
Aged Pericarpium Citri Reticulatae ‘Chachi’ Attenuates Oxidative Damage Induced by tert-Butyl Hydroperoxide (t-BHP) in HepG2 Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Samples
2.2. Chemicals and Reagents
2.3. Sample Preparation
2.4. Extract Preparation of Free and Bound Phenolics
2.5. Determination of Total Phenolics and Total Flavonoids Concentration
2.6. Determination of Free and Bound Flavonoids in PCR-C
2.7. Purification of Free Flavonoids in PCR-C
2.8. Cell Culture
2.9. MTT Assay
2.10. Cytoprotective Effects
2.11. Measurement of Intracellular ROS Levels
2.12. Assessment of MDA, T-SOD and GSH
2.13. Data Analysis and Statistics
3. Results and Discussion
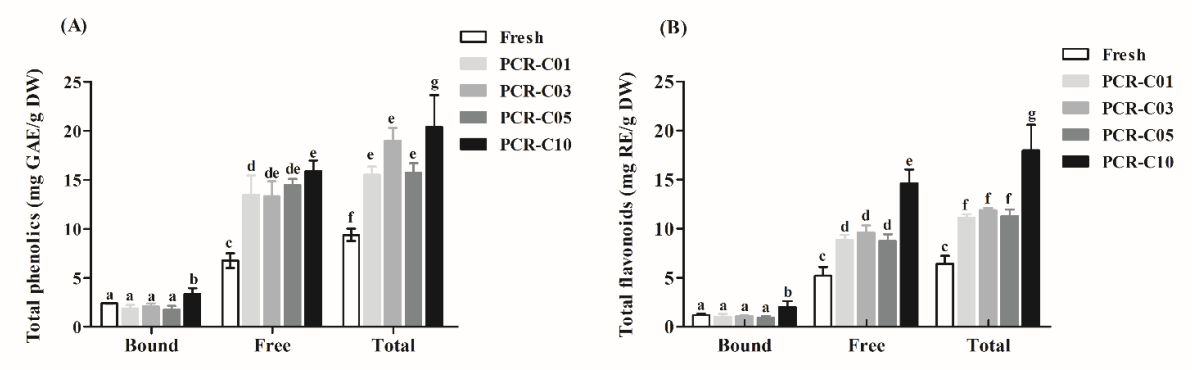
3.1. Measurement of Total Phenolics and Flavonoids Contents in PCR-C during Aging
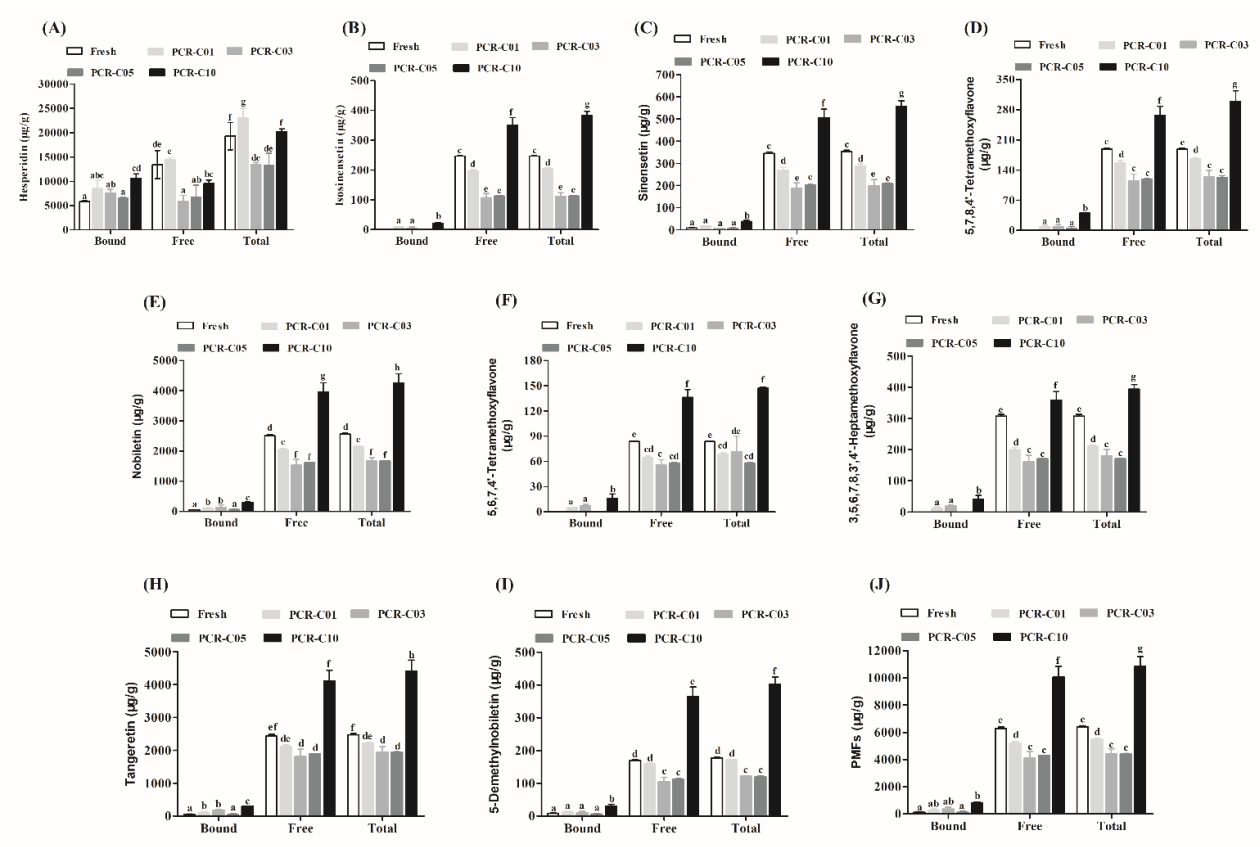
3.2. Tracing the Variation of Free and Bound Flavonoids in PCR-C during Aging
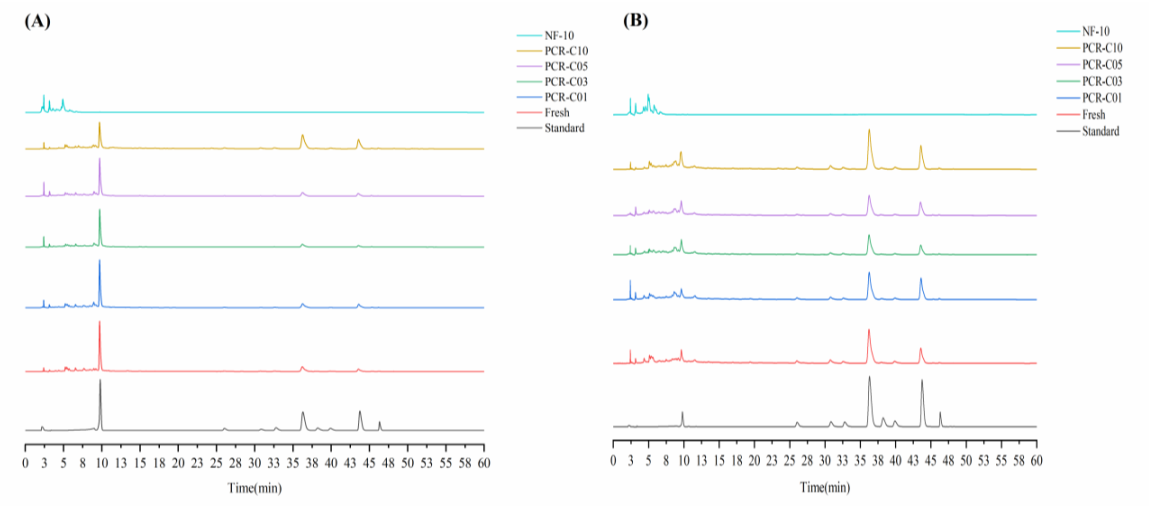
3.3. Purification of Free Flavonoids in PCR-C
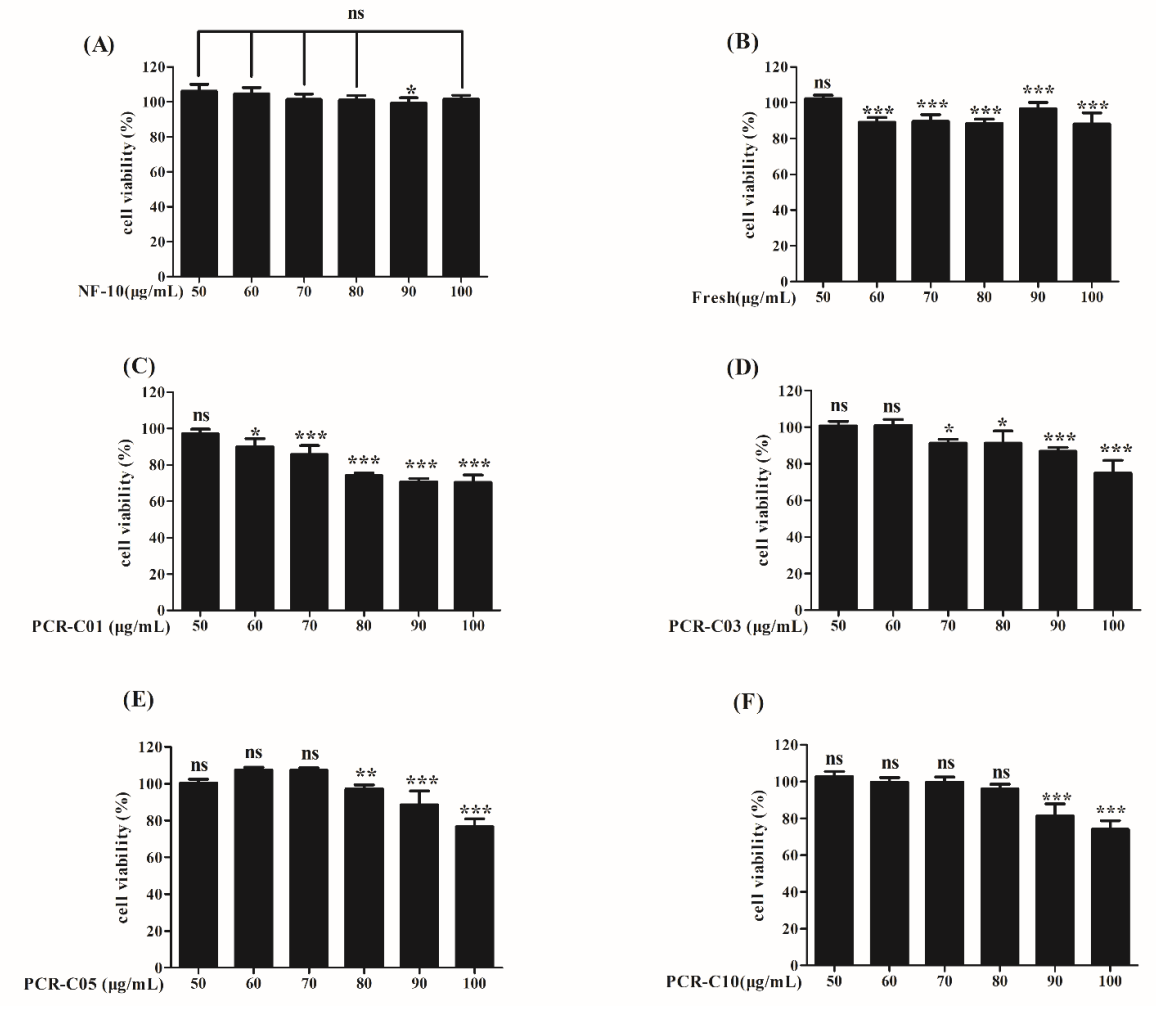
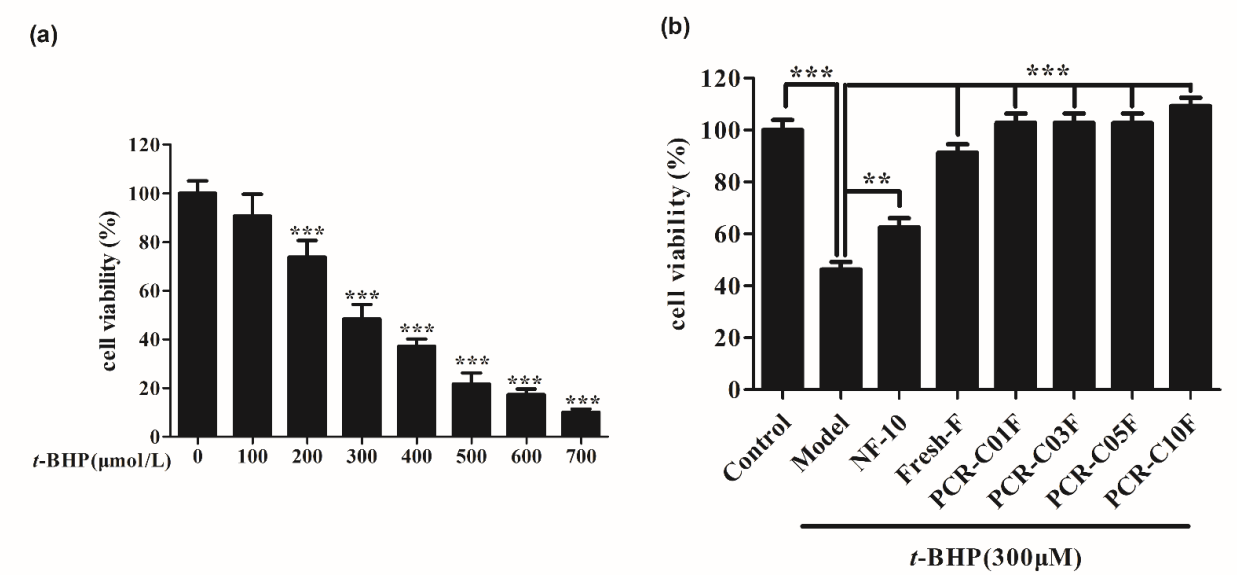
3.4. Cell Viability
3.5. Purified PCR-C Samples Attenuated t-BHP-Induced Cytotoxicity
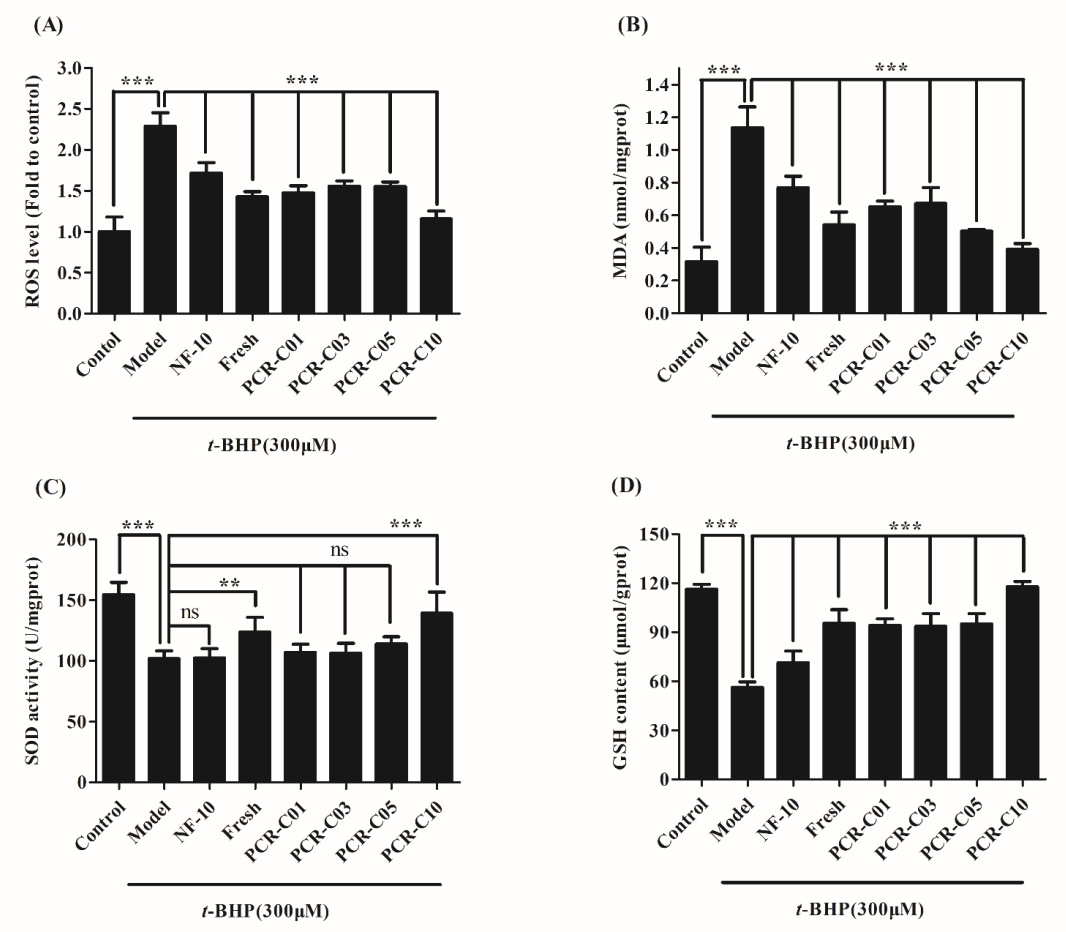
3.6. Purified PCR-C Samples Attenuated t-BHP-Induced Oxidative Damage
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liu, H.; Qiu, N.; Ding, H.; Yao, R. Polyphenols contents and antioxidant capacity of 68 Chinese herbals suitable for medical or food uses. Food Res. Int. 2008, 41, 363–370. [Google Scholar] [CrossRef]
- Lv, W.S.; Lin, T.; Ren, Z.Y.; Jiang, Y.Q.; Zhang, J.; Bi, F.J.; Gu, L.H.; Hou, H.C.; He, J.N. Rapid discrimination of Citrus reticulata ‘Chachi’ by headspace-gas chromatography-ion mobility spectrometry fingerprints combined with principal component analysis. Food Res. Int. 2020, 131, 108985. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, G.; Fu, X.; Liu, R.H. Effects of aging on the phytochemical profile and antioxidative activity of Pericarpium Citri Reticulatae ‘Chachiensis’. Rsc. Adv. 2016, 6, 105272–105281. [Google Scholar] [CrossRef]
- Choi, M.-Y.; Chai, C.; Park, J.H.; Lim, J.; Lee, J.; Kwon, S.W. Effects of storage period and heat treatment on phenolic compound composition in dried Citrus peels (Chenpi) and discrimination of Chenpi with different storage periods through targeted metabolomic study using HPLC-DAD analysis. J. Pharm. Biomed 2011, 54, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosti, V.; Bertozzi, B.; Fontana, L. Health Benefits of the Mediterranean Diet: Metabolic and Molecular Mechanisms. J. Gerontol. A-Biol. 2018, 73, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Wang, J.; Huang, X.H.; Tu, Y.; Ni, K.Y. Identification of polymethoxylated flavones from green tangerine peel (Pericarpium Citri Reticulatae Viride) by chromatographic and spectroscopic techniques. J. Pharm. Biomed 2007, 44, 63–69. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Y.; Shen, S.; Zhi, Z.; Cheng, H.; Chen, S.; Ye, X. Antioxidant and pancreatic lipase inhibitory effects of flavonoids from different citrus peel extracts: An in vitro study. Food Chem. 2020, 326, 126785. [Google Scholar] [CrossRef]
- Xican, L.; Yanping, H.; Dongfeng, C. Protective Effect against Hydroxyl-induced DNA Damage and Antioxidant Activity of Citri reticulatae Pericarpium. Adv. Pharm. Bull. 2013, 3, 175–181. [Google Scholar]
- Wang, H.; Chen, G.; Guo, X.B.; Abbasi, A.M.; Liu, R.H. Influence of the stage of ripeness on the phytochemical profiles, antioxidant and antiproliferative activities in different parts of Citrus reticulata Blanco cv. Chachiensis. LWT-Food Sci. Technol. 2016, 69, 67–75. [Google Scholar] [CrossRef]
- Fu, M.Q.; Xu, Y.J.; Chen, Y.L.; Wu, J.J.; Yu, Y.S.; Zou, B.; An, K.J.; Xiao, G.S. Evaluation of bioactive flavonoids and antioxidant activity in Pericarpium Citri Reticulatae (Citrus reticulata ‘Chachi’) during storage. Food Chem. 2017, 230, 649–656. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Structure-activity relationships of flavonoids in the cellular antioxidant activity assay. J. Agric. Food Chem. 2008, 56, 8404–8411. [Google Scholar] [CrossRef] [PubMed]
- Alia, M.; Ramos, S.; Mateos, R.; Bravo, L.; Goya, L. Response of the antioxidant Defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HepG2). J. Biochem. Mol. Toxicol. 2005, 19, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Chen, Z.; Darwish, W.S.; Terada, K.; Chiba, H.; Hui, S.P. Choline and Ethanolamine Plasmalogens Prevent Lead-Induced Cytotoxicity and Lipid Oxidation in HepG2 Cells. J. Agric. Food Chem. 2019, 67, 7716–7725. [Google Scholar] [CrossRef]
- Kimatu, B.M.; Fang, D.L.; Zhao, L.Y.; Hu, Q.H. Agaricus bisporuspeptide fractions confer cytoprotective ability against hydrogen peroxide-induced oxidative stress in HepG2 and Caco-2 cells. J. Food Meas Charact. 2020, 14, 2503–2519. [Google Scholar] [CrossRef]
- Liang, F.Q.; Cao, W.W.; Huang, Y.T.; Fang, Y.J.; Cheng, Y.X.; Pan, S.Y.; Xu, X.Y. Isoflavone biochanin A, a novel nuclear factor erythroid 2-related factor 2 (Nrf2)-antioxidant response element activator, protects against oxidative damage in HepG2 cells. Biofactors 2019, 45, 563–574. [Google Scholar] [CrossRef]
- Goya, L.; Martin, M.A.; Ramos, S.; Mateos, R.; Bravo, L. A Cell Culture Model for the Assessment of the Chemopreventive Potential of Dietary Compounds. Curr. Nutr. Food Sci. 2009, 5, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Min, B.; Gu, L.; McClung, A.M.; Bergman, C.J.; Chen, M.-H. Free and bound total phenolic concentrations, antioxidant capacities, and profiles of proanthocyanidins and anthocyanins in whole grain rice (Oryza sativa L.) of different bran colours. Food Chem. 2012, 133, 715–722. [Google Scholar] [CrossRef]
- Wang, C.; Cheng, D.; Cao, J.; Jiang, W. Antioxidant capacity and chemical constituents of Chinese jujube (Ziziphus jujuba Mill.) at different ripening stages. Food Sci. Biotechnol. 2013, 22, 639–644. [Google Scholar] [CrossRef]
- Zunli, K.; Ziyan, Z.; Yuanyuan, Z.; Xiaodan, X.; Yin, L.; Si, T.; Cheng, H.; Zhiqin, Z. PMFs-rich Citrus extract prevents the development of non-alcoholic fatty liver disease in C57BL/6J mice induced by a high-fat diet. J. Funct. Foods 2018, 47, 28–39. [Google Scholar]
- Zhang, X.; Jiang, L.; Geng, C.; Hu, C.; Yoshimura, H.; Zhong, L. Inhibition of Sudan I genotoxicity in human liver-derived HepG2 cells by the antioxidant hydroxytyrosol. Free Radic. Res. 2008, 42, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.D.; Chao, Y.X.; Luo, M.X.; Xie, B.; Zhang, D.D.; Hu, P.J.; Yang, X.J.; Yang, D.P.; Wei, M.Y. Construction and Chemical Profile on “Activity Fingerprint” of Citri Reticulatae Pericarpium from Different Cultivars Based on HPLC-UV, LC/MS-IT-TOF, and Principal Component Analysis. Evid.-Based Compl. Alt. 2020, 2020, 4736152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Zeng, W.; Huang, K.E.; Li, D.X.; Chen, W.; Yu, X.Q.; Ke, X.H. Discrimination of Citrus reticulata Blanco and Citrus reticulata ‘Chachi’ as well as the Citrus reticulata ‘Chachi’ within different storage years using ultra high performance liquid chromatography quadrupole/time-of-flight mass spectrometry based metabolomics approach. J. Pharm. BioMed 2019, 171, 218–231. [Google Scholar]
- Zhao, L.H.; Zhao, H.Z.; Zhao, X.; Kong, W.J.; Hu, Y.C.; Yang, S.H.; Yang, M.H. Simultaneous Quantification of Seven Bioactive Flavonoids in Citri Reticulatae Pericarpium by Ultra-Fast Liquid Chromatography Coupled with Tandem Mass Spectrometry. Phytochem. Anal. 2016, 27, 168–173. [Google Scholar] [CrossRef]
- Zheng, G.D.; Yang, D.P.; Wang, D.M.; Zhou, F.; Yang, X.; Jiang, L. Simultaneous Determination of Five Bioactive Flavonoids in Pericarpium Citri Reticulatae from China by High-Performance Liquid Chromatography with Dual Wavelength Detection. J. Agric. Food Chem. 2009, 57, 6552–6557. [Google Scholar] [CrossRef]
- Liu, E.H.; Zhao, P.; Duan, L.; Zheng, G.D.; Guo, L.; Yang, H.; Li, P. Simultaneous determination of six bioactive flavonoids in Citri Reticulatae Pericarpium by rapid resolution liquid chromatography coupled with triple quadrupole electrospray tandem mass spectrometry. Food Chem. 2013, 141, 3977–3983. [Google Scholar] [CrossRef]
- Yi, L.Z.; Yuan, D.L.; Liang, Y.Z.; Xie, P.S.; Zhao, Y. Fingerprinting alterations of secondary metabolites of tangerine peels during growth by HPLC-DAD and chemometric methods. Anal. Chim. Acta 2009, 649, 43–51. [Google Scholar] [CrossRef]
- Aidoo, M.K.; Sherman, T.; Lazarovitch, N.; Fait, A.; Rachmilevitch, S. A bell pepper cultivar tolerant to chilling enhanced nitrogen allocation and stress-related metabolite accumulation in the roots in response to low root-zone temperature. Physiol. Plant. 2017, 161, 196–210. [Google Scholar] [CrossRef]
- He, J.; Chen, C.C.; He, Q.X.; Li, J.Y.; Ying, F.; Chen, G. The central bacterial community in Pericarpium Citri Reticulatae ‘Chachiensis’. Food Res. Int. 2019, 125, 108624. [Google Scholar] [CrossRef]
- Cui, L.; Wu, S.Q.; Zhao, C.A.; Yin, C.R. Microbial conversion of major ginsenosides in ginseng total saponins by Platycodon grandiflorum endophytes. J. Ginseng Res. 2016, 40, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Xu, C.C.; Li, Z.; Li, J.; Dai, Y.L.; Han, H.X.; Yu, S.S.; Liu, S.Y. Microbial conversion of ginsenoside Rd from Rb1 by the fungus mutant Aspergillus niger strain TH-10a. Prep. Biochem. Biotech. 2016, 46, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.-H.; Chan, Y.-F.; Pan, M.-H.; Tung, Y.-C.; Su, Z.-Y. Aged Citrus Peel (Chenpi) Prevents Acetaminophen-Induced Hepatotoxicity by Epigenetically Regulating Nrf2 Pathway. Am. J. Chin. Med. 2019, 47, 1833–1851. [Google Scholar] [CrossRef] [PubMed]
- Qiaoyu, T.; Yifeng, Z.; Ting, Y.A.N. Study on Purification Technology of Flavonoids from Fortunella margarita Peel Using HPD300 Macropomus Resin. Food Sci. 2008, 29, 355–358. [Google Scholar]
- Jannat, S.; Ali, M.Y.; Kim, H.-R.; Jung, H.A.; Choi, J.S. Protective Effects of Sweet Orange, Unshiu Mikan, and Mini Tomato Juice Powders on t-BHP-Induced Oxidative Stress in HepG2 Cells. Prev. Nutr. Food Sci. 2016, 21, 208–220. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.-L.; Yen, G.-C. Neuroprotective effects of the citrus flavanones against H2O2-induced cytotoxicity in PC12 cells. J. Agric. Food Chem. 2008, 56, 859–864. [Google Scholar] [CrossRef]
- Liang, F.Q.; Fang, Y.J.; Cao, W.W.; Zhang, Z.; Pan, S.Y.; Xu, X.Y. Attenuation of tert-Butyl Hydroperoxide (t-BHP)-Induced Oxidative Damage in HepG2 Cells by Tangeretin: Relevance of the Nrf2-ARE and MAPK Signaling Pathways. J. Agric. Food Chem. 2018, 66, 6317–6325. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, J.; Zhang, Y.T.; Su, H.; Qiu, X.H.; Gong, L.; Huang, J.; Bai, J.Q.; Huang, Z.H.; Xu, W. Chemical and genetic discrimination of commercial Guangchenpi (Citrus reticulata ‘Chachi’) by using UPLC-QTOF-MS/MS based metabolomics and DNA barcoding approaches. RSC Adv. 2019, 9, 23373–23381. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Amarsanaa, K.; Wu, J.; Jeon, S.-C.; Cui, Y.; Jung, S.-C.; Park, D.-B.; Kim, S.-J.; Han, S.-H.; Kim, H.-W.; et al. Nobiletin attenuates neurotoxic mitochondrial calcium overload through K+ influx and Delta psi(m) across mitochondrial inner membrane. Korean J. Physiol. Pharmacol. 2018, 22, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-K.; Kim, S.-I.; Oh, S.Y.; Na, W.; Kang, Y.-H. Tangeretin Ameliorates Glucose-Induced Podocyte Injury through Blocking Epithelial to Mesenchymal Transition Caused by Oxidative Stress and Hypoxia. Int. J. Mol. Sci. 2020, 21, 8577. [Google Scholar] [CrossRef]
- Mateos, R.; Goya, L.; Bravo, L. Determination of malondialdehyde by liquid chromatography as the 2,4-dinitrophenylhydrazone derivative—A marker for oxidative stress in cell cultures of human hepatoma HepG2. J. Chromatogr. B-Anal. Technol. Biomed. Life Sci. 2004, 805, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Suttnar, J.; Masova, L.; Dyr, J.E. Influence of citrate and EDTA anticoagulants on plasma malondialdehyde concentrations estimated by high-performance liquid chromatography. J. Chromatogr. B 2001, 751, 193–197. [Google Scholar] [CrossRef]
- Wang, B.; Peng, L.; Zhu, L.; Ren, P. Protective effect of total flavonoids from Spirodela polyrrhiza (L.) Schleid on human umbilical vein endothelial cell damage induced by hydrogen peroxide. Colloids Surf. B-Biointerfaces 2007, 60, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhao, J.T.; Zu, Y.G.; Fu, Y.J.; Liang, L.; Luo, M.; Wang, W.; Efferth, T. Antioxidant properties, superoxide dismutase and glutathione reductase activities in HepG2 cells with a fungal endophyte producing apigenin from pigeon pea Cajanus cajan (L.) Millsp. Food Res. Int. 2012, 49, 147–152. [Google Scholar] [CrossRef]
- Kobayashi, S.; Wakl, T.; Nakanish, I.; Matsumoto, K.-I.; Anzai, K. Potent 2,2-Diphenyl-1-picrylhydrazyl Radical-Scavenging Activity of Novel Antioxidants, Double-Stranded Tyrosine Residues Conjugating Pyrocatechol. Chem. Pharm. Bull. 2010, 58, 1442–1446. [Google Scholar] [CrossRef] [Green Version]
- Joko, S.; Watanabe, M.; Fuda, H.; Takeda, S.; Furukawa, T.; Hui, S.-P.; Shrestha, R.; Chiba, H. Comparison of chemical structures and cytoprotection abilities between direct and indirect antioxidants. J. Funct. Foods 2017, 35, 245–255. [Google Scholar] [CrossRef]
- Swetha, S.; Rajitha, S.; Ogutu, S.; Walker, L.T.; Verghese, M. Antioxidant activity of papaya seed extracts against H2O2 induced oxidative stress in HepG2 cells. LWT-Food Sci. Technol. 2016, 66, 293–297. [Google Scholar]
- Zong-Tsi, C.; Heuy-Ling, C.; Charng-Cherng, C.; Chin-Chen, C.; Pin-Der, D. Protective effects of sweet orange (Citrus sinensis) peel and their bioactive compounds on oxidative stress. Food Chem. 2012, 135, 2119–2127. [Google Scholar]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Wojdylo, A.; Oszmianski, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- He, Z.X.; Li, X.F.; Chen, R.; He, K.; Liu, Y.M.; Gong, J.H.; Gong, J.P. Nobiletin attenuates lipopolysaccharide/D-galactosamine-induced liver injury in mice by activating the Nrf2 antioxidant pathway and subsequently inhibiting NF-kappa B-mediated cytokine production. Mol. Med. Rep. 2016, 14, 5595–5600. [Google Scholar] [CrossRef] [Green Version]
- Lakshmi, A.; Subramanian, S.P. Tangeretin ameliorates oxidative stress in the renal tissues of rats with experimental breast cancer induced by 7,12-dimethylbenz[a] anthracene. Toxicol. Lett. 2014, 229, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.-X.; Wei, X.; Li, A.-L.; Wang, A.-M.; Lu, L.-Z.; Yang, Y.; Ren, D.-M.; Wang, X.-N.; Wen, X.-S.; Lou, H.-X.; et al. Screening of traditional Chinese medicines with therapeutic potential on chronic obstructive pulmonary disease through inhibiting oxidative stress and inflammatory response. BMC Complementary Altern. Med. 2016, 16, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT (min) | Molecular Formula | [M + H]+ (m/z) (Error, ppm) | Fragment Ions in the Positive Ion Mode | [M-H]− (m/z) (Error, ppm) | Fragment Ions in the Negative Ion Mode | Identification |
|---|---|---|---|---|---|---|---|
| 1 | 5.563 | C27H30O15 | 595.1665 (−1.27) | 541; 481; 457; 409; 379; 337; 325; 295 | 593.1520 (−1.34) | 503; 473; 455; 383; 353 | Vicenin-2 |
| 2 | 5.974 | C28H32O16 | 625.1769 (−1.03) | 409; 379; 367; 355; 337; 325; 151 | 623.1621 (−0.58) | 533; 503; 413; 383 | Diosmetin-6,8-di-C-glucoside |
| 3 | 9.022 | C22H22O11 | 463.1238 (−1.26) | 445; 427; 367; 343; 313 | 461.1095 (−1.26) | 371; 341; 298 | Diosmetin-8-C-glucoside/Scoparin |
| 4 | 9.278 | C23H24O12 | 493.1345 (−0.92) | 329; 314; 299 | ND | ND | Coccinoside A |
| 5 | 10.167 | C28H34O15 | 611.1984 (−0.03) | 465; 449; 413; 303 | 609.1843 (−0.80) | 301 | Hesperidin |
| 6 | 25.064 | C20H20O8 | 389.1234 (−0.90) | 374; 359; 341 | ND | ND | Monohydroxypentamethoxyflavone (1) |
| 7 | 26.749 | C20H20O7 | 373.1288 (−1.85) | 357; 343; 315; 181 | ND | ND | Isosinensetin |
| 8 | 27.688 | C20H20O8 | 389.1238 (−1.90) | 374; 359; 355; 323; 305 | ND | ND | Monohydroxypentamethoxyflavone (2) |
| 9 | 31.559 | C20H20O7 | 373.1288 (−1.97) | 357; 343; 315; 296; 283; 153 | ND | ND | Sinensetin |
| 10 | 33.255 | C19H18O6 | 343.1182 (−2.12) | 313; 285; 181; 153 | ND | ND | 5,7,8,4′-Tetramethoxyflavone |
| 11 | 36.562 | C21H22O8 | 403.1405 (−1.35) | 373; 355; 327; 211 | ND | ND | Nobiletin |
| 12 | 38.731 | C19H18O6 | 343.1182 (−2.03) | 327; 313; 285; 267; 153 | ND | ND | 5,6,7,4′-Tetramethoxyflavone |
| 13 | 40.827 | C22H24O9 | 433.1501 (−1.83) | 416; 403; 401; 388 | ND | ND | 3,5,6,7,8,3′,4′-Heptamethoxyflavone |
| 14 | 44.243 | C20H20O7 | 373.1288 (−2.06) | 343; 328; 297; 271; 211 | ND | ND | Tangeretin |
| 15 | 46.599 | C20H20O8 | 389.1238 (−1.85) | 373; 359; 341; 313; 197 | ND | ND | 5-Demethylnobiletin |
| 16 | 47.436 | C20H20O9 | 405.1554 (0.84) | 388; 373; 301; 241 | ND | ND | Dihydroxypentamethoxyflavone |
| 17 | 48.127 | C19H18O7 | 359.1129 (−0.91) | 329; 311; 286; 227; 169 | ND | ND | Gardenin B |
| No. | RT (Min) | Molecular Formula | [M + H]+ (m/z) (Error, ppm) | Fragment Ions in the Positive Ion Mode | [M-H]− (m/z) (Error, ppm) | Fragment Ions in the Negative Ion Mode | Identification |
|---|---|---|---|---|---|---|---|
| 1 | 8.113 | C27H30O15 | 595.1660 (−0.38) | 449; 287 | 593.1504 (0.55) | 285 | Luteolin 7-O-rutinoside |
| 2 | 9.005 | C22H22O11 | 463.1232 (0.42) | 445; 427; 367; 343; 313; 177 | ND | ND | Flavone-C-glucoside |
| 3 | 9.41 | C27H30O14 | 579.1720 (0.15) | 433; 271 | 577.1553 (−1.03) | 269 | Isorhoifolin |
| 4 | 9.744 | C28H32O15 | 609.1821 (−1.11) | 301; 286; 177; 153; 71 | 607.1655 (1.03) | 301; 242; 299; 284; 164; 151 | Diosmin |
| 5 | 10.046 | C28H32O15 | 609.1820 (−0.11) | 301; 263;153; 85 | 607.1662 (−0.65) | 301; 299; 284; 164 | Neodiosmin |
| 6 | 10.453 | C22H24O10 | 449.1446 (−1.19) | 431; 413; 369; 303; 263; 195 | ND | ND | Chalcone-C-glycoside |
| 7 | 10.455 | C28H34O15 | 611.1979 (0.02) | 465; 449; 431; 303 | 609.1827 (−0.14) | 301 | Hesperidin |
| 8 | 11.252 | C27H32O14 | 581.1872 (−1.14) | 435; 273; 153 | 579.1719 (0.29) | 271; 151 | Narirutin |
| 9 | 12.493 | C28H34O15 | 611.1979 (0.13) | 465; 303; 177 | 609.1827 (−0.16) | 301; 286; 242; 164; 151 | Neohesperidin |
| 10 | 16.049 | C28H34O14 | 595.2025 (−0.38) | 449; 433; 415; 287; 129; 85 | 593.1866 (1.08) | 285; 270; 164; 151 | Didymin |
| 11 | 19.244 | C28H34O14 | 595.2026 (−0.72) | 449; 443; 287; 161 | 593.1870 (0.97) | 327; 309; 285 | Poncirin |
| 12 | 27.484 | C20H20O7 | 373.1284 (0.14) | 358; 343; 315 | ND | ND | Isosinensetin |
| 13 | 28.375 | C20H20O8 | 389.1228 (0.65) | 374; 359; 341; 313 | ND | ND | Monohydroxypentamethoxyflavone |
| 14 | 32.323 | C20H20O7 | 373.1284 (0.34) | 343; 329; 312; 297;151 | ND | ND | Sinensetin |
| 15 | 34.064 | C19H18O6 | 343.1175 (0.18) | 328; 313; 285; 153 | ND | ND | 5,7,8,4′-Tetramethoxyflavone |
| 16 | 37.922 | C21H22O8 | 403.1394 (−1.49) | 388; 373; 355; 327 | ND | ND | Nobiletin |
| 17 | 39.54 | C19H18O6 | 343.1174 (0.56) | 327; 313; 309; 282; 153 | ND | ND | 5,6,7,4′-Tetramethoxyflavone |
| 18 | 41.631 | C22H24O9 | 433.1490 (0.63) | 417; 403; 385; 357 | ND | ND | 3,5,6,7,8,3′,4′-Heptamethoxyflavone |
| 19 | 44.723 | C20H20O7 | 373.1288 (−1.52) | 343; 325; 297 | ND | ND | Tangeretin |
| 20 | 47.401 | C20H20O8 | 389.1238 (−1.25) | 373; 359; 341; 313; 197 | ND | ND | 5-Demethylnobiletin |
| Fresh | PCR-C01 | PCR-C03 | PCR-C05 | PCR-C10 | |
|---|---|---|---|---|---|
| Total flavonoids (mg RE/g dw) | 102.50 ± 3.94 a | 102.50 ± 4.90 a | 135.58 ± 5.99 b | 139.42 ± 1.61 b | 164.04 ± 8.16 c |
| Purification fold | 20.00 | 10.97 | 13.81 | 15.44 | 11.17 |
| Fresh | PCR-C01 | PCR-C03 | PCR-C05 | PCR-C10 | |
|---|---|---|---|---|---|
| Hesperidin | 113.64 ± 0.83 a (59.5) | 77.03 ± 1.00 b (51.1) | 87.13 ± 3.58 c (65.6) | 108.61 ± 0.34 a (67.6) | 99.58 ± 6.26 d (50.2) |
| Isosinensetin | 4.68 ± 0.04 a (2.5) | 3.30 ± 0.06 b (2.2) | 2.09 ± 0.02 c (1.6) | 1.93 ± 0.01 d (1.2) | 4.04 ± 0.02 e (2.0) |
| Sinensetin | 6.16 ± 0.10 a (3.2) | 4.39 ± 0.05 b (2.9) | 3.22 ± 0.03 c (2.4) | 3.24 ± 0.01 c (2.0) | 6.32 ± 0.02 d (3.2) |
| 5,7,8,4′-Tetramethoxyflavone | 3.08 ± 0.01 a (1.6) | 2.46 ± 0.03 b (1.6) | 1.89 ± 0.01 c (1.4) | 1.85 ± 0.01 d (1.2) | 3.04 ± 0.01 a (1.5) |
| Nobiletin | 39.97 ± 0.17 a (20.9) | 32.59 ± 0.33 b (21.6) | 23.76 ± 0.13 c (17.9) | 24.44 ± 0.04 d (15.2) | 48.92 ± 0.32 e (24.7) |
| 5,6,7,4′-Tetramethoxyflavone | 0.83 ± 0.05 a (0.4) | 0.82 ± 0.01 a (0.5) | 0.43 ± 0.02 b (0.3) | 0.60 ± 0.01 c (0.4) | 1.30 ± 0.02 d (0.7) |
| 3,5,6,7,8,3′,4′-Heptamethoxyflavone | 3.49 ± 0.03 a (1.8) | 2.71 ± 0.04 b (1.8) | 1.64 ± 0.02 c (1.2) | 1.97 ± 0.01 d (1.2) | 3.57 ± 0.02 e (1.8) |
| Tangeretin | 18.63 ± 0.09 a (9.8) | 26.80 ± 0.28 b (17.8) | 12.06 ± 0.05 c (9.1) | 17.46 ± 0.06 d (10.9) | 30.68 ± 0.19 e (15.5) |
| 5-Demethylnobiletin | 0.42 ± 0.01 a (0.2) | 0.73 ± 0.01 b (0.5) | 0.60 ± 0.04 c (0.5) | 0.51 ± 0.03 d (0.3) | 0.79 ± 0.00 e (0.4) |
| PMFs | 77.25 ± 0.36 a (40.5) | 73.80 ± 0.79 b (48.9) | 45.70 ± 0.25 c (34.4) | 51.99 ± 0.12 d (32.4) | 98.66 ± 0.56 e (49.8) |
| Total | 190.89 ± 1.10 a | 150.82 ± 1.78 b | 132.83 ± 3.83 c | 160.60 ± 0.44 d | 198.24 ± 6.04 e |
| Parameter | with ROS Level | with MDA Value | with SOD Activity | with GSH Content |
|---|---|---|---|---|
| Hesperidin | −0.158 | −0.626 | 0.525 | 0.147 |
| Isosinensetin | −0.685 | −0.380 | 0.655 | 0.418 |
| Sinensetin | −0.844 | −0.590 | 0.831 | 0.640 |
| 5,7,8,4′-Tetramethoxyflavone | −0.800 | −0.501 | 0.761 | 0.569 |
| Nobiletin | −0.953 * | −0.681 | 0.892 * | 0.805 |
| 5,6,7,4′-Tetramethoxyflavone | −0.970 ** | −0.751 | 0.871 | 0.879 * |
| 3,5,6,7,8,3′,4′-Heptamethoxyflavone | −0.825 | −0.593 | 0.796 | 0.607 |
| Tangeretin | −0.808 | −0.535 | 0.597 | 0.718 |
| 5-Demethylnobiletin | −0.556 | −0.159 | 0.231 | 0.619 |
| PMFs | −0.939 ** | −0.646 | 0.825 | 0.787 |
| Total flavonoids | −0.813 | −0.844 | 0.928 * | 0.690 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Q.; Tao, Y.; Huang, Y.; Zogona, D.; Wu, T.; Liu, R.; Pan, S.; Xu, X. Aged Pericarpium Citri Reticulatae ‘Chachi’ Attenuates Oxidative Damage Induced by tert-Butyl Hydroperoxide (t-BHP) in HepG2 Cells. Foods 2022, 11, 273. https://doi.org/10.3390/foods11030273
Yu Q, Tao Y, Huang Y, Zogona D, Wu T, Liu R, Pan S, Xu X. Aged Pericarpium Citri Reticulatae ‘Chachi’ Attenuates Oxidative Damage Induced by tert-Butyl Hydroperoxide (t-BHP) in HepG2 Cells. Foods. 2022; 11(3):273. https://doi.org/10.3390/foods11030273
Chicago/Turabian StyleYu, Qian, Yexing Tao, Yuting Huang, Daniel Zogona, Ting Wu, Ruiting Liu, Siyi Pan, and Xiaoyun Xu. 2022. "Aged Pericarpium Citri Reticulatae ‘Chachi’ Attenuates Oxidative Damage Induced by tert-Butyl Hydroperoxide (t-BHP) in HepG2 Cells" Foods 11, no. 3: 273. https://doi.org/10.3390/foods11030273