
Optimisation and Characterisation of Novel Angiotensin-Converting Enzyme Inhibitory Peptides Prepared by Double Enzymatic Hydrolysis from Agaricus bisporus Scraps

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Extraction of Protein from A. bisporus Scraps
2.3. Determination of ACE Inhibition Rate
2.4. Determination of the Degree of Protein Hydrolysis
2.5. Protease Screening
2.6. Single Factor Experimental Design
2.6.1. Addition Ratio of Double-Enzyme Hydrolysis
2.6.2. Determination of the Optimal Temperature for Enzymatic Digestion
2.6.3. Determination of the Optimal pH for Enzymatic Hydrolysis
2.6.4. Determination of the Optimal Enzymatic Digestion Duration
2.7. Response Surface Experimental Design
2.8. Separation and Purification
2.8.1. Purification of Mushroom ACE Inhibitor Peptides Using Macroporous Resin (Purchased from Shanghai Yuanye Bio-Technology Co., Ltd., Shanghai, China)
Screening of the Macroporous Resin
Dynamic Adsorption and Desorption Experiments
2.8.2. Gel Chromatography Separation
2.9. Determination of Molecular Mass by HPLC
2.10. Sequence Determination
2.11. Molecular Docking
2.12. Stability Verification
2.12.1. pH Stability of ACE Inhibitory Peptides
2.12.2. Temperature Stability of ACE Inhibitory Peptides
2.12.3. In Vitro Digestion of ACE Inhibitory Peptides
3. Results
3.1. Screening of Proteases
3.2. Single Factor Testing
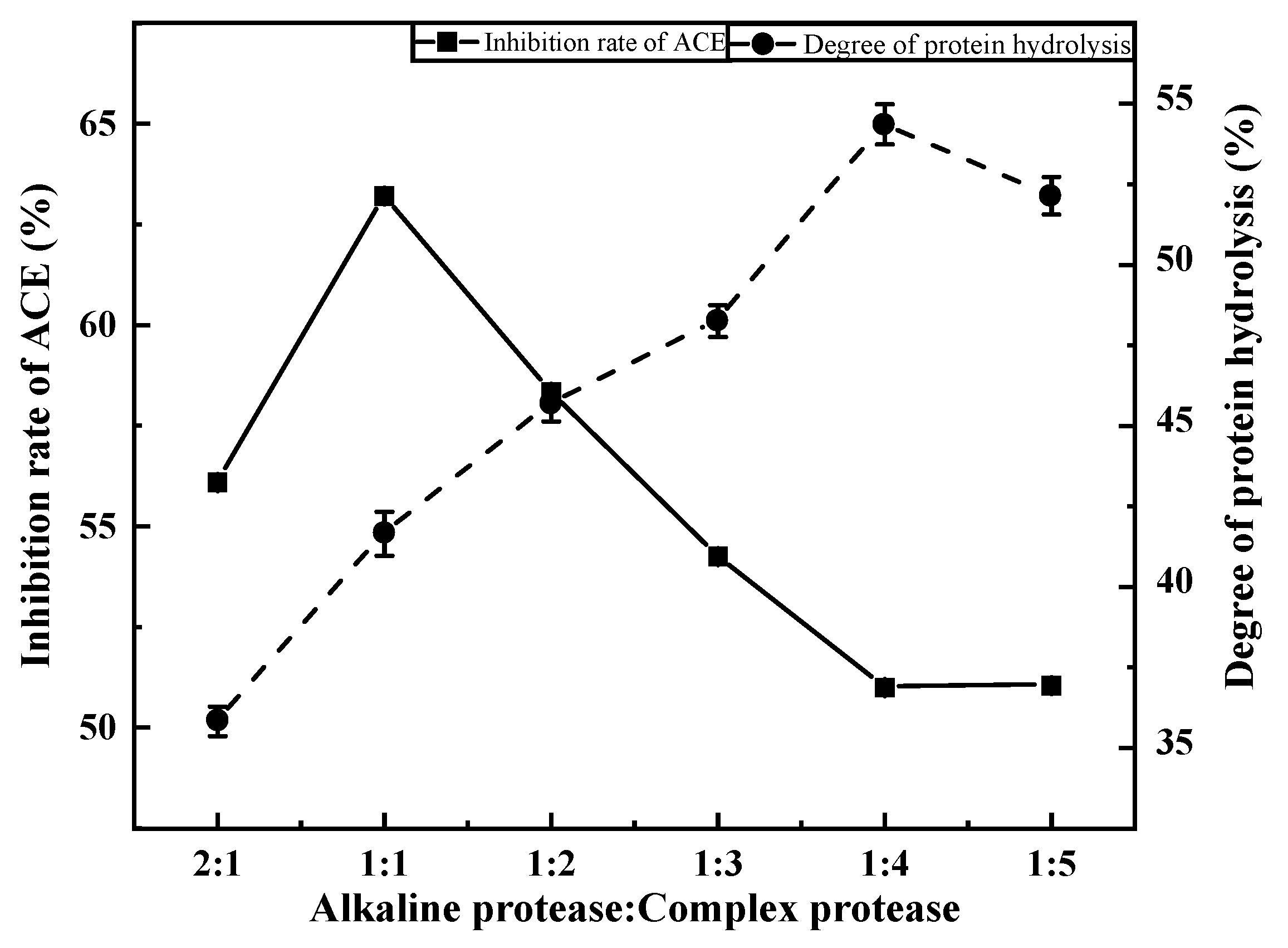
3.2.1. Effects of Compound Ratio on ACE Inhibition Rate
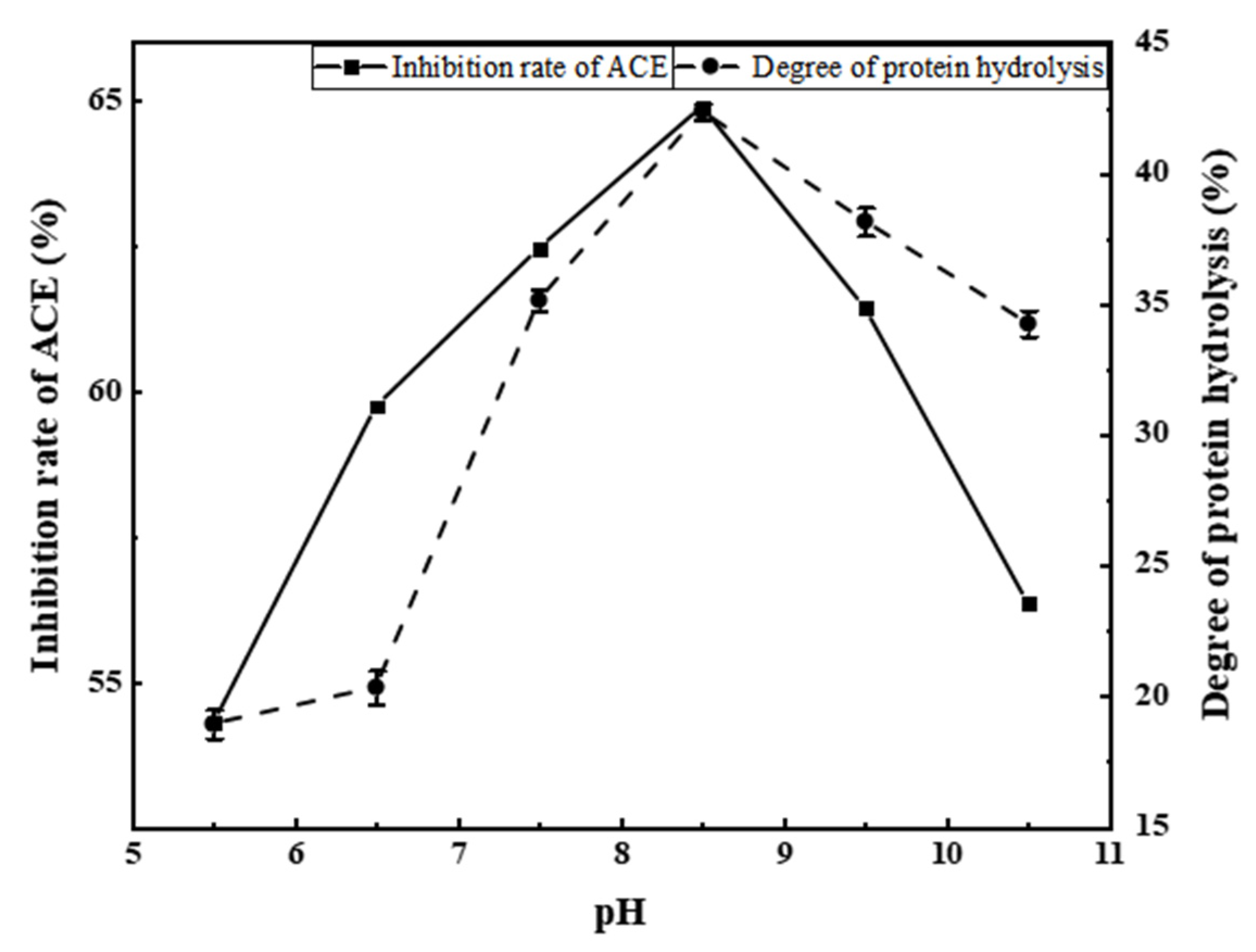
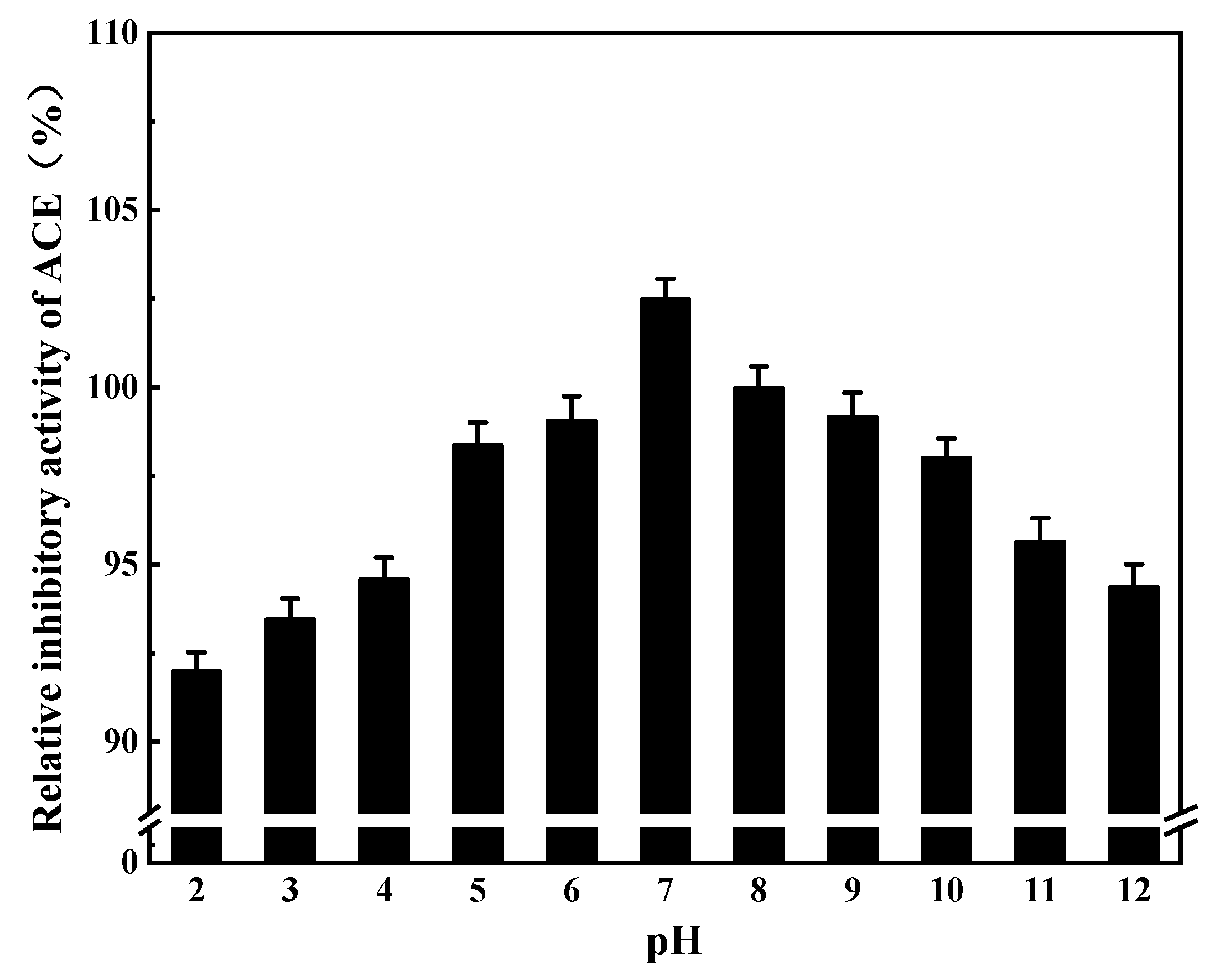
3.2.2. Effects of pH on ACE Inhibition Rate
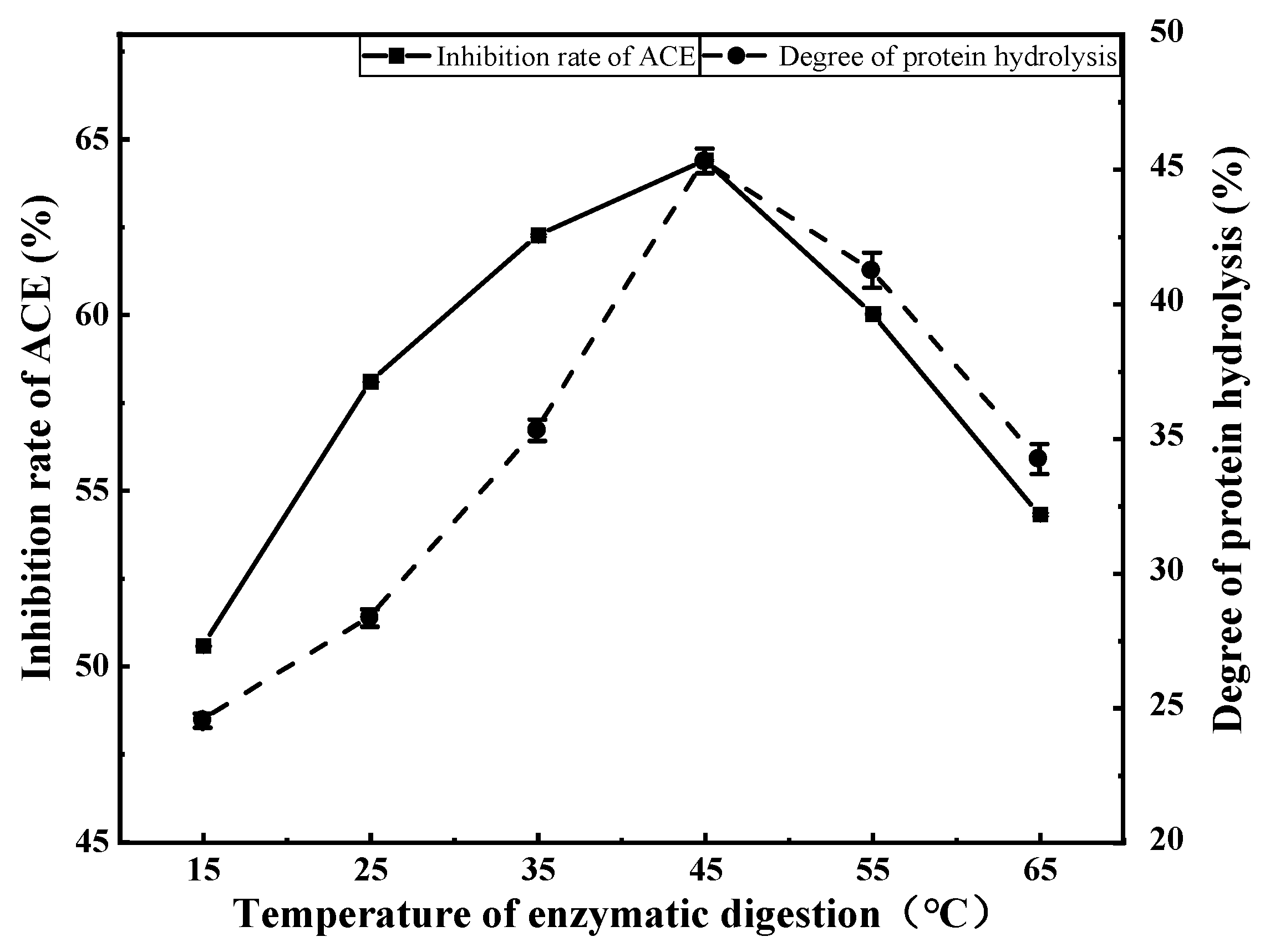
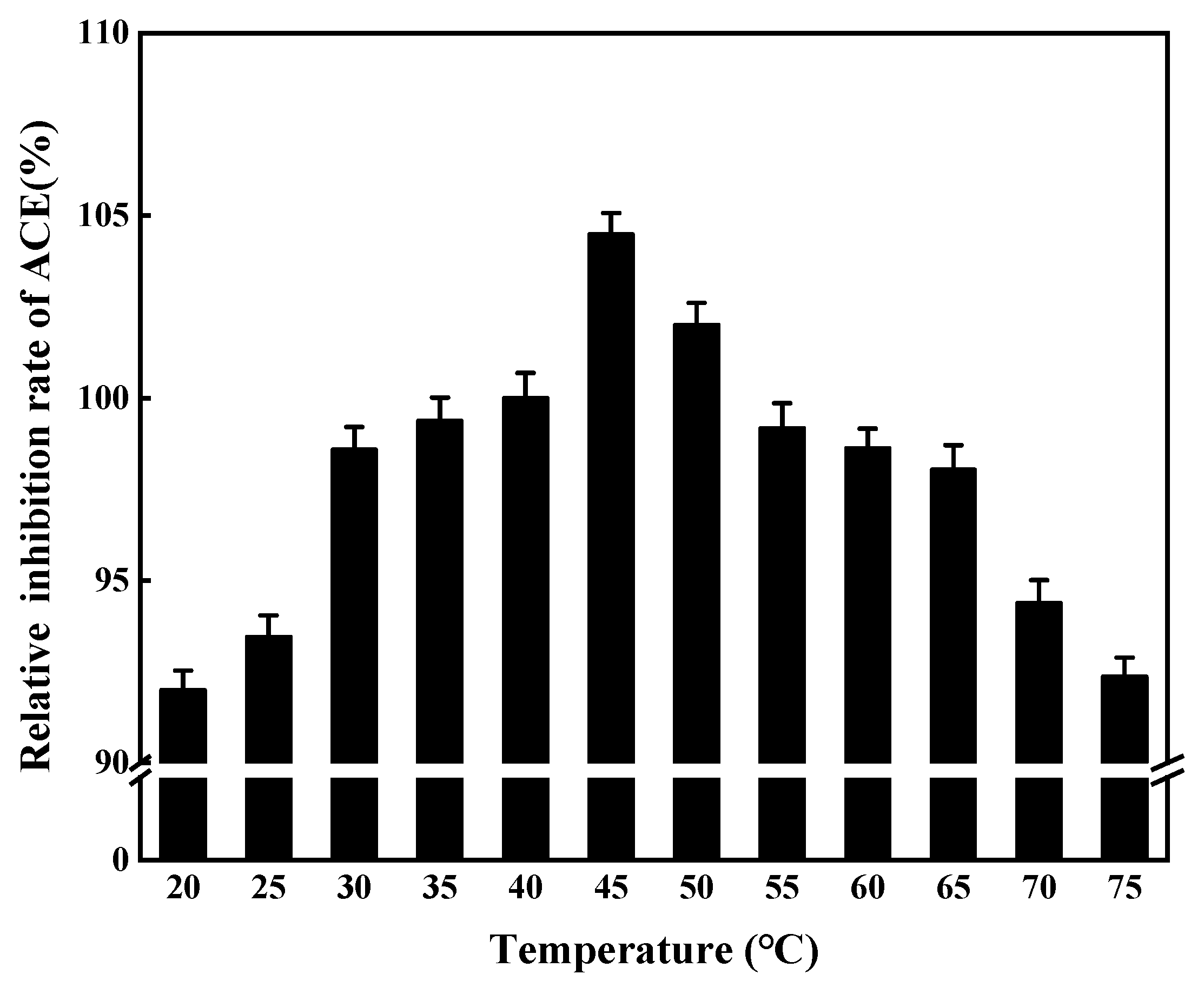
3.2.3. Effects of Enzymatic Digestion Temperature on ACE Inhibition Rate
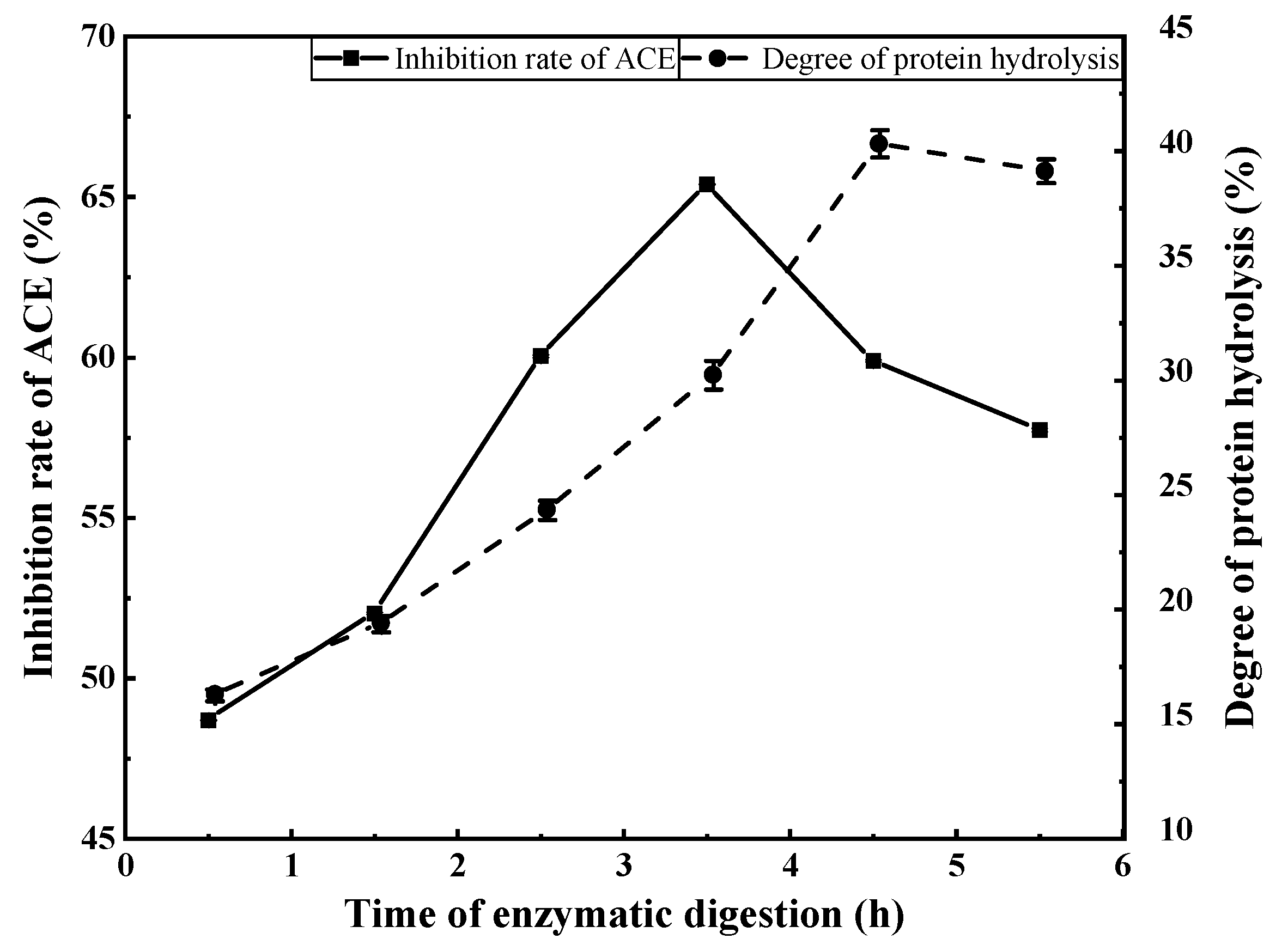
3.2.4. Effects of Enzymatic Digestion Duration on ACE Inhibition Rate
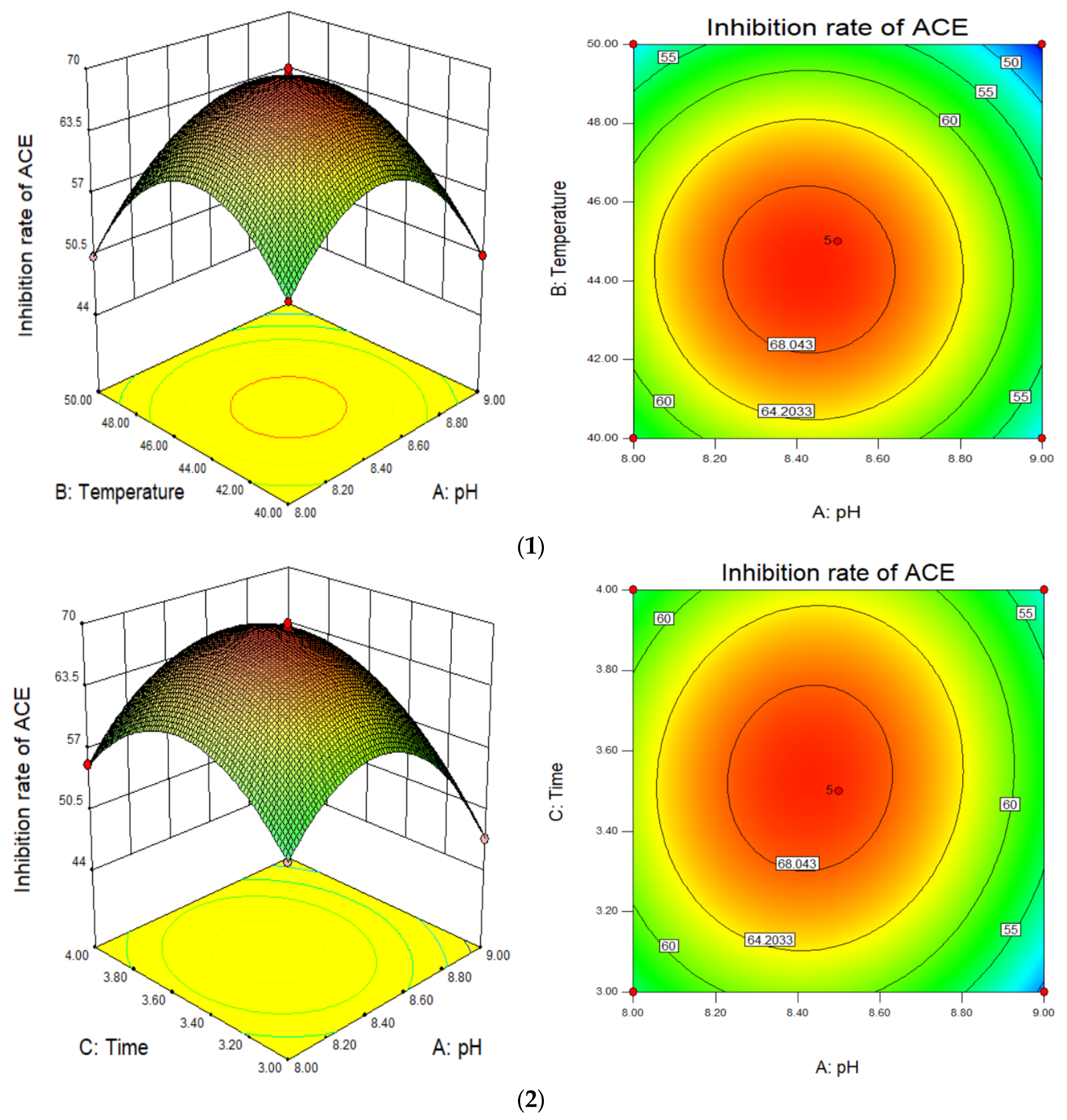
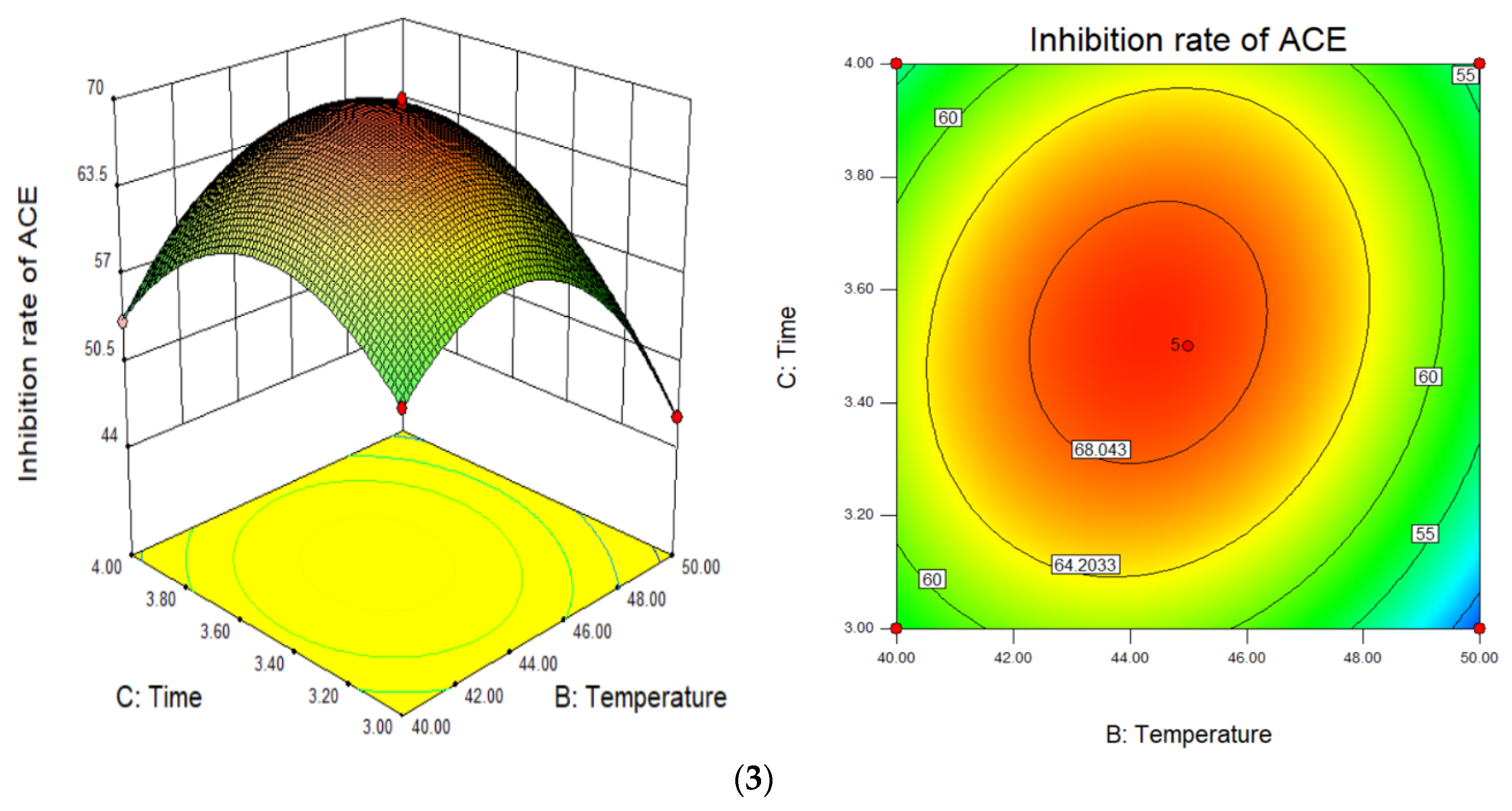
3.3. Response Surface Optimisation
3.4. Separation and Purification
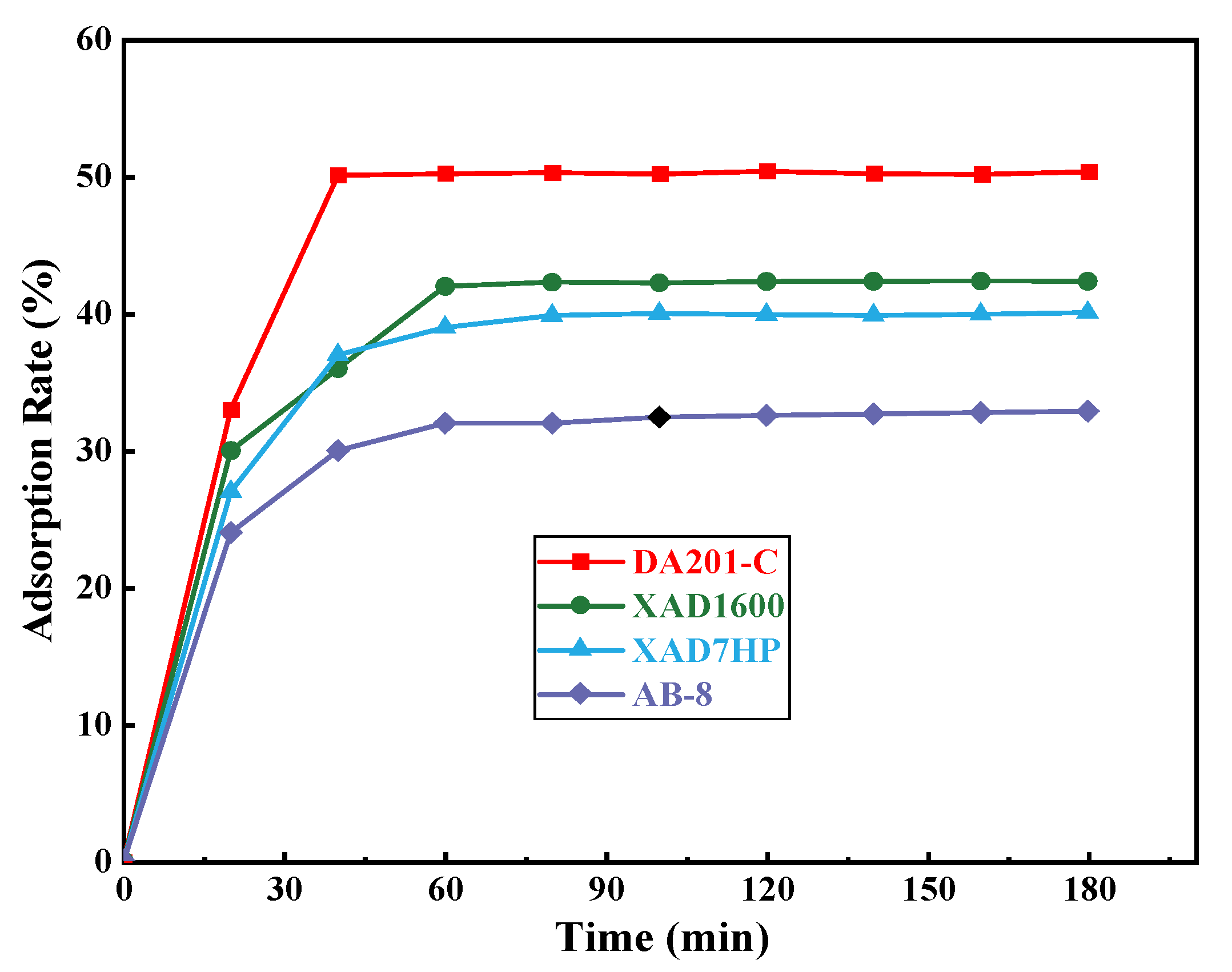
3.4.1. Purification of ACE Inhibitory Peptides Using Macroporous Resin
Screening of Macroporous Resin
Dynamic Adsorption and Washing of Enzymatic Hydrolysates
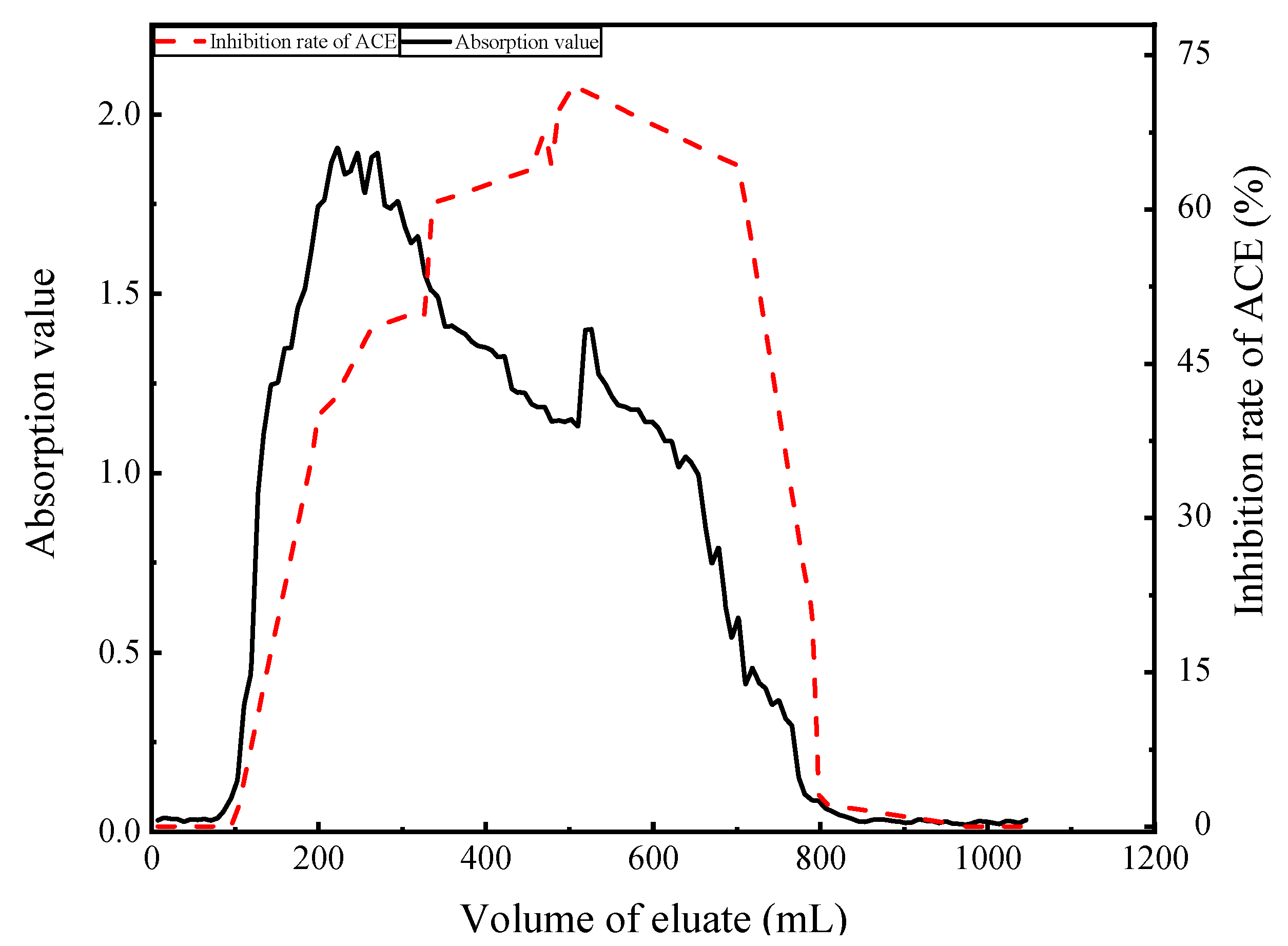
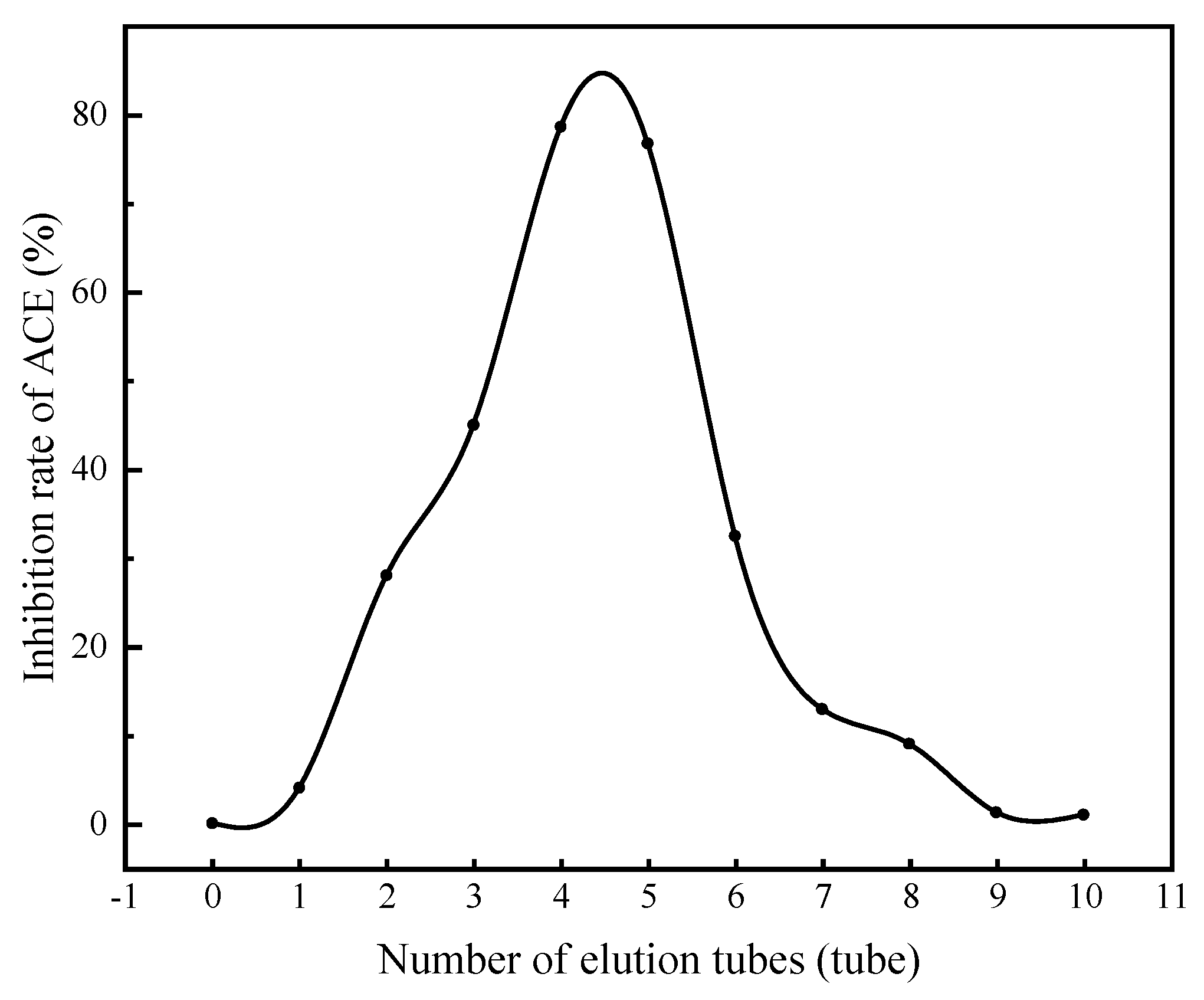
3.4.2. Separation of ACE Inhibitory Peptides by Gel Chromatography
3.5. Comparison of the Activity of ACE Inhibitory Peptides before and after Purification and Separation
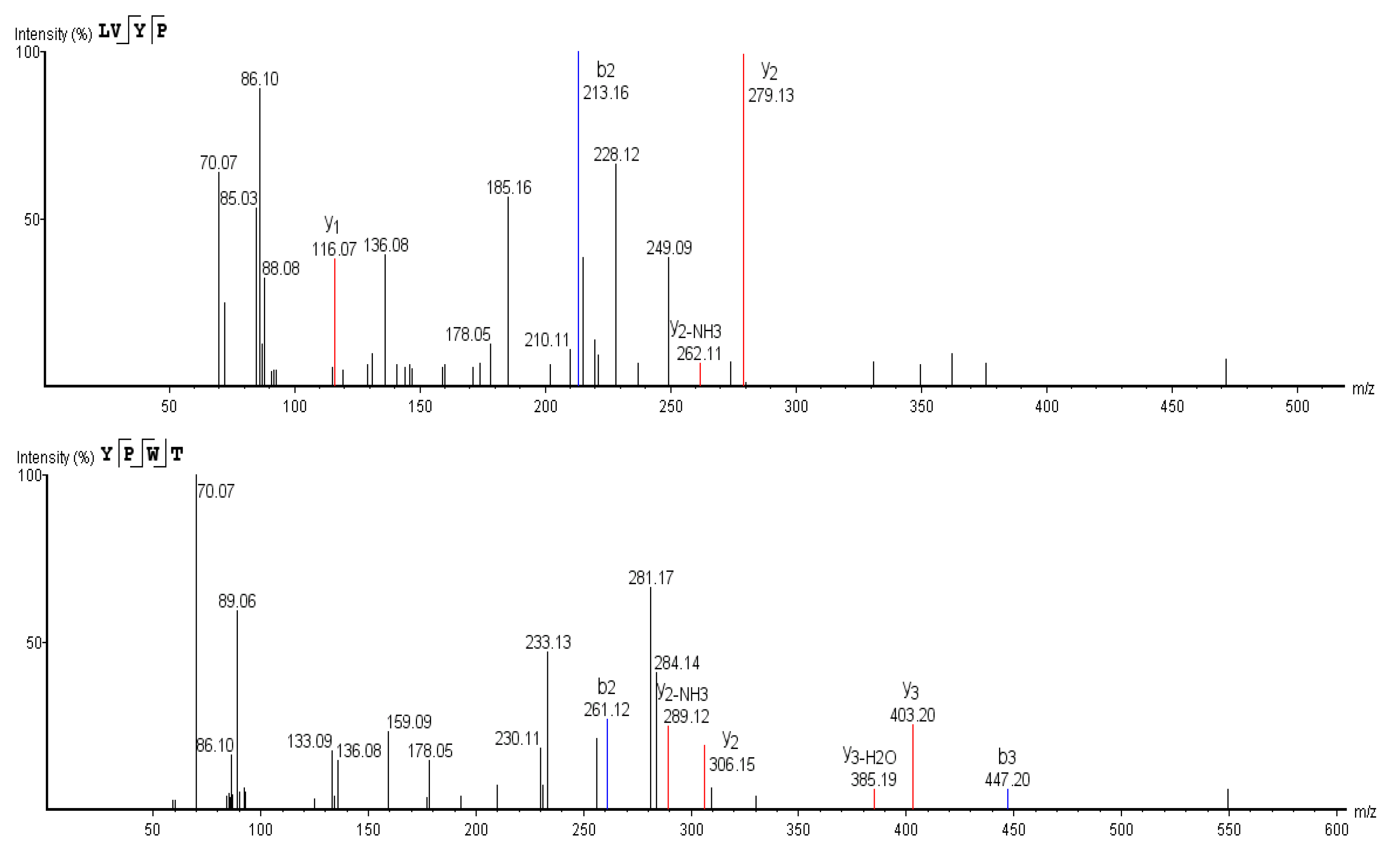
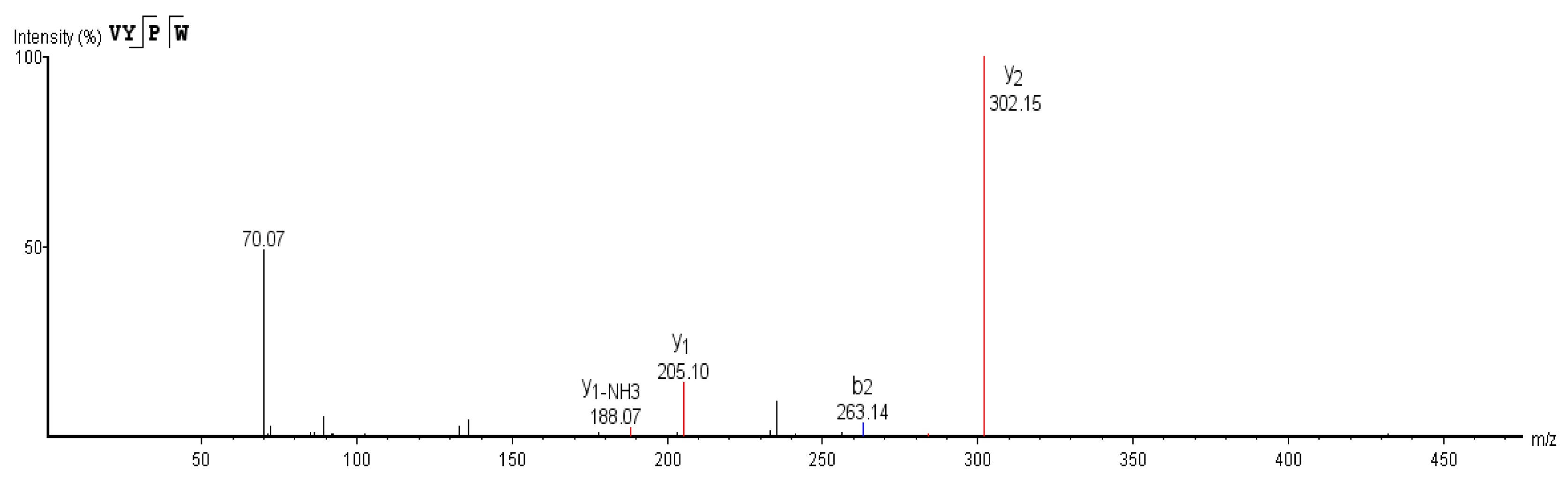
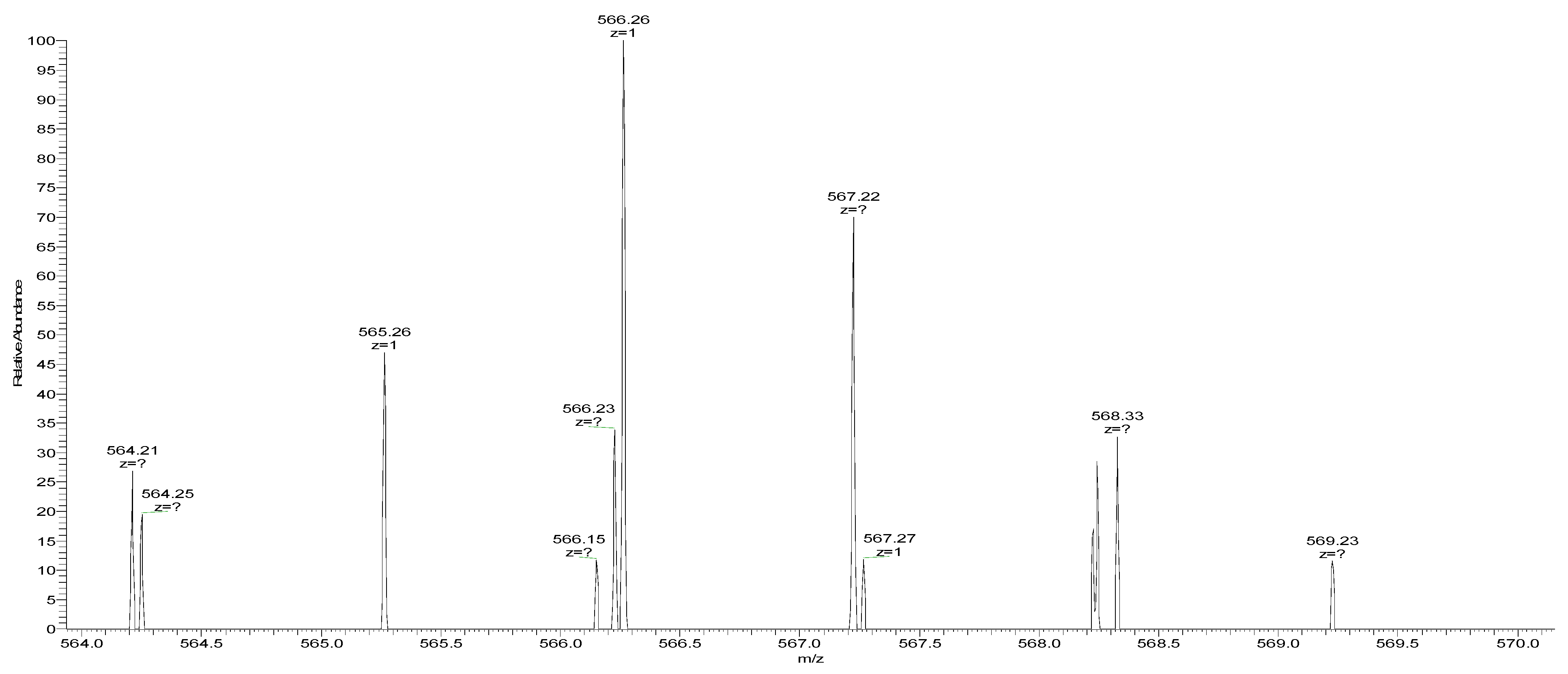
3.6. Sequence Determination of ACE Inhibitory Peptides
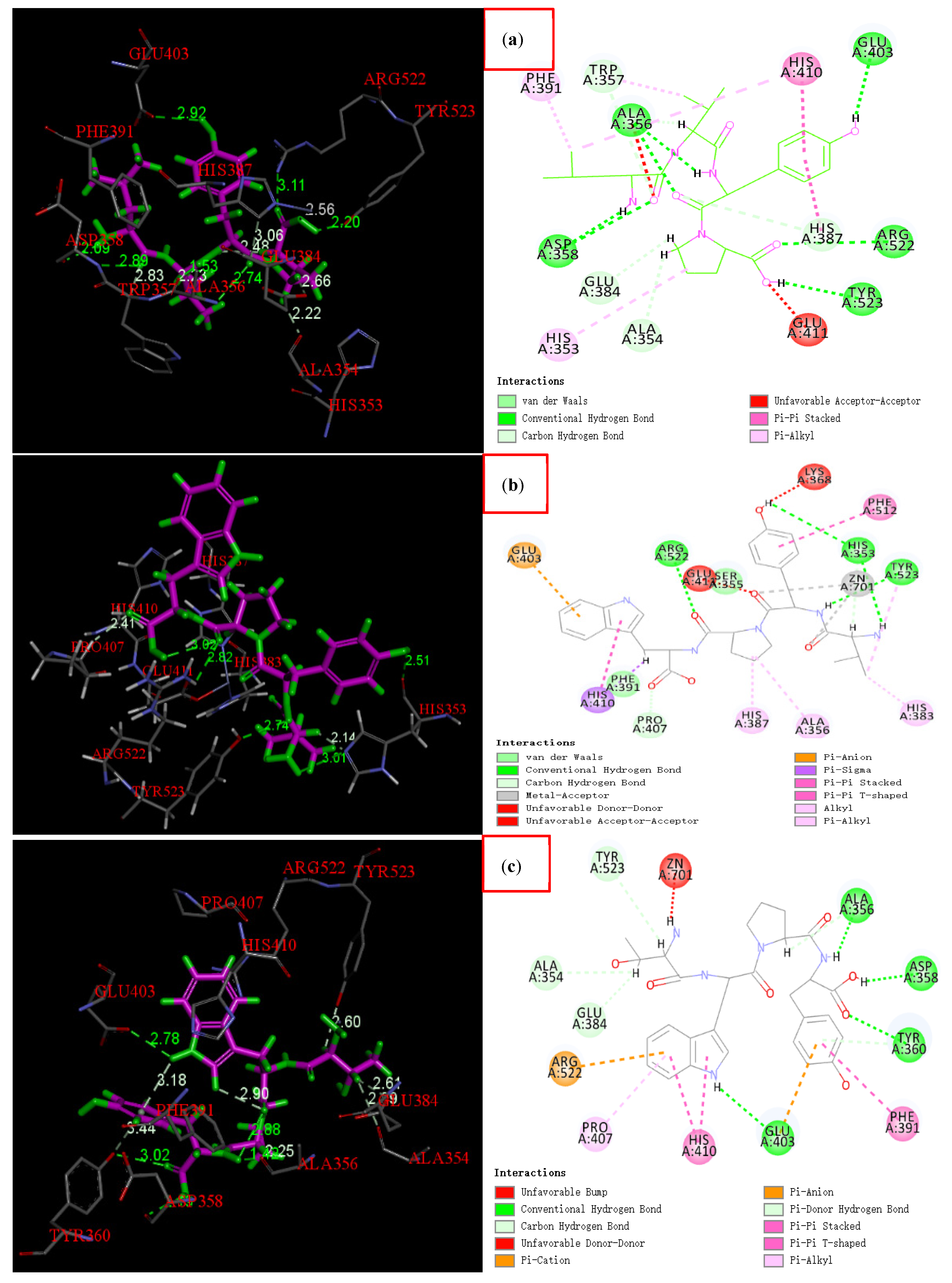
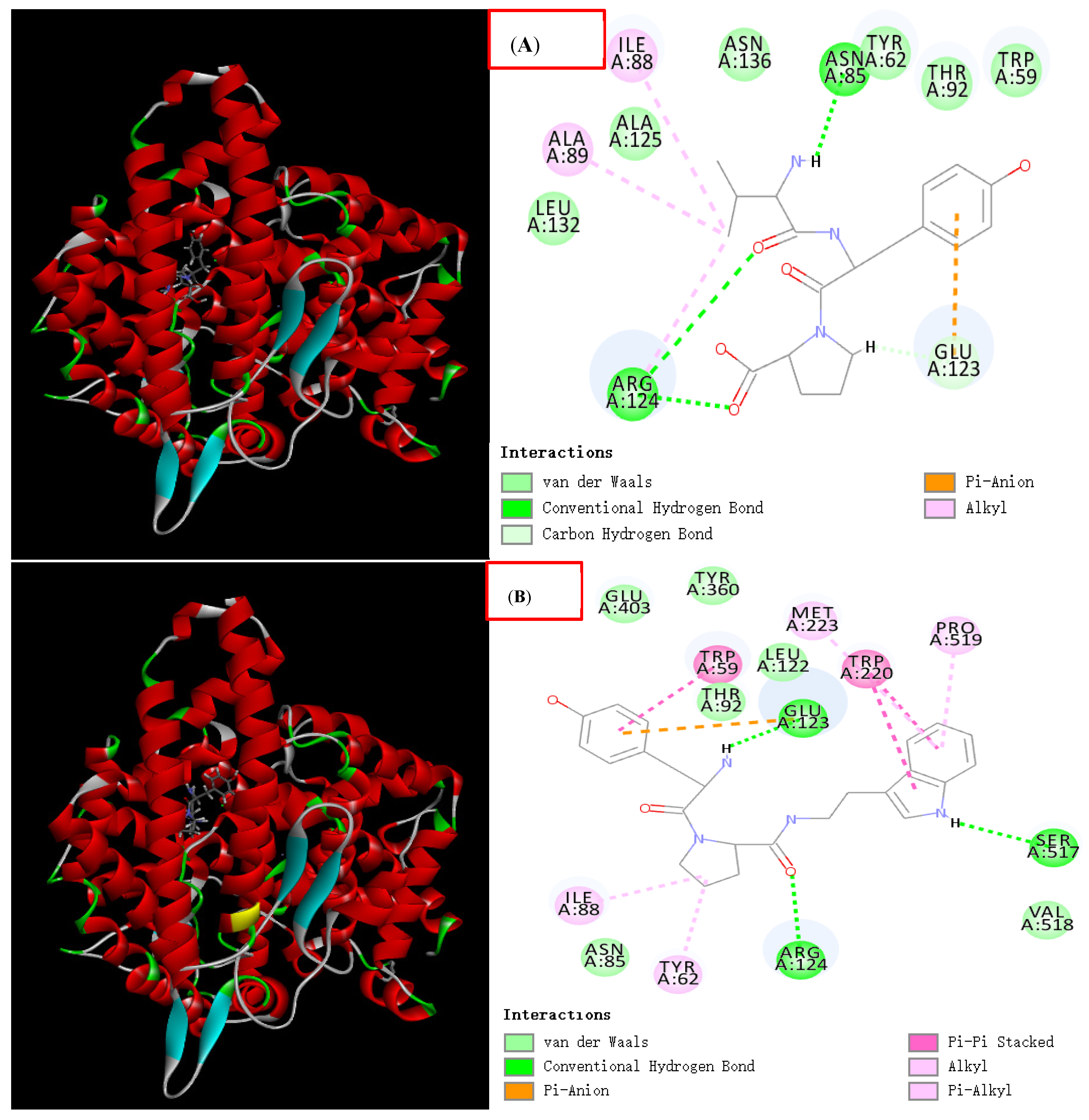
3.7. Molecular Docking
3.8. Stability Verification of ACE Inhibitory Peptides
3.8.1. pH Stability of ACE Inhibitory Peptides
3.8.2. Temperature Stability of ACE Inhibitory Peptides
3.8.3. In Vitro Digestion of ACE Inhibitory Peptides
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berendsen, R.L.; Kalkhove-Stefanie, I.C.; Lugones, L.G.; Baars-Johan, J.P.; Wösten-Han, A.B.; Bakker-Peter, A.H.M. Effects of the mushroom-volatile 1-octen-3-ol on dry bubble disease. Appl. Microbiol. Biotechnol. 2013, 97, 5535–5543. [Google Scholar] [CrossRef]
- Kimatu, B.M.; Zhao, L.Y.; Biao, Y.; Ma, G.X.; Yang, W.J.; Fei, P.; Hu, Q.H. Antioxidant potential of edible mushroom (A. bisporus.) protein hydrolysates and their ultrafiltration fractions. Food Chem. 2017, 230, 58–67. [Google Scholar] [CrossRef]
- Shen, Q. Study on Drying Characteristics and Extraction of Flavor Nucleotides from Agaricus bisporus Waste Stipe. Master’s Thesis, Henan Agricultural University, Henan, China, 2014. [Google Scholar]
- Taghizadeh, S.F.; Azizi, M.; Asili, J.; Madarshahi, F.S.; Rakhshandeh, H.; Fujii, Y. Therapeutic peptides of Mucuna pruriens L.: Anti-genotoxic molecules against human hepatocellular carcinoma and hepatitis C virus. Food Sci. Nutr. 2021, 9, 2908–2914. [Google Scholar] [CrossRef]
- Liu, C.L.; Fang, L.; Min, W.H.; Liu, J.S.; Li, H.M. Exploration of the molecular interactions between angiotensin-I-converting enzyme (ACE) and the inhibitory peptides derived from hazelnut (Corylus heterophylla Fisch). Food Chem. 2018, 245, 471–480. [Google Scholar] [CrossRef]
- Zhang, T.; Li, M.; Fu, X.D.; Mou, H.J. Purification and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides with specific structure X-Pro. Eur. Food Res. Technol. 2019, 245, 1743–1753. [Google Scholar] [CrossRef]
- Xiang, L.; Qiu, Z.C.; Zhao, R.J.; Zheng, Z.J.; Qiao, X.G. Advancement and prospects of production, transport, functional activity and structure-activity relationship of food-derived angiotensin converting enzyme (ACE) inhibitory peptides. Crit. Rev. Food Sci. Nutr. 2021, PP, 21–27. [Google Scholar] [CrossRef]
- Gen, X.R.; Tian, G.T.; Zhang, W.W.; Zhao, Y.C.; Zhao, L.Y.; Wang, H.X. A Tricholoma matsutake Peptide with Angiotensin Converting Enzyme Inhibitory and Antioxidative Activities and Antihypertensive Effects in Spontaneously Hypertensive Rats. Sci. Rep. 2016, 6, 24130. [Google Scholar] [CrossRef] [Green Version]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptistam, J. Isolation and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides from Ulva rigida C. Agardh protein hydrolysate. J. Funct. Foods. 2016, 26, 65–76. [Google Scholar] [CrossRef]
- Kang, M.G.; Kim, Y.H.; Boloramaa, Z.; Kim, M.K.; Seo, G.S.; Lee, J.S. Characterization of an antihypertensive angiotensin I-converting enzyme inhibitory peptide from the edible mushroom Hypsizygus marmoreus. BioMed. Res. Int. 2013, 2013, 283964. [Google Scholar] [CrossRef]
- Tran, H.B.; Yamsmoto, A.; Matsumoto, S.; Ito, H.; Igami, K.; Miyazaki, T.; Kondo, R.; Shimizu, K. Hypotensive effects and angiotensin-converting enzyme inhibitory peptides of reishi (Ganoderma lingzhi) auto- digested extract. Molecules 2014, 19, 13473–13485. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.G.; Sum, H.N.; Sun, J.H.; Liao, D.K. Technical optimization for extracting hypotensive active peptides from Agrocybe aegerita. J. South. Med. Univ. 2010, 30, 1264–1267. [Google Scholar]
- Zhao, X.R. Studies of Triterpenoids of Ganoderma Lucidum and Their Bioactivities. Master’s Thesis, Dalian Medical University, Dalian, China, 2016. [Google Scholar]
- Jang, J.H.; Jeong, S.C.; Kim, J.H.; Lee, Y.H. Characterisation of a new antihypertensive angiotensin I-converting enzyme inhibitory peptide from Pleurotus cornucopiae. Food Chem. 2010, 127, 412–418. [Google Scholar] [CrossRef]
- Gen, X.R. Studies on Purification, Structure and Biological Activities Ofangiotensin Converting Enzyme Inhibitors from Tricholoma Matsutake and Leiicopaxillus Tricolor. Ph.D. Thesis, China Agricultural Universal, Beijing, China, 2016. [Google Scholar]
- Guo, P.H.; Gao, D.D.; Chang, S.H.; Hou, F.J. Optimizing double-enzymatic hydrolysis conditions of alfalfa protein for the preparation of ACE inhibitory peptides. Pratac. Sci. 2020, 37, 1627–1637. [Google Scholar] [CrossRef]
- Wu, W.F.; Jia, J.; Wen, C.R.; Yu, C.P.; Zhao, Q.; Hu, J.N. Optimization of ultrasound assisted extraction of abalone viscera protein and its effect on the iron-chelating activity. Ultrason. Sonochem. 2021, 77, 105670. [Google Scholar] [CrossRef]
- Samah, M.S.; Mila, Z.; Jeanette, O. Performance of two commonly used angiotensin-converting enzyme inhibition assays using FA-PGG and HHL as substrates. J. Dairy Res. 2006, 73, 178–186. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzene sulfonic acid. J. Agric. Food Chem. 1979, 27, 59–65. [Google Scholar] [CrossRef]
- Deng, H.L.; Zheng, J.K.; An, J.Q. Macroporous Resin Adsorption for Separation and Purification of ACE Inhibitory Peptides from Porcine Hemoglobin. Food Sci. 2013, 34, 27–31. [Google Scholar] [CrossRef]
- Vermeirssen, V.; Camp, J.V.; Vertraete, W. Optimization and validation of an angiotensin-converting enzyme inhibition assay for the screening of bioactive peptides. J. Biochem. Biophys. Methods 2002, 51, 75–87. [Google Scholar] [CrossRef]
- Yu, W.C.; Gao, J.; Xue, Z.H.; Kou, X.H.; Wang, Y.F.; Zhai, L.J. Radical-scavenging activity, ACE-inhibiting capability and identification of rapeseed albumin hydrolysate. Food Sci. Hum. Well. 2013, 2, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.D.; Fang, L.J.; Ma, P.J. The angiotensin-converting enzyme inhibitor, captopril, suppressed Hepatic Stellate Cell activation via NF-kappaB or wnt3α/β-catenin pathway. Bioengineered 2021, 12, 8370–8377. [Google Scholar] [CrossRef]
- Zhang, P.; Chang, C.; Liu, H.J.; Li, B.; Yan, Q.J.; Jiang, Z.Q. Identification of novel angiotensin I-converting enzyme (ACE) inhibitory peptides from wheat gluten hydrolysate by the protease of Pseudomonas aeruginosa. J. Funct. Foods 2020, 65, 103751. [Google Scholar] [CrossRef]
- Salampessy, J.; Reddy, N.; Phillips, M.; Kailasapathy, K. Isolation and characterization of nutraceutically potential ACE -Inhibitory peptides from leatherjacket (Mechanic sp.) protein hydrolysates. LWT-Food Sci. Technol. 2017, 80, 430–436. [Google Scholar] [CrossRef]
- Xie, J.L.; Chen, X.J.; Wu, J.J.; Zhang, Y.Y.; Zhou, Y.; Zhang, L.J.; Tang, Y.J.; Wei, D.Z. Antihypertensive effects, molecular docking study and isothermal titration calorimetry assay of the angiotensin I-converting enzyme inhibitory peptides from Chlorella vulgaris. J. Agric. Food Chem. 2018, 66, 1359–1368. [Google Scholar] [CrossRef]
- Pan, D.D.; Cao, J.X.; Guo, H.Q.; Zhao, B. Studies on purification and the molecular mechanism of a novel ACE inhibitory peptide from whey protein hydrolysate. Food Chem. 2012, 130, 121–126. [Google Scholar] [CrossRef]
- Bah, C.S.; Carne, A.; McConnell, M.A.; Michelle, A.; El-Din, A. Bekhita Production of bioactive peptide hydrolysates from deer, sheep, pig and cattle red blood cell fractions using plant and fungal protease preparations. Food Chem. 2016, 202, 458–466. [Google Scholar] [CrossRef]
- Piovesana, S.; Capriotti, A.L.; Cavaliere, C.; Giorgia, L.B.; Maria, M.C.; Riccardo, Z.C.; Laganà, A. Recent trends and analytical challenges in plant bioactive peptide separation, identification and validation. Anal. Bioanal. Chem. 2018, 410, 1–20. [Google Scholar] [CrossRef]
- Yang, F.J.; Chen, X.; Huang, M.C.; Yang, Q.; Cai, X.X.; Chen, X.; Du, M.; Huang, J.L.; Wang, S.Y. Molecular characteristics and structure–activity relationships of food-derived bioactive peptides. J. Integr. Agric. 2021, 9, 2313–2332. [Google Scholar] [CrossRef]
- Daskayadikmen, C.; Yucetepe, A.; Funda, K.G.; Hayrettin, D.; Beraat, O. Angiotensin-I-converting enzyme (ACE)-Inhibitory peptides from plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef]
- Rudolph, S.; Lunow, D.; Kaiser, S.; Henle, T. Identification and quantification of ACE-inhibiting peptides in enzymatic hydrolysates of plant proteins. Food Chem. 2017, 224, 19–25. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-originating ACE inhibitors, including antihypertensive peptides, as preventive food components in blood pressure reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Sklenicka, P. Applying evaluation criteria for the land consolidation effect to three contrasting study areas in the Czech Republic. Land Use Policy 2006, 23, 502–510. [Google Scholar] [CrossRef]
- Xu, X.Y. Optimization of Ultrasound-Assisted Enzymatic Preparation of Pleurotus Eryngii Peptides for Lowering Blood Pressure. Master’s Thesis, Shandong Agricultural University, Shandong, China, 2020. [Google Scholar]
- Huang, J.C.; Peng, X.C. Two-step enzymatic hydrolysis for preparation of ACE inhibitory peptides from sardine. Sci. Technol. Food Ind. 2015, 36, 295–300. [Google Scholar] [CrossRef]
- Ishak, N.H.; Shaik, M.I.; Yellapu, N.K.; Howell, N.K.; Sarbon, N.M. Purification, characterization and molecular docking study of angiotensin-I converting enzyme (ACE) inhibitory peptide from shortfin scad (Decapterus macrosoma) protein hydrolysate. J. Food Sci. Technol. 2021, 58, 1–11. [Google Scholar] [CrossRef]
- Pang, G.C.; Chen, Q.S.; Hu, Z.H.; Xie, J.B. Bioactive Peptides: Absorption, Utilization and Functionality. Food Sci. 2013, 34, 375–391. [Google Scholar] [CrossRef]
- Zhang, B.Y. ACE-Renin Inhibitory Mechanism and Absorption Pathway of Egg White Derived Peptides. Ph.D. Thesis, Jilin University, Jilin, China, 2021. [Google Scholar]
- Sarbon, N.M.; Badii, F.; Howell, N.K. Purification and characterization of antioxidative peptides derived from chicken skingelatin hydrolysate. Food Hydrocoll. 2018, 85, 311–320. [Google Scholar] [CrossRef]
- Sarbon, N.M.; Howell, N.K. Angiotensin-I converting enzyme (ACE) inhibitory peptides from chicken skin gelatin hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Int. Food Res. J. 2019, 26, 903–911. [Google Scholar]
- Raquel, N.; Michael, L.; Wolfgang, L. Alternative high-performance liquid chromatographic peptide separation and purification concept using a new mixed-mode reversed-phase/weak anion-exchange type stationary phase. J. Chromatogr. 2005, 1089, 158–169. [Google Scholar] [CrossRef]
- Razavi, R.; Amiri, M.; Alshamsi, H.A.; Eslaminejad, T.; Salavati-Niasari, M. Green synthesis of agnanoparticles in oil-in-water nano-emulsion and evaluation of their antibacterial and cytotoxic properties as well as molecular docking. Arabian. J. Chem. 2021, 14, 1878–5352. [Google Scholar] [CrossRef]
- Fan, H.B.; Wang, J.P.; Liao, W. Identification and characterization of gastrointestinal-resistant angiotensin-converting enzyme inhibitory peptides from egg white proteins. J. Agric. Food Chem. 2019, 67, 7147–7156. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Aluko, R.E. Kinetics and molecular docking studies of the inhibitions of angiotensin converting enzyme and renin activities by hemp seed (Cannabis sativa L.) peptides. J. Agric. Food Chem. 2014, 62, 4135–4144. [Google Scholar] [CrossRef]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor Level | Factor | ||
|---|---|---|---|
| Temperature of Enzymatic Digestion | pH | Time of Enzymatic Digestion | |
| −1 | 40 °C | 8.0 | 3.0 |
| 0 | 45 °C | 8.5 | 3.5 |
| +1 | 50 °C | 9.0 | 4.0 |
| Serial Number | pH | Temperature/°C | Time/h | Inhibition Rate of ACE/% |
|---|---|---|---|---|
| 1 | 8.00 | 45.00 | 4.00 | 55.43 |
| 2 | 8.50 | 40.00 | 4.00 | 53.64 |
| 3 | 8.50 | 45.00 | 3.50 | 68.45 |
| 4 | 8.00 | 45.00 | 3.00 | 54.87 |
| 5 | 9.00 | 40.00 | 3.50 | 50.46 |
| 6 | 8.50 | 50.00 | 4.00 | 53.27 |
| 7 | 9.00 | 45.00 | 3.00 | 47.38 |
| 8 | 8.50 | 40.00 | 3.00 | 56.68 |
| 9 | 8.00 | 50.00 | 3.50 | 50.27 |
| 10 | 8.50 | 45.00 | 3.50 | 69.82 |
| 11 | 8.50 | 50.00 | 3.00 | 46.29 |
| 12 | 8.50 | 45.00 | 3.50 | 68.65 |
| 13 | 8.50 | 45.00 | 3.50 | 70.49 |
| 14 | 8.00 | 40.00 | 3.50 | 55.36 |
| 15 | 9.00 | 45.00 | 4.00 | 51.48 |
| 16 | 8.50 | 45.00 | 3.50 | 69.47 |
| 17 | 9.00 | 50.00 | 3.50 | 44.26 |
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F Value | p-Value | Significance |
|---|---|---|---|---|---|---|
| Model | 1288.34 | 9 | 143.15 | 337.78 | <0.0001 | *** |
| A | 62.44 | 1 | 62.44 | 147.34 | <0.0001 | *** |
| B | 60.78 | 1 | 60.78 | 143.41 | <0.0001 | *** |
| C | 9.24 | 1 | 9.24 | 21.81 | 0.0023 | ** |
| AB | 0.31 | 1 | 0.31 | 0.73 | 0.4221 | |
| AC | 3.13 | 1 | 3.13 | 7.39 | 0.0298 | * |
| BC | 25.10 | 1 | 25.10 | 59.23 | 0.0001 | *** |
| A2 | 398.97 | 1 | 398.97 | 941.43 | <0.0001 | *** |
| B2 | 384.35 | 1 | 384.35 | 906.94 | <0.0001 | *** |
| C2 | 227.57 | 1 | 227.57 | 536.99 | <0.0001 | *** |
| Residual | 2.97 | 7 | 0.42 | |||
| Lack of Fit | 0.14 | 3 | 0.045 | 0.064 | 0.9764 | |
| Pure Error | 2.83 | 4 | 0.71 | |||
| Cor Total | 1291.31 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Yun, J.; Wu, S.; Bi, Y.; Zhao, F. Optimisation and Characterisation of Novel Angiotensin-Converting Enzyme Inhibitory Peptides Prepared by Double Enzymatic Hydrolysis from Agaricus bisporus Scraps. Foods 2022, 11, 394. https://doi.org/10.3390/foods11030394
Wang R, Yun J, Wu S, Bi Y, Zhao F. Optimisation and Characterisation of Novel Angiotensin-Converting Enzyme Inhibitory Peptides Prepared by Double Enzymatic Hydrolysis from Agaricus bisporus Scraps. Foods. 2022; 11(3):394. https://doi.org/10.3390/foods11030394
Chicago/Turabian StyleWang, Rui, Jianmin Yun, Shujuan Wu, Yang Bi, and Fengyun Zhao. 2022. "Optimisation and Characterisation of Novel Angiotensin-Converting Enzyme Inhibitory Peptides Prepared by Double Enzymatic Hydrolysis from Agaricus bisporus Scraps" Foods 11, no. 3: 394. https://doi.org/10.3390/foods11030394