
Preparation, Characterization and In Vitro Stability of a Novel ACE-Inhibitory Peptide from Soybean Protein
 , , , ,
, , , ,  , , , , , and
, , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
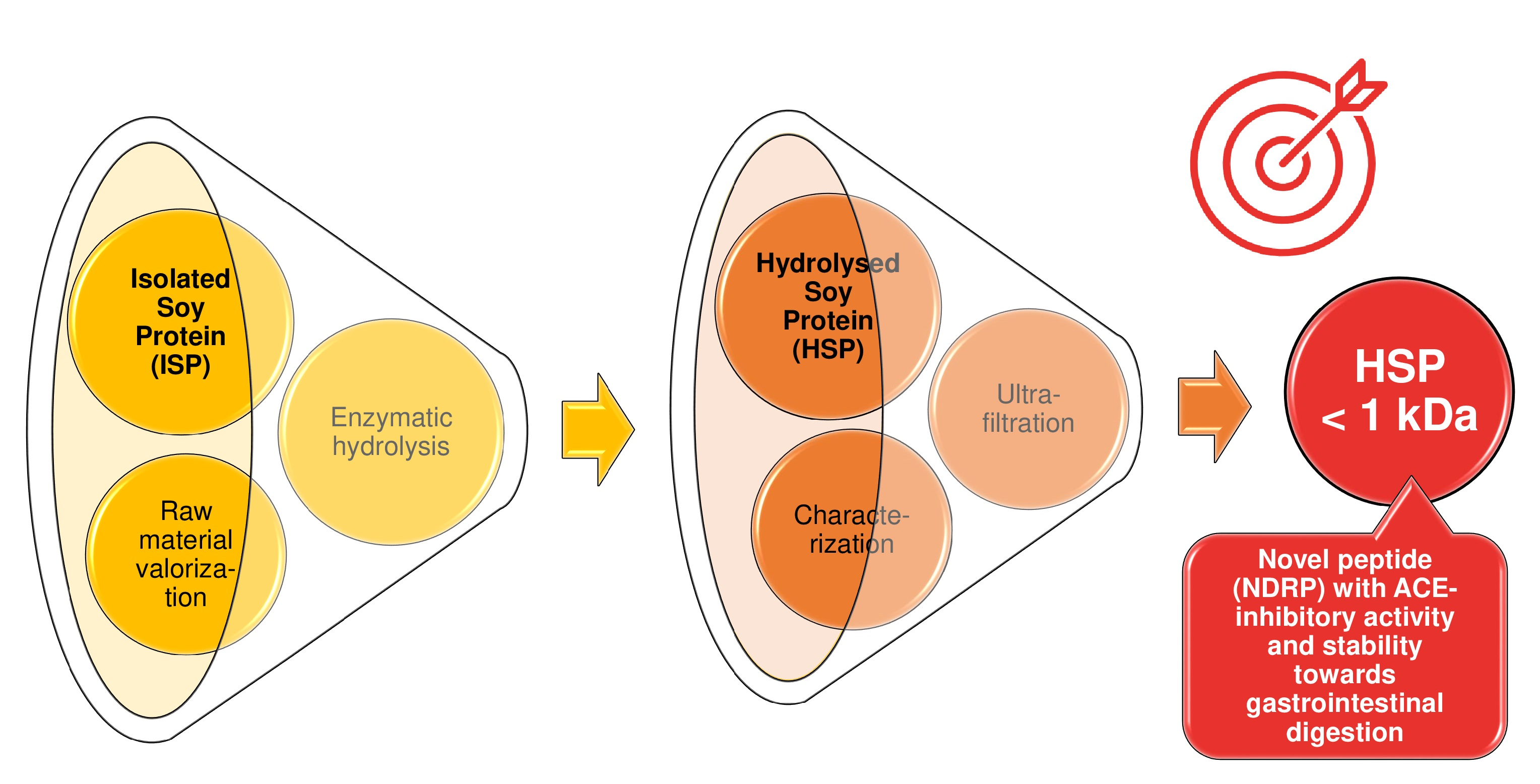
2.2. Hydrolysis of Soy Protein Isolate (SPI)
2.3. Protein and Hydrolysate Profiles
2.3.1. Degree of Hydrolysis (DH)
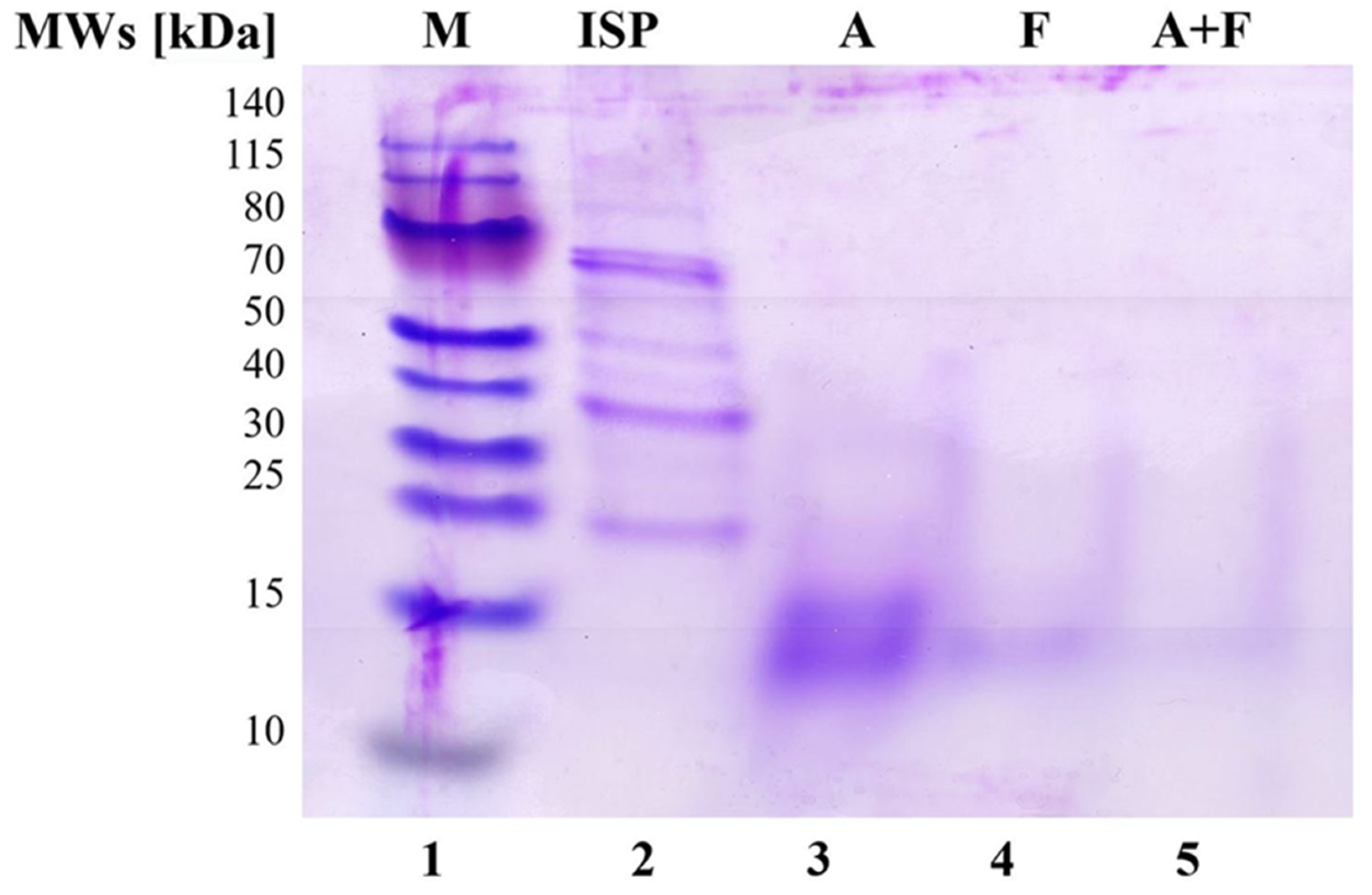
2.3.2. SDS-PAGE Analysis
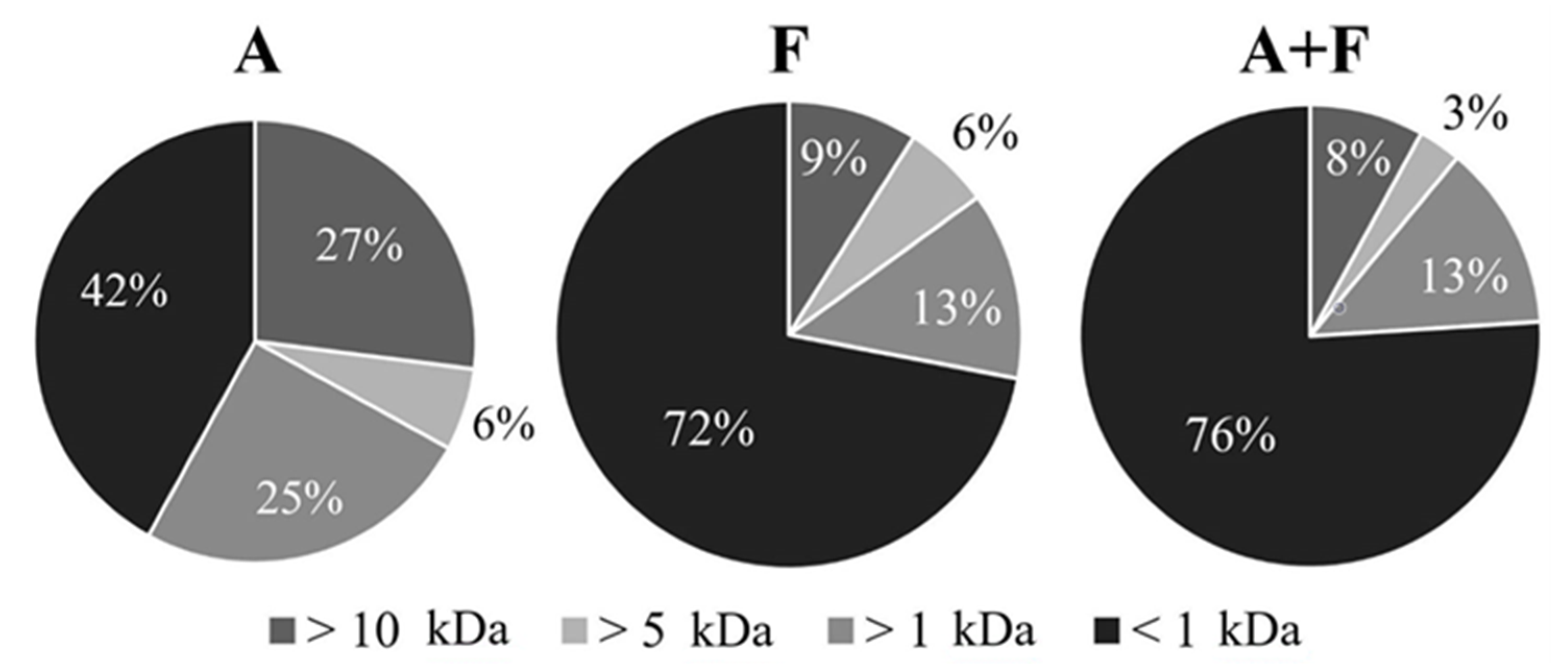
2.4. Ultrafiltration
2.5. Characterization of SPH < 1 kDa Fraction
2.5.1. ACE Inhibition Assay
2.5.2. Determination of ACE Inhibition Pattern
2.5.3. RP-HPLC
2.5.4. NMR Analysis
2.5.5. Determination of Amino Acid Sequence
2.5.6. Mass Spectrometry of Synthetic Peptides
2.6. Peptides Synthesis
2.7. Detection of Peptide NDRP in the A < 1 kDa Fraction by Multiple Reaction Monitoring (MRM)
2.8. In Vitro Simulated Digestion of NDRP
2.9. Data Treatment and Statistical Analysis
3. Results and Discussion
3.1. Enzymatic Hydrolysis of Soy Proteins and Degree of Hydrolysis
3.2. SDS-PAGE
3.3. Ultrafiltration and Evaluation of ACE-Inhibitory Activity
3.4. Purification, Characterization and Synthesis of ACE-Inhibitory Peptides
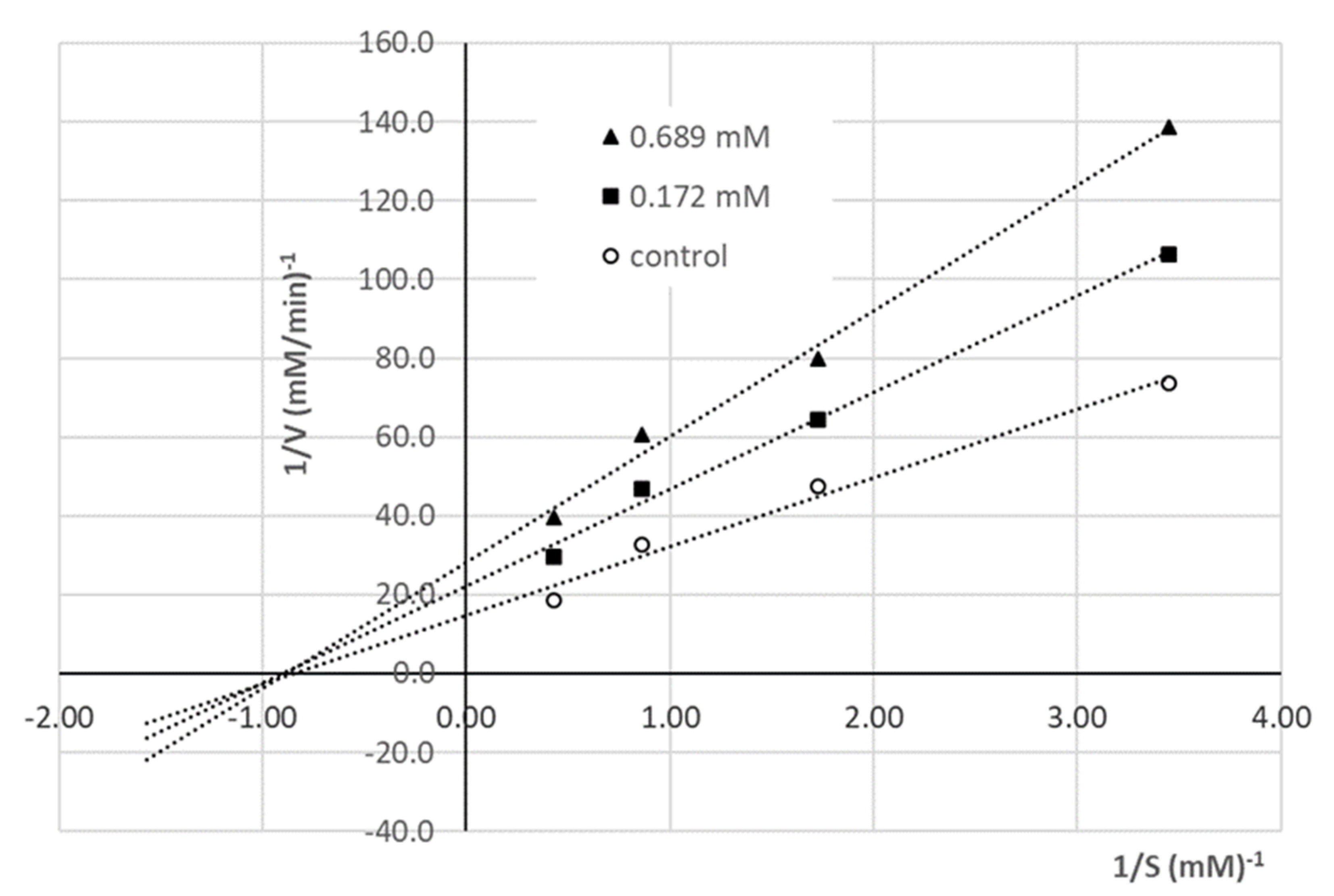
3.5. Determination of the ACE Inhibition Mechanism
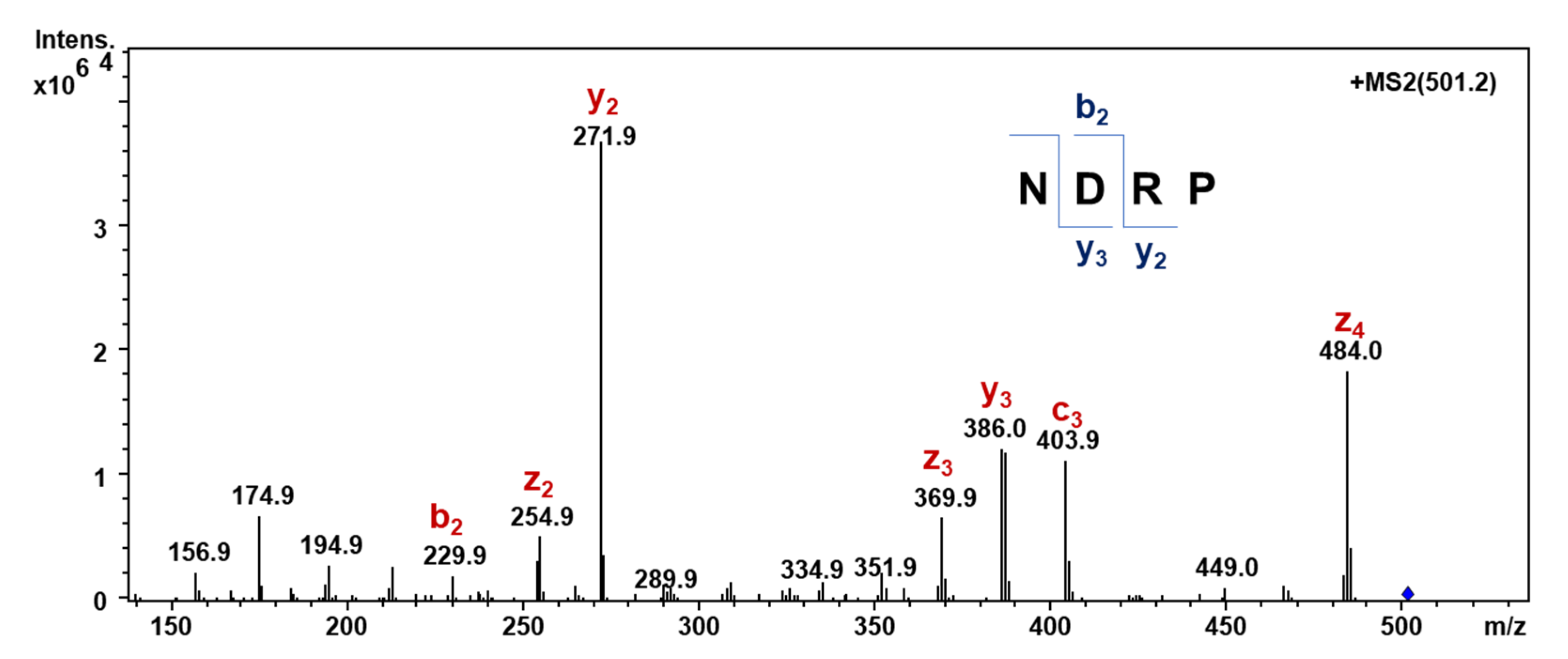
3.6. Detection of Peptide NDRP in the A < 1 kDa Fraction Assessed by MRM Mass Spectrometry Analysis
3.7. Stability of NDRP toward In Vitro Simulated Digestion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iavarone, F.; Desiderio, C.; Vitali, A.; Messana, I.; Martelli, C.; Castagnola, M.; Cabras, T. Cryptides: Latent peptides everywhere. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 246–263. [Google Scholar] [CrossRef] [PubMed]
- Ulug, S.K.; Jahandideh, F.; Wu, J. Novel technologies for the production of bioactive peptides. Trends Food Sci. Technol. 2021, 108, 27–39. [Google Scholar] [CrossRef]
- Sun, X.D. Enzymatic hydrolysis of soy proteins and the hydrolysates utilisation. Int. J. Food Sci. Technol. 2011, 46, 2447–2459. [Google Scholar] [CrossRef]
- Xue, L.; Yin, R.; Howell, K.; Zhang, P. Activity and bioavailability of food protein-derived angiotensin-I-converting enzyme–inhibitory peptides. Compr. Rev. Food. Sci. Food. Saf. 2021, 20, 1150–1187. [Google Scholar] [CrossRef] [PubMed]
- Coscueta, E.R.; Camposa, D.A.; Osório, H.; Nerli, B.B.; Pintado, M. Enzymatic soy protein hydrolysis: A tool for biofunctional food ingredient production. Food Chem. X 2019, 1, 100006. [Google Scholar] [CrossRef]
- WHO (World Health Organization). Available online: https://www.who.int/health-topics/hypertension#tab=tab_1 (accessed on 15 July 2022).
- Lee, S.H.; Qian, Z.J.; Kim, S.K. A novel angiotensin I converting enzyme inhibitory peptide from tuna frame protein hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2010, 118, 96–102. [Google Scholar] [CrossRef]
- Sica, D.A. Angiotensin-converting enzyme inhibitors side effects--physiologic and non-physiologic considerations. J. Clin. Hypertens. 2004, 6, 410–416. [Google Scholar] [CrossRef]
- Conde, J.M.; Patino, J.M.R. The effect of enzymatic treatment of a sunflower protein isolate on the rate of adsorption at the air–water interface. J. Food Eng. 2007, 78, 1001–1009. [Google Scholar] [CrossRef]
- Aluko, R.E. Structure and function of plant protein-derived antihypertensive peptides. Curr. Opin. Food Sci. 2015, 4, 444–450. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant and angiotensin I converting enzyme (ACE) inhibitory activities of date seed protein hydrolysates prepared using Alcalase, Flavourzyme and Thermolysin. J. Funct. Foods 2015, 18, 1125–1137. [Google Scholar] [CrossRef]
- Ashaolu, T.P. Applications of soy protein hydrolysates in the emerging functional foods: A review. Int. J. Food Sci. Technol. 2020, 55, 421–428. [Google Scholar] [CrossRef]
- Shobako, N. Hypotensive peptides derived from plant proteins. Peptides 2021, 142, 170573. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, C.; Sun-Waterhouse, D.; Zhao, T.; Waterhouse, G.I.N.; Zhao, M.; Su, G. Identification of post-digestion angiotensin-I converting enzyme (ACE) inhibitory peptides from soybean protein Isolate: Their production conditions and in silico molecular docking with ACE. Food Chem. 2021, 345, 128855. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.S. The Dilemma with the Soy Protein Health Claim. J. Am. Heart Assoc. 2019, 8, 13. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, L.; Sun, X.; Zhang, J.; Wang, J.; Li, Y. Study on hydrolysis conditions of flavourzyme in soy polypeptide alcalase hydrolysate and soy polypeptide refining process. Adv. J. Food Sci. Technol. 2014, 6, 1027–1032. [Google Scholar] [CrossRef]
- Rostammiry, L.; Reza Saeidi Asl, M.; Safar, R.; Javadian, R. Optimization of the Enzymatic Hydrolysis of Soy Protein Isolate by Alcalase and Trypsin. Biosci. Biotechnol. Res. Asia 2017, 14, 193–200. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the Degree of Hydrolysis of Food Protein Hydrolysates by Trinitrobenzenesulfonic Acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Barbana, C.; Boye, J.I. Angiotensin I-converting enzyme inhibitory properties of lentil protein hydrolysates: Determination of the kinetics of inhibition. Food Chem. 2011, 127, 94–101. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-Inhibitory Peptides from Hemp Seed (Cannabis sativa L.) Proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: https://www.uniprot.org/ (accessed on 15 July 2022).
- Kaur, A.; Kehinde, B.A.; Sharma, P.; Sharma, D.; Kaur, S. Recently isolated food-derived antihypertensive hydrolysates and peptides: A review. Food Chem. 2021, 346, 128719. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.H.; Tavano, O.; Berenguer-Murcia, A.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef] [PubMed]
- Meinlschmidt, P.; Sussmann, D.; Schweiggert-Weisz, U.; Eisner, P. Enzymatic treatment of soy protein isolates: Effects on the potential allergenicity, technofunctionality, and sensory properties. Food Sci. Nutr. 2016, 4, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Clemente, A. Enzymatic protein hydrolysates in human nutrition. Trends Food Sci. Technol. 2000, 11, 254–262. [Google Scholar] [CrossRef]
- Amnuaycheewa, P.; Gonzalez de Mejia, E. Purification, characterisation, and quantification of the soy allergen profilin (Gly m 3) in soy products. Food Chem. 2010, 119, 1671–1680. [Google Scholar] [CrossRef]
- Merz, M.; Eisele, T.; Berends, P.; Appel, D.; Rabe, S.; Blank, I.; Stressler, T.; Fischer, L. Flavourzyme, an Enzyme Preparation with Industrial Relevance: Automated Nine-Step Purification and Partial Characterization of Eight Enzymes. J. Agric. Food Chem. 2015, 63, 5682–5693. [Google Scholar] [CrossRef]
- Zhao, T.; Zheng, L.; Zhang, Q.; Wang, S.; Zhao, Q.; Su, G.; Zhao, M. Stability towards the gastrointestinal simulated digestion and bioactivity of PAYCS and its digestive product PAY with cognitive improving properties. Food Funct. 2019, 10, 2439–2449. [Google Scholar] [CrossRef]
- Bax, A.; Grzesiek, S. ROESY. In Encyclopedia of Nuclear Magnetic Resonance; John Wiley and Sons: Chichester, UK, 2007; Reo-Tim; Volume 7, pp. 4157–4166. [Google Scholar]
- Wang, Y.; Shortle, D. Residual helical and turn structure in the denatured state of staphylococcal nuclease: Analysis of peptide fragments. Fold Des. 1997, 2, 93–100. [Google Scholar] [CrossRef]
- Sornwatana, T.; Bangphoomi, K.; Roytrakul, S.; Wetprasit, N.; Choowongkomon, K.; Ratanapo, S. Chebulin: Terminalia chebula Retz. fruit-derived peptide with angiotensin-I–converting enzyme inhibitory activity. Biotechnol. Appl. Biochem. 2015, 62, 746–753. [Google Scholar] [CrossRef]
- Kuba, M.; Tanaka, K.; Tawata, S.; Takeda, Y.; Yasuda, M. Angiotensin I-converting enzyme inhibitory peptides isolated from tofuyo fermented soybean food. Biosci. Biotechnol. Biochem. 2003, 67, 1278–1283. [Google Scholar] [CrossRef]
- Jiang, Z.; Tian, B.; Brodcrob, A.; Huo, G. Production, analysis and in vivo evaluation of novel angiotensin-I-converting enzyme inhibitory peptides from bovine casein. Food Chem. 2010, 123, 779–786. [Google Scholar] [CrossRef]
- Jang, J.H.; Jeong, S.C.; Kim, J.H.; Lee, Y.H.; Ju, Y.C.; Lee, J.S. Characterisation of a new antihypertensive angiotensin I-converting enzyme inhibitory peptide from Pleurotus cornucopiae. Food Chem. 2011, 127, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-Converting Enzyme (ACE)-Inhibitory Peptides from Plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef]
- Quirós, A.; del Mar Contreras, M.; Ramos, M.; Amigo, L.; Recio, I. Stability to gastrointestinal enzymes and structure–activity relationship of β-casein-peptides with antihypertensive properties. Peptides 2009, 30, 1848–1853. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymatic Protocol | SPH (%) | DH (%) * |
|---|---|---|
| A | 43 | 17 ± 5.3 a |
| F | 64 | 34 ± 5.5 b |
| A + F | 77 | 36 ± 5.6 b |
| Sample | Max ACE Inhibition (%) * | IC50 (μg mL−1) * |
|---|---|---|
| A < 1 kDa | 77.01 ± 0.57 g | 296.57 ± 2.24 c |
| F < 1 kDa | 54.42 ± 3.34 d | 869.87 ± 15.46 f |
| A + F < 1 kDa | 66.54 ± 0.59 e | 558.35 ± 26.88 e |
| A1 | 92.2 ± 0.13 i | 231.75 ± 2.43 b |
| A2 | 70.63 ± 0.32 f | 361.66 ± 2.73 d |
| A3 | 75.72 ± 0.18 g | 304.61 ± 4.81 c |
| ALKPDNR | 25.72 ± 0.84 b | // |
| VVPD | 13.23 ± 1.18 a | // |
| NDRP | 87.76 ± 0.61 h | 148.28 ± 9.83 a |
| NDTP | 46.15 ± 1.72 c | // |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangiorgio, S.; Vidović, N.; Boschin, G.; Aiello, G.; Arcidiaco, P.; Arnoldi, A.; Morelli, C.F.; Rabuffetti, M.; Recca, T.; Scarabattoli, L.; et al. Preparation, Characterization and In Vitro Stability of a Novel ACE-Inhibitory Peptide from Soybean Protein. Foods 2022, 11, 2667. https://doi.org/10.3390/foods11172667
Sangiorgio S, Vidović N, Boschin G, Aiello G, Arcidiaco P, Arnoldi A, Morelli CF, Rabuffetti M, Recca T, Scarabattoli L, et al. Preparation, Characterization and In Vitro Stability of a Novel ACE-Inhibitory Peptide from Soybean Protein. Foods. 2022; 11(17):2667. https://doi.org/10.3390/foods11172667
Chicago/Turabian StyleSangiorgio, Sara, Nikolina Vidović, Giovanna Boschin, Gilda Aiello, Patrizia Arcidiaco, Anna Arnoldi, Carlo F. Morelli, Marco Rabuffetti, Teresa Recca, Letizia Scarabattoli, and et al. 2022. "Preparation, Characterization and In Vitro Stability of a Novel ACE-Inhibitory Peptide from Soybean Protein" Foods 11, no. 17: 2667. https://doi.org/10.3390/foods11172667