technol. 2018, 53, 219–227. [Google Scholar] [CrossRef]
- Guadix, A.; Camacho, F.; Guadix, E.M. Production of whey protein hydrolysates with reduced allergenicity in a stable membrane reactor. J. Food Eng. 2006, 72, 398–405. [Google Scholar] [CrossRef]
- Cheison, S.C.; Wang, Z.; Xu, S.Y. Use of response surface methodology to optimise the hydrolysis of whey protein isolate in a tangential flow filter membrane reactor. J. Food Eng. 2007, 80, 1134–1145. [Google Scholar] [CrossRef]
- Cheison, S.C.; Wang, Z.; Xu, S.Y. Hydrolysis of whey protein isolate in a tangential flow filter membrane reactor. II. Characterisation for the fate of the enzyme by multivariate data analysis. J. Membr. Sci. 2006, 286, 322–332. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| t (min) | % Acetonitrile |
|---|---|
| 0 | 3.0 |
| 20 | 38.4 |
| 26 | 38.4 |
| 28 | 44.0 |
| 31 | 52.0 |
| 33 | 90.0 |
| 37 | 90.0 |
| 38 | 3.0 |
| E:S 1:10 | E:S 1:30 | E:S 1:50 | E:S 1:100 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time (h) | %DH | α-LA (%) | β-LG (%) | %DH | α-LA (%) | β-LG (%) | %DH | α-LA (%) | β-LG (%) | %DH | α-LA (%) | β-LG (%) |
| 1 | 5.71 ± 0.19 | 26.84 ± 2.61 | 86.46 ± 1.87 | 3.93 ± 0.26 | 7.43 ± 1.83 | 70.36 ± 3.10 | 2.82 ± 0.14 | 0.05 ± 5.79 | 58.67 ± 1.51 | 1.30 ± 0.24 | 2.39 ± 2.93 | 30.63 ± 1.58 |
| 2 | 7.00 ± 0.22 | 31.02 ± 2.15 | 97.49 ± 0.76 | 5.07 ± 0.32 | 9.91 ± 3.27 | 80.63 ± 0.57 | 4.01 ± 0.13 | 0.84 ± 2.34 | 73.72 ± 1.67 | 2.14 ± 0.32 | 2.54 ± 2.23 | 47.34 ± 1.53 |
| 3 | 7.83 ± 0.17 | 33.18 ± 1.67 | 100.00 | 5.79 ± 0.37 | 11.83 ± 0.93 | 86.55 ± 1.06 | 4.64 ± 0.15 | 1.51 ± 5.20 | 81.04 ± 0.81 | 2.78 ± 0.35 | 3.52 ± 2.53 | 53.15 ± 2.87 |
| 4 | 6.43 ± 0.38 | 11.59 ± 0.93 | 90.54 ± 1.11 | 5.08 ± 0.20 | 3.83 ± 6.92 | 86.19 ± 0.19 | 3.32 ± 0.36 | 3.47 ± 4.99 | 63.90 ± 0.84 | |||
| 5 | 6.88 ± 0.44 | 9.82 ± 2.88 | 93.14 ± 0.40 | 5.46 ± 0.23 | 4.11 ± 3.44 | 88.32 ± 0.44 | 3.70 ± 0.34 | 3.00 ± 1.82 | 69.81 ± 2.19 | |||
| 6 | 7.34 ± 0.43 | 11.06 ± 4.43 | 95.38 ± 0.60 | 5.88 ± 0.19 | 4.43 ± 3.47 | 91.71 ± 0.45 | 4.08 ± 0.26 | 3.13 ± 2.28 | 73.27 ± 1.35 | |||
| 7 | 7.86 ± 0.48 | 14.10 ± 3.31 | 97.22 ± 0.50 | 6.17 ± 0.20 | 4.68 ± 2.88 | 93.85 ± 0.31 | 4.39 ± 0.23 | 4.48 ± 3.22 | 77.54 ± 1.01 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáez, L.; Murphy, E.; FitzGerald, R.J.; Kelly, P. Exploring the Use of a Modified High-Temperature, Short-Time Continuous Heat Exchanger with Extended Holding Time (HTST-EHT) for Thermal Inactivation of Trypsin Following Selective Enzymatic Hydrolysis of the β-Lactoglobulin Fraction in Whey Protein Isolate. Foods 2019, 8, 367. https://doi.org/10.3390/foods8090367
Sáez L, Murphy E, FitzGerald RJ, Kelly P. Exploring the Use of a Modified High-Temperature, Short-Time Continuous Heat Exchanger with Extended Holding Time (HTST-EHT) for Thermal Inactivation of Trypsin Following Selective Enzymatic Hydrolysis of the β-Lactoglobulin Fraction in Whey Protein Isolate. Foods. 2019; 8(9):367. https://doi.org/10.3390/foods8090367
Chicago/Turabian StyleSáez, Laura, Eoin Murphy, Richard J. FitzGerald, and Phil Kelly. 2019. "Exploring the Use of a Modified High-Temperature, Short-Time Continuous Heat Exchanger with Extended Holding Time (HTST-EHT) for Thermal Inactivation of Trypsin Following Selective Enzymatic Hydrolysis of the β-Lactoglobulin Fraction in Whey Protein Isolate" Foods 8, no. 9: 367. https://doi.org/10.3390/foods8090367