
Comparative Transcriptome Analysis of the Accumulation of Anthocyanins Revealed the Underlying Metabolic and Molecular Mechanisms of Purple Pod Coloration in Okra (Abelmoschus esculentus L.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Chemicals and Reagents
2.3. RNA Preparation and RNA-Seq
2.4. De Novo Transcriptome Assembly and Annotation
2.5. Identification of Differentially Expressed Genes (DEGs)
2.6. Extraction and UHPLC-Q-Orbitrap HRMS Analysis of Flavonoids
2.7. Statistical Analysis
3. Results
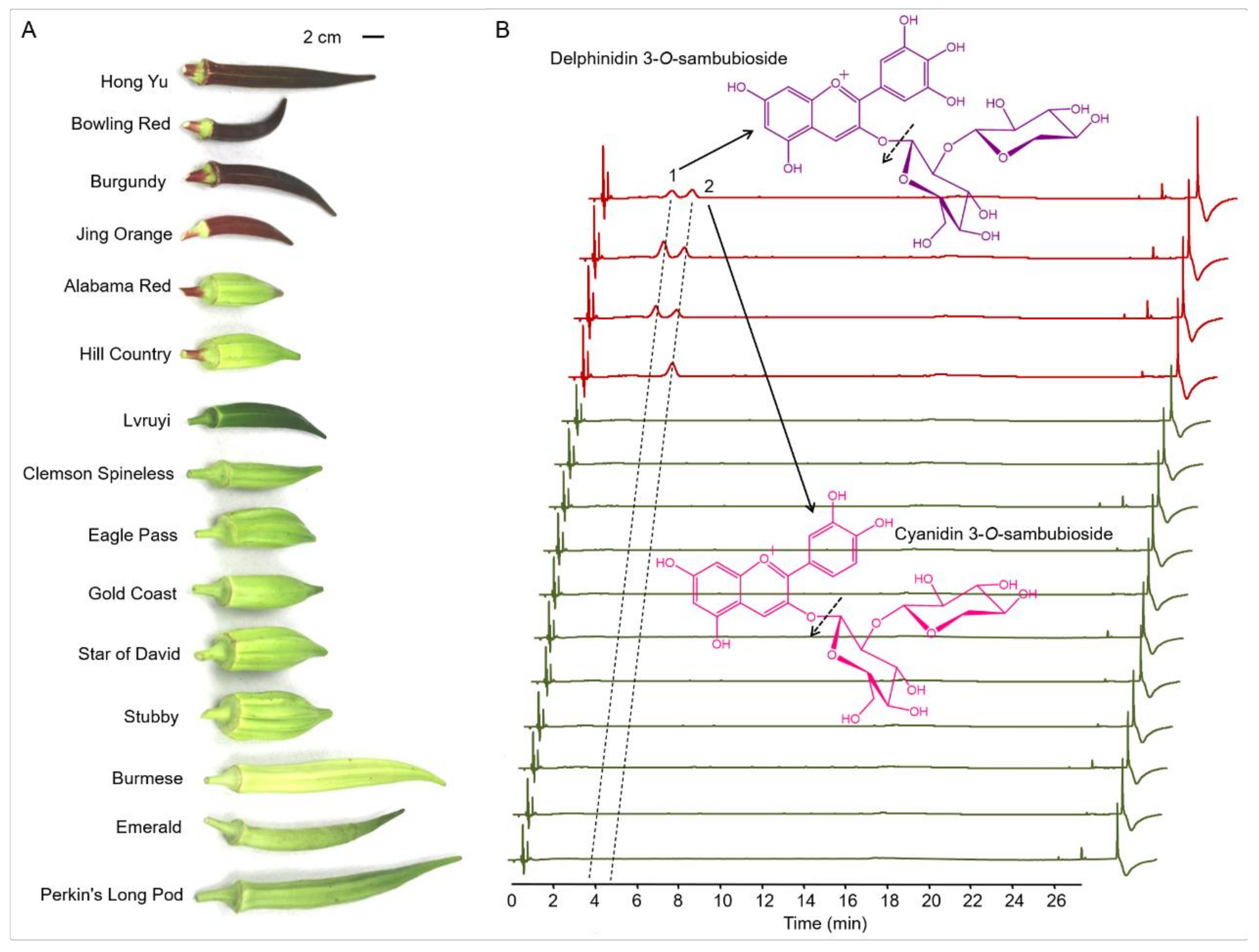
3.1. Identification and Quantitation of Anthocyanins in Pods of 15 Okra Cultivars
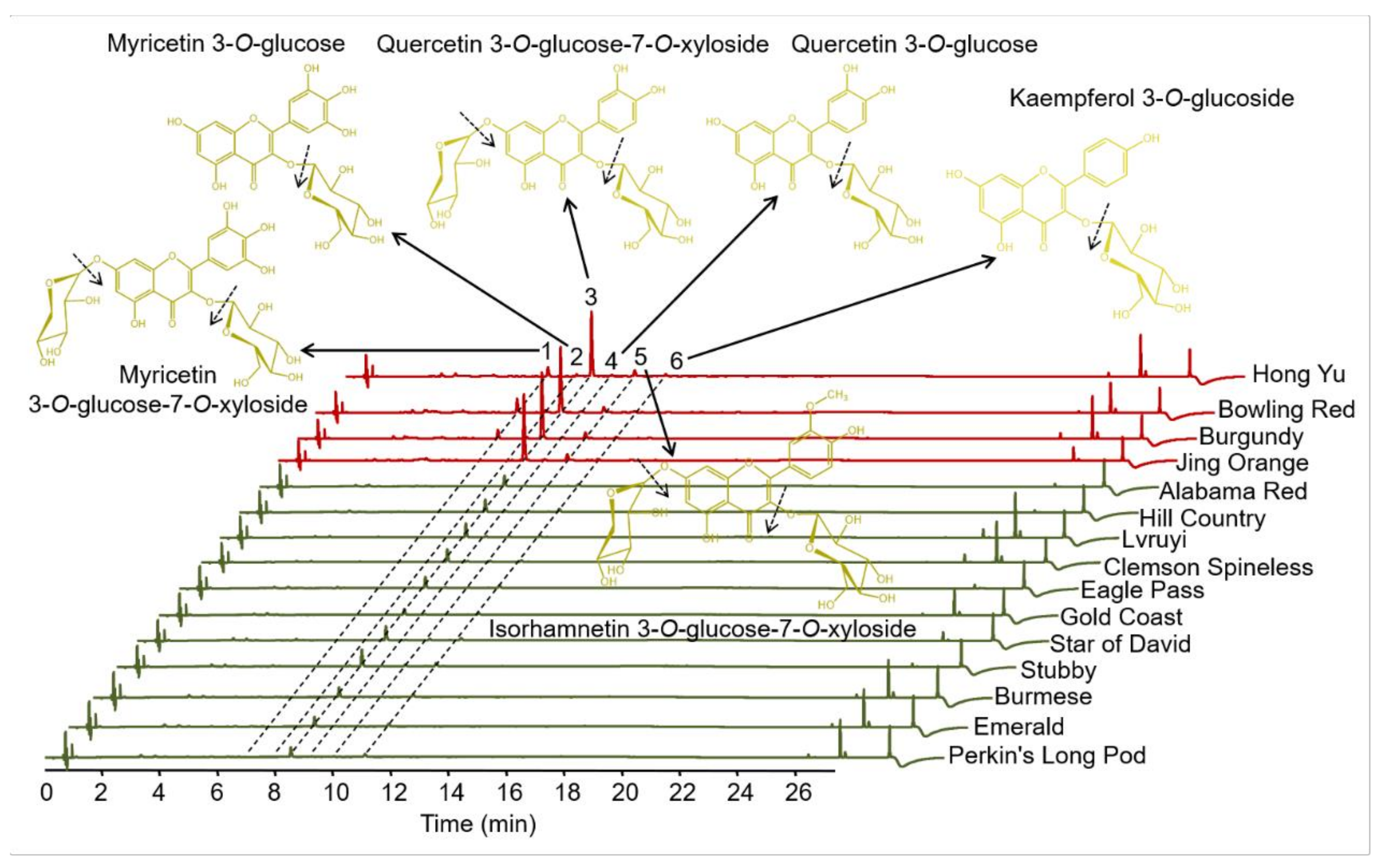
3.2. Identification and Quantitation of Flavonols in Various Okra Pods
3.3. RNA-Seq, De Novo Assembly, and Functional Annotation
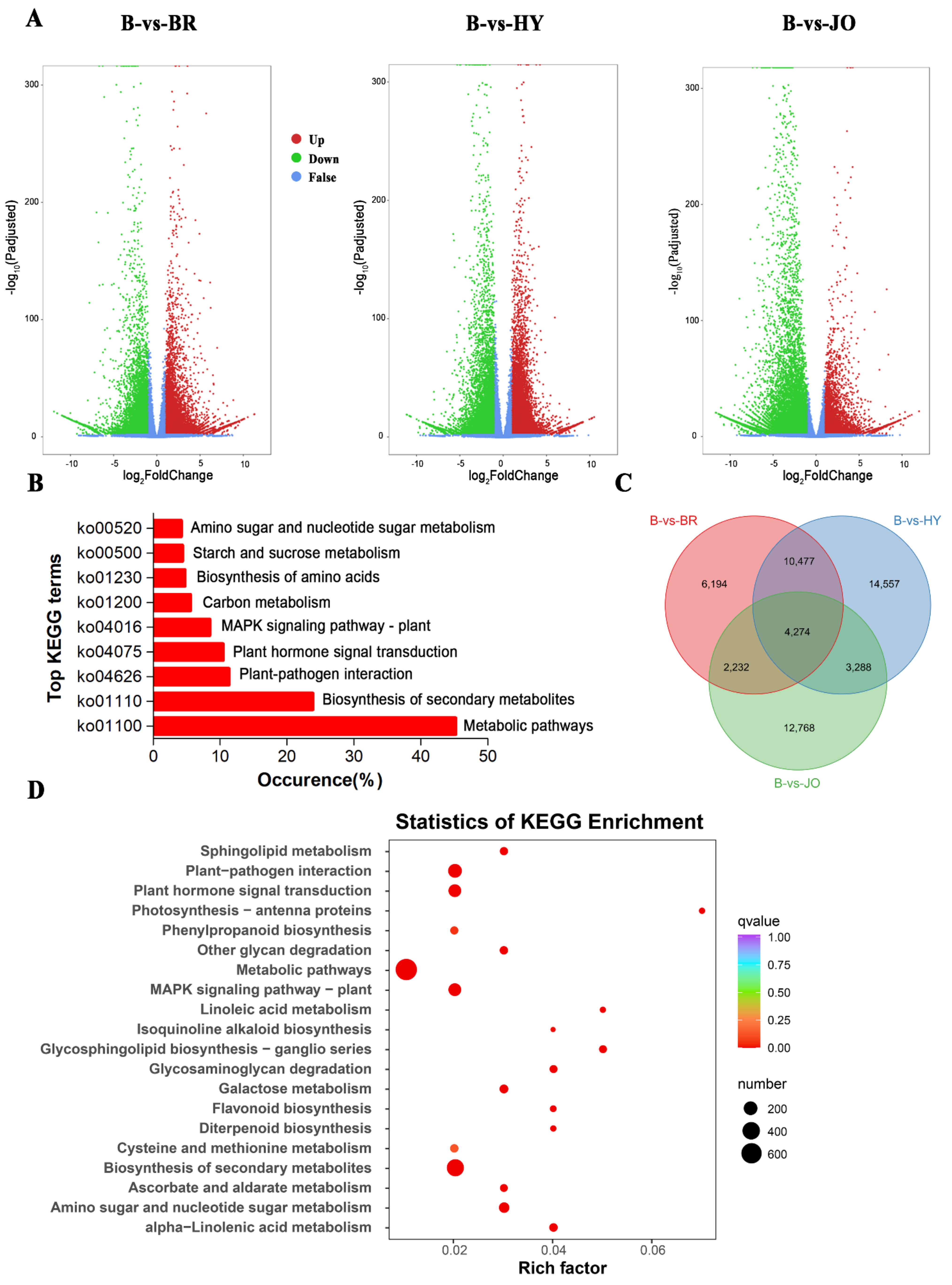
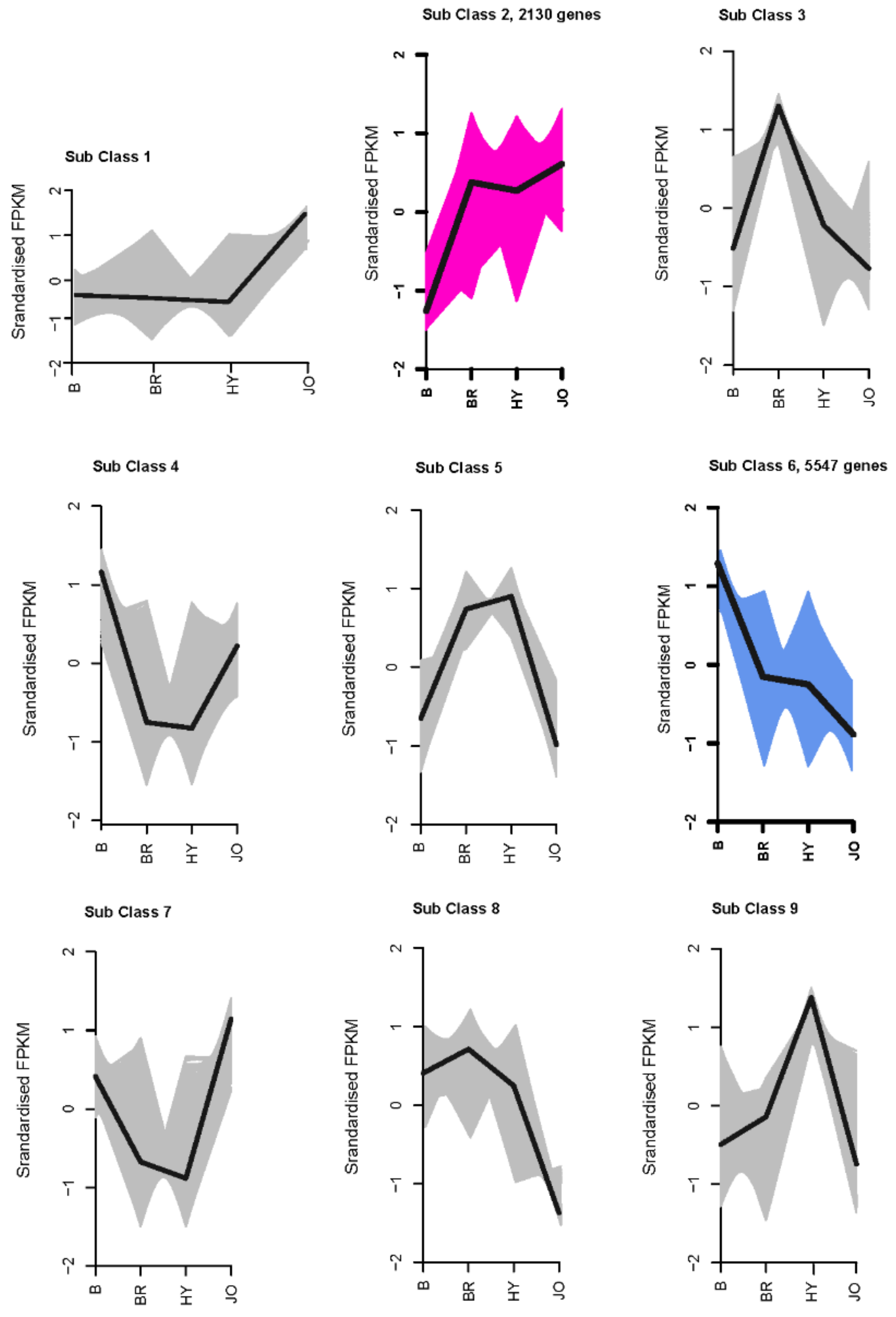
3.4. Differentially Expressed Genes between Pod Skins of the Green and Purple Okra Cultivars
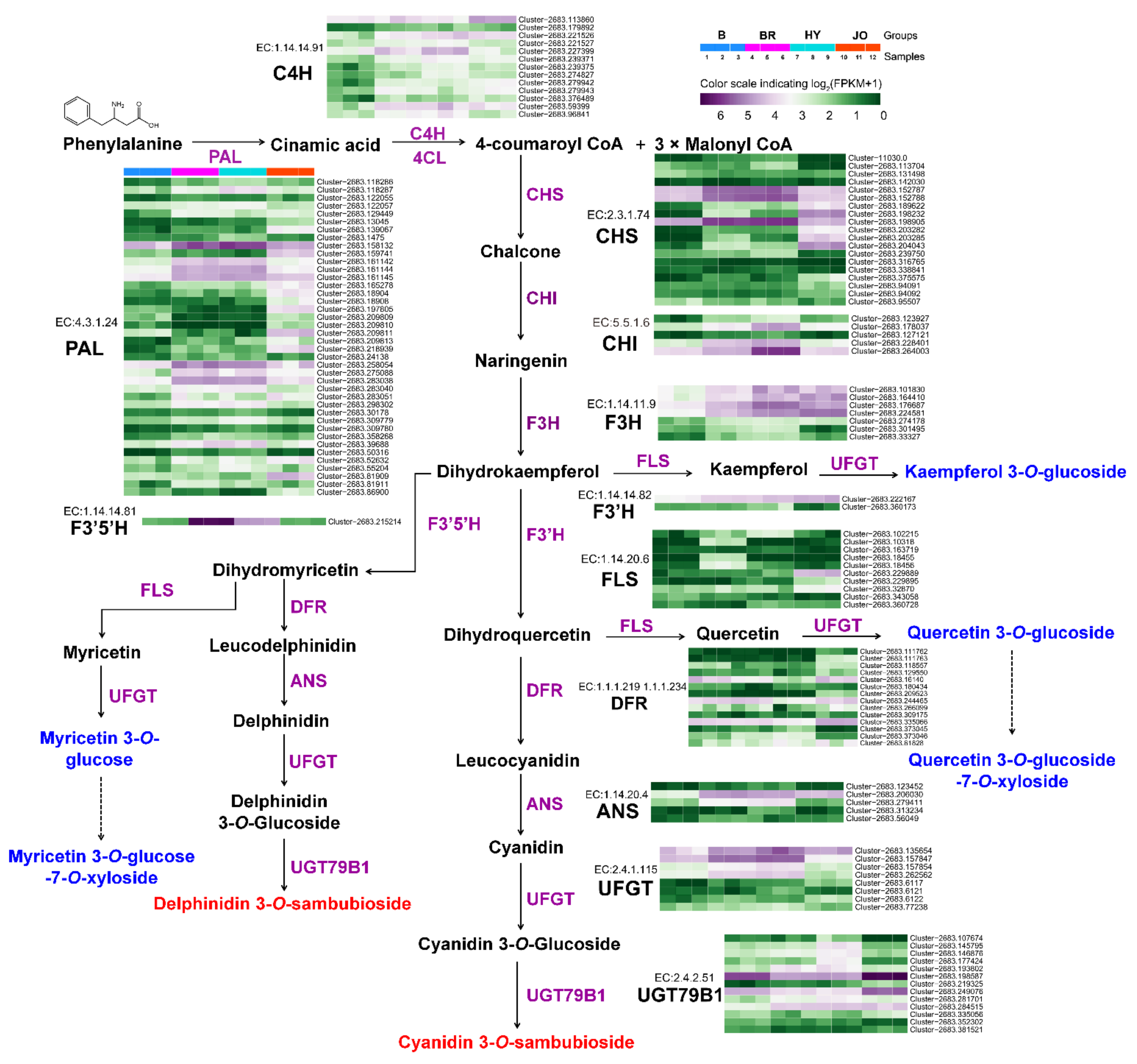
3.5. Expression Analysis of Anthocyanin and Flavonol Biosynthetic Genes in Pod Skins of the Four Okra Cultivars
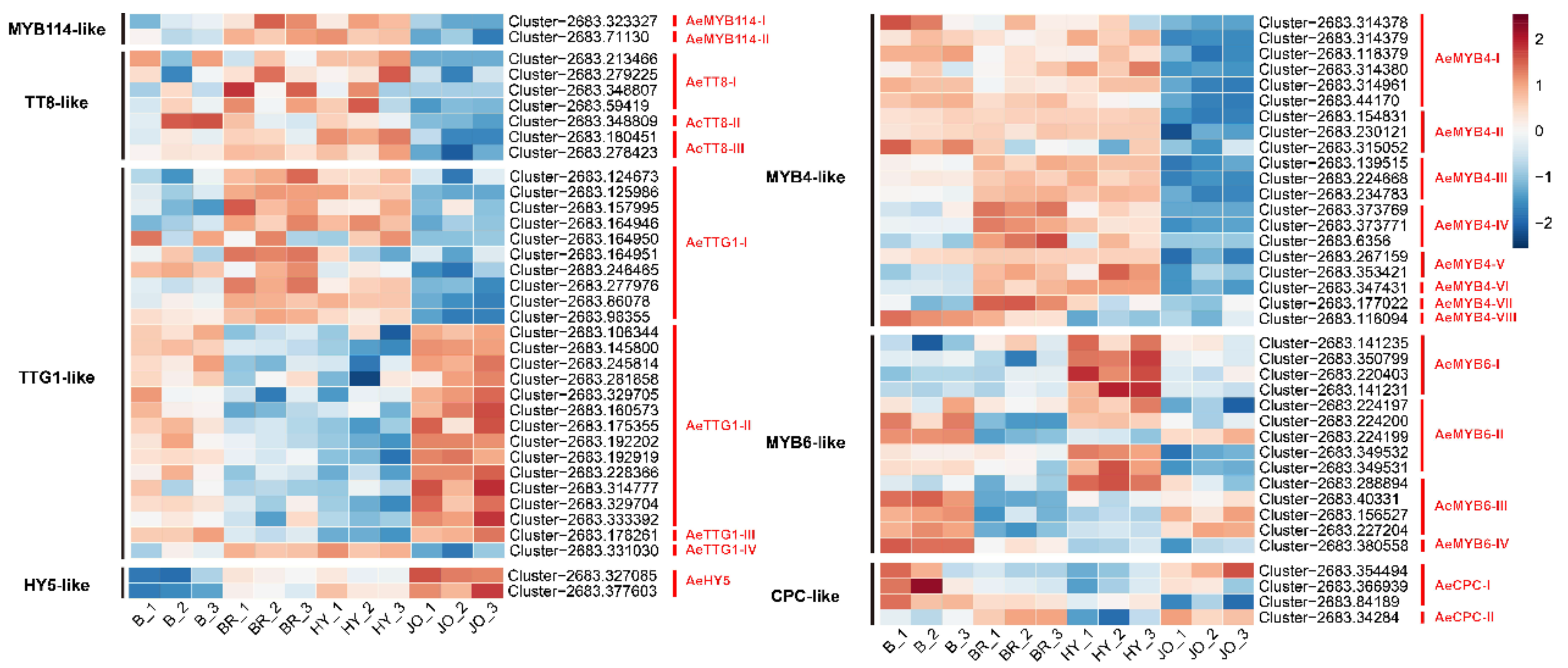
3.6. Expression Analysis of Anthocyanin Biosynthesis Regulatory Proteins in Skins of the Four Okra Cultivars
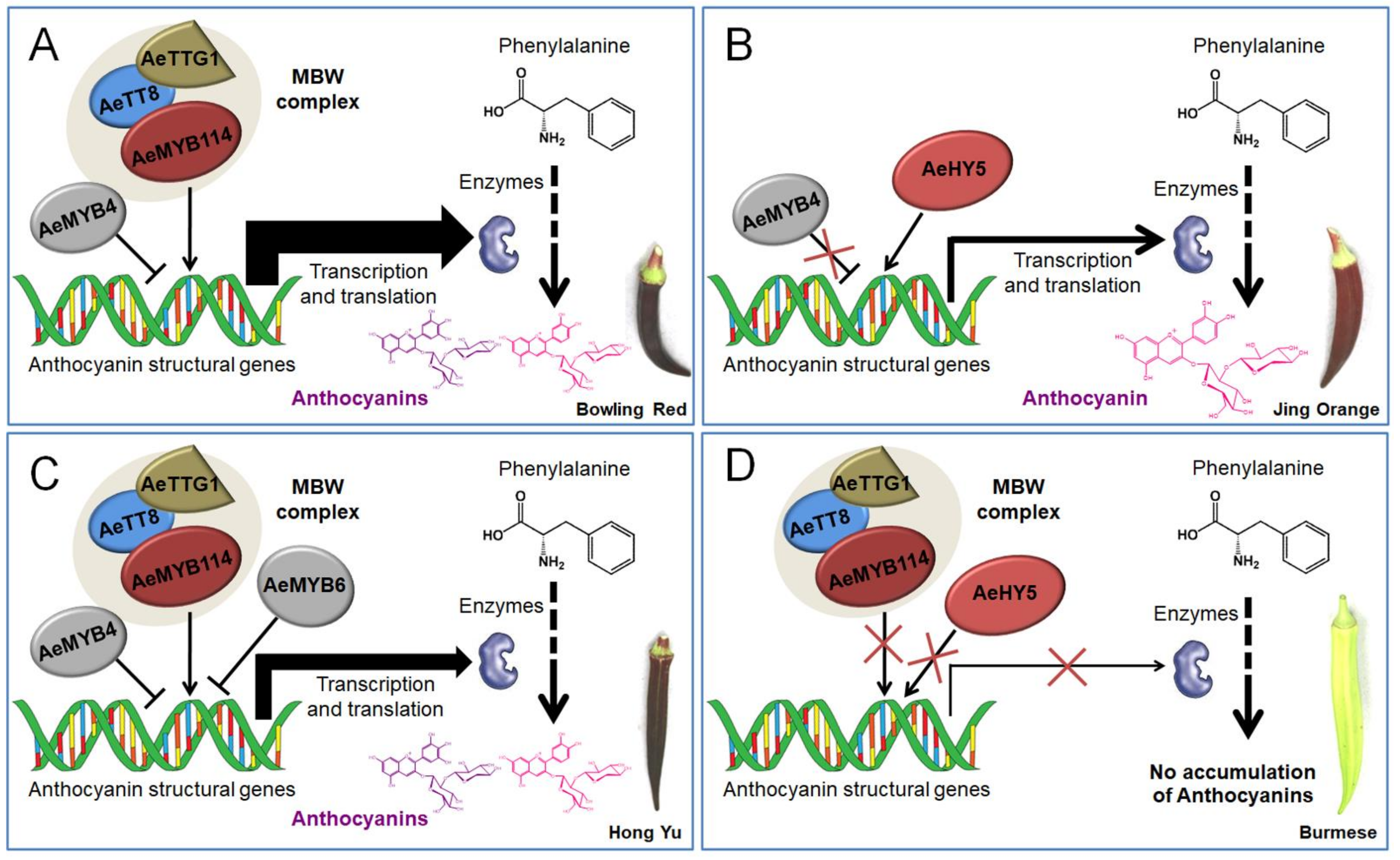
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; Di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 2007, 30, 1381–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenc-Kukula, K.; Jafra, S.; Oszmianski, J.; Szopa, J. Ectopic expression of anthocyanin 5-O-glucosyltransferase in potato tuber causes increased resistance to bacteria. J. Agric. Food Chem. 2005, 53, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Olsen, K.M.; Slimestad, R.; Lea, U.S.; Brede, C.; Lovdal, T.; Ruoff, P.; Verheul, M.; Lillo, C. Temperature and nitrogen effects on regulators and products of the flavonoid pathway: Experimental and kinetic model studies. Plant Cell Environ. 2009, 32, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.P.; Matros, A.; Peterek, S.; Schijlen, E.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef]
- Wallace, T.C.; Giusti, M.M. Anthocyanins-Nature’s Bold, Beautiful, and Health-Promoting Colors. Foods 2019, 8, 550. [Google Scholar] [CrossRef] [Green Version]
- de Pascual-Teresa, S.; Moreno, D.A.; Garcia-Viguera, C. Flavanols and Anthocyanins in Cardiovascular Health: A Review of Current Evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, P.; Bomser, J.A.; Schwartzt, S.J.; He, J.; Magnuson, B.A.; Giusti, M.M. Structure-Function Relationships of Anthocyanins from Various Anthocyanin-Rich Extracts on the Inhibition of Colon Cancer Cell Growth. J. Agric. Food Chem. 2008, 56, 9391–9398. [Google Scholar] [CrossRef]
- Ghosh, D.; Konishi, T. Anthocyanins and anthocyanin-rich extracts: Role in diabetes and eye function. Asia Pac. J. Clin. Nutr. 2007, 16, 200–208. [Google Scholar]
- Li, D.T.; Wang, P.P.; Luo, Y.H.; Zhao, M.Y.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons Control Fruit-Specific, Cold-Dependent Accumulation of Anthocyanins in Blood Oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 repeat protein. Plant Cell 1999, 11, 1337–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.B.; Yuan, L.; Zheng, M.Y.; Xi, Z.M. Differences in Anthocyanin Accumulation Profiles between Teinturier and Non-Teinturier Cultivars during Ripening. Foods 2021, 10, 1073. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Fukushima, A.; Nakabayashi, R.; Hanada, K.; Matsuda, F.; Sugawara, S.; Inoue, E.; Kuromori, T.; Ito, T.; Shinozaki, K.; et al. Two glycosyltransferases involved in anthocyanin modification delineated by transcriptome independent component analysis in Arabidopsis thaliana. Plant J. 2012, 69, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.J.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Osbourn, A.; Ma, P.D. MYB Transcription Factors as Regulators of Phenylpropanoid Metabolism in Plants. Mol. Plant 2015, 8, 689–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracke, R.; Ishihara, H.; Barsch, G.H.A.; Mehrtens, F.; Niehaus, K.; Weisshaar, B. Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J. 2007, 50, 660–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.W.; Constabel, C.P. MYB Repressors as Regulators of Phenylpropanoid Metabolism in Plants. Trends Plant Sci 2019, 24, 275–289. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef] [Green Version]
- Albert, N.W.; Davies, K.M.; Lewis, D.H.; Zhang, H.B.; Montefiori, M.; Brendolise, C.; Boase, M.R.; Ngo, H.; Jameson, P.E.; Schwinn, K.E. A Conserved Network of Transcriptional Activators and Repressors Regulates Anthocyanin Pigmentation in Eudicots. Plant Cell 2014, 26, 962–980. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.F.; Fitzsimmons, K.; Khandelwal, A.; Kranz, R.G. CPC, a Single-Repeat R3 MYB, Is a Negative Regulator of Anthocyanin Biosynthesis in Arabidopsis. Mol. Plant 2009, 2, 790–802. [Google Scholar] [CrossRef]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 2008, 55, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L.; Poole, M.; Jones, M.O.; Kamarainen-Karppinen, T.; Koskimaki, J.J.; Hohtola, A.; Haggman, H.; Fraser, P.D.; Manning, K.; King, G.J.; et al. A SQUAMOSA MADS Box Gene Involved in the Regulation of Anthocyanin Accumulation in Bilberry Fruits. Plant Physiol. 2010, 153, 1619–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. TRANSPARENT TESTA GLABRA2, a trichome and seed coat development gene of Arabidopsis, encodes a WRKY transcription factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Park, E.; Choi, G. PIF3 regulates anthocyanin biosynthesis in an HY5-dependent in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. Plant J. 2007, 49, 981–994. [Google Scholar] [CrossRef]
- Schafleitner, R.; Kumar, S.; Lin, C.Y.; Hegde, S.G.; Ebert, A. The okra (Abelmoschus esculentus) transcriptome as a source for gene sequence information and molecular markers for diversity analysis. Gene 2013, 517, 27–36. [Google Scholar] [CrossRef]
- Sheu, S.C.; Lai, M.H. Composition analysis and immuno-modulatory effect of okra (Abelmoschus esculentus L.) extract. Food Chem. 2012, 134, 1906–1911. [Google Scholar] [CrossRef]
- Huang, J.Y.; Xing, M.H.; Li, Y.; Cheng, F.; Gu, H.H.; Yue, C.P.; Zhang, Y.J. Comparative Transcriptome Analysis of the Skin-Specific Accumulation of Anthocyanins in Black Peanut (Arachis hypogaea L.). J. Agric. Food Chem. 2019, 67, 1312–1324. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Q.Y.; Xie, X.D.; Cai, Y.; Li, J.F.; Feng, Y.L.; Zhang, Y.J. Integrated Metabolomics and Transcriptomics Analyses Reveal the Molecular Mechanisms Underlying the Accumulation of Anthocyanins and Other Flavonoids in Cowpea Pod (Vigna unguiculata L.). J. Agric. Food Chem. 2020, 68, 9260–9275. [Google Scholar] [CrossRef] [PubMed]
- Merken, H.M.; Beecher, G.R. Measurement of food flavonoids by high-performance liquid chromatography: A review. J. Agric. Food Chem. 2000, 48, 577–599. [Google Scholar] [CrossRef]
- El Majdoub, Y.O.; Diouri, M.; Arena, P.; Arigo, A.; Cacciola, F.; Rigano, F.; Dugo, P.; Mondello, L. Evaluation of the availability of delphinidin and cyanidin-3-O-sambubioside from Hibiscus sabdariffa and 6-gingerol from Zingiber officinale in colon using liquid chromatography and mass spectrometry detection. Eur. Food Res. Technol. 2019, 245, 2425–2433. [Google Scholar] [CrossRef]
- Jabeur, I.; Pereira, E.; Caleja, C.; Calhelha, R.C.; Sokovic, M.; Catarino, L.; Barros, L.; Ferreira, I. Exploring the chemical and bioactive properties of Hibiscus sabdariffa L. calyces from Guinea-Bissau (West Africa). Food Funct. 2019, 10, 2234–2243. [Google Scholar] [CrossRef] [Green Version]
- Malacarne, G.; Costantini, L.; Coller, E.; Battilana, J.; Velasco, R.; Vrhovsek, U.; Grando, M.S.; Moser, C. Regulation of flavonol content and composition in (SyrahxPinot Noir) mature grapes: Integration of transcriptional profiling and metabolic quantitative trait locus analyses. J. Exp. Bot. 2015, 66, 4441–4453. [Google Scholar] [CrossRef]
- Boulton, R. The copigmentation of anthocyanins and its role in the color of red wine: A critical review. Am. J. Enol. Viticult. 2001, 52, 67–87. [Google Scholar]
- Medic-Saric, M.; Rastija, V.; Bojic, M. Recent Advances in the Application of High Performance Liquid Chromatography in the Analysis of Polyphenols in Wine and Propolis. J. AOAC Int. 2011, 94, 32–42. [Google Scholar] [CrossRef]
- Shen, D.D.; Li, X.; Qin, Y.L.; Li, M.T.; Han, Q.H.; Zhou, J.; Lin, S.; Zhao, L.; Zhang, Q.; Qin, W.; et al. Physicochemical properties, phenolic profiles, antioxidant capacities, and inhibitory effects on digestive enzymes of okra (Abelmoschus esculentus) fruit at different maturation stages. J. Food Sci. Technol. 2019, 56, 1275–1286. [Google Scholar] [CrossRef]
- Park, H.L.; Lee, J.C.; Lee, K.; Lee, J.M.; Nam, H.J.; Bhoo, S.H.; Lee, T.H.; Lee, S.W.; Cho, M.H. Biochemical Characterization of a Flavonoid O-methyltransferase from Perilla Leaves and Its Application in 7-Methoxyflavonoid Production. Molecules 2020, 25, 4455. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morishita, T.; Kojima, Y.; Maruta, T.; Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. Arabidopsis NAC Transcription Factor, ANAC078, Regulates Flavonoid Biosynthesis under High-light. Plant Cell Physiol. 2009, 50, 2210–2222. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Chi, C.; Jin, L.J.; Zhu, J.; Yu, J.Q.; Zhou, Y.H. The bZip transcription factor HY5 mediates CRY1a-induced anthocyanin biosynthesis in tomato. Plant Cell Environ. 2018, 41, 1762–1775. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. I | RT II (min) | [M + H]+ (m/z) | MS/MS (m/z) | Compound | Hong Yu | Bowling Red | Burgundy | Jing Orange |
|---|---|---|---|---|---|---|---|---|
| 1 | 4.01 | 597.1500 | 303.05 | Delphinidin 3-O-sambubioside | 3.47 ± 0.13 c | 7.00 ± 0.22 a | 4.29 ± 0.11 b | Nd d |
| 2 | 4.97 | 581.1580 | 287.06 | Cyanidin 3-O-sambubioside | 3.52 ± 0.14 c | 4.05 ± 0.12 b | 3.06 ± 0.10 d | 4.41 ± 0.17 a |
| Total | 6.99 ± 0.16 | 11.05 ± 0.25 | 7.35 ± 0.16 | 4.41 ± 0.17 |
| No. a | RT b (min) | λ Max: Band I, Band II (nm) | [M+H]+ (m/z) | MS/MS (m/z) | Compound |
|---|---|---|---|---|---|
| 1 | 7.07 | 258, 354 | 613.1729 | 481.10/319.05 | Myricetin 3-O-glucose-7-O-xyloside |
| 2 | 8.03 | 258, 358 | 481.1045 | 319.05 | Myricetin 3-O-glucoside |
| 3 | 8.54 | 258, 354 | 597.0500 | 465.11/303.05 | Quercetin 3-O-glucose-7-O-xyloside |
| 4 | 10.02 | 258, 354 | 465.1090 | 303.05 | Quercetin 3-O-glucoside |
| 5 | 11.07 | 254, 354 | 611.1694 | 479.12/317.07 | Isorhamnetin 3-O-glucose-7-O-xyloside |
| 6 | 12.22 | 266, 358 | 449.1140 | 287.06 | Kaempferol 3-O-glucoside |
| Okra Cultivar | Flavonoid | Total | |||||
|---|---|---|---|---|---|---|---|
| Myricetin 3-O-Glucose-7-O-xyloside | Myricetin 3-O-Glucoside | Quercetin 3-O-Glucose-7-O-xyloside | Quercetin 3-O-Glucoside | Isorhamnetin 3-O-Glucose-7-O-xyloside | Kaempferol 3-O-Glucoside | ||
| Hongyu | 0.52 ± 0.01 b | 0.08 ± 0.01 | 3.72 ± 0.07 c | 0.34 ± 0.01 c | 0.14 ± 0.01 c | 0.05 ± 0.01 | 4.85 ± 0.02 |
| Bowling Red | 0.80 ± 0.02 a | 0.10 ± 0.01 | 4.07 ± 0.07 b | 0.32 ± 0.01 c | 0.13 ± 0.01 c | 0.05 ± 0.01 | 5.47 ± 0.03 |
| Burgundy | 0.56 ± 0.02 c | 0.07 ± 0.01 | 4.42 ± 0.11 a | 0.36 ± 0.01 b | 0.13 ± 0.01 c | 0.05 ± 0.01 | 5.59 ± 0.04 |
| Jing Orange | Nd | Nd | 4.55 ± 0.16 a | 0.39 ± 0.01 a | 0.10 ± 0.05 c | 0.04 ± 0.01 | 5.08 ± 0.08 |
| Alabama Red | Nd | Nd | 0.66 ± 0.03 e | 0.04 ± 0.01 e | 0.18 ± 0.02 b | Nd | 0.88 ± 0.02 |
| Hill Country | Nd | Nd | 0.74 ± 0.06 e | 0.05 ± 0.01 e | 0.23 ± 0.03 ab | Nd | 1.02 ± 0.03 |
| Lvruyi | Nd | Nd | 0.74 ± 0.05 e | 0.06 ± 0.01 e | 0.10 ± 0.01 d | Nd | 0.90 ± 0.02 |
| Clemson Spineless | Nd | Nd | 0.47 ± 0.02 e | 0.03 ± 0.01 ef | 0.19 ± 0.01 b | Nd | 0.69 ± 0.01 |
| Eagle Pass | Nd | Nd | 0.64 ± 0.02 e | 0.03 ± 0.01 ef | 0.20 ± 0.01 b | Nd | 0.87 ± 0.01 |
| Gold Coast | Nd | Nd | 0.37 ± 0.04 f | 0.02 ± 0.01 f | 0.14 ± 0.02 c | Nd | 0.53 ± 0.02 |
| Star of David | Nd | Nd | 0.91 ± 0.06 c | 0.09 ± 0.01 d | 0.19 ± 0.02 b | Nd | 1.19 ± 0.03 |
| Stubby | Nd | Nd | 0.99 ± 0.02 c | 0.10 ± 0.01 d | 0.26 ± 0.01 a | Nd | 1.35 ± 0.01 |
| Burmese | Nd | Nd | 0.29 ± 0.02 f | Nd | 0.08 ± 0.01 d | Nd | 0.37 ± 0.01 |
| Emerald | Nd | Nd | 0.37 ± 0.05 f | 0.01 ± 0.01 f | 0.16 ± 0.02 bc | Nd | 0.54 ± 0.02 |
| Perkins’ Long Pod | Nd | Nd | 0.94 ± 0.02 c | 0.08 ± 0.01 d | 0.12 ± 0.01 cd | Nd | 1.14 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, T.; Zhao, Q.; Xie, X.; Li, Y.; Chen, Q.; Cheng, F.; Tian, J.; Gu, H.; Huang, J. Comparative Transcriptome Analysis of the Accumulation of Anthocyanins Revealed the Underlying Metabolic and Molecular Mechanisms of Purple Pod Coloration in Okra (Abelmoschus esculentus L.). Foods 2021, 10, 2180. https://doi.org/10.3390/foods10092180
Zhang Y, Zhang T, Zhao Q, Xie X, Li Y, Chen Q, Cheng F, Tian J, Gu H, Huang J. Comparative Transcriptome Analysis of the Accumulation of Anthocyanins Revealed the Underlying Metabolic and Molecular Mechanisms of Purple Pod Coloration in Okra (Abelmoschus esculentus L.). Foods. 2021; 10(9):2180. https://doi.org/10.3390/foods10092180
Chicago/Turabian StyleZhang, Yanjie, Tianjiao Zhang, Qing Zhao, Xiaodong Xie, Yan Li, Qiyan Chen, Fang Cheng, Jianwei Tian, Huihui Gu, and Jinyong Huang. 2021. "Comparative Transcriptome Analysis of the Accumulation of Anthocyanins Revealed the Underlying Metabolic and Molecular Mechanisms of Purple Pod Coloration in Okra (Abelmoschus esculentus L.)" Foods 10, no. 9: 2180. https://doi.org/10.3390/foods10092180