
The Environment, Farm Animals and Foods as Sources of Clostridioides difficile Infection in Humans

Teagasc Food Research Centre, Ashtown, D15 DY05 Dublin, Ireland
*
Author to whom correspondence should be addressed.
Foods 2023, 12(5), 1094; https://doi.org/10.3390/foods12051094
Submission received: 14 November 2022
/
Revised: 21 February 2023
/
Accepted: 28 February 2023
/
Published: 4 March 2023
(This article belongs to the Special Issue Foodborne Pathogens Management: From Farm and Pond to Fork)
Abstract
:The recent discovery of the same Clostridioides difficile ribotypes associated with human infection in a broad range of environments, animals and foods, coupled with an ever-increasing rate of community-acquired infections, suggests this pathogen may be foodborne. The objective of this review was to examine the evidence supporting this hypothesis. A review of the literature found that forty-three different ribotypes, including six hypervirulent strains, have been detected in meat and vegetable food products, all of which carry the genes encoding pathogenesis. Of these, nine ribotypes (002, 003, 012, 014, 027, 029, 070, 078 and 126) have been isolated from patients with confirmed community-associated C. difficile infection (CDI). A meta-analysis of this data suggested there is a higher risk of exposure to all ribotypes when consuming shellfish or pork, with the latter being the main foodborne route for ribotypes 027 and 078, the hypervirulent strains that cause most human illnesses. Managing the risk of foodborne CDI is difficult as there are multiple routes of transmission from the farming and processing environment to humans. Moreover, the endospores are resistant to most physical and chemical treatments. The most effective current strategy is, therefore, to limit the use of broad-spectrum antibiotics while advising potentially vulnerable patients to avoid high-risk foods such as shellfish and pork.
1. Introduction
Clostridioides difficile is a Gram-positive, endospore-forming anaerobic bacterium often carried asymptomatically in the human gastrointestinal tract [1,2,3,4,5]. However, when conditions are favourable, the endospores germinate in the colon, vegetative cells multiply, and toxins are produced [6], resulting in watery, non-bloody diarrhoea with abdominal pain, toxic megacolon and/or pseudomembranous colitis, which may be fatal [7,8,9].
The most common risk factor associated with CDI is the use/misuse of broad-spectrum antibiotics. C. difficile is often resistant to a wide range of antibiotics [10], and the administration of antibiotics like clindamycin, cephalosporins, penicillins and fluoroquinolones eliminate competitive bacteria in the colon and promote C. difficile outgrowth [11]. The elderly, infants, other immune compromised, and patients on antibiotic therapies are therefore most at risk [1,2,4], although the incidence of CDI in pregnant women, children and patients with inflammatory bowel disease (IBD) has also increased [12].
The generally accepted route for human CDI is transmission from the healthcare environment [13]. However, in recent years the proportion of community-acquired CDI, where the patient has no association with a healthcare facility, has increased [14]. At the same time, non-human reservoirs, including the natural environment (soil, rivers and lakes) [15] and animals, including domestic pets [16,17], food animals [18,19,20] and wild fauna [21] have been reported. Moreover, food may be contaminated [22,23].
The link between C. difficile and animals has been known for at least 60 years. In 1960, McBee [24] isolated this bacterium from the large intestine of a seal in Antarctica. By 1974 C. difficile had also been detected in animal faeces (donkeys, horses, cows and camels) and in the environment (hay, soil, sand and mud) [25]. In the early 1980s, C. difficile reservoirs were reported in healthy pigs and cattle [26,27] and in asymptomatic domestic pets, such as dogs, cats and birds, which had a prevalence of 21%, 30% and 33%, respectively [28]. Thus it was suggested that animals could be a vehicle of transmission to humans [29]. Interestingly, a common human pathogenic C. difficile ribotype (ribotype 078) was also isolated from pigs, cattle, and horses later, providing additional evidence of zoonotic transmission of C. difficile between animals and humans [30,31,32,33]. In more recent years, several studies have reported C. difficile in animals, on carcasses [21,34], in food processing facilities and in both raw and cooked foods [35,36,37,38,39,40,41].
Despite the increase in community-acquired CDI and data on C. difficile in the food chain, it is difficult to prove the source of infection in a given patient or outbreak as the same ribotypes and strains are common to both healthcare and food chain sources. Moreover, the patient may have acquired C. difficile sometime before the conditions in the colon changed to promote outgrowth. The objective of this review was to examine the evidence (CDI, virulence, ribotypes, environment, food animal and food sources and the current epidemiology of CDI in humans) supporting the hypothesis that C. difficile may be foodborne.
2. C. difficile Infection (CDI) in Humans
Elderly people are especially vulnerable to CDI, and cases are more likely to result in severe outcomes [42], possibly due to a decreased immune response or changes in the intestinal microbiota with age [43,44]. An underlying condition, chemotherapy or gastrointestinal surgery can increase susceptibility to CDI [45], which may become recurrent, leading to increased morbidity and mortality [46,47]. Broad-spectrum antibiotics significantly reduce the gut microflora diversity and alter the bile composition in the colon, facilitating CDI and recurrent infection in humans [48]. Treatment with acid suppression medication to prevent ulcers or treat acid-related diseases is also a risk factor for recurrence [49,50,51].
Metronidazole is used to treat mild to moderate CDI, while vancomycin is used in more severe cases, although the combination of both may be used when there are complications [52]. When these are ineffective, fidaxomicin has been proposed as an alternative to vancomycin [53,54] and has proven effective in preventing recurrent infection [55].
3. Virulence
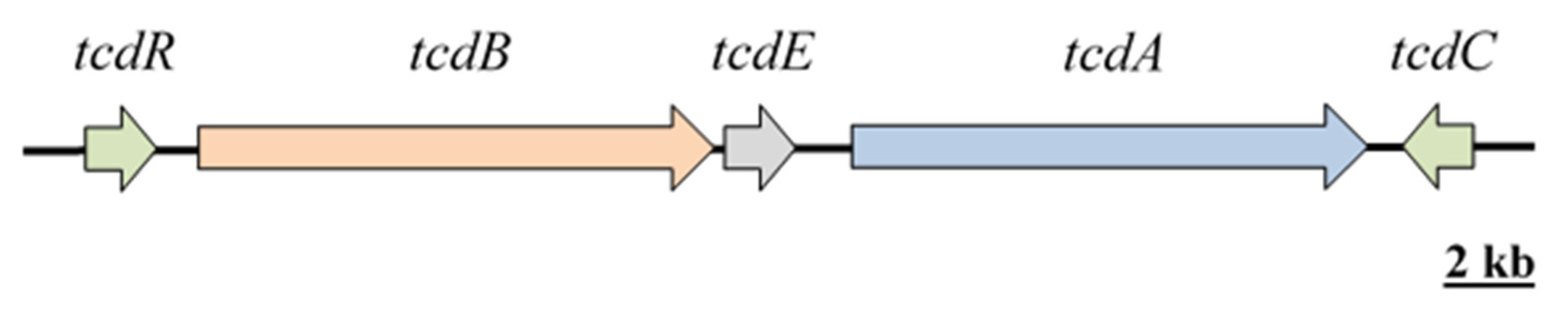
Within the host, C. difficile endospores germinate into vegetative cells, colonise the intestinal tract and produce toxins resulting in disease [56,57], which causes intestinal inflammation, perforation, toxic megacolon and pseudomembranous colitis [58,59]. Mortality rates range from less than 2% to 17% [60,61]. The main virulence factors in C. difficile are toxin A and toxin B, encoded by the tcdA (308 kDa) and tcdB (270 kDa) genes located on a pathogenicity locus (PaLoc) (Figure 1 and Table 1). Both are large clostridial glycosylation toxins and are activated in response to environmental signals during the late log and stationary phases. In addition to the toxins, two regulatory proteins (TcdR and TcdC) and a protein whose function remains unclear (TcdE) complete the PaLoc [62,63]. TcdR (also referred to as TcdD) is a positive regulator activated in stationary phase growth, while TcdC is a negative regulator produced during the exponential phase. Mutations, such as deletions in the tcdC gene, may cause increased production of toxins A and B [62,64].
TcdA and TcdB possess the same biological activities, among which is the disruption of the cytoskeleton that leads to cytopathic effects in cultured cells. They also possess proinflammatory activity and can stimulate intestinal epithelial cells and immune cells to produce cytokines and chemokines [66,67]. Even low doses of toxins A and B damage the tight junctions of the gut epithelial barrier, facilitating the translocation of commensal bacteria, inflammation and cell apotheosis [66,67,68]. Sequence variations, deletions, and duplications within the pathogenicity locus account for different toxinotypes of C. difficile, with 27 currently identified. Certain strains can present only one of the toxins genes (A−B+ or A+B−), however, they reportedly still cause severe disease in humans [62]. In addition, the cytotoxicitybetween toxins that belong to different toxinotypes may vary, making the relation between strain type and CDI severity even more complex [59]. Strains lacking toxin A are more frequently reported due to deletions in the receptor-binding repetitive regions of TcdA caused by the recombination between short repetitive sequences highly conserved in this toxin gene [63]. Donta et al. [66] reported TcdB to be 4 to 200-fold more cytotoxic than TcdA in a mouse model. Therefore, strains producing toxin B have a higher severity in humans.
Up to a third of C. difficile isolates also produce the transferase C. difficile binary toxin (CDT) [69,70]. CDT, composed of CDTa (biological activity) and CDTb (binding), inhibits the protein actin, damaging the cytoskeleton of the gastrointestinal tract (GIT) cells [71]. The presence of the full-length CDT locus implies the potential expression of the binary toxin, and although some strains contain portions of the CDT locus, these are predicted as non-binary toxin-producing strains [68,70]. CDT-producing strains have been previously associated with a higher production of toxins A and B, leading to an increased disease severity [71,72]. However, CDT is not always present in severe cases [73,74]. In addition, CDT can also be produced by only B+ and non-toxigenic strains (A−B−) [72]. Although CDT production is commonly associated with higher severity of C. difficile infection, the role of this toxin during infection and its mechanism of secretion is still not well understood.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The virulence factors in C. difficile and their function.
| Virulence Factor | Encoding Genes | Role in CDI | References |
|---|---|---|---|
| Toxin A | tcdA | Multiple cytopathic and cytotoxic effects on the targeted cells include disruption of Rho, Rac and Cdc42-dependent signalling, the actin cytoskeleton and the tight adherence junctions, increasing epithelial permeability, allowing commensal bacterial translocation, inflammation, diarrhoea and sometimes death. | [66,67,68,75] |
| Toxin B | tcdB | ||
| TcdR | tcdR | TcdR is a positive regulator (produced in response environmental conditions) that triggers the induction of transcription of the toxin genes (tcdA and tcdB). | [76,77] |
| TcdC | tcdC | TcdC is a negative regulator that inhibits the expression of tcdA and tcdB. Mutations may cause increased production of toxins A and B. | [62,64] |
| TcdE | tcdE | TcdE may function as a lytic protein to facilitate the release of toxins A and B to the extracellular environment by a phage-like system, as these toxins lack signal peptides. | [78,79] |
| CDT | cdtA & cdtB | C. difficile binary toxin (CDT) is a transferase that disrupts the normal cytoskeletal function of cells by inhibiting the protein actin. The altered actin cytoskeleton causes an imbalance between actin and microtubules. | [69,70,71] |
4. Ribotypes
There are in excess of 800 C. difficile ribotypes (RT), some of which are associated with increased virulence [6,80,81], including RT027 and RT078 [82,83]. These ribotypes are also more prevalent in human cases. RT027 (toxinotype III) has a mutation in tcdC, resulting in significantly increased production of toxins A and B while also carrying the genes encoding CDT production and fluoroquinolone resistance [84,85]. Although prevalence has decreased in Europe in recent years, RT027 is associated with a higher mortality and morbidity rate than other ribotypes [86]. The fluoroquinolone resistance, which emerged in two genetically distinct epidemiological lineages (FQR1 and FQR2), was a key driver in the rapid emergence of RT027 [57]. Moreover, this is essential to the increased severity of this ribotype, as this strain typically infects elderly hospital patients on fluoroquinolone treatment [5].
RT078 carries a 39 bp deletion in the tcdC gene and therefore overproduces toxins A and B in addition to the binary toxin CDT. In contrast to RT027, which is mostly hospital-acquired, RT078 is more prevalent in younger people and is generally associated with the community [87]. RT078 strains are resistant to fluoroquinolones and erythromycin, which has contributed to their higher prevalence in CDI [88]. Ribotype 126 has the same mutation in its tcdC gene found in RT078, is resistant to moxifloxacin and tetracycline and is also considered hypervirulent [89,90,91]. Other significant ribotypes from a public health perspective include RT017 and RT018. Although the former only produces toxin B, it is resistant to fluoroquinolones and rifampicin and has been associated with numerous outbreaks [92,93,94]. RT018 has high toxin production capacity, increased cell adhesion, is multidrug-resistant (erythromycin, clindamycin and moxifloxacin) and has become endemic in several countries, including Italy, Spain, Austria and Slovenia [95,96,97].
5. C. difficile in the Environment, Farm Animals and Food
5.1. Water
Toxigenic C. difficile has been isolated from a variety of aquatic environments, including drinking water, rivers, sewage effluent and swimming pools [98,99]. Coastal beaches and river sediments are also contaminated [98,99], in some cases by runoff from fields or effluents from wastewater treatment plants [100]. Indeed, C. difficile is often detected in water from treatment plants [101], and contamination of drinking water was the source of at least one C. difficile outbreak in Finland [102]. Thus, C. difficile survives in water and through the effluent treatment process [100].
5.2. Soil, Manure and Silage
C. difficile is commonly found in soil on farms as well as in forests, recreational parks, residential gardens, etc. [103,104,105,106,107]. These authors reported the highest prevalence in urban settings (57%), followed by farms (31%) and forests (28%). Shivaperumal et al. [108] found prevalence rates of 62%, 13% and 15% in garden soil, manure and compost, respectively, while Fröschle et al. [109] reported C. difficile to be the most prevalent Clostridium spp. in grass silage and cattle manure.
5.3. Farm Environment and Animals
Marcos et al. [110] reported that C. difficile were widespread in soil, water and faeces on beef, sheep and broiler farms, with the prevalence ranging from 7% to 83% and counts from 2.9 to 8.4 log10 cfu/g or /mL, depending on the animal species and sample type being tested. Other studies also found C. difficile in the faeces of a range of farm animals, including cattle, sheep, poultry and pigs [111,112,113,114,115,116]. Of these, pigs are the most important source of C. difficile [113,116], with the relative prevalence by age being 45%, 3% and 1% in suckling piglets, post-weaning piglets and finishing pigs, respectively [114]. Although these animals may show symptoms (diarrhoea), most are asymptomatic [114]. Other similar studies have reported a prevalence of 37% [115] and 78% [111] in piglets and 4% [115], 62% [117] and 9% [16] in mature pigs.
C. difficile are also found in cattle, especially younger animals. Rodriguez et al. [113] reported a prevalence of 11% in calves and 6% in adult cattle. Other studies have found these bacteria in 11% [118], 14% [117] and 22% of calves [111] and 7% of mature animals [16]. Sheep, including lambs, are also potential carriers, with 0.6 to 2% in the former and 7% reported in the latter [16,119].
5.4. C. difficile at the Animal Slaughter Stage
Pathogenic bacteria in faeces on the hide/fleece or in the gastrointestinal tract are readily transferred to the carcass during slaughter and dressing [125]. C. difficile was found in 1%, 3% and 28% of porcine gut contents at slaughter in Belgium [126], Austria [18] and the Netherlands [127], respectively. Reported carcass contamination rates include 7% in Belgium [126], 15% in Canada [128] and 23% in Taiwan [129]. The prevalence of bovine carcass contamination ranges from 7–8% [111,126] but may be as high as 34% [130]. Ovine carcass contamination rates of 15% and 25% have been reported in Iran and Turkey, respectively [130,131]. While poultry carcass data is lacking, Candel-Pérez et al. [132] found C. difficile in 28% of gizzard and 6% of liver samples collected in a poultry processing plant in Spain. In Ireland, beef, sheep and broiler carcass contamination rates ranged from 40% to 100%, 40% to 60% and 10% to 40%, respectively, depending on the sampling stage during carcass processing [16].
Ribotypes 002, 005, 013, 014, 015, 019, 035, 062, 081, 087 and 126 have been identified in porcine faeces and rectal swabs at slaughter plants in Europe [18,111,126,127,133]. The C. difficile ribotypes isolated from other animal carcasses include 027 from cattle and IR46 from ovine carcasses [131]. Poultry slaughter data is lacking, although Koene et al. [16] found toxigenic ribotypes 056, 014 and 003 in faecal samples from poultry in Dutch slaughter plants.
5.5. C. difficile in Retail Foods
C. difficile has been reported in a range of foods at the retail stage. Thus, the consumption of contaminated retail foods, especially ready-to-eat (RTE) foods, is a risk factor for human infection [134]. Marcos et al. tested meat, dairy and vegetable retail foods and detected C. difficile in 9 out of the 240 samples tested [110]. These include corned beef (1), spinach leaves (2), iceberg and little gem lettuce (1 sample each), wild rocket, coleslaw, whole milk yoghurt and cottage cheese (also 1 sample each). Of these samples, direct counts were obtained for the spinach leaves (5.8 log10 cfu/g), coleslaw (4.3 log10 cfu/g) and cottage cheese (6.8 log10 cfu/g).
5.6. C. difficile in Meat and Seafood
Both raw and RTE meat and seafood are frequently contaminated with C. difficile [35,118], and the prevalence, including toxin gene profiles and ribotypes, is summarised in Table 2. The reported contamination rates include 41% [35] and 20% [135] for raw pork meat, 12% for ground pork meat [36] and up to 29% for pork sausages and RTE pork products [135]. A beef contamination rate of 42% was reported by Rodriguez-Palacios et al. [118], while ground beef rates include 2% [37], 12% [36], 20% [118] and 50% [35]. In one study, de Boer et al. [38] detected C. difficile in 6% of raw lamb samples. Reported poultry contamination rates include 1% [38], 3% [39], 8% [136,137], 13% [36,120] and 44% [35]. C. difficile has also been detected in shellfish and fish in several countries, with prevalence ranging from 4% to 49% [138,139,140,141].
5.7. C. difficile in Vegetables
The information on C. difficile isolated from vegetables is summarised in Table 3, with overall prevalence rates of 2% to 5% [22,103,142]. Lim et al. detected C. difficile in 56% of organic and 50% of non-organic potatoes, 22% of organic beetroots, 56% of organic onions and 53% of organic carrots [143]. Tkalec et al. found this pathogen in 9% of leaf vegetables, 7% of ginger, 26% and 60% of potatoes, and 14.3% of homegrown leaf vegetables [144]. RTE salads contamination rates included 2% [41], 3% [142], 3.3% [145] and 8% (153].
All of these ribotypes have toxin genes associated with illness in humans. Many have been isolated directly from patients with CDI (Table 4), including 001, 002, 003, 010, 011, 012, 014, 015, 017, 018, 020, 023, 027, 029, 070, 071, 072, 077, 078, 087, 101, 126, 137 and 150. Of these, 002, 003, 012, 014, 027, 029, 070, 078 and 126 have been reported in confirmed community-acquired CDI, while 001, 017, 027, 072, 078 and 126 are hypervirulent.
5.8. Meta-Analysis
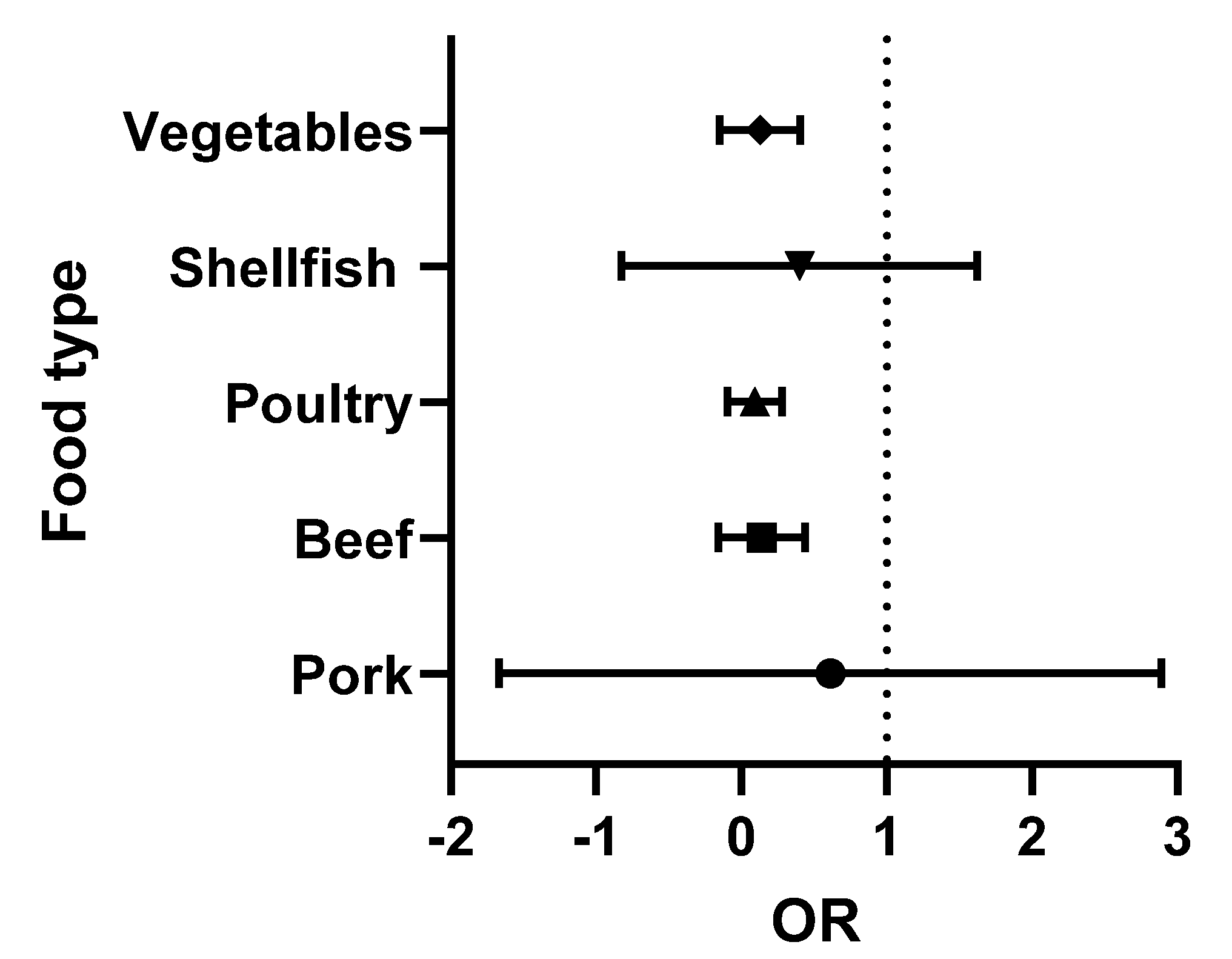
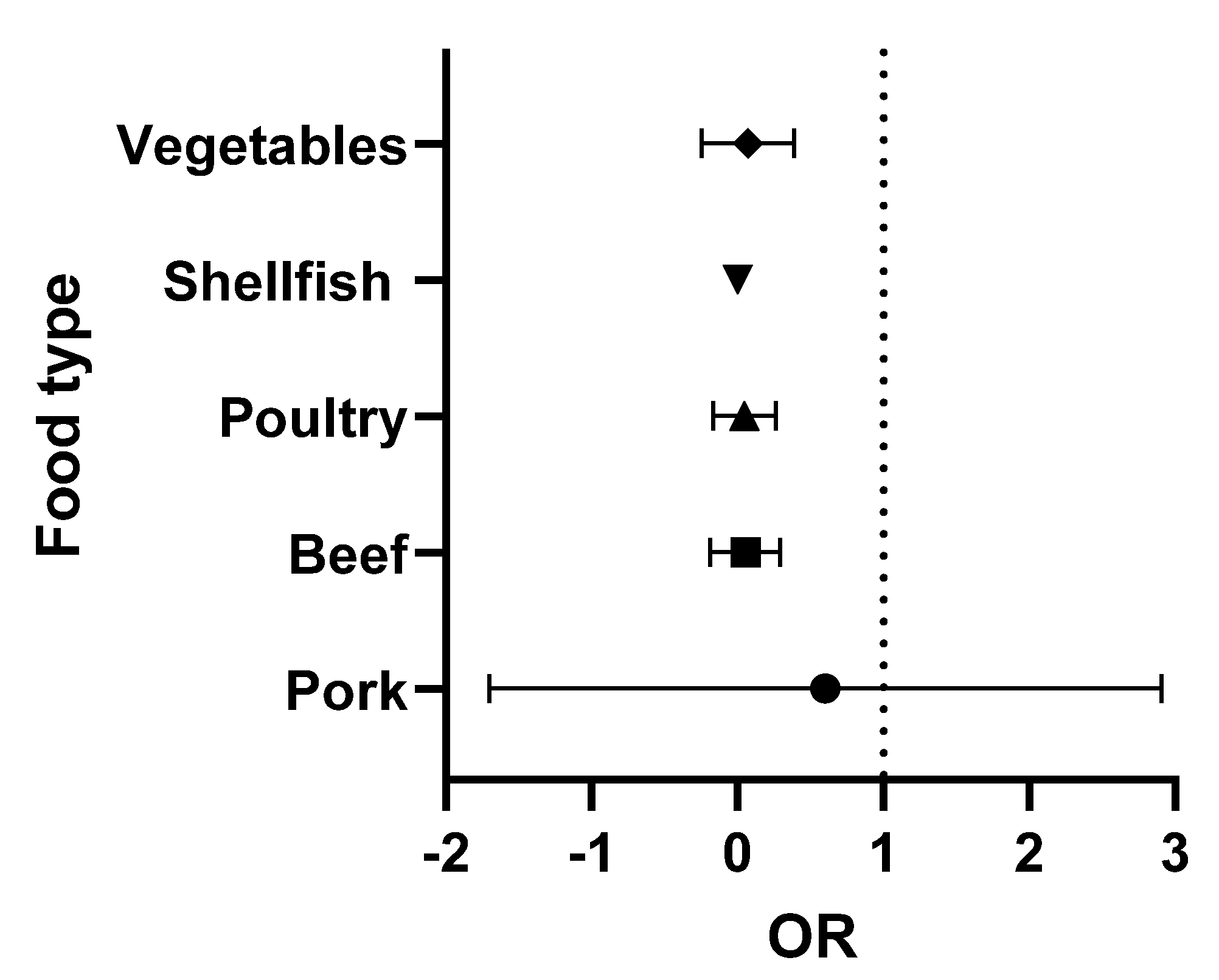
The data presented in Table 2 and Table 3 were analysed using Graphpad Prism version 9.3.1. The odds ratios (OR) (the odds of consuming a contaminated product) were calculated for each food type. Briefly, the OR was calculated as the number of positive over negative samples reported for each study. Turkey (with only two studies) was combined with the chicken data (poultry category), while the single lamb study was omitted. The medians and 95% confidence intervals were obtained and were then used to prepare the forest plots. In these Figures, the vertical line is set at an OR = 1 (50:50 chance of the food being contaminated). When all ribotypes are considered, shellfish and pork present a higher risk to the consumer (Figure 2). However, when the analysis is repeated, focusing exclusively on ribotypes 027 and 078 (the 2 hypervirulent strains most commonly associated with human infection), the increased risk is only associated with the consumption of pork (Figure 3).
6. The Epidemiology of Foodborne Infection
In 1978, C. difficile was recognised as the causative agent of pseudomembranous colitis and diarrhoea in patients on antimicrobial therapy and it was a hospital-associated disease [157]. In the 1980s and 1990s, the incidence of CDI increased significantly, driven by the use of broad-spectrum third-generation cephalosporins (to which C. difficile is intrinsically resistant), but the disease was rarely fatal [158,159]. There was a further increase in CDI in the first 10 years of this century driven by the emergence and epidemic spread of the hypervirulent strain, ribotype 027 [160]. The epidemiology of CDI also changed in terms of clinical presentation, response to treatment, and disease outcome. Community-acquired CDI, defined as cases with symptom onset in the community with no history of hospitalisation in the previous 12 weeks or symptom onset within 48 h of hospital admission [161], also emerged. Since then, the incidence of CDI has remained high in developed countries [159,162], and rates of community acquired CDI have increased, accounting for 41%, 30% and 14% of total CDI in the USA, Australia and the EU, respectively [96,159,163]. Furthermore, community acquired CDI patients are generally younger, healthy, often female and lack the traditional risk factors of CDI, including a history of antimicrobial usage [164].
The natural habitat of C. difficile is the mammalian gastrointestinal tract (GIT). These bacteria colonise the neonatal GIT, proliferate and are excreted in the faeces to which other newborn animals are exposed, and the cycle recommences. As mammals develop, other bacterial species colonise the GIT, and the prevalence of C. difficile decreases [165]. The GIT microbiota inhibit germination, vegetative growth and toxin production, thus protecting against C. difficile [48]. However, in the 1990s, this protection was removed when cephalosporins were used in animal husbandry, and food animals became a major reservoir and amplification host for C. difficile [119,166], resulting in the contamination of the environment and a range of foods [100,119,166].
Once the environment is contaminated, there are multiple direct and indirect routes to humans, including via food (as illustrated in Figure 4). It is all but impossible to provide incontrovertible proof of foodborne transmission because of the ubiquitous nature of C. difficile, delayed onset of symptoms, ability to persist for extended periods as an endospore, etc. However, it has been shown that C. difficile endospores in animal waste, wastewater treatment sludge, soil, manure and compost may survive for extended periods of time, facilitating direct contamination of vegetables and fruit or meat via cross-contamination of carcasses during slaughter and processing [108,147]. Water also frequently contains C. difficile endospores [99,100,148], and food production may also be contaminated via water used for irrigation or food processing [100,144]. Moreover, the presence of endospores in rivers may contaminate fish and seafood [100,138,139,141]. Transfer from food and wild animals and from domestic pets has also been described [116,149].
Of particular interest, from the public health perspective, is the detection of similar C. difficile isolates in farm animals and in humans suffering from CDI, suggesting this pathogen may be zoonotic [150]. Whole genome sequencing (WGS) analysis has shown that ribotypes 078, 126 and 066, commonly found in pigs and/or cattle, are genetically identical to those in humans [151,152,153,154,155]. Although ribotype data for sheep is limited, ribotypes 014, 010 and 045 are common to both humans and ovine sources [119,156,167], while human-related ribotypes 001, 014 and 039 are also found in broilers [120,121,122,168].
7. Control Strategies
CDI can be controlled in hospitals using deep environmental cleaning, appropriate hand hygiene, stringent infection control and antimicrobial stewardship [169]. However, the same strategies cannot be used in agriculture and food processing [15]. Reduced usage of antibiotics in food animal production would reduce C. difficile amplification but is unlikely as increasing global food demand is driving increased antimicrobial usage in animal husbandry, which is projected to rise by 67% by 2030 [15,140]. In 2006 the EU banned the use of antibiotics as growth promoters, followed by the USA in 2017, but other major food-producing countries still allow this practice [170].
Preventing the recycling and dissemination of C. difficile endospores in animal slurries applied to land as organic fertilisers would also facilitate reduced environmental contamination and animal carriage. However, research is required to develop effective treatments [171]. Vaccination of food-producing animals is another possible control strategy, but an effective vaccine has not been developed yet [15]. Controlling C. difficile in food is dependent on reducing or eliminating the endospores, which are resistant to chilled (4 °C) and freezing (−18 °C and −80 °C) temperatures [172,173]. Although the endospores are resistant at 80 °C [172,173,174] and will survive the recommended cooking time temperature combinations recommended for meat [174], they are eliminated at 98 °C for 2 min [175]. The same authors suggested microwave irradiation (800 W/60 s) also achieved complete inactivation by denaturing the outer coat.
C. difficile endospores are also resistant to desiccation, hydrostatic pressure [37,176,177,178,179] and a range of food preservatives, including sodium nitrite, sodium nitrate and sodium metabisulfite, at permitted concentrations [180]. In contrast, nisin [181], black seed oil, myrrh water [182], garlic juice, peppermint oil, trans-cinnamaldehyde, allicin, menthol and zingerone [183] have a potential application, but validation studies are required before they can be used in controlling C. difficile in food.
8. Conclusions
Based on the information provided, it was concluded that C. difficile is widespread in the environment and along the food chain. Many food isolates carry the virulence factors required for human infection, and there is no conceivable reason why food is not a source of these pathogens. This conclusion is further supported by the presence of the same ribotypes in food and humans suffering from community-acquired CDI. Based on our analysis, potentially vulnerable consumers should be advised not to handle or consume shellfish or pork.
Author Contributions
Conceptualization, D.B.; investigation, D.B. and P.M.; data curation, P.M.; writing—original draft preparation, D.B.; writing—review and editing, D.B. and P.M.; supervision, D.B.; project administration, D.B.; funding acquisition, D.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Food Institutional Research Measure (FIRM) administered by the Department for Agriculture, Food and the Marine (DAFM) (Grant number 17F206). Pilar Marcos was supported by the Teagasc Walsh Scholarship Scheme (number 2018210).
Data Availability Statement
Not applicable.
Acknowledgments
The pictographic symbols used are the property of the Government of Aragón and have been created by Sergio Palao for the Aragonese Portal of Augmentative and Alternative Communication (ARASAAC) (http://www.arasaac.org; accessed on 27 September 2022) that distributes them under Creative Commons License BY-NC-SA.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Heinlen, L.; Ballard, J.D. Clostridium difficile infection. Am. J. Med. Sci. 2010, 340, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, C.; Poilane, I.; De Pontual, L.; Maherault, A.C.; Le Monnier, A.; Collignon, A. Clostridium difficile carriage in healthy infants in the community: A potential reservoir for pathogenic strains. Clin. Inf. Dis. 2012, 55, 1209–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galdys, A.L.; Curry, S.R.; Harrison, L.H. Asymptomatic Clostridium difficile colonization as a reservoir for Clostridium difficile infection. Expert Rev. Anti-Infect. Ther. 2014, 12, 967–980. [Google Scholar] [CrossRef]
- Martin, J.S.; Monaghan, T.M.; Wilcox, M.H. Clostridium difficile infection: Epidemiology, diagnosis and understanding transmission. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Czepiel, J.; Dróżdż, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultańska, D.; Garlicki, A.; Biesiada, G. Clostridium difficile infection: Review. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Vedantam, G.; Clark, A.; Chu, M.; McQuade, R.; Mallozzi, M.; Viswanathan, V.K. Clostridium difficile infection: Toxins and non-toxin virulence factors, and their contributions to disease establishment and host response. Gut Microbes 2012, 3, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Manabe, Y.C.; Vinetz, J.M.; Moore, R.D.; Merz, C.; Charache, P.; Bartlett, J.G. Clostridium difficile colitis: An efficient clinical approach to diagnosis. Ann. Intern. Med. 1995, 123, 835–840. [Google Scholar] [CrossRef]
- Borriello, S.P. Pathogenesis of Clostridium difficile infection. J. Antimicrobial. Chemo. 1998, 41, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Brazier, J.S. Clostridium difficile: From obscurity to superbug. Brit. J. Biomed. Sci. 2008, 65, 39–44. [Google Scholar] [CrossRef]
- Peng, Z.; Jin, D.; Kim, H.B.; Stratton, C.W.; Wu, B.; Tang, Y.-W.; Sun, X. Update on Antimicrobial Resistance in Clostridium difficile. J. Clin. Microbiol. 2017, 55, 1998–2008. [Google Scholar] [CrossRef] [Green Version]
- Owen, R.C.; Donskey, C.J.; Gaynes, R.P.; Loo, V.G.; Muto, C.A. Antimicrobial-associated risk factors for Clostridium difficile infection. Clin. Infect. Dis. 2008, 46, 19–31. [Google Scholar]
- Ananthakrishnan, A. Clostridium difficile infection: Epidemiology, risk factors and management. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Enoch, D.A.; Aliyu, S.H. Is Clostridium difficile infection still a problem for hospitals? Can. Med. Assoc. J. 2012, 184, 17–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.; Zhu, N.A. Community-acquired Clostridium difficile infection. Can Fam Physician. 2017, 63, 131–132. [Google Scholar]
- Lim, S.C.; Knight, D.R.; Riley, T.V. Clostridium difficile and One Health. Clin. Microbiol. Infect. 2020, 26, 857–863. [Google Scholar] [CrossRef]
- Koene, M.; Mevius, D.; Wagenaar, J.; Harmanus, C.; Hensgens, M.; Meetsma, A.; Putirulan, F.; van Bergen, M.; Kuijper, E. Clostridium difficile in Dutch animals: Their presence, characteristics and similarities with human isolates. Clin. Microbiol. Infect. 2012, 18, 778–784. [Google Scholar] [CrossRef] [Green Version]
- Schneeberg, A.; Rupnik, M.; Neubauer, H.; Seyboldt, C. Prevalence and distribution of Clostridium difficile PCR ribotypes in cats and dogs from animal shelters in Thuringia, Germany. Anaerobe 2012, 18, 484–488. [Google Scholar] [CrossRef]
- Indra, A.; Lassnig, H.; Baliko, N.; Much, P.; Fiedler, A.; Huhulescu, S. Clostridium difficile: A new zoonotic agent? Wien. Klin. Wochenschr. 2009, 121, 91–95. [Google Scholar] [CrossRef]
- Schmid, A.; Messelhäusser, U.; Hörmansdorfer, S.; Sauter-Louis, C.; Mansfeld, R. Occurrence of zoonotic Clostridia and Yersinia in healthy cattle. J. Food Protect. 2013, 76, 1697–1703. [Google Scholar] [CrossRef]
- Noren, T.; Johansson, K.; Unemo, M. Clostridium difficile PCR-ribotype 046 is common among neonatal pigs and humans in Sweden. Clin. Microbiol. Infect. 2014, 20, O2–O6. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Palacios, A.; Borgmann, S.; Kline, T.R.; LeJeune, J.T. Clostridium difficile in foods and animals: History and measures to reduce exposure. Animal Health Res. Rev. 2013, 14, 11–29. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, D.; Costa, M.C.; Dew, W.M.; Weese, J.S. Clostridium difficile in vegetables, Canada. Lett. Appl. Microbiol. 2010, 51, 600–602. [Google Scholar] [CrossRef] [PubMed]
- Weese, J. Clostridium difficile in food—Innocent bystander or serious threat? Clin. Microbiol. Infect. 2010, 16, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBee, R.H. Intestinal flora of some antarctic birds and mammals. J. Bacteriol. 1960, 79, 311–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafiz, S. Clostridium difficile and Its Toxins. Ph.D. Thesis, Department of Microbiology, University of Leeds, Leeds, UK, 1974. [Google Scholar]
- Princewell, T.J.T.; Agba, M.I. Examination of bovine faeces for the isolation and identification of Clostridium species. J. Appl. Bacteriol. 1982, 52, 97–102. [Google Scholar] [CrossRef]
- Nagy, J.; Bilkei, G. Neonatal piglet losses associated with Escherichia coli and Clostridium difficile infection in a Slovakian outdoor production unit. Vet. J. 2003, 166, 98–100. [Google Scholar] [CrossRef]
- Borriello, S.P.; Honour, P.; Turner, T.; Barclay, F. Household pets as a potential reservoir for Clostridium difficile infection. J. Clin. Pathol. 1983, 36, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Weber, A.; Kroth, P.; Heil, G. Domestic animals as excreters of Clostridium difficile. Dtsch. Med. Wochenschr. 1988, 113, 1617–1618. [Google Scholar]
- Goorhuis, A.; Debast, S.B.; van Leengoed, L.A.; Harmanus, C.; Notermans, D.W.; Bergwerff, A.A.; Kuijper, E.J. Clostridium difficile PCR ribotype 078: An emerging strain in humans and in pigs? J. Clin. Microbiol. 2008, 46, 1157. [Google Scholar] [CrossRef] [Green Version]
- Jhung, M.A.; Thompson, A.D.; Killgore, G.E.; Zukowski, W.E.; Songer, G.; Warny, M.; Johnson, S.; Gerding, D.N.; McDonald, L.C.; Limbago, B.M. Toxinotype V Clostridium difficile in humans and food animals. Emer. Infect. Dis. 2008, 14, 1039–1045. [Google Scholar] [CrossRef]
- Rupnik, M.; Widmer, A.; Zimmermann, O.; Eckert, C.; Barbut, F. Clostridium difficile toxinotype V, ribotype 078, in animals and humans. J. Clin. Microbiol. 2008, 46, 2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debast, S.B.; van Leengoed, L.A.; Goorhuis, A.; Harmanus, C.; Kuijper, E.J.; Bergwerff, A.A. Clostridium difficile PCR ribotype 078 toxinotype V found in diarrhoeal pigs identical to isolates from affected humans. Environ. Microbiol. 2009, 11, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Susick, E.K.; Putnam, M.; Bermudez, D.M.; Thakur, S. Longitudinal study comparing the dynamics of Clostridium difficile in conventional and antimicrobial free pigs at farm and slaughter. Vet. Microbiol. 2012, 157, 172–178. [Google Scholar] [CrossRef]
- Songer, J.G.; Trinh, H.T.; Killgore, G.E.; Thompson, A.D.; McDonald, L.C.; Limbago, B.M. Clostridium difficile in Retail Meat Products, USA, 2007. Emer. Infect. Dis. 2009, 15, 819–821. [Google Scholar] [CrossRef]
- Weese, J.; Avery, B.P.; Rousseau, J.; Reid-Smith, J. Detection and Enumeration of Clostridium difficile Spores in Retail Beef and Pork. Appl. Environ. Microbiol. 2009, 15, 5009–5011. [Google Scholar] [CrossRef] [Green Version]
- Bouttier, S.; Barc, M.C.; Felix, B.; Lambert, S.; Collignon, A.; Barbut, F. Clostridium difficile in ground meat, France. Emer. Infect. Dis. 2010, 16, 733–735. [Google Scholar] [CrossRef]
- de Boer, E.; Zwartkruis-Nahuis, A.; Heuvelink, A.E.; Harmanus, C.; Kuijper, E.J. Prevalence of Clostridium difficile in retailed meat in The Netherlands. Int. J. Food Microbiol. 2011, 144, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Gomez, C.; Mulvey, M.R.; Vargas, P.; Gamboa-Coronado Mde, l.M.; Rodriguez, C.; Rodriguez-Cavillini, E. Isolation of a toxigenic and clinical 25 genotype of Clostridium difficile in retail meats in Costa Rica. J. Food Pro. 2013, 76, 348–351. [Google Scholar] [CrossRef]
- Kouassi, K.A.; Dadie, A.T.; N’Guessan, K.F.; Dje, K.M.; Loukou, Y.G. Clostridium perfringens and Clostridium difficile in cooked beef sold in Côte d’Ivoire and their antimicrobial susceptibility. Anaerobe 2014, 28, 90–94. [Google Scholar] [CrossRef]
- Primavilla, S.; Farneti, S.; Petruzzelli, A.; Drigo, I.; Scuota, S. Contamination of hospital food with Clostridium difficile in Central Italy. Anaerobe 2019, 55, 8–10. [Google Scholar] [CrossRef]
- Miller, M.; Gravel, D.; Mulvey, M.; Taylor, G.; Boyd, D.; Simor, A.; Gardam, M.; McGeer, A.; Hutchinson, J.; Moore, D.; et al. Health Care-Associated Clostridium difficile Infection in Canada: Patient Age and Infecting Strain Type Are Highly Predictive of Severe Outcome and Mortality. Clin. Infect. Dis 2010, 50, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.P. Immune response to Clostridium difficile infection. Eur. J. Gastroenterol. Hepatol 1996, 8, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Ogra, P.L. Ageing and its possible impact on mucosal immune responses. Ageing Res. Rev. 2010, 9, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.A.; Stahl, T.J. Persistent and Recurrent Clostridium difficile Colitis. Clin. Colon Rectal Sur. 2015, 28, 65–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dharbhamulla, N.; Abdelhady, A.; Domadia, M.; Patel, S.; Gaughan, J.; Roy, S. Risk Factors Associated with Recurrent Clostridium difficile Infection. J. Clin. Med. Res. 2019, 11, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Song, J.H.; Kim, Y.S. Recurrent Clostridium difficile Infection: Risk Factors, Treatment, and Prevention. Gut Liver 2019, 13, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Britton, R.A.; Young, V.B. Role of the intestinal microbiota in resistance to colonization by Clostridium difficile. Gastroenterology 2014, 146, 1547–1553. [Google Scholar] [CrossRef] [Green Version]
- Min, J.H.; Kim, Y.S. Proton pump inhibitors should be used with caution in critically Ill patients to prevent the risk of Clostridium difficile infection. Gut Liver 2016, 10, 493–494. [Google Scholar] [CrossRef] [Green Version]
- Tariq, R.; Singh, S.; Gupta, A.; Pardi, D.S.; Khanna, S. Association of gastric acid suppression with recurrent Clostridium difficile infection: A systematic review and meta-analysis. JAMA Internal Med. 2017, 177, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; Nahass, R.G.; Brunetti, L. Acid suppression medications during hospitalization as a risk factor for recurrence of Clostridioides difficile infection: Systematic review and meta-analysis. Clin. Infect. Dis. 2021, 545, 1–7. [Google Scholar] [CrossRef]
- Surawicz, C.M.; Brandt, L.J.; Binion, D.G.; Ananthakrishnan, A.N.; Curry, S.R.; Gilligan, P.H.; McFarland, L.V.; Mellow, M.; Zuckerbraun, B.S. Guidelines for diagnosis, treatment, and prevention of Clostridium difficile infections. Am. J. Gastroenterol. 2013, 108, 478–498. [Google Scholar] [CrossRef] [PubMed]
- Louie, T.J.; Miller, M.A.; Mullane, K.M.; Weiss, K.; Lentnek, A.; Golan, Y.; Gorbach, S.; Sears, P.; Shue, Y.-K. Fidaxomicin versus vancomycin for Clostridium difficile infection. N. Eng. J. Med. 2011, 364, 422–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaparro-Rojas, F.; Mullane, K.M. Emerging therapies for Clostridium difficile infection—Focus on fidaxomicin. Infect. Drug Res. 2013, 6, 41–53. [Google Scholar]
- Iv, E.C.O.; Iii, E.C.O.; Johnson, D.A. Clinical update for the diagnosis and treatment of Clostridium difficile infection. World J. Gastro. Pharmacol. Ther. 2014, 5, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Dyer, C.; Hutt, L.P.; Burky, R.; Joshi, L.T. Biocide Resistance and Transmission of Clostridium difficile Spores Spiked onto Clinical Surfaces from an American Health Care Facility. Appl. Environ. Microbiol. 2019, 85, e01090-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depestel, D.D.; Aronoff, D.M. Epidemiology of Clostridium difficile infection. J. Pharm. Pract. 2013, 26, 464–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbut, F.; Petit, J.C. Epidemiology of Clostridium difficile-associated infections. Clin. Microbiol. Infect. 2001, 7, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Akerlund, T.; Svenungsson, B.; Lagergren, A.; Burman, L.G. Correlation of disease severity with fecal toxin levels in patients with Clostridium difficile-associated diarrhea and distribution of PCR ribotypes and toxin yields in vitro of corresponding isolates. J. Clin. Microbiol. 2006, 44, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.T.; Safdar, N. Current trends in the epidemiology and outcomes of Clostridium difficile infection. Clin Infect Dis. 2015, 60 (Suppl. S2), S66–S71. [Google Scholar] [CrossRef] [Green Version]
- Czepiel, J.; Krutova, M.; Mizrahi, A.; Khanafer, N.; Enoch, D.A.; Patyi, M.; Deptuła, A.; Agodi, A.; Nuvials, X.; Pituch, H.; et al. Mortality Following Clostridioides difficile Infection in Europe: A Retrospective Multicenter Case-Control Study. Antibiotic 2021, 10, 299. [Google Scholar] [CrossRef]
- Voth, D.; Ballard, J.D. Clostridium difficile Toxins: Mechanism of Action and Role in Disease. Clin. Microbiol. Rev. 2005, 2, 247–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupnik, M.; Janezic, S. An update on Clostridium difficile toxinotyping. J. Clin. Microbiol. 2016, 54, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundsberger, T.; Braun, V.; Weidmann, M.; Leuke, P.; Sauerborn, M.; von Eichel-Streiber, C. Transcription analysis of the genes tcdA-E of the pathogenicity locus of Clostridium difficile. Eur. J. Biochem. 1997, 244, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Matamouros, S.; England, P.; Dupuy, B. Clostridium difficile toxin expression is inhibited by the novel regulator TcdC. Mol. Microbiol. 2007, 64, 1274–1288. [Google Scholar] [CrossRef] [PubMed]
- Donta, S.T.; Sullivan, N.; Wilkins, T.D. Differential effects of Clostridium difficile toxins on tissue-cultured cells. J. Clin. Microbiol. 1982, 15, 1157–1158. [Google Scholar] [CrossRef] [Green Version]
- Feltis, B.A.; Wiesner, S.M.; Kim, A.S.; Erlandsen, S.L.; Lyerly, D.L.; Wilkins, T.D.; Wells, C.L. Clostridium difficile toxins A and B can alter epithelial permeability and promote bacterial paracellular migration through HT-29 enterocytes. Shock 2000, 14, 629–634. [Google Scholar] [CrossRef]
- Riggs, M.M.; Sethi, A.K.; Zabarsky, T.F.; Eckstein, E.C.; Jump, R.L.P.; Donskey, C.J. Asymptomatic carriers are a potential source for transmission of epidemic and nonepidemic Clostridium difficile strains among long-term care facility residents. Clin. Infect. Dis. 2007, 45, 992–998. [Google Scholar] [CrossRef] [Green Version]
- Stiles, B.G.; Wigelsworth, D.J.; Popoff, M.R.; Barth, H. Clostridial binary toxins: Iota and c2 family portraits. Front. Cell. Infect. Microbiol. 2011, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Gerding, D.; Johnson, S.; Rupnik, M.; Aktories, K. Clostridium difficile binary toxin CDT: Mechanism, epidemiology, and potential clinical importance. Gut Microbes 2014, 5, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Aktories, K.; Papatheodorou, P.; Schwan, C. Binary Clostridium difficile toxin (CDT)—A virulence factor disturbing the cytoskeleton. Anaerobe 2018, 53, 21–29. [Google Scholar] [CrossRef]
- Kuehne, S.A.; Collery, M.M.; Kelly, M.L.; Cartman, S.T.; Cockayne, A.; Minton, N.P. Importance of toxin A, toxin B, and CDT in virulence of an epidemic Clostridium difficile strain. J. Infect. Dis. 2014, 209, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.; Cheek, L.; Satta, G.; Walker-Bone, K.; Cubbon, M.; Citron, D. Predictors of death after Clostridium difficile infection: A report on 128 strain-typed cases from a teaching hospital in the United Kingdom. Clin. Infect. Dis. 2010, 50, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldenberg, S.D.; French, G.L. Lack of association of tcdC type and binary toxin status with disease severity and outcome in toxigenic Clostridium Difficile. J. Infect. 2011, 62, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, S.; Ascenzi, P.; Siarakas, S.; Petrosillo, N.; di Masi, A. Clostridium difficile Toxins A and B: Insights into Pathogenic Properties and Extraintestinal Effects. Toxins 2016, 8, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, T.; Kaibuchi, K.; Yamamoto, T.; Kawamura, M.; Sakoda, T.; Fujioka, H.; Matsuura, Y.; Takai, Y. A stimulatory GDP/GTP exchange protein for smg p21 is active on the post-translationally processed form of c-Ki-ras p21 and rhoA p21. Proc. Natl. Acad. Sci. USA 1991, 88, 6442–6446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, N.; Dupuy, B. Regulation of toxin synthesis in Clostridium difficile by an alternative RNA polymerase sigma factor. Proc. Natl. Acad. Sci. USA 2001, 98, 5844–5849. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.S.; Wee, B.Y.; Song, K.P. Evidence for holin function of tcdE gene in the pathogenicity of Clostridium difficile. J. Med. Microbiol. 2001, 50, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Govind, R.; Dupuy, B. Secretion of Clostridium difficile toxins A and B requires the holin-like protein TcdE. PLoS Pathog. 2012, 8, e1002727. [Google Scholar] [CrossRef]
- Dayananda, P.; Wilcox, M.H. A Review of Mixed Strain Clostridium difficile colonization and infection. Front. Microbiol. 2019, 10, 692. [Google Scholar] [CrossRef] [Green Version]
- Azimirad, M.; Krutova, M.; Yadegar, A.; Shahrokh, S.; Olfatifar, M.; Aghdaei, H.A.; Fawley, W.N.; Wilcox, M.H.; Zali, M.R. Clostridioides difficile ribotypes 001 and 126 were predominant in Tehran healthcare settings from 2004 to 2018: A 14-year-long cross-sectional study. Emerg. Microbes Infect. 2020, 1, 1432–1443. [Google Scholar] [CrossRef]
- Neely, F.; Lambert, M.L.; Van Broeck, J.; Delmée, M. Clinical and laboratory features of the most common Clostridium difficile ribotypes isolated in Belgium. J. Hosp. Infect. 2017, 4, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Stein, K.; Egan, S.; Lynch, H.; Harmanus, C.; Kyne, L.; Herra, C.; McDermott, S.; Kuijper, E.; Fitzpatrick, F.; FitzGerald, S.; et al. PCR-ribotype distribution of Clostridium difficile in Irish pigs. Anaerobe 2017, 48, 237–241. [Google Scholar] [CrossRef]
- Kuijper, E.J.; Coignard, B.; Tüll, P. Emergence of Clostridium difficile-associated disease in North America and Europe. Clinical microbiology and infection: The official publication of the European Society of Clinical Microbiology and Infectious Diseases. Clin. Microbiol. Infect. 2006, 6, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hookman, P.; Barkin, J.S. Clostridium difficile associated infection, diarrhea and colitis. World J. Gastroenterol. 2009, 15, 1554–1580. [Google Scholar] [CrossRef] [PubMed]
- Janezic, S.; Garneau, J.R.; Monot, M. Comparative Genomics of Clostridium difficile. Adv. Exp. Med. Biol. 2018, 1050, 59–75. [Google Scholar] [PubMed]
- Jones, A.M.; Kuijper, E.J.; Wilcox, M.H. Clostridium difficile: A European perspective. J. Infect. 2013, 66, 115–128. [Google Scholar] [CrossRef]
- Baldan, R.; Trovato, A.; Bianchini, V.; Biancardi, A.; Cichero, P.; Mazzotti, M.; Nizzero, P.; Moro, M.; Ossi, C.; Scarpellini, P.; et al. Clostridium difficile PCR ribotype 018, a successful epidemic genotype. J. Clin. Microbiol. 2015, 53, 2575–2580. [Google Scholar] [CrossRef] [Green Version]
- Knetsch, C.W.; Hensgens, M.P.M.; Harmanus, C.; van der Bijl, M.W.; Savelkoul, P.H.M.; Kuijper, E.J.; Corver, J.; van Leeuwen, H.C. Genetic markers for Clostridium difficile lineages linked to hypervirulence. Microbiology 2011, 157, 3113–3123. [Google Scholar] [CrossRef] [Green Version]
- Schneeberg, A.; Neubauer, H.; Schmoock, G.; Baier, S.; Harlizius, J.; Nienhoff, H.; Brase, K.; Zimmermann, S.; Seyboldt, C. Clostridium difficile genotypes in piglet populations in Germany. J. Clin. Microbiol. 2013, 51, 3796–3803. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Pérez, S.; Blanco, J.L.; Harmanus, C.; Kuijper, E.; Garcia, M.E. Subtyping and antimicrobial susceptibility of Clostridium difficile PCR ribotype 078/126 isolates of human and animal origin. Vet. Microbiol. 2017, 199, 15–22. [Google Scholar] [CrossRef]
- Cairns, M.D.; Stabler, R.A.; Shetty, N.; Wren, B.W. The continually evolving Clostridium difficile species. Future Microbiol. 2012, 7, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Cairns, M.D.; Preston, M.D.; Lawley, T.D.; Clark, T.G.; Stabler, R.A.; Wren, B.W. Genomic epidemiology of a protracted hospital outbreak caused by a toxin A-negative Clostridium difficile sublineage PCR ribotype 017 strain in London, England. J. Clin. Microbiol. 2015, 53, 3141–3147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, D.A.; Hawkey, P.M.; Riley, T.V. Epidemiology of Clostridium difficile infection in Asia. Antimicrob. Resist. Infect. Control. 2013, 2, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spigaglia, P.; Barbanti, F.; Dionisi, A.M.; Mastrantonio, P. Clostridium difficile isolates resistant to fluoroquinolones in Italy: Emergence of PCR ribotype 018. J. Clin. Microbiol. 2010, 48, 2892–2896. [Google Scholar] [CrossRef] [Green Version]
- Bauer, M.P.; Notermans, D.W.; van Benthem, B.H.; Brazier, J.S.; Wilcox, M.H.; Rupnik, M.; Monnet, D.L.; van Dissel, J.T.; Kuijper, E.J. Clostridium difficile infection in Europe: A hospital-based survey. Lancet 2011, 377, 63–73. [Google Scholar] [CrossRef]
- Barbanti, F.; Spigaglia, P. Characterization of Clostridium difficile PCR-ribotype 018: A problematic emerging type. Anaerobe 2016, 42, 123–129. [Google Scholar] [CrossRef]
- Zidaric, V.; Beigot, S.; Lapajne, S.; Rupnik, M. The occurrence and high diversity of Clostridium difficile genotypes in rivers. Anaerobe 2010, 16, 371–375. [Google Scholar] [CrossRef]
- Bazaid, F. Distribution and Sources of Clostridium difficile Present in Water Sources; MSc University of Guelph: Guelph, Canada, 2012. [Google Scholar]
- Warriner, K.; Xu, C.; Habash, M.; Sultan, S.; Weese, S.J. Dissemination of Clostridium difficile in food and the environment: Significant sources of C. difficile community acquired infection? J. Appl. Microbiol. 2016, 122, 542–555. [Google Scholar] [CrossRef]
- Moradigaravand, D.; Gouliouris, T.; Ludden, C.; Reuter, S.; Jamrozy, D.; Blane, B.; Naydenova, P.; Judge, K.H.; Aliyu, S.F.; Hadjirin, N.A.; et al. Genomic survey of Clostridium difficile reservoirs in the East of England implicates environmental contamination of wastewater treatment plants by clinical lineages. Microb. Genom. 2018, 4, e000162. [Google Scholar] [CrossRef] [Green Version]
- Kotila, S.M.; Pitkänen, T.; Brazier, J.; Eerola, E.; Jalava, J.; Kuusi, M.; Könönen, E.; Laine, J.; Miettinen, I.T.; Vuento, R.; et al. Clostridium difficile contamination of public tap water distribution system during a waterborne outbreak in Finland. Scand. J. Public Health 2013, 41, 541–545. [Google Scholar] [CrossRef]
- Al Saif, N.; Brazier, J.S. The distribution of Clostridium difficile in the environment of South Wales. J. Med. Microbiol. 1996, 45, 133–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baverud, V.; Gustafsson, A.; Franklin, A.; Aspan, A.; Gunnarsson, A. Clostridium difficile: Prevalence in horses and environment, and antimicrobial susceptibility. Equine Vet. J. 2003, 35, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Higazi, T.B.; AL-Saghir, M.; Burkett, M.; Pusok, R. PCR detection of Clostridium difficile and its toxigenic strains in public places in Southeast Ohio. Int. J. Microbiol. Res. 2011, 2, 105–111. [Google Scholar]
- Rodriguez, C.; Bouchafa, L.; Soumillion, K.; Ngyuvula, E.; Taminiau, B.; VanBroeck, J.; Delmée, M.; Daube, G. Seasonality of Clostridium difficile in the natural environment. Transbound. Emerg. Dis. 2019, 66, 2440–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dharmasena, M.; Jiang, X. Isolation of toxigenic Clostridium difficile from animal manure and composts being used as biological soil amendments. Appl. Environ. Microbiol. 2018, 84, e00738-18. [Google Scholar] [CrossRef] [Green Version]
- Shivaperumal, N.; Chang, B.J.; Riley, T.V. High Prevalence of Clostridium difficile in Home Gardens in Western Australia. Appl. Environ. Microbiol. J. 2020, 87, e01572-20. [Google Scholar] [CrossRef]
- Fröschle, B.; Messelhäusser, U.; Höller, C.; Lebuhn, M. Fate of Clostridium botulinum and incidence of pathogenic clostridia in biogas processes. J. Appl. Microbiol. 2015, 119, 936–947. [Google Scholar] [CrossRef]
- Marcos, P.; Whyte, P.; Rogers, T.; McElroy, M.; Fanning, S.; Frias, J.; Bolton, D. The prevalence of Clostridioides difficile on farms, in abattoirs and in retail foods in Ireland. Food Microbiol. 2021, 98, 103781. [Google Scholar] [CrossRef]
- Rodriguez, C.; Taminiau, B.; Van Broeck, J.; Avesani, V.; Delmee, M.; Daube, G. Clostridium difficile in young farm animals and slaughter animals in Belgium. Anaerobe 2012, 18, 621–625. [Google Scholar] [CrossRef]
- Wu, Y.C.; Lee, J.J.; Tsai, B.Y.; Liu, Y.F.; Chen, C.M.; Tien, N.; Tsai, P.J.; Chen, T.H. Potentially hypervirulent Clostridium difficile PCR ribotype 078 lineage isolates in pigs and possible implications for humans in Taiwan. Int. J. Med. Microbiol. 2016, 306, 115–122. [Google Scholar] [CrossRef]
- Rodriguez, C.; Hakimi, D.E.; Vanleyssem, R.; Taminiau, B.; Van Broeck, J.; Delmee, M.; Korsak, N.; Daube, G. Clostridium difficile in beef cattle farms, farmers and their environment: Assessing the spread of the bacterium. Vet. Microbiol. 2017, 210, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Cho, A.; Kim, J.W.; Kim, H.; Kim, B. High prevalence of Clostridium difficile PCR ribotype 078 in pigs in Korea. Anaerobe 2018, 51, 42–46. [Google Scholar] [CrossRef]
- Krutova, M.; Zouharova, M.; Matejkova, J.; Tkadlec, J.; Krejci, J.; Faldyna, M.; Nyc, O.; Bernardy, J. The emergence of Clostridium difficile PCR ribotype 078 in piglets in the Czech Republic clusters with Clostridium difficile PCR ribotype 078 isolates from Germany, Japan and Taiwan. Int. J. Med. Microbiol. 2018, 308, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Taminiau, B.; van Broeck, J.; Daube, G. Clostridium difficile in Food and Animals: A Comprehensive Review. Adv. Exp. Med. Biol. 2016, 932, 65–92. [Google Scholar] [PubMed] [Green Version]
- Keel, K.; Brazier, J.S.; Post, K.W.; Weese, S.; Songer, J.G. Prevalence of PCR Ribotypes among Clostridium difficile Isolates from Pigs, Calves, and Other Species. J. Clin. Microbiol. 2007, 6, 1963–1964. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Palacios, A.; Staempfli, H.R.; Duffield, T.; Weese, J.S. Clostridium difficile in Retail Ground Meat, Canada. Emerg. Infect. Dis. 2007, 3, 485–487. [Google Scholar] [CrossRef]
- Knight, D.R.; Riley, T.V. Prevalence of Gastrointestinal Clostridium difficile Carriage in Australian Sheep and Lambs. Appl. Environ. Microbiol. 2013, 79, 5689–5692. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.; Norman, K.; Andrews, K.; Hume, M.; Scanlan, C.; Callaway, T. Clostridium difficile in poultry and poultry meat. Foodborne Path. Dis. 2011, 8, 1321–1323. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Glil, M.Y.; Thomas, P.; Schmoock, G.; Abou-El-Azm, K.; Wieler, L.H.; Neubauer, H.; Seyboldt, C. Presence of Clostridium difficile in poultry and poultry meat in Egypt. Anaerobe 2018, 51, 21–25. [Google Scholar] [CrossRef]
- Hussain, I.; Borah, P.; Sharma, R.K.; Rajkhowa, S.; Rupnik, M.; Saikia, D.P. Molecular characteristics of Clostridium difficile isolates from human and animals in the North Eastern region of India. Mol. Cell Probes 2016, 30, 306–311. [Google Scholar] [CrossRef]
- Simango, C.; Mwakurudza, S. Clostridium difficile in broiler chickens sold at market places in Zimbabwe and their antimicrobial susceptibility. Int. J. Food Microbiol. 2008, 124, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Zidaric, V.; Zemljic, M.; Janezic, S.; Kocuvan, A.; Rupnik, M. High diversity of Clostridium difficile genotypes isolated from a single poultry farm producing replacement laying hens. Anaerobe 2008, 14, 325–327. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion on the public health hazards to be covered by inspection of meat (bovine animals). Eur. Food Saf. Aut. J. 2013, 11, 1–261. [Google Scholar]
- Rodriguez, C.; Avesani, V.; Van Broeck, J.; Taminiau, B.; Delmée, M.; Daube, G. Presence of Clostridium difficile in pigs and cattle intestinal contents and carcass contamination at the slaughterhouse in Belgium. Int. J. Food Microbiol. 2013, 166, 256–262. [Google Scholar] [CrossRef]
- Hopman, N.E.; Oorburg, D.; Sanders, I.; Kuijper, E.J.; Lipman, L.J. High occurrence of various Clostridium difficile PCR ribotypes in pigs arriving at the slaughterhouse. Vet. Quart. 2011, 31, 179–181. [Google Scholar] [CrossRef] [Green Version]
- Hawken, P.; Weese, J.S.; Friendship, R.; Warriner, K. Carriage and dissemination of Clostridium difficile and methicillin resistant Staphylococcus aureus in pork processing. Food Control 2013, 31, 433–437. [Google Scholar] [CrossRef]
- Wu, Y.C.; Chen, C.M.; Kuo, C.J.; Lee, J.J.; Chen, P.C.; Chang, Y.C. Prevalence and molecular characterization of Clostridium difficile isolates from a pig slaughterhouse, pork, and humans in Taiwan. Int. J. Food Microbiol. 2013, 242, 37–44. [Google Scholar] [CrossRef]
- Hampikyan, H.; Bingol, E.B.; Muratoglu, K.; Akkaya, E.; Cetin, O.; Colak, H. The prevalence of Clostridium difficile in cattle and sheep carcasses and the antibiotic susceptibility of isolates. Meat Sci. 2018, 139, 120–124. [Google Scholar] [CrossRef]
- Esfandiari, Z.; Weese, J.S.; Ezzatpanah, H.; Chamani, M.; Shoaei, P.; Yaran, M.; Ataei, B.; Maracy, M.R.; Ansariyan, A.; Ebrahimi, F.; et al. Isolation and characterization of Clostridium difficile in farm animals from slaughterhouse to retail stage in Isfahan, Iran. Food. Path. Dis. 2015, 10, 864–866. [Google Scholar] [CrossRef]
- Candel-Pérez, C.; Santaella-Pascual, J.; Ros-Berruezo, G.; Martínez-Gracia, C. Occurrence of Clostridioides [Clostridium] difficile in poultry giblets at slaughter and retail pork and poultry meat in southeastern Spain. J. Food Prot. 2021, 84, 310–314. [Google Scholar] [CrossRef]
- Keessen, E.C.; van den Berkt, A.J.; Haasjes, N.H.; Hermanus, C.; Kuijper, E.J.; Lipman, L.J. The relation between farm specific factors and prevalence of Clostridium difficile in slaughter pigs. Vet. Microbiol. 2011, 54, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Lund, B.M.; Peck, M.W. A possible route for foodborne transmission of Clostridium difficile? Food. Path. Dis. 2015, 12, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, R.; Norman, K.; Andrews, K.; Norby, B.; Hume, M.; Scanlan, C.; Hardin, M.; Scott, H. Clostridium difficile in retail meat and processing plants in Texas. J. Vet. Diag. Investig. 2011, 23, 807–811. [Google Scholar] [CrossRef] [Green Version]
- Varshney, J.B.; Very, K.J.; Williams, J.L.; Hegarty, J.P.; Stewart, D.B.; Lumadue, J. Characterization of Clostridium difficile isolates from human fecal samples and retail meat from Pennsylvania. Food. Path. Dis. 2014, 11, 822–829. [Google Scholar] [CrossRef]
- Guran, H.S.; Ilhak, O.I. Clostridium difficile in retail chicken meat parts and liver in the Eastern Region of Turkey. J. Con. Prot. Food Saf. 2015, 10, 359–364. [Google Scholar] [CrossRef]
- Metcalf, D.; Avery, B.P.; Janecko, N.; Matic, N.; Reid-Smith, R.; Weese, J.S. Clostridium difficile in seafood and fish. Anaerobe 2011, 17, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Pasquale, V.; Romano, V.; Rupnik, M.; Capuano, F.; Bove, D.; Aliberti, F.; Krovacek, K.; Dumontet, S. Occurrence of toxigenic Clostridium difficile in edible bivalve molluscs. Food Microbiol. 2012, 31, 309–312. [Google Scholar] [CrossRef]
- Troiano, T.; Harmanus, C.; Sanders, I.; Pasquale, V.; Dumontet, S.; Capuano, F.; Romano, V.; Kuijper, E.J. Toxigenic Clostridium difficile PCR ribotypes in ediblemarine bivalve molluscs in Italy. Int. J. Food Microbiol. 2015, 208, 30–34. [Google Scholar] [CrossRef]
- Agnoletti, F.; Arcangelia, G.; Barbantib, F.; Barcoa, L.; Brunettaa, R.; Cocchia, M.; Conederaa, G.; D’Estea, L.; Drigoa, I.; Spigagliab, P.; et al. Survey, characterization and antimicrobial susceptibility of Clostridium difficile from marine bivalve shellfish of North Adriatic Sea. Int. J. Food Microbiol. 2019, 298, 74–80. [Google Scholar] [CrossRef]
- Eckert, C.; Burghoffer, B.; Barbut, F. Contamination of ready-to-eat raw vegetables with Clostridium difficile, France. J. Med. Microbiol. 2013, 62, 1435–1438. [Google Scholar] [CrossRef]
- Lim, S.C.; Foster, N.F.; Elliott, B.; Riley, T.V. High prevalence of Clostridium difficile on retail root vegetables, Western Australia. J. Appl. Microbiol. 2018, 124, 585–590. [Google Scholar] [CrossRef]
- Tkalec, V.; Janezica, S.; Skoka, B.; Simonica, T.; Mesarica, S.; Vrabica, T.; Rupnik, M. High Clostridium difficile contamination rates of domestic and imported potatoes compared to some other vegetables in Slovenia. Food Microbiol. 2019, 78, 194–200. [Google Scholar] [CrossRef]
- Marcos, P.; Whyte, P.; Burgess, C.; Ekhlas, D.; Bolton, D. Detection and Genomic Characterisation of Clostridioides difficile from Spinach Fields. Pathogens 2022, 11, 1310. [Google Scholar] [CrossRef]
- Bakri, M.M.; Brown, D.J.; Butcher, J.P.; Sutherland, A.D. Clostridium difficile in ready-to-eat salads, Scotland. Emerg. Infect. Dis. 2009, 15, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Dharmasena, M.; Wang, H.; Wei, T.; Bridges, W.C., Jr.; Jiang, X. Survival of Clostridioides difficile in finished dairy compost under controlled conditions. J. Appl. Microbiol. 2021, 131, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Weese, J.S.; Flemming, C.; Odumeru, J.; Warriner, K. Fate of Clostridium difficile during wastewater treatment and incidence in Southern Ontario watersheds. J. Appl. Microbiol. 2014, 117, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Sun, M.; Zhang, Y.; Gao, J.; Kong, F.; Liu, D.; Yu, H.; Du, J.; Tang, R. Prevalence, genotype and antimicrobial resistance of Clostridium difficile isolates from healthy pets in Eastern China. BMC Infect. Dis. 2019, 19, 46. [Google Scholar] [CrossRef] [Green Version]
- Weese, J.S. Clostridium (Clostridioides) difficile in animals. J. Vet. Diagn.Investig. Off. Publ. Am. Assoc. Vet. Lab. Diag. 2020, 32, 213–221. [Google Scholar] [CrossRef]
- Hammitt, M.C.; Bueschel, D.M.; Keel, M.K.; Glock, R.D.; Cuneo, P.; DeYoung, D.W.; Reggiardo, C.; Trinh, H.T.; Songer, J.G. A possible role for Clostridium difficile in the etiology of calf enteritis. Vet. Microbiol. 2008, 127, 343–352. [Google Scholar] [CrossRef]
- Costa, M.C.; Stämpfli, H.R.; Arroyo, L.G.; Pearl, D.L.; Weese, J.S. Epidemiology of Clostridium difficile on a veal farm: Prevalence, molecular characterization and tetracycline resistance. Vet. Microbiol. 2011, 152, 379–384. [Google Scholar] [CrossRef]
- Costa, M.C.; Reid-Smith, R.; Gow, S.; Hannon, S.J.; Booker, C.; Rousseau, J.; Benedict, K.M.; Morley, P.S.; Weese, J.S. Prevalence and molecular characterization of Clostridium difficile isolated from feedlot beef cattle upon arrival and mid-feeding period. BMC Vet. Res. 2012, 8, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keessen, E.C.; Harmanus, C.; Dohmen, W.; Kuijper, E.J.; Lipman, L.J. Clostridium difficile infection associated with pig farms. Emerg. Infect. Dis. 2013, 19, 1032–1034. [Google Scholar] [CrossRef] [PubMed]
- Magistrali, C.F.; Maresca, C.; Cucco, L.; Bano, L.; Drigo, I.; Filippini, G.; Dettori, A.; Broccatelli, S.; Pezzotti, G. Prevalence and risk factors associated with Clostridium difficile shedding in veal calves in Italy. Anaerobe 2015, 33, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Romano, V.; Albanese, F.; Dumontet, S.; Krovacek, K.; Petrini, O.; Pasquale, V. Prevalence and genotypic characterization of Clostridium difficile from ruminants in Switzerland. Zoo. Pub. Health 2012, 59, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Larson, H.E.; Price, A.B.; Honour, P.; Borriello, S.P. Clostridium difficile and the aetiology of pseudomembranous colitis. Lancet 1978, 1, 1063–1066. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.M.; Shanholtzer, C.J.; Lee, J.T., Jr.; Gerding, D.N. Ten years of prospective Clostridium difficile-associated disease surveillance and treatment at the Minneapolis VA Medical Center, 1982–1991. Infect. Control Hosp. Epidemiol. 1994, 15, 371–381. [Google Scholar] [CrossRef]
- Slimings, C.; Riley, T. Antibiotics and hospital-acquired Clostridium difficile infection: Update of systematic review and meta-analysis. J. Antimicrob. Chemother. 2014, 69, 881–891. [Google Scholar] [CrossRef]
- Freeman, J.; Vernon, J.; Morris, K.; Nicholson, S.; Todhunter, S.; Longshaw, C.; Wilcox, M.H. Pan-European Longitudinal Surveillance of Antibiotic Resistance among Prevalent Clostridium difficile Ribotypes’ Study Group. Clin. Microbiol. Infect. 2015, 21, 248.e9–248.e16. [Google Scholar] [CrossRef] [Green Version]
- McDonald, L.C.; Coignard, B.; Dubberke, E.; Song, X.; Horan, T.; Kutty, P.K.; Ad Hoc Clostridium difficile Surveillance Working Group. Recommendations for surveillance of Clostridium difficile-associated disease. Infect. Control. Hosp. Epidemiol. 2007, 28, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Lessa, F.C.; Mu, Y.; Bamberg, W.M.; Beldavs, Z.G.; Dumyati, G.K.; Dunn, J.R.; Farley, M.M.; Holzbauer, S.M.; Meek, J.I.; Phipps, E.C.; et al. Burden of Clostridium difficile infection in the United States. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Khanna, S. Microbiota restoration for recurrent Clostridioides difficile: Getting one step closer every day! J. Intern. Med. 2021, 290, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, J.L.; Chrischilles, E.A.; Pendergast, J.F.; Herwaldt, L.A.; Polgreen, P.M. Incidence of and risk factors for community-associated Clostridium difficile infection: A nested case-control study. BMC Infect. Dis. 2011, 11, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moono, P.; Lim, S.C.; Riley, T.V. High prevalence of toxigenic Clostridium difficile in public space lawns in Western Australia. Sci. Rep. 2017, 7, 41196. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez Diaz, C.; Seyboldt, C.; Rupnik, M. Non-human C. difficile reservoirs and sources: Animals, food, environment. Adv. Exp. Med. Biol. 2018, 1050, 227–243. [Google Scholar]
- Avberšek, J.; Pirš, T.; Pate, M.; Rupnik, M.; Ocepek, M. Clostridium difficile in goats and sheep in Slovenia: Characterisation of strains and evidence of age-related shedding. Anaerobe 2014, 28, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Palacios, A.; Barman, T.; LeJeune, J.T. Three-week summer period prevalence of Clostridium difficile in farm animals in a temperate region of the United States (Ohio). Can. Vet. J. 2014, 55, 786–789. [Google Scholar] [PubMed]
- Cataldo, M.A.; Granata, G.; Petrosillo, N. Clostridium difficile infection: New approaches to prevention, non-antimicrobial treatment, and stewardship. Expert Rev. Anti-Infect. Ther. 2017, 15, 1027–1040. [Google Scholar] [CrossRef]
- Maron, D.F.; Smith, T.J.; Nachman, K.E. Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Glob. Health 2013, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Squire, M.M.; Riley, T.V. Clostridium difficile infection in humans and piglets: A ‘One Health’ opportunity. Curr. Top. Microbiol. Immunol. 2013, 365, 299–314. [Google Scholar]
- Deng, K.; Plaza-Garrido, A.; Torres, J.A.; Paredes-Sabja, D. Survival of Clostridium difficile spores at low temperatures. Food Microbiol. 2015, 46, 218–221. [Google Scholar] [CrossRef]
- Flock, G.; Chen, C.-H.; Yin, H.-B.; Fancher, S.; Mooyottu, S.; Venkitanarayanan, K. Effect of chilling, freezing and cooking on survivability of Clostridium difficile spores in ground beef. Meat Sci. 2016, 112, 161. [Google Scholar] [CrossRef]
- Rodriguez-Palacios, A.; Reid-Smith, R.J.; Staempfli, H.R.; Weese, J.S. Clostridium difficile survives minimal temperature recommended for cooking ground meats. Anaerobe 2010, 16, 540–542. [Google Scholar] [CrossRef] [PubMed]
- Ojha, S.C.; Chankhamhaengdecha, S.; Singhakaew, S.; Ounjai, P.; Janvilisri, T. Inactivation of Clostridium difficile spores by microwave irradiation. Anaerobe 2016, 38, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Doona, C.J.; Feeherry, F.E.; Setlow, B.; Wang, S.; Li, W.; Nichols, F.C. Effects of high-pressure treatment on spores of Clostridium species. Appl. Environ. Microbiol. 2016, 82, 5287–5297. [Google Scholar] [CrossRef] [Green Version]
- Deng, K.; Talukdar, P.K.; Sarker, M.R.; Paredes-Sabja, D.; Torres, J.A. Survival of Clostridium difficile spores at low water activity. Food Microbiol. 2017, 65, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Broda, D.M.; Delacy, K.M.; Bell, R.G.; Terry, J.; Cook, R.L. Psychrotrophic Clostridium spp. associated with ‘blown pack’ spoilage of chilled vacuum-packed red meats and dog rolls in gas-impermeable plastic casings. Int. J. Food Microbiol. 1996, 29, 335–352. [Google Scholar] [CrossRef]
- Atasoy, F.; Gücükoğlu, A. Detection of Clostridium difficile and toxin genes in samples of modified atmosphere packaged (MAP) minced and cubed beef meat. Ank. ÜNiversitesi Vet. Fakültesi Derg. 2017, 64, 165–170. [Google Scholar]
- Lim, S.C.; Foster, N.F.; Riley, T.V. Susceptibility of Clostridium difficile to the food preservatives sodium nitrite, sodium nitrate and sodium metabisulphite. Anaerobe 2016, 37, 67–71. [Google Scholar] [CrossRef]
- Le Lay, C.; Dridi, L.; Bergeron, M.G.; Ouellette, M.; Fliss, I. Nisin is an effective inhibitor of Clostridium difficile vegetative cells and spore germination. J. Med. Microbiol. 2016, 65, 169–175. [Google Scholar] [CrossRef]
- Aljarallah, K.M. Inhibition of Clostridium difficile by natural herbal extracts. J. Taibah Univ. Med. Sci. 2016, 11, 427–431. [Google Scholar] [CrossRef] [Green Version]
- Roshan, N.; Riley, T.V.; Hammer, K.A. Antimicrobial activity of natural products against Clostridium difficile in vitro. J. Appl. Microbiol. 2017, 123, 92–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Illustration of the C. difficile Pathogenicity locus (PaLoc). Adapted from [65].
Figure 1.
Illustration of the C. difficile Pathogenicity locus (PaLoc). Adapted from [65].

Figure 2.
Forest plot of the OR of C. difficile (all ribotypes) in each food type.

Figure 3.
Forest plot of the OR of C. difficile 027 and 078 in each food type.

Figure 4.
The cycle of community-associated CDI infections from zoonotic, environmental or foodborne sources. Adapted from [100] using ARASAAC pictograms.
Figure 4.
The cycle of community-associated CDI infections from zoonotic, environmental or foodborne sources. Adapted from [100] using ARASAAC pictograms.

Table 2.
Meat and seafood retail foods contaminated with C. difficile, including toxin gene profiles (toxins A, B and CDT) and ribotypes.
Table 2.
Meat and seafood retail foods contaminated with C. difficile, including toxin gene profiles (toxins A, B and CDT) and ribotypes.
| Product | Raw or RTE | Total No. (%) Positive | Toxin Gene Profile | Ribotype(s) | Reference |
|---|---|---|---|---|---|
| Ground pork | Raw | 3/7 (41.3%) | A+ B+ CDT+ | 027 078 | [35] |
| Ground pork | Raw | 14/115 (12%) | A+ B+ CDT+ | 027 078 | [36] |
| Ground pork | Raw | 2/66 (3.0%) | A+ B+ CDT− | 029 | [39] |
| Pork meat | Raw | 35/303 (11.5%) | A+ B+ CDT+ | 078 | [136] |
| Pork sausages | RTE | 10/16 (62.5%) | A+ B+ CDT+ | 027 078 | [35] |
| Ground beef | Raw | 13/26 (42.4%) | A+ B+ CDT+ | 027 078 | [35] |
| Ground beef | Raw | 11/53 (20.8%) | A+ B+ CDT+ | M31 | [118] |
| A+ B+ CDT− | 014 077 | ||||
| Ground beef | Raw | 14/115 (12%) | A+ B+ CDT+ | 027 078 | [36] |
| Ground beef | Raw | 2/105 (1.9%) | A+ B+ CDTND | 012 | [37] |
| Ground beef | Raw | 21/303 (6.9%) | A+ B+ CDT+ | PA22 | [136] |
| Beef | Raw | 1/67 (1.5%) | A+ B+ CDT− | 029 | [39] |
| Beef sausages | RTE | 1/7 (14.3%) | A+ B+ CDT+ | 027 | [35] |
| Corned beef | RTE | 1/10 (10%) | AND B+ CDTND | ND 1 | [110] |
| Ground veal | Raw | 1/7 (14.3%) | A+ B+ CDT+ | M31 | [118] |
| Turkey | Raw | 44/303 (14.5%) | A+ B+ CDT+ | PA01 PA05 PA16 | [136] |
| Ground turkey | Raw | 4/9 (44.4%) | A+ B+ CDT+ | 078 | [35] |
| Lamb | Raw | 1/16 (6.3%) | A+ B+ CDT+ | 045 | [38] |
| Chicken | Raw | 7/257 (2.7%) | A+ B+ CDT− | 001 003 071 087 | [38] |
| Chicken | Raw | 1/67 (1.5%) | A+ B+ CDT− | 029 | [39] |
| Chicken | Raw | 25/310 (8.0%) | A+ B+ CDT− | ND 1 | [137] |
| Chicken | Raw | 26/203 (12.8%) | A+ B+ CDT+ | 078 | [23] |
| Chicken | Raw | 24/303 (7.8%) | A+ B+ CDT+ | PA05 PA16 | [136] |
| Chicken | Raw | 4/32 (12.5%) | A+ B+ CDT+ | 078 | [110] |
| Chicken | RTE | 1/130 (0.8%) | A+ B+ CDT− | 014 020 | [41] |
| Shellfish | Raw | 118/702 (16.8%) | A+ B+ CDT+ | 126 475 | [141] |
| Bivalve molluscs | Raw | 26/53 (49%) | A+ B+ CDT+ | 078 | [139] |
| A+ B+ CDT− | 002 012 014/020 018 001 | ||||
| Bivalve molluscs | Raw | 36/925 (3.9%) | A+ B+ CDT+ | 078 126 010 | [140] |
| A− B+ CDT− | 017 001 |
+: Positive; −: Negative; 1 ND: Not determined.
Table 3.
Vegetable retail foods contaminated with C. difficile, including toxin gene profiles (toxins A, B and CDT) and ribotypes.
Table 3.
Vegetable retail foods contaminated with C. difficile, including toxin gene profiles (toxins A, B and CDT) and ribotypes.
| Product | Raw or RTE | Total No. (%) Positive | Toxin Gene Profile | Ribotype(s) | Reference |
|---|---|---|---|---|---|
| Root vegetables (potatoes, beetroots, onions and carrots) | Raw | 30/100 (30%) | A+ B+ CDT+ | QX 274 | [143] |
| A+ B+ CDT− | 002 137 QX519 QX049 101 | ||||
| A− B+ CDT+ | 070 237 584 | ||||
| A− B− CDT+ | 033 | ||||
| Root vegetables (potatoes, ginger) and leaf vegetables | Raw and RTE | 28/154 (18.2%) | A+ B+ CDT− | 001/072 011/049 014/020 012 070 150 394 SLO129 SLO187 SLO279 | [144] |
| A+B+CDT+ | 027 244 126 023 | ||||
| Lettuce | RTE | 1/54 (1.9%) | A+ B+ CDT+ | 126 | [41] |
| Vegetables (potato, onion, mushroom, carrot, radish and cucumber) | Raw | 7/300 (2.4%) | A+ BND CDT ND | ND 1 | [103] |
| Salad (lettuce, lamb’s lettuce) and vegetable (pea sprouts) | RTE | 3/104 (2.8%) | A+ B+ CDT− | 014/020 001 015 | [142] |
| Vegetables (carrots, potatoes, garlic, ginger, beets, mushrooms, lettuce, green onions, radishes, etc.) | Raw and RTE | 5/111 (4.5%) | A+ B+ CDT+ | 078 | [22] |
| A+ B+ CDT− | ND 1 | ||||
| Salad (baby leaf spinach) | RTE | 2/60 (3.3%) | A+ B+ CDT+ | 078 126 | [145] |
| Salad (baby leaf spinach, organic mixed leaf salad, organic lettuce) | RTE | 3/40 (7.5%) | A+ B+ CDT ND | 001 | [146] |
| A− B+ CDT ND | 017 | ||||
| Spinach leaves | RTE | 2/10 (20%) | A−B+ CDT− | ND 1 | [110] |
| Iceberg lettuce leaves | RTE | 1/10 (10%) | A−B+ CDT− | ND 1 | [110] |
| Little Gem lettuce leaves | RTE | 1/10 (10%) | A−B+ CDT− | ND 1 | [110] |
| Wild rocket leaves | RTE | 1/10 (10%) | A−B+ CDT+ | ND 1 | [110] |
| Coleslaw | RTE | 1/10 (10%) | A−B+ CDT− | ND 1 | [110] |
+: Positive; −: Negative; 1 ND: Not determined.
Table 4.
Further characterisation (pathogenicity, hypervirulence and association with community-acquired CDI) of the ribotypes isolated from foods (Table 2 and Table 3).
| Ribotype | Pathogenic | Hypervirulent | CA CDI 1 | Reference(s) | ||||
|---|---|---|---|---|---|---|---|---|
| yes | no | unk 2 | yes | no | unk | |||
| 001 | ✓ | ✓ | [81,99,147,148,149,150] | |||||
| 002 | ✓ | ✓ | ✓ | [81,99,148,149,151,152] | ||||
| 003 | ✓ | ✓ | ✓ | [81,99] | ||||
| 010 | ✓ | ✓ | [150] | |||||
| 011 | ✓ | ✓ | [148] | |||||
| 012 | ✓ | ✓ | ✓ | [81,148,149,150,153] | ||||
| 014 | ✓ | ✓ | ✓ | [81,99,148,151,153] | ||||
| 015 | ✓ | ✓ | [148,149,151] | |||||
| 017 | ✓ | ✓ | [145,148,149,154] | |||||
| 018 | ✓ | ✓ | [148,149] | |||||
| 020 | ✓ | ✓ | [148,149,151] | |||||
| 023 | ✓ | ✓ | [148,149,151] | |||||
| 027 | ✓ | ✓ | ✓ | [72,73,147,148,149,153,155] | ||||
| 029 | ✓ | ✓ | ✓ | [99,153] | ||||
| 070 | ✓ | ✓ | ✓ | [81,149] | ||||
| 071 | ✓ | ✓ | [149] | |||||
| 072 | ✓ | ✓ | [99,148,149,156] | |||||
| 077 | ✓ | ✓ | [149] | |||||
| 078 | ✓ | ✓ | ✓ | [72,73,148,149,153,154,155] | ||||
| 087 | ✓ | ✓ | [148,149] | |||||
| 101 | ✓ | ✓ | [149] | |||||
| 126 | ✓ | ✓ | ✓ | [6,80,81,99,149,153] | ||||
| 137 | ✓ | ✓ | [149] | |||||
| 150 | ✓ | ✓ | [149] | |||||
| 033, 045, 049, 237, 244, 394, 475, 584, M31, PA01, PA05, PA16, PA22, QX049, QX274, QX519, SLO129, SLO187, SLO279 | No information | |||||||
1 CA CDI = community acquired C. difficile infection; 2 unk = unknown.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bolton, D.; Marcos, P. The Environment, Farm Animals and Foods as Sources of Clostridioides difficile Infection in Humans. Foods 2023, 12, 1094. https://doi.org/10.3390/foods12051094
AMA Style
Bolton D, Marcos P. The Environment, Farm Animals and Foods as Sources of Clostridioides difficile Infection in Humans. Foods. 2023; 12(5):1094. https://doi.org/10.3390/foods12051094
Chicago/Turabian StyleBolton, Declan, and Pilar Marcos. 2023. "The Environment, Farm Animals and Foods as Sources of Clostridioides difficile Infection in Humans" Foods 12, no. 5: 1094. https://doi.org/10.3390/foods12051094
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.