
Understanding In Vivo Mastication Behaviour and In Vitro Starch and Protein Digestibility of Pulsed Electric Field-Treated Black Beans after Cooking

 ,
,  , and
, and
Abstract
:1. Introduction
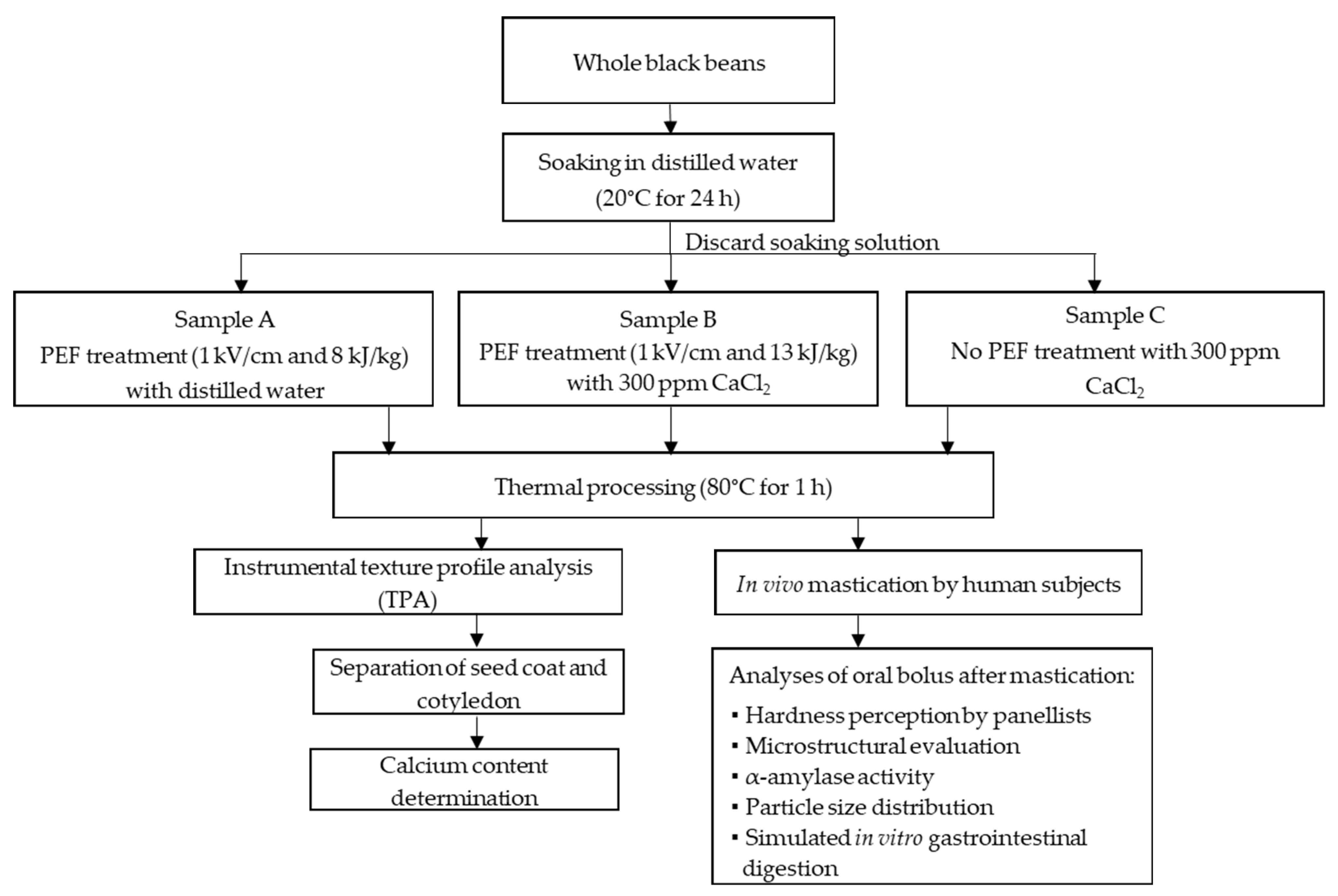
2. Materials and Methods
2.1. Raw Material
2.2. Selection of PEF Treatment Parameters and Other Processing Variables
2.3. Preparation of Black Bean Samples for In Vivo Mastication Study
2.4. In Vivo Mastication Study
2.5. Particle Size Distribution of Oral Boluses Using Image Analysis
2.5.1. Image Capturing and Processing of the Oral Bolus
2.5.2. Modelling the Particle Area Distribution of Oral Bolus
2.6. Determination of α-Amylase Activity in Oral Boluses
2.7. Microstructural Evaluation of Oral Boluses
2.8. Simulated In Vitro Human Gastric Intestinal Digestion Assay and Determination of Starch and Protein Digestibility
2.8.1. Starch and Protein Digesta Collection and Measurement
2.8.2. Kinetic Modelling of In Vitro Starch and Protein Digestibility at the Small Intestinal Phase
2.9. Statistical Data Analysis
3. Results and Discussion
3.1. Selection of PEF Treatment Parameters and Other Processing Variables
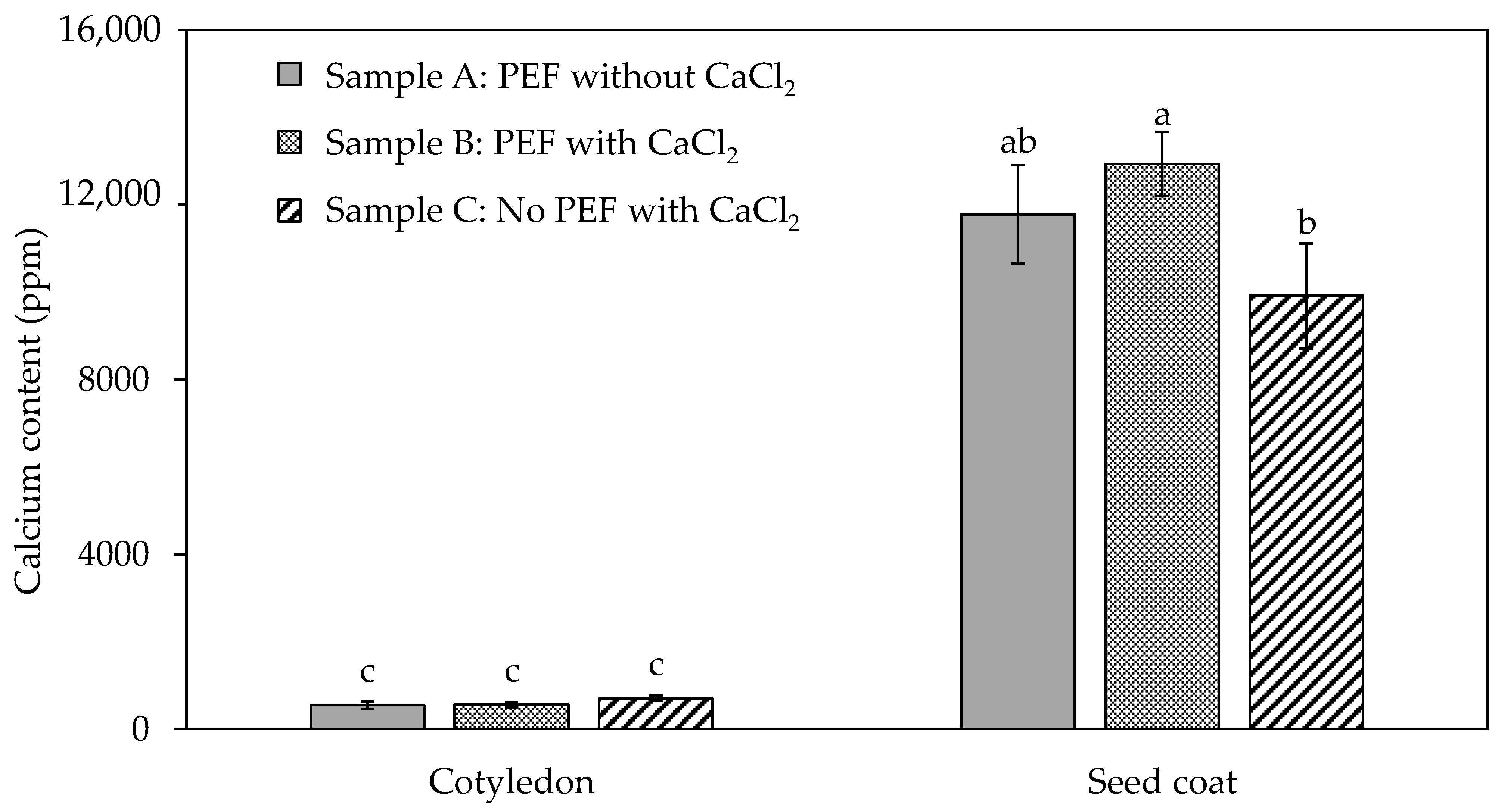
3.2. Texture Profile and Calcium Content of Differently Processed Black Beans Used for the In Vivo Mastication Study
3.3. Characterisation of Mastication Behaviour of Participants for PEF and Calcium Pre-Treated Cooked Black Beans
3.3.1. Texture Perception of Cooked Beans Rated by the Participants
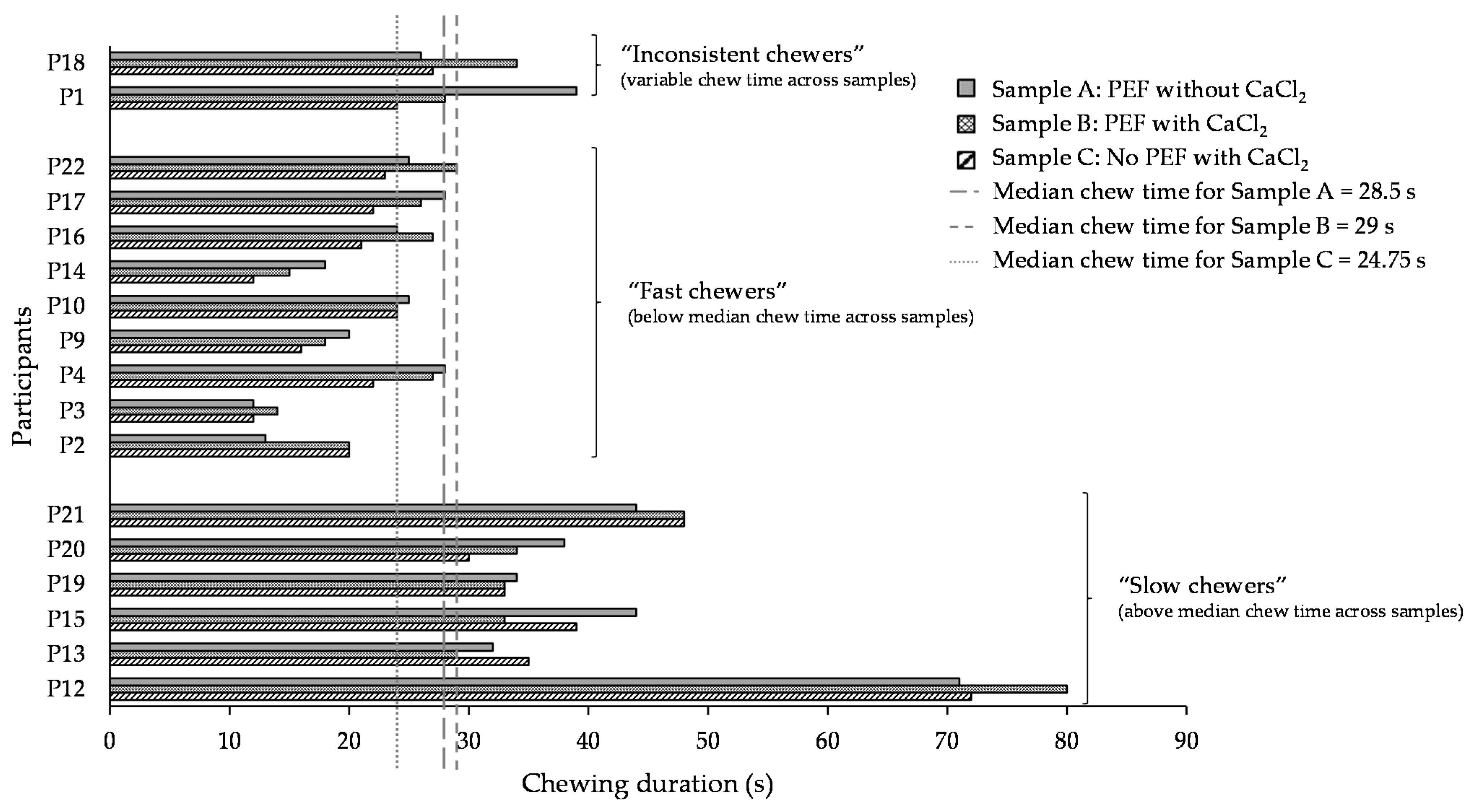
3.3.2. Chewing Duration of Cooked Beans before Ready for Swallowing
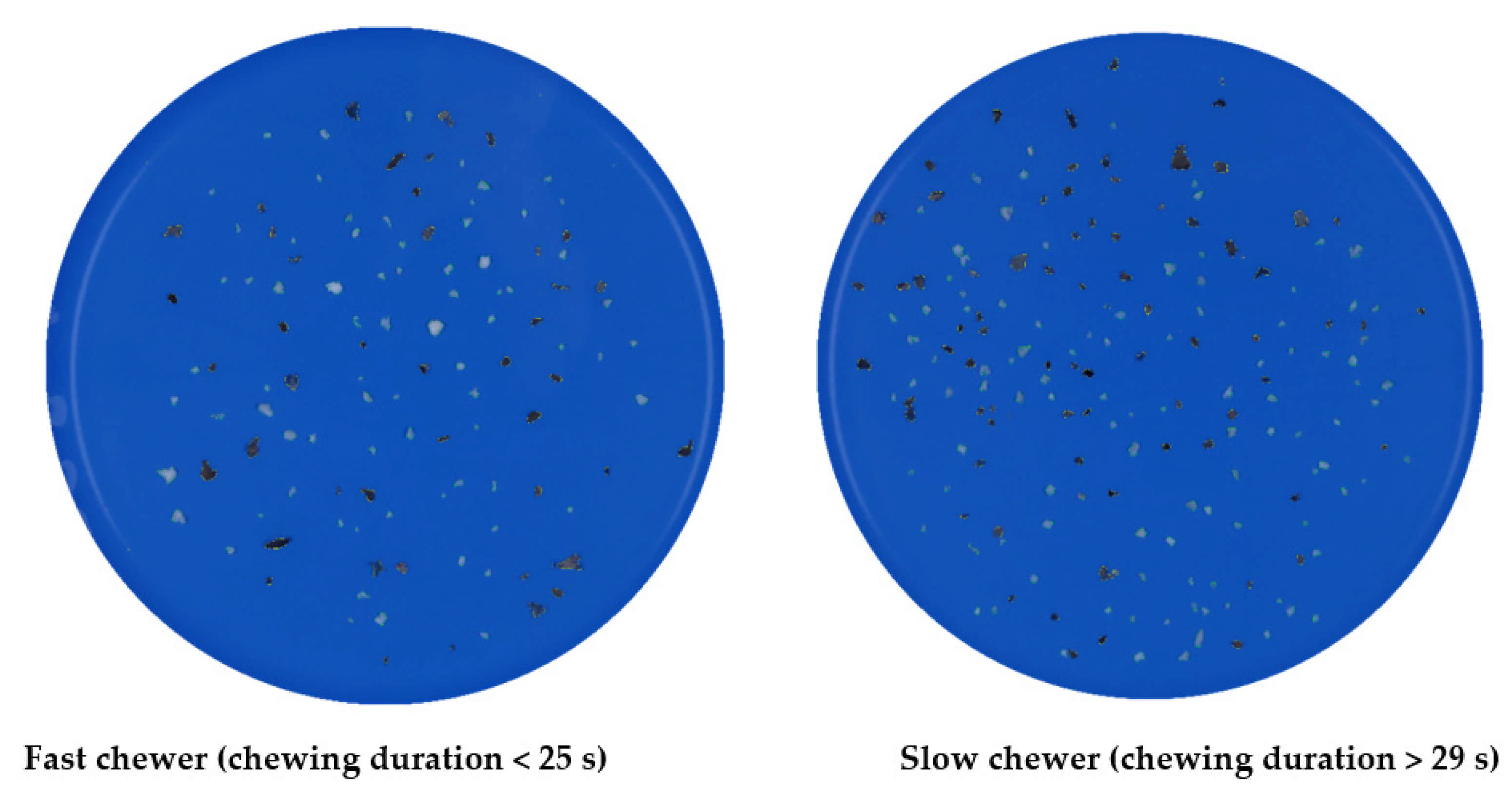
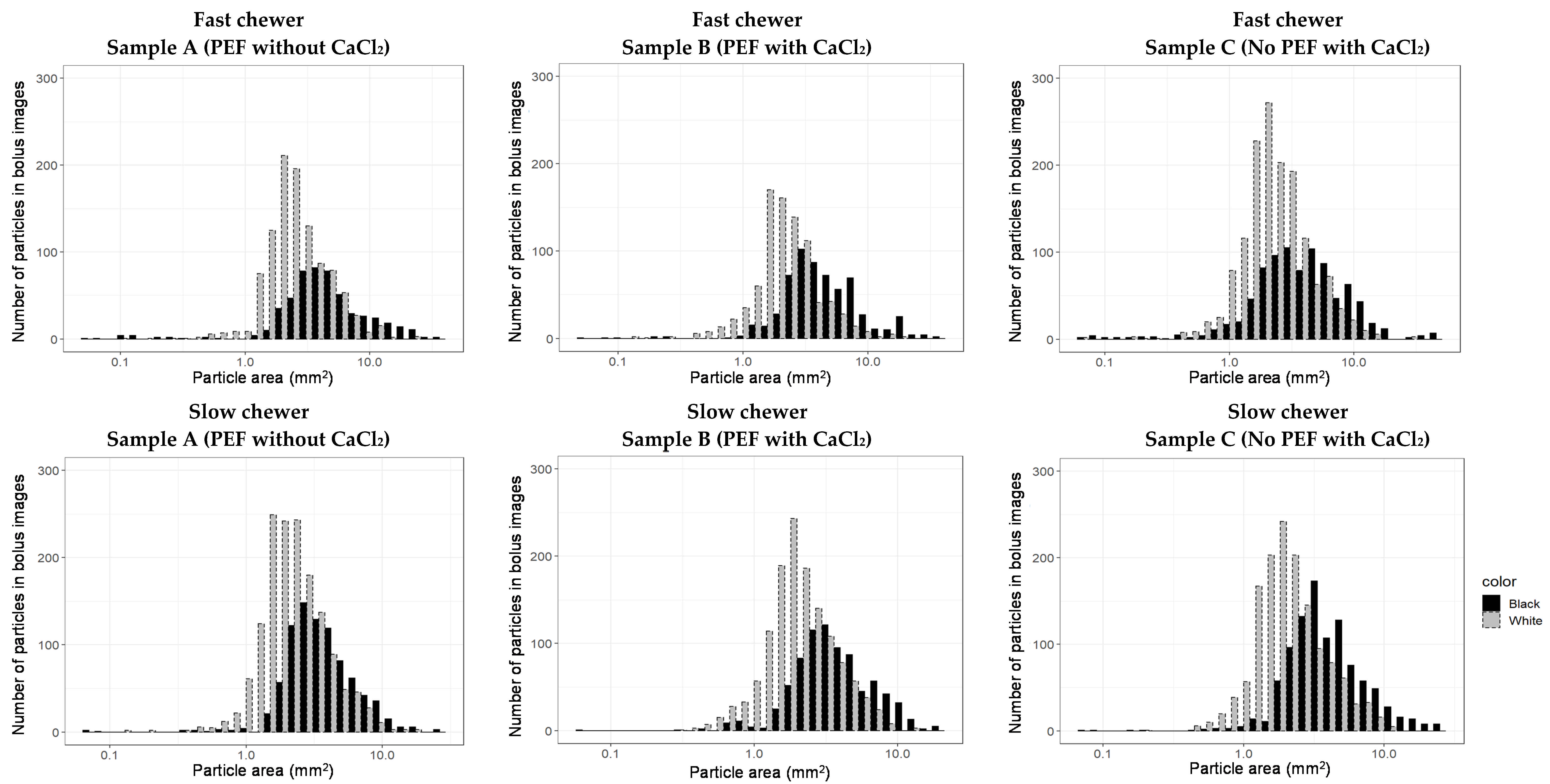
3.3.3. Particle Size Distribution after In Vivo Mastication of Differently Processed Black Beans
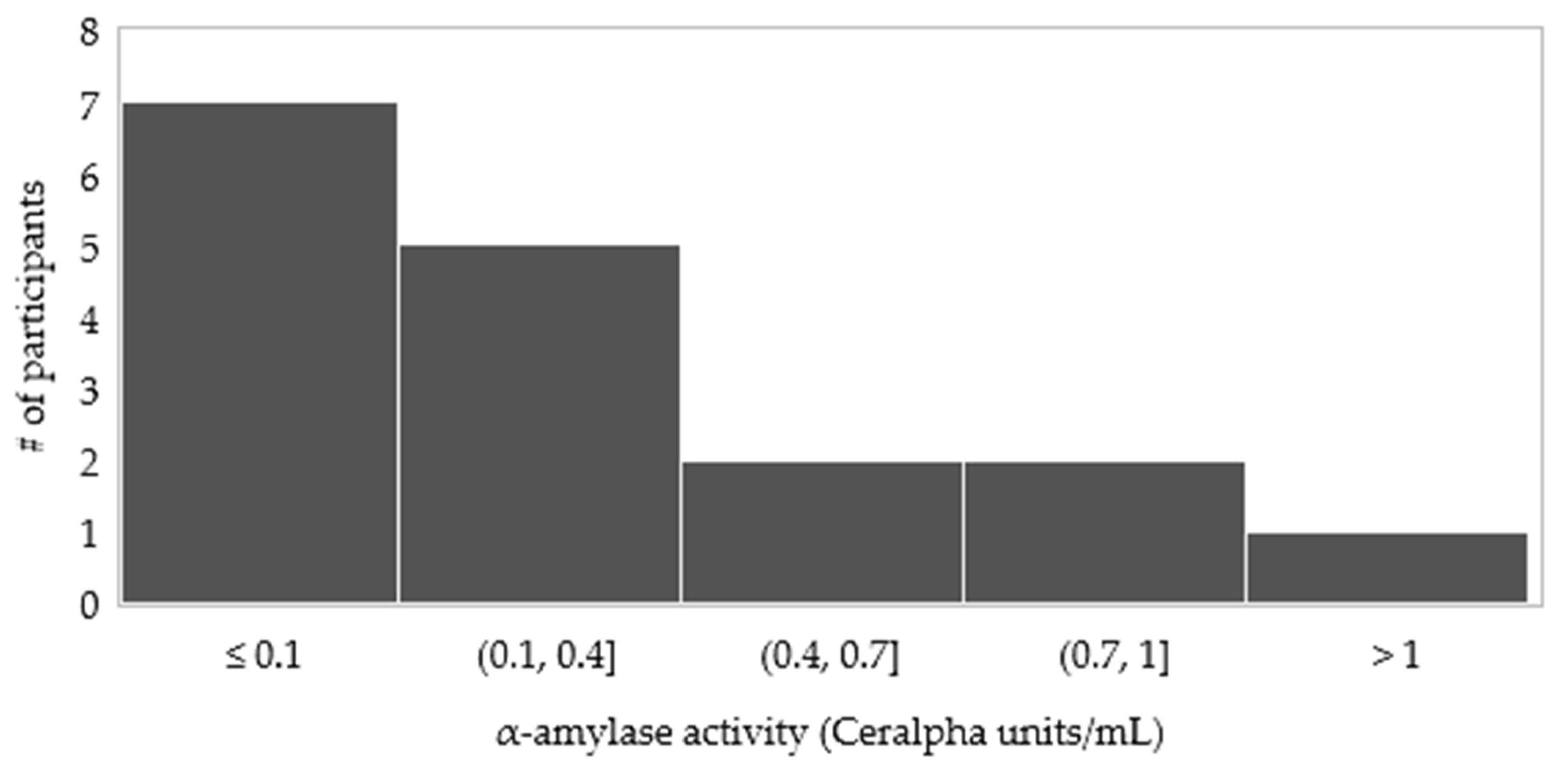
3.3.4. The Activity of α-Amylase in Oral Boluses
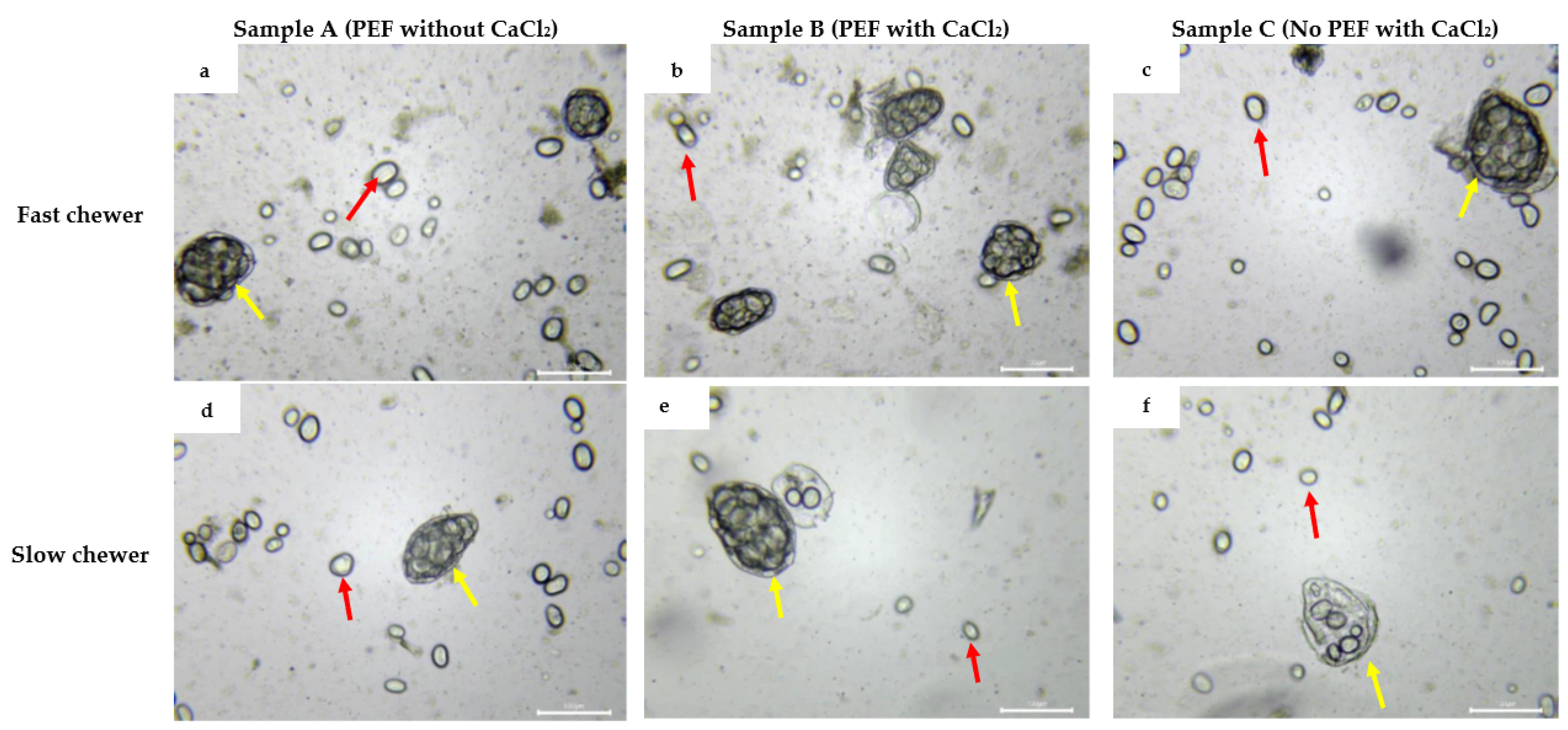
3.3.5. Microstructural Changes of Black Bean Oral Bolus
3.4. Characterisation of the Extent of In Vitro Starch Digestibility of In Vivo Masticated Black Beans by Different Participants
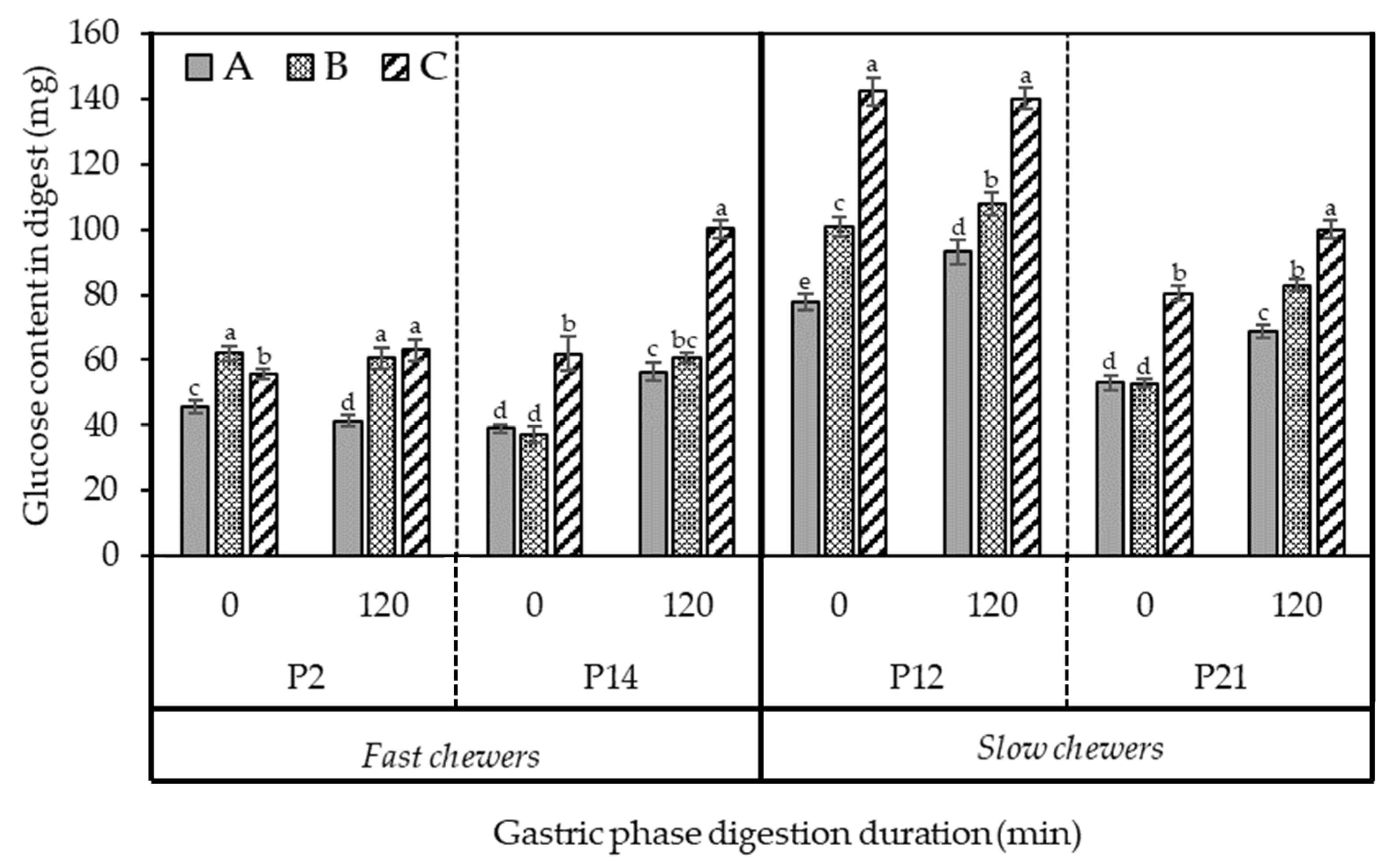
3.4.1. The Extent of Starch Digestibility during the In Vitro Gastric Phase
3.4.2. The Proportion of Different Starch Fractions (RDS, SDS and RS) Digested during the In Vitro Small Intestinal Phase
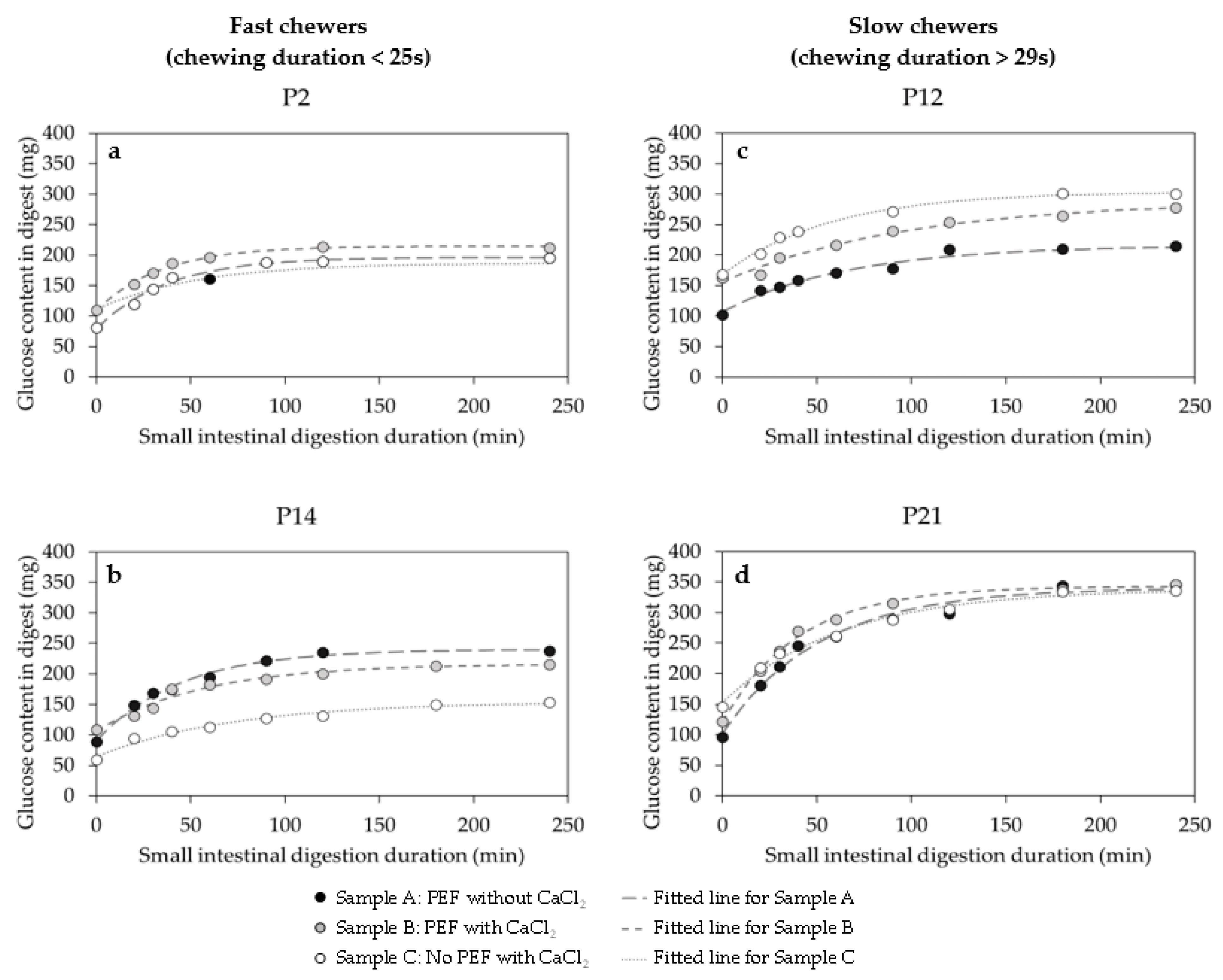
3.4.3. The Kinetics of Starch Digestibility during the In Vitro Small Intestinal Phase
3.5. Characterisation of the Extent of In Vitro Protein Digestibility of In Vivo Masticated Black Beans by Different Participants
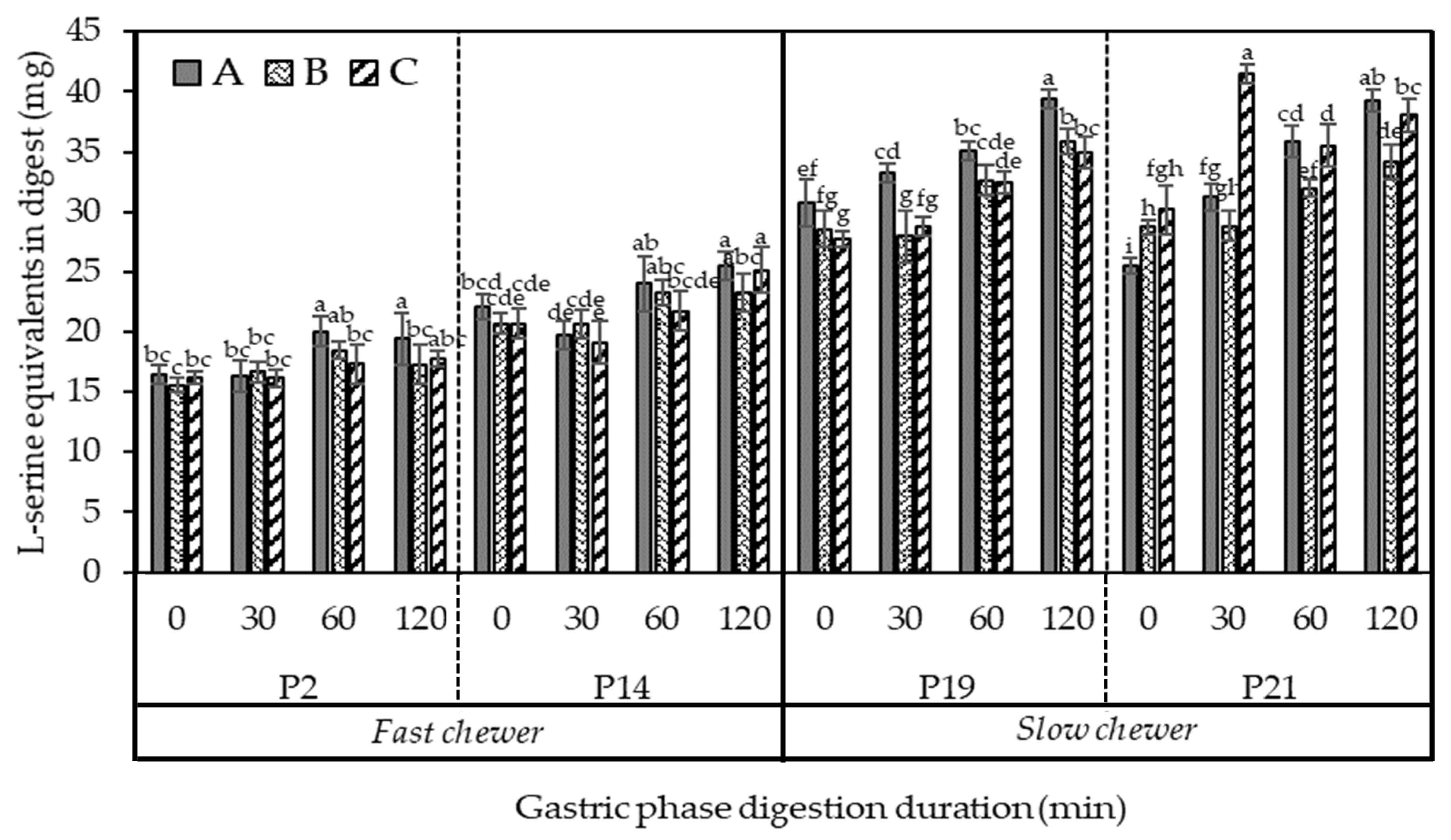
3.5.1. The Extent of Protein Digestibility during the In Vitro Gastric Phase
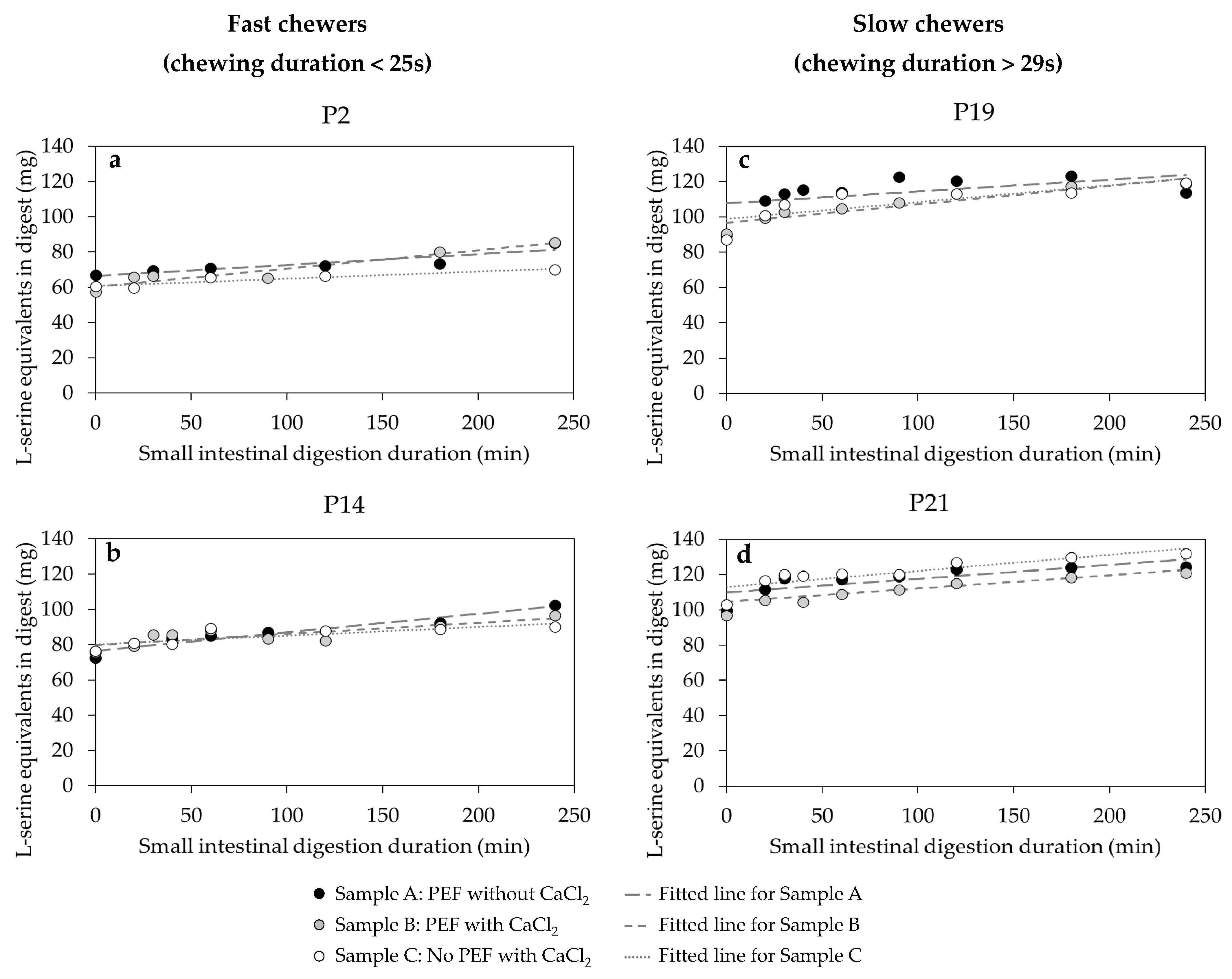
3.5.2. The Kinetic Behaviour of Protein Digestibility during the In Vitro Small Intestinal Phase
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Van Buggenhout, S.; Sila, D.; Duvetter, T.; Van Loey, A.; Hendrickx, M. Pectins in processed fruits and vegetables: Part III—Texture engineering. Compr. Rev. Food Sci. Food Saf. 2009, 8, 105–117. [Google Scholar] [CrossRef]
- Van Buren, J.P. The chemistry of texture in fruits and vegetables. J. Texture Stud. 1979, 10, 1–23. [Google Scholar] [CrossRef]
- Van der Sman, R. Impact of processing factors on quality of frozen vegetables and fruits. Food Eng. Rev. 2020, 12, 399–420. [Google Scholar] [CrossRef]
- Moens, L.G.; Huang, W.; Van Loey, A.M.; Hendrickx, M.E.G. Effect of pulsed electric field and mild thermal processing on texture-related pectin properties to better understand carrot (Daucus carota) texture changes during subsequent cooking. Innov. Food Sci. Emerg. Technol. 2021, 70, 102700. [Google Scholar] [CrossRef]
- Leong, S.Y.; Du, D.; Oey, I. Pulsed Electric Fields enhances calcium infusion for improving the hardness of blanched carrots. Innov. Food Sci. Emerg. Technol. 2018, 47, 46–55. [Google Scholar] [CrossRef]
- Lebovka, N.I.; Praporscic, I.; Vorobiev, E. Effect of moderate thermal and pulsed electric field treatments on textural properties of carrots, potatoes and apples. Innov. Food Sci. Emerg. Technol. 2004, 5, 9–16. [Google Scholar] [CrossRef]
- Toepfl, S.; Heinz, V.; Knorr, D. Applications of pulsed electric fields technology for the food industry. In Pulsed Electric Fields Technology for the Food Industry: Fundamentals and Applications; Raso, J., Heinz, V., Eds.; Springer: New York, NY, USA, 2006; pp. 197–221. [Google Scholar]
- Bornhorst, G.M.; Singh, R.P. Bolus formation and disintegration during digestion of food carbohydrates. Compr. Rev. Food Sci. Food Saf. 2012, 11, 101–118. [Google Scholar] [CrossRef]
- Chen, J.; Khandelwal, N.; Liu, Z.; Funami, T. Influences of food hardness on the particle size distribution of food boluses. Arch. Oral Biol. 2013, 58, 293–298. [Google Scholar] [CrossRef]
- Tharanathan, R.N.; Mahadevamma, S. Grain legumes—A boon to human nutrition. Trends Food Sci. Technol. 2003, 14, 507–518. [Google Scholar] [CrossRef]
- Pallares, A.P.; Loosveldt, B.; Karimi, S.N.; Hendrickx, M.; Grauwet, T. Effect of process-induced common bean hardness on structural properties of in vivo generated boluses and consequences for in vitro starch digestion kinetics. Br. J. Nutr. 2019, 122, 388–399. [Google Scholar] [CrossRef]
- Pangborn, R.M.; Lundgren, B. Salivary secretion in response to mastication of crisp bread. J. Texture Stud. 1977, 8, 463–472. [Google Scholar] [CrossRef]
- Bornhorst, G.M.; Kostlan, K.; Singh, R.P. Particle size distribution of brown and white rice during gastric digestion measured by image analysis. J. Food Sci. 2013, 78, E1383–E1391. [Google Scholar] [CrossRef] [PubMed]
- Etzler, F.M.; Sanderson, M.S. Particle size analysis: A comparative study of various methods. Part. Part. Syst. Charact. 1995, 12, 217–224. [Google Scholar] [CrossRef]
- Mora, C.; Kwan, A.; Chan, H. Particle size distribution analysis of coarse aggregate using digital image processing. Cem. Concr. Res. 1998, 28, 921–932. [Google Scholar] [CrossRef]
- Alpos, M.; Leong, S.Y.; Oey, I. Combined effects of calcium addition and thermal processing on the texture and in vitro digestibility of starch and protein of black beans (Phaseolus vulgaris). Foods 2021, 10, 1368. [Google Scholar] [CrossRef]
- Lombardi-Boccia, G.; Lucarini, M.; Di Lullo, G.; Del Puppo, E.; Ferrari, A.; Carnovale, E. Dialysable, soluble and fermentable calcium from beans (Phaseolus vulgaris L.) as model for in vitro assessment of the potential calcium availability. Food Chem. 1998, 61, 167–171. [Google Scholar] [CrossRef]
- Wesley, R.; Rousselle, J.; Schwan, D.; Stadelman, W. Improvement in quality of scrambled egg products served from steam table display. Poult. Sci. 1982, 61, 457–462. [Google Scholar] [CrossRef]
- Swackhamer, C.; Zhang, Z.; Taha, A.Y.; Bornhorst, G.M. Fatty acid bioaccessibility and structural breakdown from in vitro digestion of almond particles. Food Funct. 2019, 10, 5174–5187. [Google Scholar] [CrossRef] [Green Version]
- McCleary, B.V.; McNally, M.; Monaghan, D.; Mugford, D.C. Measurement of α-amylase activity in white wheat flour, milled malt, and microbial enzyme preparations, using the Ceralpha assay: Collaborative study. J. AOAC Int. 2002, 85, 1096–1102. [Google Scholar] [CrossRef] [Green Version]
- Gwala, S.; Wainana, I.; Pallares Pallares, A.; Kyomugasho, C.; Hendrickx, M.; Grauwet, T. Texture and interlinked post-process microstructures determine the in vitro starch digestibility of Bambara groundnuts with distinct hard-to-cook levels. Food Res. Int. 2019, 120, 1–11. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrire, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Abduh, S.; Leong, S.Y.; Agyei, D.; Oey, I. Understanding the properties of starch in potatoes (Solanum tuberosum var. Agria) after being treated with pulsed electric field processing. Foods 2019, 8, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallares, A.P.; Miranda, B.A.; Truong, N.Q.A.; Kyomugasho, C.; Chigwedere, C.M.; Hendrickx, M.; Grauwet, T. Process-induced cell wall permeability modulates the in vitro starch digestion kinetics of common bean cotyledon cells. Food Funct. 2018, 9, 6544–6554. [Google Scholar] [CrossRef]
- Piecyk, M.; Wołosiak, R.; Drużynska, B.; Worobiej, E. Chemical composition and starch digestibility in flours from Polish processed legume seeds. Food Chem. 2012, 135, 1057–1064. [Google Scholar] [CrossRef]
- Wani, I.A.; Sogi, D.S.; Wani, A.A.; Gill, B.S. Physical and cooking characteristics of some Indian kidney bean (Phaseolus vulgaris L.) cultivars. J. Saudi Soc. Agric. Sci. 2017, 16, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Guiné, R.P.F.; Roque, A.R.F.; Seiça, F.F.A.; Batista, C.E.O. Effect of chemical pretreatments on the physical properties of kiwi. Int. J. Food Eng. 2016, 2, 90–95. [Google Scholar] [CrossRef]
- Liu, X.; Lu, K.; Yu, J.; Copeland, L.; Wang, S.; Wang, S. Effect of purple yam flour substitution for wheat flour on in vitro starch digestibility of wheat bread. Food Chem. 2019, 284, 118–124. [Google Scholar] [CrossRef]
- Englyst, H.N.; Veenstra, J.; Hudson, G.J. Measurement of rapidly available glucose (RAG) in plant foods: A potential in vitro predictor of the glycaemic response. Br. J. Nutr. 1996, 75, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-F.; Oey, I.; Bremer, P.; Silcock, P.; Carne, A. In vitro peptic digestion of ovomucin-depleted egg white affected by pH, temperature and pulsed electric fields. Food Chem. 2017, 231, 165–174. [Google Scholar] [CrossRef]
- Gwala, S.; Pallares, A.P.; Pälchen, K.; Hendrickx, M.; Grauwet, T. In vitro starch and protein digestion kinetics of cooked Bambara groundnuts depend on processing intensity and hardness sorting. Food Res. Int. 2020, 137, 109512. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-Cánovas, G.V.; Pothakamury, U.R.; Gongora-Nieto, M.M.; Swanson, B.G. Preservation of Foods with Pulsed Electric Fields; Academic Press: Cambridge, UK, 1999. [Google Scholar]
- Parniakov, O.; Bals, O.; Lebovka, N.; Vorobiev, E. Effects of pulsed electric fields assisted osmotic dehydration on freezing-thawing and texture of apple tissue. J. Food Eng. 2016, 183, 32–38. [Google Scholar] [CrossRef]
- Yi, J.; Njoroge, D.M.; Sila, D.N.; Kinyanjui, P.K.; Christiaens, S.; Bi, J.; Hendrickx, M.E. Detailed analysis of seed coat and cotyledon reveals molecular understanding of the hard-to-cook defect of common beans (Phaseolus vulgaris L.). Food Chem. 2016, 210, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.N.; Galindo, F.G.; Vicente, A.A.; Dejmek, P. Effects of pulsed electric field on the viscoelastic properties of potato tissue. Food Biophys. 2009, 4, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, P.; Yang, R.; Wang, X.; Gu, Z. Ca2+ influxes and transmembrane transport are essential for phytic acid degradation in mung bean sprouts: Mechanism of the enhanced phytic acid degradation in mung bean sprouts under CaCl2 treatment. J. Sci. Food Agric. 2017, 98, 1968–1976. [Google Scholar] [CrossRef]
- Mishellany, A.; Woda, A.; Labas, R.; Peyron, M.-A. The challenge of mastication: Preparing a bolus suitable for deglutition. Dysphagia 2006, 21, 87–94. [Google Scholar] [CrossRef]
- Olthoff, L.; Van der Bilt, A.; Bosman, F.; Kleizen, H. Distribution of particle sizes in food comminuted by human mastication. Arch. Oral Biol. 1984, 29, 899–903. [Google Scholar] [CrossRef]
- Gavião, M.B.D.; Engelen, L.; Van Der Bilt, A. Chewing behavior and salivary secretion. Eur. J. Oral Sci. 2004, 112, 19–24. [Google Scholar] [CrossRef]
- Engelen, L.; Fontijn-Tekamp, A.; van der Bilt, A. The influence of product and oral characteristics on swallowing. Arch. Oral Biol. 2005, 50, 739–746. [Google Scholar] [CrossRef]
- Pedersen, A.; Bardow, A.; Jensen, S.B.; Nauntofte, B. Saliva and gastrointestinal functions of taste, mastication, swallowing and digestion. Oral Dis. 2002, 8, 117–129. [Google Scholar] [CrossRef]
- Fried, M.; Abramson, S.; Meyer, J. Passage of salivary amylase through the stomach in humans. Dig. Dis. Sci. 1987, 32, 1097–1103. [Google Scholar] [CrossRef]
- Rosenblum, J.L.; Irwin, C.L.; Alpers, D.H. Starch and glucose oligosaccharides protect salivary-type amylase activity at acid pH. Am. J. Physiol. Gastrointest. Liver Physiol. 1988, 254, G775–G780. [Google Scholar] [CrossRef]
- Leong, S.Y.; Duque, S.M.; Abduh, S.B.M.; Oey, I. Carbohydrates. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Barba, F.J., Saraiva, J.M.A., Cravotto, G., Lorenzo, J.M., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 171–206. [Google Scholar]
- Osorio-Díaz, P.; Bello-Pérez, L.A.; Sáyago-Ayerdi, S.G.; Benítez-Reyes, M.D.P.; Tovar, J.; Paredes-López, O. Effect of processing and storage time on in vitro digestibility and resistant starch content of two bean (Phaseolus vulgaris L.) varieties. J. Sci. Food Agric. 2003, 83, 1283–1288. [Google Scholar] [CrossRef]
- Jenkins, A.L.; Marchie, A.; Kendall, C.W.C.; Augustin, L.S.A.; Hamidi, M.; Axelsen, M.; Franceschi, S.; Jenkins, D.J.A. Glycemic index: Overview of implications in health and disease. Am. J. Clin. Nutr. 2002, 76, 266S–273S. [Google Scholar] [CrossRef]
- Rovalino-Córdova, A.M.; Fogliano, V.; Capuano, E. The effect of cell wall encapsulation on macronutrients digestion: A case study in kidney beans. Food Chem. 2019, 286, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wu, Q.-Y.; Jiang, W.; Qian, J.-Y.; Zhang, L.; Wu, M.; Rao, S.-Q.; Wu, C.-S. Effect of pulsed electric field on structural properties and digestibility of starches with different crystalline type in solid state. Carbohydr. Polym. 2019, 207, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Silk, D.; Grimble, G.; Rees, R.G. Protein digestion and amino acid and peptide absorption. Proc. Nutr. Soc. 1985, 44, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahir, M.; Fogliano, V.; Capuano, E. Food matrix and processing modulate in vitro protein digestibility in soybeans. Food Funct. 2018, 9, 6326–6336. [Google Scholar] [CrossRef] [PubMed]
- Tinus, T.; Damour, M.; van Riel, V.; Sopade, P.A. Particle size-starch–protein digestibility relationships in cowpea (Vigna unguiculata). J. Food Eng. 2012, 113, 254–264. [Google Scholar] [CrossRef]
- Paz-Yépez, C.; Peinado, I.; Heredia, A.; Andrés, A. Influence of particle size and intestinal conditions on in vitro lipid and protein digestibility of walnuts and peanuts. Food Res. Int. 2019, 119, 951–959. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Morton, J.D.; Mason, S.L.; Jayawardena, S.R.; Bekhit, A.E.-D.A. Pulsed electric field: A new way to improve digestibility of cooked beef. Meat Sci. 2019, 155, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; Becker, K. Dietary roles of phytate and phytase in human nutrition: A review. Food Chem. 2010, 120, 945–959. [Google Scholar] [CrossRef]
- Deshpande, S.; Damodaran, S. Effect of phytate on solubility, activity and conformation of trypsin and chymotrypsin. J. Food Sci. 1989, 54, 695–699. [Google Scholar] [CrossRef]
- Knuckles, B.; Kuzmicky, D.; Betschart, A.A. Effect of phytate and partially hydrolyzed phytate on in vitro protein digestibility. J. Food Sci. 1985, 50, 1080–1082. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Oral Bolus † | |
|---|---|---|
| x50 (mm2) | b | |
| A | 5.0 ± 1.3 a | 1.63 ± 0.19 a |
| B | 5.2 ± 1.2 a | 1.60 ± 0.18 a |
| C | 5.4 ± 0.9 a | 1.53 ± 0.14 a |
| Sample | Cotyledon (“White”) | Seed Coat (“Black”) | ||
|---|---|---|---|---|
| x50 (mm2) | b | x50 (mm2) | b | |
| A | 3.5 ± 0.9 a | 1.9 ± 0.3 a | 6.7 ± 1.8 a | 1.7 ± 0.2 a |
| B | 3.5 ± 0.8 a | 1.9 ± 0.3 a | 7.1 ± 2.0 a | 1.7 ± 0.3 a |
| C | 3.7 ± 0.6 a | 1.8 ± 0.3 a | 7.6 ± 1.4 a | 1.58 ± 0.16 a |
| Participants | RDS (%) | SDS (%) | RS (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample A | Sample B | Sample C | Sample A | Sample B | Sample C | Sample A | Sample B | Sample C | |
| Fast chewers (chewed < 25 s) | |||||||||
| P2 | 32.8 ± 0.7 dC | 42.91 ± 1.70 dA | 38.6 ± 0.9 dB | 19.5 ± 2.4 cA | 17.6 ± 1.8 bA | 18.6 ± 7.0 cdA | 47.8 ± 1.9 bA | 39.52 ± 1.80 cB | 42.9 ± 6.8 cAB |
| P14 | 37.7 ± 1.9 cA | 37.0 ± 2.0 eA | 27.7 ± 1.1 fB | 21.8 ± 0.9 cA | 19.3 ± 0.9 bB | 10.9 ± 1.7 eC | 40.5 ± 1.4 cC | 43.71 ± 2.50 bB | 61.32 ± 1.10 aA |
| P16 | 20.81 ± 0.70 eC | 29.6 ± 0.6 fB | 34.0 ± 1.3 eA | 13.6 ± 1.3 dB | 18.6 ± 1.8 bA | 13.2 ± 1.7 deB | 65.6 ± 1.1 aA | 51.8 ± 1.7 aB | 52.8 ± 1.2 bB |
| Slow chewers (chewed > 29 s) | |||||||||
| P12 | 39.0 ± 1.5 cC | 47.8 ± 1.3 cB | 57.4 ± 2.3 bA | 26.2 ± 3.5 bA | 24.5 ± 0.9 aA | 19.6 ± 1.9 bcB | 33.4 ± 3.3 dA | 27.7 ± 0.9 dB | 23.0 ± 2.5 dC |
| P19 | 56.4 ± 1.0 aB | 69.1 ± 1.1 aA | 51.6 ± 0.9 cC | 26.4 ± 1.4 bA | 19.1 ± 2.2 bB | 24.3 ± 2.3 abA | 17.2 ± 1.0 fB | 13.6 ± 1.8 fC | 24.2 ± 1.7 dA |
| P21 | 46.2 ± 1.4 bC | 54.9 ± 2.2 bB | 61.8 ± 1.5 aA | 29.9 ± 3.1 aA | 27.4 ± 3.2 aA | 28.3 ± 3.9 aA | 24.0 ± 1.9 eA | 17.7 ± 2.1 eB | 11.7 ± 1.5 eC |
| Participants | S0 (mg) | Sf (mg) | ks (×10−2 min−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample A | Sample B | Sample C | Sample A | Sample B | Sample C | Sample A | Sample B | Sample C | |
| Fast chewers (chewed < 25 s) | |||||||||
| P2 | 78.4 ± 5.6 | 108.5 ± 3.3 | 110.6 ± 7.2 | 196.4 ± 4.4 | 214.6 ± 2.7 | 186.9 ± 6.7 | 2.7 ± 0.4 | 3.0 ± 0.2 | 1.9 ± 0.6 |
| P14 | 90.5 ± 5.2 | 105.9 ± 6.6 | 64.4 ± 4.9 | 240.0 ± 4.7 | 216.0 ± 6.1 | 154.6 ± 6.1 | 2.3 ± 0.2 | 1.8 ± 0.3 | 1.4 ± 0.3 |
| P16 | 50.6 ± 4.4 | 112.5 ± 4.2 | 103.4 ± 5.0 | 133.8 ± 4.5 | 209.5 ± 4.8 | 200.9 ± 4.6 | 1.6 ± 0.3 | 1.4 ± 0.2 | 1.8 ± 0.3 |
| Average | 73.18 ± 5.06 a | 109.0 ± 4.7 a | 92.8 ± 5.7 a | 190.1 ± 4.6 a | 213.4 ± 4.5 a | 180.8 ± 5.8 a | 2.2 ± 0.3 a | 2.1 ± 0.3 a | 1.7 ± 0.4 a |
| Slow chewers (chewed > 29 s) | |||||||||
| P12 | 107.3 ± 5.4 | 156.2 ± 6.1 | 168.0 ± 4.7 | 216.0 ± 6.2 | 289.2 ± 12.3 | 303.8 ± 4.5 | 1.5 ± 0.2 | 1.0 ± 0.2 | 1.8 ± 0.2 |
| P19 | 152.3 ± 7.8 | 145.1 ± 11.9 | 128.6 ± 11.8 | 324.8 ± 5.7 | 306.40 ± 9.80 | 306.5 ± 9.3 | 2.3 ± 0.3 | 2.1 ± 0.4 | 2.1 ± 0.4 |
| P21 | 102.6 ± 11.6 | 121.6 ± 5.2 | 152.9 ± 7.1 | 340.7 ± 9.9 | 343.0 ± 3.9 | 338.8 ± 7.7 | 1.9 ± 0.3 | 2.5 ± 0.1 | 1.6 ± 0.2 |
| Average | 120.7 ± 8.3 a | 141.0 ± 7.7 a | 149.8 ± 6.3 a | 293.8 ± 7.3 a | 312.9 ± 6.5 a | 316.4 ± 7.2 a | 1.9 ± 0.3 a | 1.9 ± 0.3 a | 1.8 ± 0.3 a |
| Participants | Sample A | Sample B | Sample C |
|---|---|---|---|
| Fast chewers (chewed < 25 s) | |||
| P2 | 6.2 ± 1.5 | 10.4 ± 1.5 | 4.1 ± 0.9 |
| P4 | 5.5 ± 1.6 | 9.2 ± 1.0 | 4.3 ± 1.9 |
| P16 | 4.7 ± 1.5 | 8.4 ± 1.3 | 5.4 ± 1.8 |
| Average | 5.5 ± 1.5 b | 9.3 ± 1.3 a | 4.6 ± 1.5 b |
| Slow chewers (chewed > 29 s) | |||
| P12 | 10.2 ± 1.5 | 16.1 ± 1.2 | 11.5 ± 2.7 |
| P19 | 6.7 ± 4.0 | 10.5 ± 1.7 | 9.6 ± 3.3 |
| P21 | 7.7 ± 2.5 | 7.5 ± 2.6 | 9.0 ± 2.1 |
| Average | 8.2 ± 2.7 a | 11.3 ± 1.8 a | 10.0 ± 2.7 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alpos, M.; Leong, S.Y.; Liesaputra, V.; Martin, C.E.; Oey, I. Understanding In Vivo Mastication Behaviour and In Vitro Starch and Protein Digestibility of Pulsed Electric Field-Treated Black Beans after Cooking. Foods 2021, 10, 2540. https://doi.org/10.3390/foods10112540
Alpos M, Leong SY, Liesaputra V, Martin CE, Oey I. Understanding In Vivo Mastication Behaviour and In Vitro Starch and Protein Digestibility of Pulsed Electric Field-Treated Black Beans after Cooking. Foods. 2021; 10(11):2540. https://doi.org/10.3390/foods10112540
Chicago/Turabian StyleAlpos, Marbie, Sze Ying Leong, Veronica Liesaputra, Candace E. Martin, and Indrawati Oey. 2021. "Understanding In Vivo Mastication Behaviour and In Vitro Starch and Protein Digestibility of Pulsed Electric Field-Treated Black Beans after Cooking" Foods 10, no. 11: 2540. https://doi.org/10.3390/foods10112540