
Optimization of Selective Hydrolysis of Cruciferins for Production of Potent Mineral Chelating Peptides and Napins Purification to Valorize Total Rapeseed Meal Proteins
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Enzymatic Proteolysis and Membrane Fractionation
2.2.1. Batch Hydrolysis Experiments
2.2.2. Ultrafiltration of Prolyve® Hydrolysates
2.3. Modelling and Optimizing Methodology
2.3.1. Simulation Methodology
2.3.2. Kinetics Modelling
2.3.3. Multicriteria Optimization
2.4. Analytical Methods
2.4.1. Protein Analysis and Quantification
2.4.2. Peptide Analysis and Quantification
2.5. Iron (II) Chelating Activity
2.6. Statistical Analysis
3. Results
3.1. Study of the Selective Hydrolysis Mechanism of Rapeseed Proteins
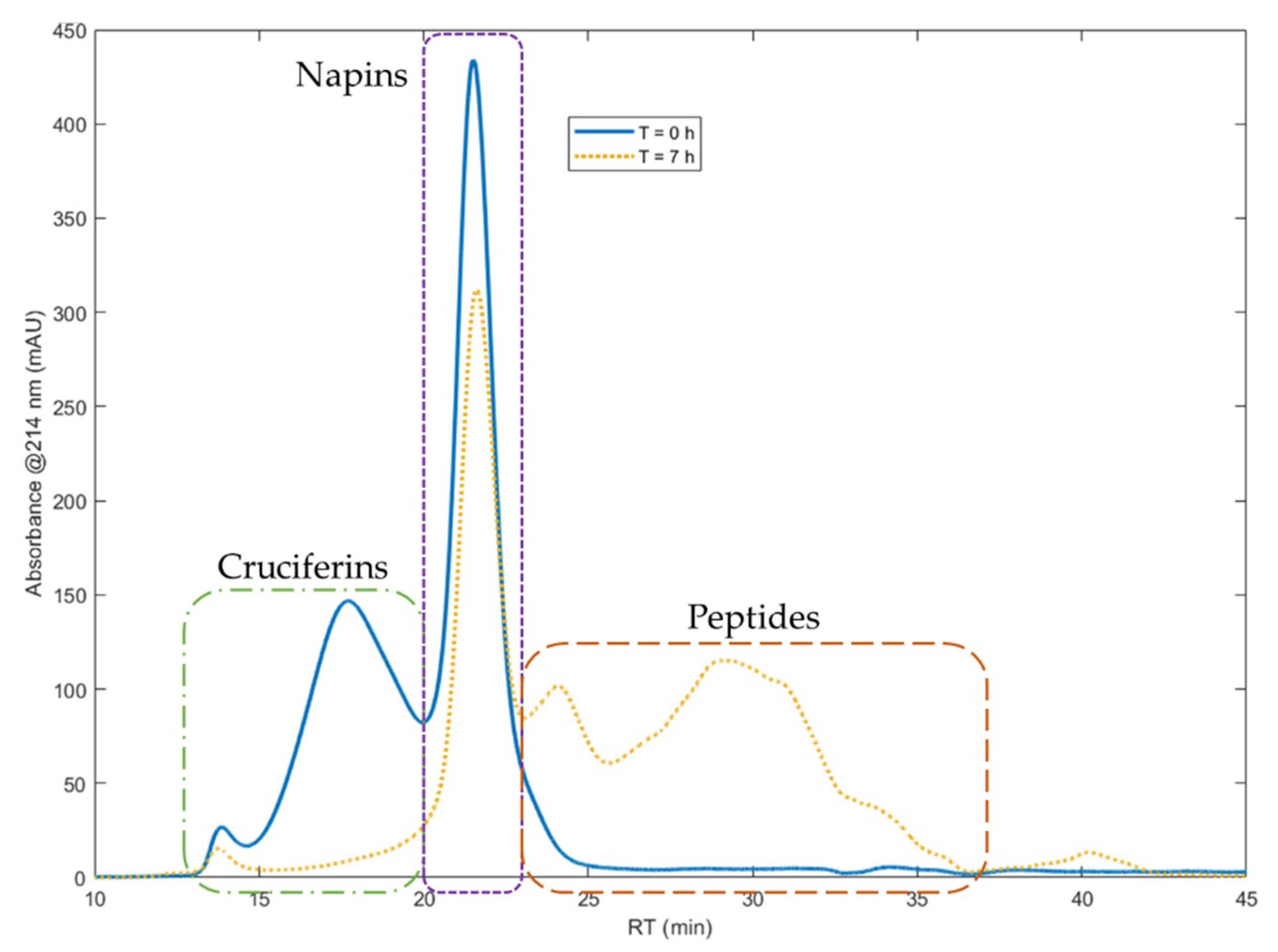
3.1.1. Highlighting of Selective Hydrolysis of Cruciferins
3.1.2. Impact of Reaction Conditions on the Selective Hydrolysis Mechanism
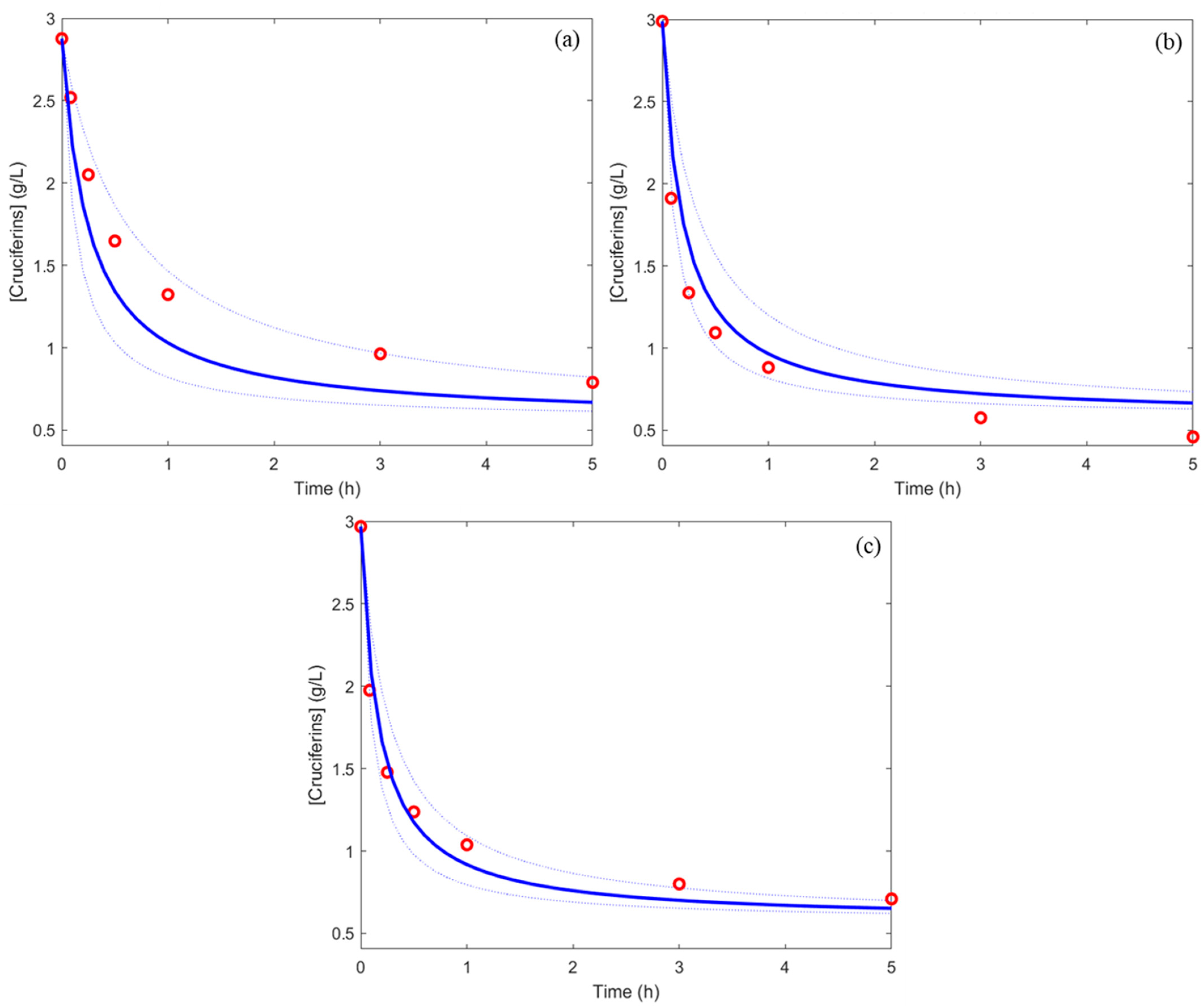
3.2. Modelling of the Kinetic Reactions of Selective Hydrolysis
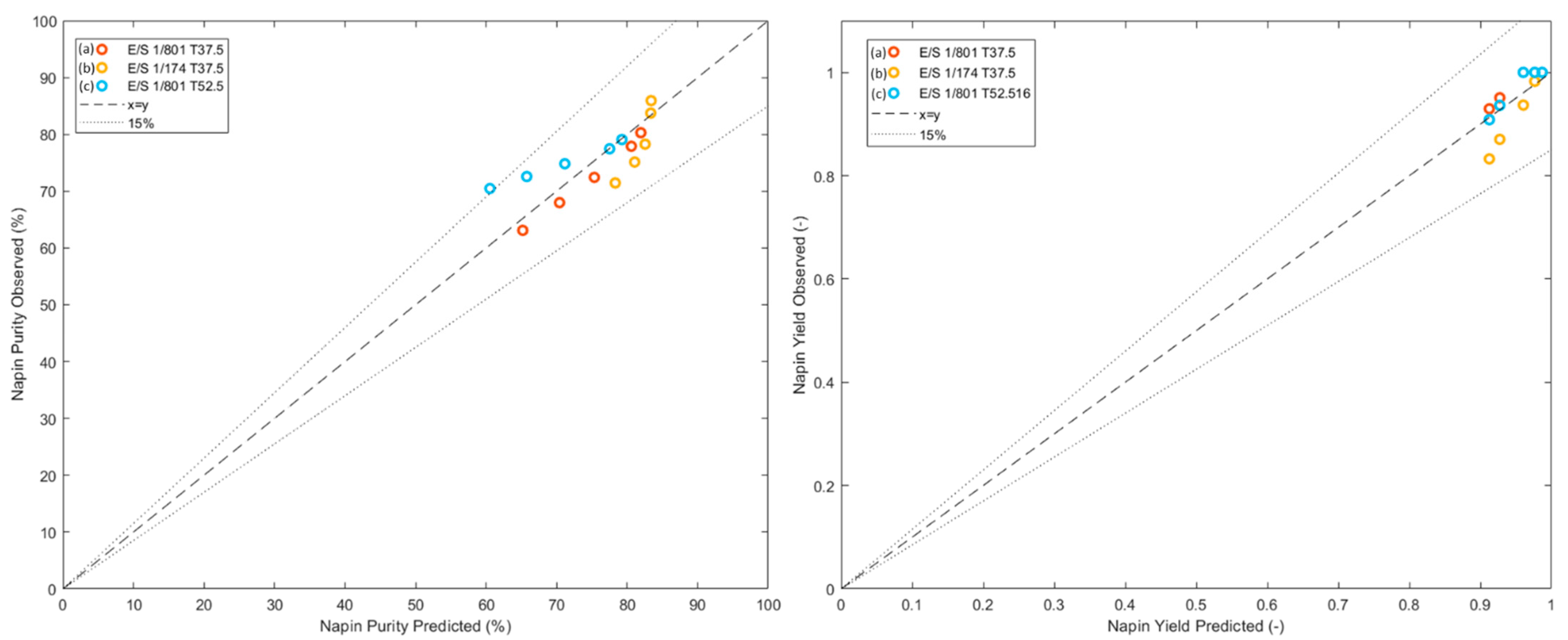
3.3. Multicriteria Optimization of the Selective Hydrolysis
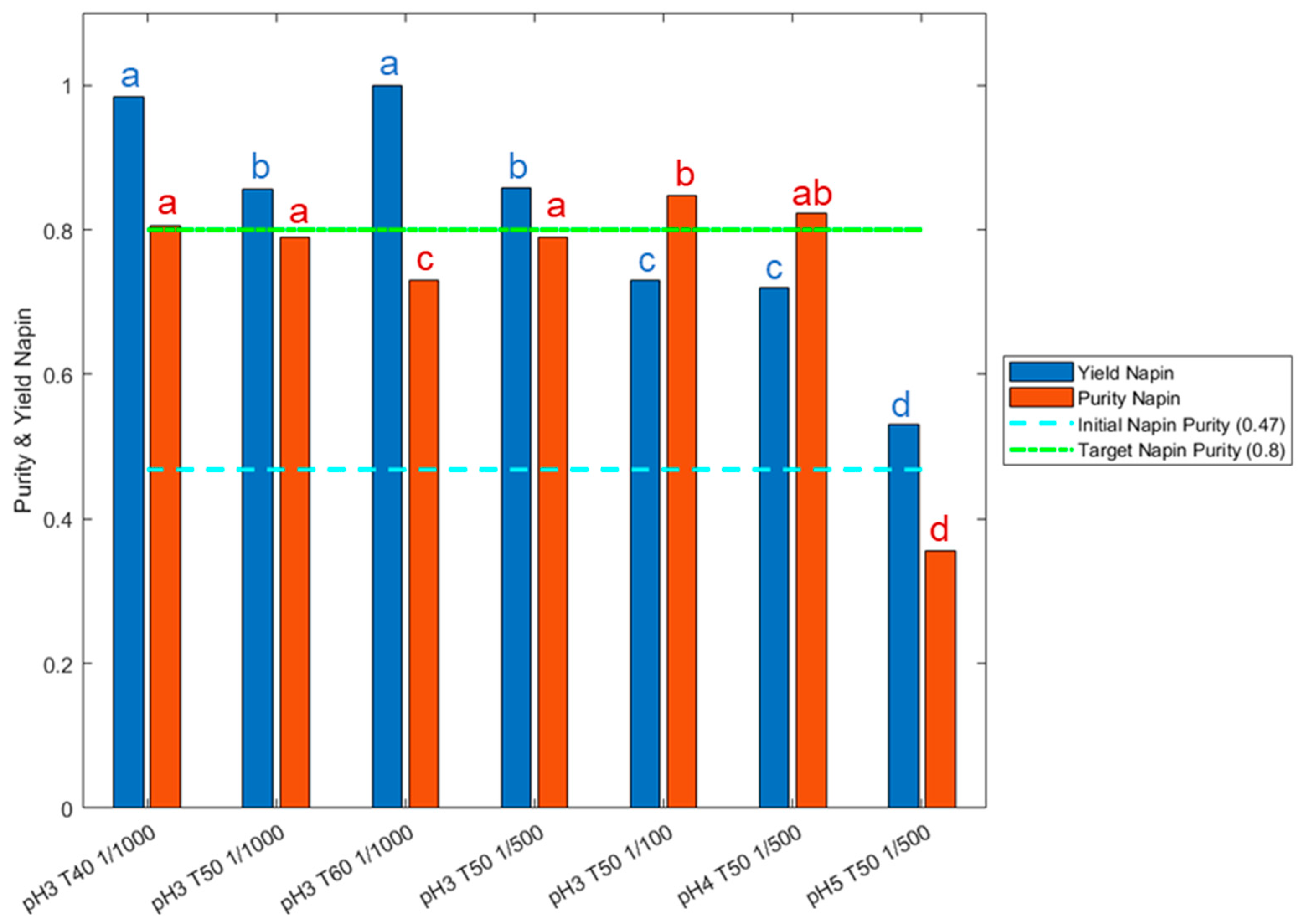
3.3.1. Search for the Best Duration/Enzymatic Cost Trade-Offs
3.3.2. Validation of the Bioactivity Preservation in Optimized Hydrolysis Conditions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization. Food Outlook—Biannual Report on Global Food Markets; FAO: Rome, Italy, 2020; ISBN 978-92-5-132848-4. [Google Scholar]
- Haar, D.V.D.; Müller, K.; Bader-Mittermaier, S.; Eisner, P. Rapeseed Proteins—Production Methods and Possible Application Ranges. OCL 2014, 21, D104. [Google Scholar] [CrossRef]
- Wanasundara, J.P.D. Proteins of Brassicaceae Oilseeds and Their Potential as a Plant Protein Source. Crit. Rev. Food Sci. Nutr. 2011, 51, 635–677. [Google Scholar] [CrossRef] [PubMed]
- Crouch, M.L.; Sussex, I.M. Development and Storage-Protein Synthesis in Brassica napus L. Embryos in Vivo and in Vitro. Planta 1981, 153, 64–74. [Google Scholar] [CrossRef] [PubMed]
- UniProtKB/Swiss-Prot. Available online: www.uniprot.org (accessed on 15 February 2022).
- Josefsson, L.G.; Lenman, M.; Ericson, M.L.; Rask, L. Structure of a Gene Encoding the 1.7 S Storage Protein, Napin, from Brassica Napus. J. Biol. Chem. 1987, 262, 12196–12201. [Google Scholar] [CrossRef]
- Ericson, M.L.; Rödin, J.; Lenman, M.; Glimelius, K.; Josefsson, L.G.; Rask, L. Structure of the Rapeseed 1.7 S Storage Protein, Napin, and Its Precursor. J. Biol. Chem. 1986, 261, 14576–14581. [Google Scholar] [CrossRef]
- Raab, B.; Leman, H.; Schwenke, K.D.; Kozlowska, H. Comparative Study of the Protein Patterns of Some Rapeseed (Brassica napus L.) Varieties by Means of Polyacrylamide Gel Electrophoresis and High-Performance Liquid Chromatography. Food/Nahrung 1992, 36, 239–247. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed Storage Proteins: Structures ’and Biosynthesis. The Plant Cell. 1995, 7, 945–956. [Google Scholar] [CrossRef]
- Dalgalarrondo, M.; Robin, J.-M.; Azanza, J.-L. Subunit Composition of the Globulin Fraction of Rapeseed (Brassica napus L.). Plant Sci. 1986, 43, 115–124. [Google Scholar] [CrossRef]
- Aider, M.; Barbana, C. Canola Proteins: Composition, Extraction, Functional Properties, Bioactivity, Applications as a Food Ingredient and Allergenicity—A Practical and Critical Review. Trends Food Sci. Technol. 2011, 22, 21–39. [Google Scholar] [CrossRef]
- Nioi, C.; Kapel, R.; Rondags, E.; Marc, I. Selective Extraction, Structural Characterisation and Antifungal Activity Assessment of Napins from an Industrial Rapeseed Meal. Food Chem. 2012, 134, 2149–2155. [Google Scholar] [CrossRef]
- Rahman, M.; Browne, J.J.; Van Crugten, J.; Hasan, M.F.; Liu, L.; Barkla, B.J. In Silico, Molecular Docking and In Vitro Antimicrobial Activity of the Major Rapeseed Seed Storage Proteins. Front. Pharmacol. 2020, 11, 1340. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Protein and Amino Acid Requirements in Human Nutrition; FAO, Vereinte Nationen, Eds.; WHO Technical Report Series; WHO: Geneva, Switzerland, 2007; ISBN 978-92-4-120935-9.
- Tan, S.H.; Mailer, R.J.; Blanchard, C.L.; Agboola, S.O. Canola Proteins for Human Consumption: Extraction, Profile, and Functional Properties. J. Food Sci. 2011, 76, R16–R28. [Google Scholar] [CrossRef] [PubMed]
- Albe-Slabi, S.; Defaix, C.; Beaubier, S.; Galet, O.; Kapel, R. Selective Extraction of Napins: Process Optimization and Impact on Structural and Functional Properties. Food Hydrocoll. 2022, 122, 107105. [Google Scholar] [CrossRef]
- Beaubier, S.; Defaix, C.; Albe-Slabi, S.; Aymes, A.; Galet, O.; Fournier, F.; Kapel, R. Multiobjective Decision Making Strategy for Selective Albumin Extraction from a Rapeseed Cold-Pressed Meal Based on Rough Set Approach. Food Bioprod. Processing 2022, 133, 34–44. [Google Scholar] [CrossRef]
- Tranter, H.S.; Board, R.G. The Antimicrobial Defense of Avian Eggs: Biological Perspective and Chemical Basis. J. Appl. Biochem. 1982, 4, 295–338. [Google Scholar]
- Xue, Z.; Yu, W.; Wu, M.; Wang, J. In Vivo Antitumor and Antioxidative Effects of a Rapeseed Meal Protein Hydrolysate on an S180 Tumor-Bearing Murine Model. Biosci. Biotechnol. Biochem. 2009, 73, 2412–2415. [Google Scholar] [CrossRef]
- Karami, Z.; Peighambardoust, S.H.; Hesari, J.; Akbari-Adergani, B.; Andreu, D. Antioxidant, Anticancer and ACE-Inhibitory Activities of Bioactive Peptides from Wheat Germ Protein Hydrolysates. Food Biosci. 2019, 32, 100450. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Fujita, H.; Matoba, N.; Takenaka, Y.; Yamamoto, T.; Yamauchi, R.; Tsuruki, H.; Takahata, K. Bioactive Peptides Derived from Food Proteins Preventing Lifestyle-Related Diseases. BioFactors 2000, 12, 143–146. [Google Scholar] [CrossRef]
- Durand, E.; Beaubier, S.; Ilic, I.; Fine, F.; Kapel, R.; Villeneuve, P. Production and Antioxidant Capacity of Bioactive Peptides from Plant Biomass to Counteract Lipid Oxidation. Curr. Res. Food Sci. 2021, 4, 365–397. [Google Scholar] [CrossRef]
- Durand, E.; Beaubier, S.; Fine, F.; Villeneuve, P.; Kapel, R. High Metal Chelating Properties from Rapeseed Meal Proteins to Counteract Lipid Oxidation in Foods: Controlled Proteolysis and Characterization. Eur. J. Lipid Sci. Technol. 2021, 123, 2000380. [Google Scholar] [CrossRef]
- Beaubier, S.; Framboisier, X.; Fournier, F.; Galet, O.; Kapel, R. A New Approach for Modelling and Optimizing Batch Enzymatic Proteolysis. Chem. Eng. J. 2021, 405, 126871. [Google Scholar] [CrossRef]
- Deng, Y.; van der Veer, F.; Sforza, S.; Gruppen, H.; Wierenga, P.A. Towards Predicting Protein Hydrolysis by Bovine Trypsin. Process Biochem. 2018, 65, 81–92. [Google Scholar] [CrossRef]
- Muniglia, L.; Nandor Kiss, L.; Fonteix, C.; Marc, I. Multicriteria Optimization of a Single-Cell Oil Production. Eur. J. Oper. Res. 2004, 153, 360–369. [Google Scholar] [CrossRef]
- Zio, E.; Baraldi, P.; Pedroni, N. Optimal Power System Generation Scheduling by Multi-Objective Genetic Algorithms with Preferences. Reliab. Eng. Syst. Saf. 2009, 24, 432–444. [Google Scholar] [CrossRef]
- Defaix, C.; Aymes, A.; Albe Slabi, S.; Basselin, M.; Mathé, C.; Galet, O.; Kapel, R. A New Size-Exclusion Chromatography Method for Fast Rapeseed Albumin and Globulin Quantification. Food Chem. 2019, 287, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, B.J.H.; Gruppen, H. Prediction of Molar Extinction Coefficients of Proteins and Peptides Using UV Absorption of the Constituent Amino Acids at 214 Nm To Enable Quantitative Reverse Phase High-Performance Liquid Chromatography−Mass Spectrometry Analysis. J. Agric. Food Chem. 2007, 55, 5445–5451. [Google Scholar] [CrossRef] [PubMed]
- Beaubier, S.; Framboisier, X.; Ioannou, I.; Galet, O.; Kapel, R. Simultaneous Quantification of the Degree of Hydrolysis, Protein Conversion Rate and Mean Molar Weight of Peptides Released in the Course of Enzymatic Proteolysis. J. Chromatogr. B 2019, 1105, 1–9. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymic Hydrolysis of Proteins for Increased Solubility. J. Agric. Food Chem. 1976, 24, 1090–1093. [Google Scholar] [CrossRef]
- Das, K.P.; Kinsella, J.E. PH DEPENDENT EMULSIFYING PROPERTIES OF B-LACTOGLOBULIN. J. Dispers. Sci. Technol. 1989, 10, 77–102. [Google Scholar] [CrossRef]
- Kella, N.K.D.; Kinsella, J.E. Enhanced Thermodynamic Stability of β-Lactoglobulin at Low pH. Biochem. J. 1988, 255, 6. [Google Scholar] [CrossRef]
- Laskowski, M.; Scheraga, H.A. Thermodynamic Considerations of Protein Reactions. 1,2 I. Modified Reactivity of Polar Groups. J. Am. Chem. Soc. 1954, 76, 6305–6319. [Google Scholar] [CrossRef]
- Joehnke, M.S.; Lametsch, R.; Sørensen, J.C. Improved in Vitro Digestibility of Rapeseed Napin Proteins in Mixtures with Bovine Beta-Lactoglobulin. Food Res. Int. 2019, 123, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Jyothi, T.C.; Sinha, S.; Singh, S.A.; Surolia, A.; Appu Rao, A.G. Napin from Brassica Juncea: Thermodynamic and Structural Analysis of Stability. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2007, 1774, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Perera, S.P.; McIntosh, T.C.; Wanasundara, J.P.D. Structural Properties of Cruciferin and Napin of Brassica Napus (Canola) Show Distinct Responses to Changes in PH and Temperature. Plants 2016, 5, 36. [Google Scholar] [CrossRef]
- Schmidt, I.; Renard, D.; Rondeau, D.; Richomme, P.; Popineau, Y.; Axelos, M.A.-V. Detailed Physicochemical Characterization of the 2S Storage Protein from Rape (Brassica napus L.). J. Agric. Food Chem. 2004, 52, 5995–6001. [Google Scholar] [CrossRef]
- Folawiyo, Y.L.; Apenten, R.K.O. The Effect of Heat- and Acid-Treatment on the Structure of Rapeseed Albumin (Napin). Food Chem. 1997, 58, 237–243. [Google Scholar] [CrossRef]
- Islam, M.M.; Kobayashi, K.; Kidokoro, S.-I.; Kuroda, Y. Hydrophobic Surface Residues Can Stabilize a Protein through Improved Water–Protein Interactions. FEBS J. 2019, 286, 4122–4134. [Google Scholar] [CrossRef]
- Malabat, C.; Nchez-Vioque, R.I.S.; Rabiller, C.; Guen, J.G. Emulsifying and Foaming Properties of Native and Chemically Modified Peptides from the 2S and 12S Proteins of Rapeseed (Brassica napus L.). J. Am. Oil Chem. Soc. 2001, 78, 235–242. [Google Scholar] [CrossRef]
- Wanasundara, J.P.D.; McIntosh, T.C.; Perera, S.P.; Withana-Gamage, T.S.; Mitra, P. Canola/Rapeseed Protein-Functionality and Nutrition. OCL 2016, 23, D407. [Google Scholar] [CrossRef]
- Linderstrom-Lang, K. The initial phases of the enzymatic degradation of proteins. Bull. Soc. Chim. Biol. 1953, 35, 100–116. [Google Scholar]
- Lisak Jakopović, K.; Cheison, S.C.; Kulozik, U.; Božanić, R. Comparison of Selective Hydrolysis of α-Lactalbumin by Acid Protease A and Protease M as Alternative to Pepsin: Potential for β-Lactoglobulin Purification in Whey Proteins. J. Dairy Res. 2019, 86, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.; Amoroso, L. Combating Micronutrient Deficiencies: Food-Based Approaches; CABI: Wallingford, UK, 2011; ISBN 978-1-84593-714-0. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | T (°C) | E/S (g Enzyme/g Substrate) |
|---|---|---|
| −1 | 35 | 1/2000 |
| 0 | 45 | 1/285 |
| 1 | 55 | 1/154 |
| Experience Name | T (°C) | E/S (g Enzyme/g Substrate) | ||
|---|---|---|---|---|
| a | −0.75 | 37.5 | −0.75 | 1/801 |
| b | −0.75 | 37.5 | 0.75 | 1/174 |
| c | 0.75 | 52.5 | −0.75 | 1/801 |
| Variables | ||
|---|---|---|
| Significant Coefficients | Constant | 10.16 |
| 8.49 | ||
| T | 9.11 | |
| 9.76 | ||
| ES2 | - | |
| T2 | 4.078 | |
| Model analysis | RSD | 1.306 |
| R² | 0.99 | |
| Q² | 0.96 | |
| Model (p-value) | 2.58 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaugarenia, N.; Beaubier, S.; Durand, E.; Aymes, A.; Villeneuve, P.; Lesage, F.; Kapel, R. Optimization of Selective Hydrolysis of Cruciferins for Production of Potent Mineral Chelating Peptides and Napins Purification to Valorize Total Rapeseed Meal Proteins. Foods 2022, 11, 2618. https://doi.org/10.3390/foods11172618
Kaugarenia N, Beaubier S, Durand E, Aymes A, Villeneuve P, Lesage F, Kapel R. Optimization of Selective Hydrolysis of Cruciferins for Production of Potent Mineral Chelating Peptides and Napins Purification to Valorize Total Rapeseed Meal Proteins. Foods. 2022; 11(17):2618. https://doi.org/10.3390/foods11172618
Chicago/Turabian StyleKaugarenia, Nastassia, Sophie Beaubier, Erwann Durand, Arnaud Aymes, Pierre Villeneuve, François Lesage, and Romain Kapel. 2022. "Optimization of Selective Hydrolysis of Cruciferins for Production of Potent Mineral Chelating Peptides and Napins Purification to Valorize Total Rapeseed Meal Proteins" Foods 11, no. 17: 2618. https://doi.org/10.3390/foods11172618