
Cold Microfiltration as an Enabler of Sustainable Dairy Protein Ingredient Innovation

School of Food and Nutritional Sciences, University College Cork, T12 TP07 Cork, Ireland
*
Author to whom correspondence should be addressed.
Foods 2021, 10(9), 2091; https://doi.org/10.3390/foods10092091
Submission received: 30 July 2021
/
Revised: 29 August 2021
/
Accepted: 1 September 2021
/
Published: 4 September 2021
(This article belongs to the Special Issue Recent Advances and Trends in the Dairy Field)
Abstract
:Classically, microfiltration (0.1–0.5 µm) of bovine skim milk is performed at warm temperatures (45–55 °C), to produce micellar casein and milk-derived whey protein ingredients. Microfiltration at these temperatures is associated with high initial permeate flux and allows for the retention of the casein fraction, resulting in a whey protein fraction of high purity. Increasingly, however, the microfiltration of skim milk and other dairy streams at low temperatures (≤20 °C) is being used in the dairy industry. The trend towards cold filtration has arisen due to associated benefits of improved microbial quality and reduced fouling, allowing for extended processing times, improved product quality and opportunities for more sustainable processing. Performing microfiltration of skim milk at low temperatures also alters the protein profile and mineral composition of the resulting processing streams, allowing for the generation of new ingredients. However, the use of low processing temperatures is associated with high mechanical energy consumption to compensate for the increased viscosity, and thermal energy consumption for inline cooling, impacting the sustainability of the process. This review will examine the differences between warm and cold microfiltration in terms of membrane performance, partitioning of bovine milk constituents, microbial growth, ingredient innovation and process sustainability.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction to Microfiltration
Membranes contain small pores of a defined average size, which permit molecules smaller than the pore size to pass through into the permeate stream, while larger molecules that cannot pass through the membrane are concentrated in the retentate. Membrane separation processes have numerous industrial applications, with the pressure-driven processes of microfiltration (MF), ultrafiltration (UF), nanofiltration (NF) and reverse osmosis (RO) being the most well established in the dairy industry with pore sizes of 10–0.1 µm, 0.10–0.01 µm, 10–0.1 nm and <0.1 nm, respectively [1]. Membrane filtration technology is widely used in dairy processing to concentrate, fractionate, isolate, demineralise, defat, and purify different target macro-molecules, such as proteins, phospholipids and lactose.
MF is a key unit operation in dairy processing, in which micrometre-scale particles (e.g., fat globules and bacterial spores) are retained while smaller molecules such as water, minerals, lactose and whey proteins permeate through the membrane. MF membranes cover a wide range of pore sizes, ranging from 0.1–10 µm, and are also associated with the lowest processing pressures (0.01–0.20 MPa) [1,2]. In comparison, higher operating pressures are required for UF, NF and RO with pressures ranging from 0.1–1.0, 1.5–3.0 and 3.0–5.0 MPa, respectively [1]. MF is increasingly being used in the dairy industry to reduce the bacterial load by removing bacteria cells (0.4–2.0 µm) and spores from skim milk prior to subsequent processing [3,4]. Based on the size difference between casein micelles (50–500 nm) and whey proteins (4–8 nm), MF technology is capable of fractionating the major proteins in milk in a chemical-free (e.g., no acids or bases required for pH adjustment) manner which is perceived as a “clean” approach when compared to alternative methods for protein fractionation (e.g., precipitation techniques). Thus, tight MF membranes (e.g., 0.1 µm) allow for the retention of casein micelles while serum proteins are removed, producing a casein-rich concentrate containing micelles in their native form, known as micellar casein concentrate (MCC), and a permeate enriched in whey proteins in their native form, known as ‘ideal’ or ‘native’ whey. This review will examine the effects of performing MF at low temperatures and how altering processing temperature ultimately affects membrane performance, fouling, protein and mineral partitioning, as well as microbial biodiversity and growth. The potential for ingredient innovation and the overall sustainability of the process are also considered. The studies cited throughout the review report the pasteurisation of feed material prior to filtration, with filtration operated under concentration mode, unless otherwise stated.
2. Choice of Membranes
There are a wide variety of MF membranes available to the dairy industry, differing both in the configuration and membrane construction material. Both the choice of membrane material and configuration depend on the intended application.
2.1. Membrane Structures and Configurations
The internal structure of MF membranes can be symmetric (isotropic) or asymmetric (anisotropic); isotropic membranes have pores of uniform size over the entire cross-section of the membrane, while anisotropic membranes have multiple layers, each with different structures and pore size (Figure 1) [5,6]. Due to the uniform structure of isotropic membranes, particles are often retained within the internal structure, resulting in reduced flux due to plugging of the pores [7]. Asymmetric membranes consist of a ‘skin’ layer on the surface of the membrane which acts as the actual selective barrier; below this thin ‘skin’ layer is a thicker porous sub-layer which has little effect on separation but provides mechanical support as well as allowing improved flux [7]. Asymmetric membranes are the most common commercially available membranes [8].
Membranes are available in hollow fibre, tubular, capillary and flat sheet geometries. Hollow fibre and tubular membranes have similar geometries, differing in terms of the tube diameters, with the former having outer diameters ranging from 0.05–0.5 mm while the latter have outer diameters of 5–25 mm [9]. For continuous membrane operations, these membranes are arranged into membrane elements/modules, with the type of module design depending on the membrane type (e.g., flat sheet membranes can be arranged into plate and frame configuration or spiral wound modules). There are four general categories of membrane configuration/design: spiral-wound, tubular, plate and frame and hollow fibre with both ceramic and polymeric materials available for use [8]. The choice of membrane module used depends on multiple factors, such as the capital costs associated with the module, resistance to fouling, ease of cleaning and the properties of the feed material (e.g., viscosity).
2.2. Choice of Membrane Material
The membrane material used in the MF of dairy streams is primarily polymeric, although ceramic materials are also used. There are a wide variety of organic (polymeric) and inorganic (ceramic) materials that can be used for the manufacture of membranes, although the choice of material used depends on the intended application, as each has specific advantages and disadvantages. Besides, the choice of membrane material, the cost associated with purchasing the membrane, cost of installation and operation, and the expected membrane lifetime are often the main factors determining membrane choice. The differences between polymeric and ceramic membranes in terms of milk protein fractionation has received some attention in recent years [10,11], with the efficiency of removal of serum proteins often being higher for ceramic membranes [12,13].
2.2.1. Polymeric
The main polymers used to create MF membranes include polysulphone/polyethersulphone (PS/PES) and polyvinylidene fluoride (PVDF). PS and PES membranes have wide pH, temperature and chemical tolerances, as well as having the advantage of being able to be manufactured into a wide variety of configurations with a wide range of pore sizes. However, they are often limited to low operating pressures due to low mechanical strength. PVDF membranes possess many of the advantages of PES membranes, while they have narrower temperature and pH tolerances. Polymeric membranes have a broader pore size distribution compared to ceramic membranes and are relatively hydrophobic, which can facilitate protein adsorption [14]. The surface of these membranes can also be modified to alter the surface chemistry, modifying the way in which the membrane interacts with components of the feed system. Alternatively, hydrophobic polymers such as PVDF can be directly modified during the membrane production process to reduce hydrophobicity through the addition of hydrophilic polymers [15]. A hydrophilic membrane is most desirable in the processing of dairy streams to minimise binding of hydrophobic proteins to the membrane surface.
2.2.2. Ceramic
Ceramic, also known as “inorganic”, membranes are formed by combining metals such as titanium or aluminium in different forms (e.g., oxides) with support materials (e.g., aluminium, stainless steel, silicon carbide). Generally, ceramic membranes are available in tubular form as either a single-channel tube or, more commonly, a multichannel element, with the latter design increasing packing density and mechanical strength. However, ceramic membranes have fewer disadvantages when compared to polymeric membranes. Due to the inorganic nature of the materials used to manufacture these membranes, they are inert to many chemicals and have wide temperature (>100 °C) and pH (0.5–13.5) tolerances [14]. As ceramic membranes can tolerate high temperatures, they can be sterilised, unlike polymeric membranes. However, ceramic membranes are both more brittle and heavier than their polymeric counterparts. In addition, the higher capital and running costs for ceramic membranes have made polymeric membranes an attractive alternative for milk processers. Inorganic membranes have longer life spans than polymeric membranes, with the life span of polymeric membranes depending on the frequency of use and the nature of cleaning (e.g., temperature at which cleaning is performed, type and strength of chemicals). Ceramic membranes also have the capability of allowing backflushing in which backpressure is applied to remove foulants from the surface of the membrane, helping to restore flux. In addition to this, to reduce fouling on ceramic MF membranes during filtration, a high cross-flow velocity can be used (5–7 m/s), which increases wall shear stress and therefore improves the efficiency of removal of loose foulant material [16].
3. Cold Microfiltration in Dairy Processing
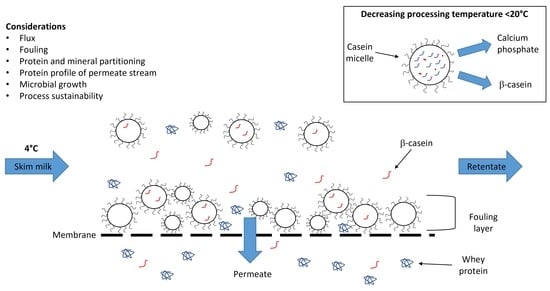
Both MF and UF at low temperatures (i.e., <20 °C) have become increasingly common in the dairy industry due to evidence of reduced fouling, which may allow for longer run times [17,18,19], the opportunity to produce new ingredients with targeted protein profiles (i.e., β-casein-enriched ingredients), and reduced levels of denatured whey protein (i.e., higher levels of native whey protein) [20]. The differences between warm and cold MF in terms of operating conditions, membrane performance and partitioning are discussed below.
3.1. Temperature Range
Traditionally, both MF and UF of milk are performed at temperatures of 45–55 °C [21], with associated advantages of higher initial fluxes, resulting from lower viscosity and efficient removal of whey proteins [19,22]. However, the filtration of milk within this temperature range can cause issues with fouling and the growth of thermophilic bacteria. As a result, some dairy processors are transitioning from traditional warm MF (45–55 °C) towards cold MF. The term “cold”, in the dairy industry, when applied to MF (and also UF), covers a broad temperature range, with temperatures <20 °C generally being considered as low temperature filtration [23]. Selecting the temperature at which MF is performed is an important consideration, as it can have significant effects on membrane performance, the degree and type of fouling, partitioning of milk constituents and energy requirements of the plant, as well as the microbial quality of the ingredients produced.
3.2. Membrane Performance and Fouling
Operating at low temperatures is associated with lower permeate flux due to higher viscosity of the permeate stream, as stated by Darcy’s law:
where J is the permeate flux, TMP is the transmembrane pressure, η is the viscosity of the fluid permeating the membrane at a specific processing temperature and R is the overall hydraulic resistance consisting of resistance of the clean membrane, reversible and irreversible fouling. Thus, when performing filtration at cold temperatures, longer run times may be required to achieve the same volume concentration factor compared to operating at higher temperatures. One of the biggest challenges of membrane filtration is fouling, in which solutes accumulate at the membrane surface or within the membrane itself [24]. Although, during warm MF, initial flux levels are higher than for cold MF, fouling is more extensive with the former process [19]. McCarthy et al. [19] filtered unpasteurised skim milk using a 0.14-µm pore size ceramic membrane at 8.9 and 50 °C and reported that permeate flux was lower for cold MF compared to warm MF, although flux decreased rapidly during the warm MF process due to higher levels of fouling. Hartinger et al. [25] reported that MF of skim milk at 50 °C, under total recirculation mode, using a 0.1-µm spiral-wound PVDF membrane, resulted in a more rapid decline in flux and a more significant reduction in protein permeation throughout filtration, compared to MF at 10 °C. Similar findings have been reported for UF membranes, with Luo et al. [17] and Ng et al. [26] observing lower fouling at low processing temperatures (i.e., 10–15 °C) compared to higher temperatures (i.e., 50 °C). Barukčić et al. [27] measured water flux before and after MF of whey under total recirculation mode, using ceramic membranes with different pore sizes (0.1, 0.5 and 0.8 µm), at 20 and 50 °C, and reported that fouling intensity was lower at the lower processing temperature for each pore size. Therefore, even though initial flux levels are lower for cold MF (Figure 2), longer processing times with a more stable flux over time, as a result of lower fouling, could result in a reduced number of cleaning cycles and therefore reduced downtime during cleaning in place (CIP) and consumption of water and cleaning chemicals [28].
These studies all indicate that more extensive fouling occurs when filtration is performed at high processing temperatures (i.e., 50 °C) compared to low processing temperatures (i.e., 10 °C). Microfiltration of dairy streams at these high processing temperatures (45–55 °C) can also result in partial unfolding of whey proteins, particularly β-lactoglobulin, which can lead to more extensive fouling. β-lactoglobulin has a free cysteine group which becomes accessible as the protein unfolds. At temperatures up to 60 °C, reversible dissociation of β-lactoglobulin dimers to monomers and reversible unfolding occurs, resulting in increased sulfhydryl group reactivity [29], which can increase protein–protein interactions. Studies have shown that performing filtration at higher temperatures (i.e., 50 °C) results in a more compact fouling layer compared to low temperatures (i.e., 10 °C; Figure 2) [18,25,26]. Steinhauer et al. [18] reported that MF of β-lactoglobulin suspensions at 10 °C, with a 0.1-µm pore size, resulted in the formation of a loosely packed and highly permeable fouling layer; on the other hand, MF at temperatures ≥40 °C resulted in the formation of a thicker and more compact fouling layer with a higher fouling resistance, which was attributed to increased protein hydrophobicity and sulfhydryl group reactivity.
The differences in membrane performance and the extent of fouling at high and low temperatures is also evident when comparing permeate flux before and after increasing TMP. Hartinger et al. [25] examined the effect of a stepwise increase in TMP and the subsequent stepwise reduction in TMP on filtration resistance during the MF of skim milk at 10 and 50 °C. At 50 °C, a distinct hysteresis was observed in the flux data, indicating extensive accumulation of deposits on the membrane, whereas at 10 °C there was no distinct hysteresis in flux after the pressure cycle. The results suggest that the compression of the deposit layer at 10 °C is largely reversible, in contrast to the deposit layer formed at 50 °C. The high pressure, coupled with the high processing temperature, may allow for increased protein–protein interactions and crosslinking of foulants, resulting in a more compressed fouling layer than that generated at lower processing temperatures. Thus, although warm MF is associated with higher initial fluxes, extensive fouling can reduce membrane life and alter filtration performance, while cold MF can provide stable flux with less extensive fouling, albeit with lower flux.
3.3. Composition of the Fouling Layer
During the MF of skim milk and other dairy streams (e.g., whey), fouling occurs, resulting in the formation of a cake layer varying in composition, with proteins and minerals (e.g., calcium phosphate) being the principal fouling materials [5,24,30,31]. Protein fouling is caused by the deposition and adsorption of proteins (whey proteins and casein) onto the surface of the membrane or within the pores of the membrane [31,32], while mineral fouling is largely due to calcium, facilitating protein–protein and protein-membrane interactions. Tan et al. [24] found both whey protein and casein to be the primary foulants during cold MF of raw skim milk using a 1.4 µm ceramic membrane, while the contribution of minerals to membrane fouling was minor. The solubility of calcium phosphate is inversely proportional to temperature; thus, calcium phosphate deposition/scaling on the membrane surface or within pores during cold MF is likely to be less significant than during warm MF. Extensive membrane fouling by proteins during MF not only reduces the permeation of proteins through the membrane but is also associated with reduced membrane selectivity, reduced yield, and increased membrane cleaning times [25,33].
3.4. Microbial Impact
MF with nominal pore sizes of 0.8–1.4 µm can be used to reduce the microbial load of milk, such as in the production of extended shelf-life milk [34] and in the production of infant formula-grade milk powders [35]. It also provides the dairy industry with MF permeate with improved microbial quality prior to the generation of milk-derived protein ingredients. However, if this is not performed, and membranes with pore sizes of 0.1 µm are employed in the fractionation of milk proteins, bacteria present are retained and concentrated in the retentate. In the dairy industry, MF systems are operated in continuous mode for long periods of time, which can result in the accumulation of microorganisms and support the formation of biofouling on the MF membrane. Filtration in the dairy industry is often performed at temperatures suitable for the growth of microorganisms; in addition to this, high TMP, lack of mechanical cleaning and constant flow of feed through the membrane make membranes susceptible to biofilm formation. Therefore, the temperature at which filtration is performed is an important consideration for processors, as it influences microbial growth and the bacterial community that can grow (Figure 3).
In order to delay and/or limit biofilm formation and microbial growth, the temperature at which MF is performed can be reduced. Filtration at high temperatures (i.e., 45–55 °C) can provide optimum conditions for the growth of thermophilic bacteria, which can proliferate at temperatures between 45–70 °C [36,37]. Common thermophilic bacteria such as Anoxybacillus flavithermus, Bacillus spp. and Geobacillus spp. can produce heat-stable proteases and lipases, with important implications for the quality of dairy products; their spore-forming and biofilm-forming abilities also present challenges, as they are difficult to remove and/or inactivate and are also capable of producing acid on growth, reducing pH [28,36,38]. Studies have indicated that operating at high temperatures can result in a faster rate of microbial growth and biofilm formation than at low temperatures. Chamberland et al. [39] reported that the rate of biofilm formation on UF membranes during skim milk processing at 15 °C was slower than at 50 °C. These authors found that bacterial growth was significantly higher at 50 °C, with the number of 16S rRNA gene copies on the membrane increasing from 3.21 ± 0.12 to 8.83 ± 1.58 log10 gene copies per cm2 after 15 h of processing, while, at 15 °C, the number of gene copies on the membrane only increased from 3.31 ± 0.24 to 3.86 ± 0.58 gene copies per cm2 after 48 h of filtration.
During filtration of milk at low temperatures (<15 °C), psychrophiles and psychrotrophic bacterial genera, such as Pseudomonas, Psychrobacter and Corynebacterium are capable of growing [39]. Facultative psychrotrophs can grow within a broad temperature range (0–40 °C), occurring at higher numbers in milk compared to obligate psychrophiles (0–15 °C) [37]. The optimal growth temperature for most psychrotrophic bacteria is between 20–30 °C; therefore, the extent of proliferation during cold MF is low [40]. Operating at the upper end of the cold MF range can result in the growth of some mesophilic organisms [37]. However, the growth rate of such mesophilic bacteria (e.g., lactic acid bacteria) is low at these temperatures (i.e., 15 °C). As these higher temperatures (15–20 °C) facilitate the growth of mesophilic bacteria and are close to the optimal metabolic activity of psychrotrophic bacteria, such temperatures are generally avoided for MF in dairy processing. Schiffer and Kulozik [28] examined the effect of temperature (10, 14, 16, 20 °C and 55 °C) on microbial growth during MF of skim milk under feed-and-bleed mode (permeate and 10 L h−1 of retentate was continuously removed during MF and replaced with skim milk), using a 0.1 µm PES membrane; the authors reported that filtration at 55 °C was deliberately stopped after 10 h due to a sudden drop in pH, attributing this to microbial activity. Within the cold MF range, microbial growth was highest at 20 °C, followed by 16 °C and 14 °C taking 8, 10 and 17 h, respectively, to reach the critical colony forming unit level of 105 cfu mL−1; at 10 °C, microbial counts remained almost constant over 24 h of MF. The bacterial communities responsible for the increasing microbial counts at 16 and 20 °C in the aforementioned study were not determined, and so it was not clear whether mesophilic bacteria made a considerable contribution towards microbial counts at the upper end of the cold MF range.
In order to improve the microbial quality of ingredients produced using filtration, there has been a trend towards cold MF, as the rate of microbial growth and biofilm formation is lower than that of warm MF. Therefore, performing MF at low temperatures (i.e., ≤10 °C) may allow for longer run times before microbial growth becomes an issue for hygiene standards. However, reducing the temperature at which filtration is performed results in reduced flux and can also increase energy demands. Therefore, research on finding the optimum temperature for maintaining microbial growth and biofilm formation low while also optimising filtration performance (e.g., permeate flux) is still required.
3.5. Protein Partitioning
During the MF of dairy streams, a cake layer is formed, which results in the membrane becoming less permeable; this fouling layer is known to act as a secondary membrane, altering the permeate flux and the selectivity of the membrane. There have been numerous studies comparing the effect of temperature, as well as differences between ceramic and polymeric membranes, on the efficiency of protein fractionation [11,12,18,25,41]. Hartinger et al. [25] studied the influence of temperature and TMP on protein fractionation during MF of skim milk using a 0.1-µm membrane. The authors reported that, at 50 °C, the initial rate of reduction in protein permeation was higher than that at 10 °C, with MF at 50 °C also being associated with a higher mean reduction in the permeation of protein (2.8% h−1 vs. 1.2% h−1 at 50 and 10 °C, respectively) between 40 and 180 min of MF; this was attributed to a more extensively fouled membrane and temperature-induced aging of the deposit layer at 50 °C.
Although many of these studies have examined the effects of temperature on protein partitioning at either one temperature or at two different temperatures (often involving a comparison between low and high temperatures), there is limited information available on the effect of different temperatures within the cold MF range on protein partitioning and fouling. Jarto et al. [41] reported that MF of skim milk at 13 °C resulted in higher whey protein permeation compared to MF at 5 °C. Similar results were reported by France et al. [33], who observed that MF of skim milk at 12 °C resulted in higher concentrations of β-lactoglobulin and α-lactalbumin in permeate, compared to at 4 and 8 °C, although differences between temperatures were not significant. Thus, temperature affects protein partitioning during filtration primarily through differences in the form and extent of fouling. These studies indicate that processing temperature influences whey protein permeation. MF of milk at low temperature (i.e., 4 °C) also results in the permeation of casein, in particular β-casein, through the membrane and into the permeate, resulting in the generation of a β-casein-enriched whey, a phenomenon that is not associated with MF performed warm. Therefore, not only does the temperature at which MF is performed influence whey protein permeation, but also the protein profile of the resulting permeate and retentate.
The choice of membrane material can also impact the efficiency of whey protein permeation and the composition of the final permeate and retentate streams. Zulewska et al. [12] examined the efficiency of serum protein removal from the MF of skim milk at 50 °C, using both spiral-wound polymeric (0.3 µm) and two ceramic membranes (uniform transmembrane pressure and graded permeability MF systems) of equivalent pore size (0.1 µm). The authors reported that permeate generated using ceramic membranes had a higher concentration of protein than that generated using the polymeric membrane. The use of ceramic MF membranes resulted in higher levels of β-lactoglobulin in the permeates, with an overall higher efficiency of whey protein removal than the spiral-wound polymeric membrane (64.4–61.0% vs. 38.6%). The authors suggested that the difference observed were, in part, due to the hydrophilic nature of ceramic membranes resulting in lower protein adsorption to the membrane when compared to the hydrophobic polymeric membrane. Similar results have since been reported by Carter et al. [13] and Beckman et al. [42].
3.6. Mineral Partitioning
Performing MF at low temperatures changes the composition of final permeate, not just in terms of the protein profile, but also of the mineral profile. As temperature decreases, colloidal calcium phosphate (CCP) within casein micelles solubilises, increasing the concentration of calcium and other minerals in the serum phase [43]. Therefore, when MF is performed at low temperatures (i.e., 4 °C), there is a higher concentration of soluble minerals (e.g., calcium) that can permeate the membrane, resulting in a higher concentration of ash in the permeate [44]. There have been numerous studies detailing the effect of processing temperature on the partitioning of calcium during both UF and MF of skim milk; the authors of these studies report that reducing the temperature at which filtration is performed increases the concentration of total and/or ionic calcium in the permeate [17,44,45,46]. Crowley et al. [47] analysed MCC powder produced by MF of skim milk at low (<10 °C) or high temperature (50 °C) and reported that the MCC powder generated at cold temperatures had lower levels of calcium and phosphorus compared to that made from warm MF, while levels of monovalent ions were relatively unchanged. The authors also studied the effect of MF on the rehydration characteristics of MCC powders and reported that MCC generated from cold MF had superior dispersion characteristics, with 50–60% less sedimentation following centrifugation; this was partially attributed to lower Ca content.
The presence of high concentrations of calcium in β-casein-enriched whey following cold MF (0.1–0.5 µm) of skim milk can lead to challenges with subsequent downstream processing of such novel whey streams. On warming the β-casein-enriched whey, the strength of hydrophobic interactions increases, causing the self-association of molecular β-casein (7–8 nm) and an increase in particle size, ultimately resulting in the formation of β-casein micelles (20–30 nm); this phenomenon is normally reversible on cooling, with β-casein returning to a monomeric state [48,49,50]. However, in the presence of calcium (e.g., 8–15 mM at 37 °C), these β-casein micelles can aggregate via calcium-mediated crosslinking, resulting in a particle size of >1 µm, and leading to precipitation of the aggregated β-casein [48,51]. Therefore, following cold MF of skim milk, a demineralisation step is often applied to the permeate stream to reduce the concentration of soluble minerals, allowing for stable downstream processing or indeed allowing for the controlled, reversible, thermal aggregation of β-casein without incurring precipitation [20]. Thus, MF of skim milk at low temperatures results in increased concentrations of total divalent cations (e.g., calcium) in the permeate, altering the mineral composition of both the permeate and retentate compared to that produced from warm MF, which can have important implications for ingredient functionality.
3.7. Enzyme Partitioning
The principal indigenous milk proteinase, plasmin, can result in challenges for the proteolytic stability of filtration-derived milk protein ingredients. During warm MF of milk using a 0.1-µm membrane, casein micelles are concentrated in the retentate, while whey proteins are removed in the permeate, with such permeate containing many inhibitors of the plasmin system, including β-lactoglobulin. As plasmin is primarily associated with casein micelles through lysine binding and to a lesser extent through electrostatic interactions [52,53], it becomes concentrated in the retentate. Consequently, plasmin activity in the retentate increases, with subsequent diafiltration resulting in further increases in plasmin activity as inhibitors are further removed [54].
During cold MF, β-casein dissociates from the casein micelles into the serum phase, where it passes through the membrane into the permeate stream. Therefore, there remains the question whether temperature-induced changes in casein micelles during cold MF also affects the association of plasmin with the casein micelles. As the principal substrate for plasmin is β-casein, plasmin activity in such permeates may lead to issues with β-casein hydrolysis on further downstream processing and in products incorporating the ingredient. France et al. [44] studied the effect of temperature (4, 8, 12, 16 or 20 °C) on enzyme partitioning during MF of skim milk under total recirculation mode and reported that temperature had no significant effect on plasmin activity in β-casein-enriched permeate; therefore, the effect of temperature on plasmin activity in permeates is not as pronounced as that seen for β-casein. Furthermore, in the purification of β-casein, the serum phase containing many of the inhibitors are removed; thus, in the absence of such inhibitors, plasmin activity may be even greater. The presence of plasmin in permeate streams containing β-casein could result in negative consequences, with plasmin-induced hydrolysis of β-casein potentially reducing levels of intact β-casein, thus reducing its functional properties of value in food formulations.
4. Microfiltration as an Enabler of Dairy Ingredient Innovation
MF of skim milk allows for the generation of specific ingredients, with the temperature at which MF is performed strongly influencing the composition and, therefore, the functionality and applications of the ingredients generated, as discussed below.
4.1. Warm Microfiltration Applications in the Dairy Industry
MF of dairy streams has many different applications depending on the feed material, processing temperature, and pore size. One of the first applications of warm MF in the dairy industry was the removal of residual fat and denatured whey proteins from whey in the production of whey protein isolate (WPI). MF membranes with a pore size of less than 1.0 µm retain any residual fat (i.e., that from the cheesemaking process in sweet whey), phospholipids, and denatured protein, thereby allowing for a higher protein content to be achieved in the final WPI product. This MF process creates a co-product with a high content of protein, fat and phospholipid which can be further processed to produce a whey protein phospholipid concentrate (WPPC). A standard for the composition of WPPC was established in 2015 by the American Dairy Products Institute which state that such products must have a minimum of 50% protein on a dry basis, minimum 12% fat, maximum of 8% ash and maximum of 6% moisture. WPPC is a functional dairy ingredient that can add functionality to ingredients at a low cost (e.g., in cakes as an egg replacement) [55], with its biggest potential use being in nutritional applications due to its rich phospholipid profile. Although MF for WPI production has been traditionally performed at 45–55 °C, transitioning to cold processes could allow for reduced fouling and higher native whey protein levels.
Warm MF in the dairy industry has also been used for the removal of bacteria from milk using a typical pore size of ~1.4 µm [2,56]. Here, the objective is to remove bacterial cells, somatic cells and endospores, while minimising the retention of proteins to maintain the composition of the MF permeate as close to that of the milk feed. Therefore, 1.4-µm membranes are most widely used, allowing for the right balance between high flux, bacteria removal and the permeation of other milk components with little or no rejection. A reduction in the pore size (e.g., 0.5–0.8 µm) of MF membranes results in improved reduction in bacteria but can lead to a more rapid decline in flux and increased retention of casein micelles [57]. In the dairy industry, the removal of bacteria is primarily performed at high temperatures (i.e., 50 °C). Such temperatures allow for the growth of thermophilic spore-formers and can lead to the formation of resistant biofilm formation, while using low-temperature MF can reduce the risk of bacterial growth and proliferation of thermophilic spore-formers. Studies have shown that cold MF of skim milk using ceramic membranes effectively reduces bacterial cells and somatic cells [4,58,59]; therefore, cold MF can be used to reduce bacterial load and somatic cell count while reducing the risk of thermophilic bacterial growth and biofilm formation, which could reduce final product quality. Within the dairy industry, cold MF is now also used to improve the microbial quality of the feed material prior to the production of ingredients, e.g., Micelate PrestigeTM, a product currently produced by FrieslandCampina, is a cold-processed, native micellar casein isolate (MCI) which has undergone cold MF to improve the microbial quality of the ingredient.
Following MF of whole milk using a 1.4-µm membrane, as described above, fat globules (0.1–15 µm) are also retained in the retentate. The milk fat globule membrane (MFGM) retained in this MF retentate can be isolated to produce MFGM material. A recent study reported that MF of raw milk resulted in the production of high purity MFGM material [60]. As MFGM material is often isolated from heavily processed dairy streams (e.g., buttermilk and high fat retentate generated following MF of WPC), the process mentioned offers the dairy industry a practical method to produce MFGM material from a less heavily processed stream.
Warm MF of skim milk, when performed with a 0.1-µm pore size membrane, results in two streams, a casein micelle-enriched retentate and a whey protein-enriched permeate (Figure 4). Performing MF at these high temperatures ensures that β-casein does not dissociate from the micelle and into the serum phase as hydrophobic interactions remain strong. The permeate produced from warm MF of milk is often referred to as ‘ideal’ or ‘native’ whey, with a composition similar to that of sweet whey but is free of many of the contaminants present in sweet whey, such as glycomacropeptide, starter culture, colour residues, lipid material, lactic acid and coagulant. In addition, ideal whey has lower fat content, improved protein quality and higher levels of native whey proteins when compared to sweet whey, as sweet whey is typically subjected to more intense thermal treatment [61,62]. The retentate stream or co-product following diafiltration (UF permeate or RO water) is known as MCC and is enriched in native micellar casein. The production of MCC has multiple applications in nutritional and clinical products and in sports nutrition products, providing a slow release of amino acids [63]. MCC can also be used as a starting feed material in the production of β-casein. Warm MF of cheese milk is also used in the manufacture of cheese, enhancing cheese yield and output. MF instead of UF also provides the added benefit of depleting most of the serum proteins prior to reaching the cheese vat and are therefore the whey is not contaminated with colour residues (e.g., annatto) and other residues from the cheese making process [64].
4.2. Cold Microfiltration
There has been an increasing trend in the use of cold MF in the dairy industry. One of the opportunities cold MF offers is the production of new ingredients with selected protein profiles (e.g., β-casein ingredients).
4.2.1. Influence of Temperature on β-Casein Dissociation
The adoption of cold MF offers dairy processors the opportunity to produce ingredients from skim milk with differentiated protein profiles. Of the four caseins (αs1, αs2, β and κ), β-casein enrichment/isolation is the most viable and widely practised. Bovine β-casein is held within the casein micelle primarily via hydrophobic interactions [65]. Reducing temperature results in a decrease in the strength of hydrophobic interactions, resulting in the dissociation of β-casein from the interior to the surface of the micelle, from where it dissociates into the serum phase of milk [66]. At low temperature (i.e., ≤4 °C), β-casein primarily exists in a monomeric state [67,68], although even at these temperatures polymeric structures are not entirely absent [69]. This phenomenon can be exploited in the production of β-casein-enriched whey during cold MF of milk, because at these low temperatures (i.e., ≤4 °C), monomeric β-casein can permeate through the MF membrane, producing a β-casein-enriched permeate and a β-casein-depleted retentate (Figure 4).
The temperature at which filtration is performed at within the cold MF range has a significant impact on serum β-casein concentration, with increasing processing temperature significantly reducing β-casein concentration in the permeate stream. A study by France et al. [44] reported that increasing processing temperature in the range 4–20 °C during the cold MF of skim milk, under total recirculation mode, reduced the concentration of β-casein in the permeate stream, from 2.02 to 0.53 mg/mL at 4 and 16 °C, respectively, under sub-critical flux conditions. By controlling temperature, the strength of hydrophobic interactions can be manipulated, ultimately influencing the concentration of β-casein in the serum phase that can permeate across the membrane.
The complete retention of caseins is often desired in milk protein fractionation to obtain a whey protein stream free from serum casein. Therefore, the increasing trend towards cold MF presents challenges to dairy processors in the production of native whey, as the purity of the whey protein fraction is affected. A recent study by Schiffer et al. [70] examined the effect of added calcium and pH on milk protein fractionation during the MF of skim milk, under total recirculation mode, using a 0.1-µm ceramic membrane at different temperatures within the cold MF range (10–20 °C). The authors hypothesised, and the results confirmed, that the addition of calcium to skim milk prior to cold MF would reduce casein concentration in the permeate as it has been shown that calcium concentration influences the ratio of soluble and micellar casein, partly by increasing milk protein hydrophobicity [71,72], and thereby reducing the amount of casein in the permeate. At 10 and 14 °C, the strongest effect for β-casein was observed at a level of 5 mM calcium and above; however, addition of >5 mM calcium negatively affected membrane performance and resulted in enhanced deposit formation on the membrane, as well as reducing whey protein permeation.
4.2.2. Production of β-Casein-Enriched Ingredients via Cold Microfiltration
There are different approaches that can be used to generate β-casein and β-casein-enriched whey. There have been numerous studies investigating the permeation of β-casein during cold MF of different feed materials [20,73,74]. The most common starting material for the generation of β-casein-enriched whey is skim milk; however, other materials/ingredients can also be used, such as whole milk, MCC and sodium caseinate, although the latter is rarely used.
There are two common MF approaches that can be utilized in the manufacture of β-casein (Figure 4). One approach is referred to as ‘cold-then-warm’ MF, in which skim milk is first subjected to cold MF often holding at ≤4 °C to facilitate, and maximise, the release of β-casein into the serum phase. Most commonly, a 0.1-µm membrane is used, allowing the serum proteins and serum β-casein to permeate the membrane while the casein micelles are retained, generating a β-casein-depleted retentate [20]. The β-casein-enriched whey can be used as an ingredient without further fractionation, or it can undergo further processing to produce β-casein concentrate or isolate [75]. The β-casein-enriched whey can be ultrafiltered and diafiltered to further enrich and demineralise the permeate; for example, O’Mahony et al. [20] achieved this at low temperature (i.e., 1–6 °C) using a 10-kDa molecular weight cut-off membrane and chilled water as the diafiltration media. As previously mentioned, the removal of calcium and other soluble minerals prevents extensive calcium-mediated crosslinking of β-casein and subsequent precipitation. Following this, the concentrated and demineralised permeate is heated (25–50 °C) to increase hydrophobic interactions, resulting in the controlled self-association of β-casein and an increase in particle size [50]. O’Mahony et al. [20] warmed the permeate to 25 °C and passed it through a 0.5-µm polymeric membrane, resulting in the removal of serum proteins while the micellised β-casein was retained, yielding two streams, a β-casein concentrate retentate and an ‘ideal’ whey permeate.
The second approach can be referred to as ‘warm-then-cold’ MF, wherein warm MF (~45–55 °C; 0.1–0.5 µm) of the skim milk is performed, removing the serum proteins (permeate) while retaining the casein micelles (retentate) and the β-casein due to hydrophobic interactions within the micelles. Following this, cold MF (i.e., 4 °C) of the micellar casein retentate is performed, yielding a β-casein-depleted MCC retentate and a β-casein permeate. The application of membrane filtration allows for the production of high purity β-casein (up to 96% of total casein); however, yields are typically low (≤20%), although yields can be increased through additional β-casein dissociation and recovery of the β-casein depleted retentate [76].
If the feed material for cold MF is MCC rather than skim milk, a β-casein permeate relatively free of serum proteins can be obtained. Schäfer et al. [74] microfiltered MCC at ≤5 °C using a 0.1-µm pore size ceramic membrane, heated the β-casein permeate to 50 °C to achieve self-association of the β-casein, and then subjected the heated permeate to UF (10 kDa) to achieve further enrichment of β-casein. As the original permeate contained little or no whey proteins, an additional MF step to remove whey proteins was not required. Thus, cold MF can be exploited by dairy processors to generate innovative ingredients with tailored protein profiles when feed material containing casein is used.
Polymeric membranes such as PVDF, polyacrylonitrile (PAN) or PES are commonly used for the fractionation of caseins. Crowley et al. [23] studied the processing and protein fractionation characteristics of different polymeric membranes during cold MF of skim milk. The authors reported that PES and PVDF membranes, with equivalent pore size, had essentially identical selectivity; however, the PES membrane resulted in higher flux and lower fouling.
5. Sustainability
The sustainability of a process must consider its effects on environmental, social, economic, and technical aspects with capital and operational costs, consumption of energy, footprints, consumption of chemicals, product loss, valorisation of co-products and membrane fouling some examples of the various criteria that must be considered when assessing the sustainability of membrane processing [77,78]. In terms of protein fractionation, MF creates a green image for the process and the ingredients generated, as alternative techniques involve the addition of chemicals (e.g., pH adjustment) or the generation of large quantities of waste buffer (e.g., chromatography). Considering the factors mentioned and the increasing demand for more sustainable dairy processing, cold MF offers both opportunities and challenges in meeting such requirements. Maximising the active production time of a membrane filtration plant reduces the frequency of cleaning and reduces the consumption of cleaning chemicals and the associated environmental impact. As mentioned throughout this review, cold MF of skim milk offers increased processing times due to reduced fouling and a slower rate of decline in permeate flux. Schiffer and Kulozik [28] used the data obtained from their study regarding flux stability, pH, and bacterial counts at different temperatures (10, 14, 16, 20 and 55 °C) during the MF of skim milk to approximate the number of production cycles and overall annual process times. The authors reported that possible production times were highest for MF at 10 °C (>24 h), while operation for only 7 h was possible for MF at 55 °C; thus, the number of annual production cycles and therefore the number of cleaning cycles was calculated to decrease with decreasing processing temperature. The study highlights that, even though permeate flux at low temperatures is not comparable to that at higher temperatures, the improved microbial quality and sustainable flux enables longer run times and reduced cleaning cycles. Reducing the number of CIP cycles reduces the quantity of water and cleaning agents consumed, reducing the impact of the process on the environment, resulting in a more sustainable process. In addition to this, other factors such as product loss resulting from mixing of product and cleaning solutions and energy requirements need to be considered.
The extent and type of fouling also affects the severity of cleaning required to restore membrane performance to acceptable limits. As discussed, performing MF at high temperatures can result in extensive fouling, as well as the formation of biofilms produced by thermophilic bacteria such as Bacillus spp., which have been shown to be one of the species most resistant to CIP solutions [79]. Therefore, cold MF of dairy streams may result in shorter cleaning times and use of reduced amounts or strengths of cleaning agents, ultimately reducing the environmental impact of the process, as a more loosely packed fouling layer is produced.
Another important consideration in the sustainability of a process is the energy requirement. One of the factors influencing the mechanical and thermal energy requirements during membrane filtration is the temperature at which filtration is performed. As the temperature of skim milk is decreased, the apparent viscosity increases, which increases the pumping energy requirements for feed and retentate recirculation. Méthot-Hains et al. [80] reported that UF of skim milk required to reach a volume concentration factor of 3.6 at 10 °C resulted in a higher total energy consumption compared to at 50 °C; the authors also reported that, at 10 °C, pumping energy was 2.3 times higher than at 50 °C, which can be attributed to the higher viscosity of the feed and retentate during filtration at low temperatures. France et al. [33] studied the effect of temperature on the mechanical and thermal energy consumption during cold MF of skim milk, and reported that operating at 4 °C resulted in considerably higher (~10.6%) mechanical and thermal energy requirements compared to 12 °C. Therefore, although cold MF offers opportunities to improve the sustainability of the process through reduced CIP cycles and therefore reduced consumption of chemicals and water as a result of reduced protein and bio-fouling, increased electrical input will contribute to increased operational costs.
6. Conclusions
MF has become an integral unit operation within the dairy industry, with the emergence of cold MF driven by increased demand for novel ingredients with enhanced functional properties, improved microbial quality, and improved membrane performance. However, a better understanding of the effect of temperature on deposit formation on membranes, in particular different temperatures within the cold MF range, which can ultimately lead to improved protein permeation and flux, is required. Cold MF can result in improved microbiological quality, longer, more sustainable, processing times and reduced fouling, extending the life span of membranes as well as providing dairy processors opportunities to produce new functional milk-derived protein ingredients, all acting as an incentive to shift from warm to cold MF. However, although increased processing times and reduced frequency of CIP can be achieved through cold MF, high operational costs associated with higher energy consumption, means research is required to reduce the energy consumption of the process. Recent literature reports promising concepts for improving the MF of dairy streams, such as developments in surface modification (e.g., charged membranes), offering the potential to improve issues relating to fouling. For the MF of dairy streams, these and other approaches will form a future role in improving efficiency and therefore the sustainability of cold MF in dairy processing.
Author Contributions
Conceptualization, T.C.F. and J.A.O.; investigation, T.C.F.; writing—original draft preparation, T.C.F.; writing—review and editing, T.C.F., A.L.K., S.V.C. and J.A.O.; visualization, T.C.F., A.L.K., S.V.C. and J.A.O.; supervision, A.L.K., S.V.C. and J.A.O.; project administration, T.C.F.; funding acquisition, J.A.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Dairy Processing Technology Centre (DPTC), an Enterprise Ireland initiative, Grant Number TC/2014/0016.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Pouliot, Y. Membrane processes in dairy technology—From a simple idea to worldwide panacea. Int. Dairy J. 2008, 18, 735–740. [Google Scholar] [CrossRef]
- Saboya, L.V.; Maubois, J.L. Current developments of microfiltration technology in the dairy industry. Lait 2000, 80, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Gésan-Guiziou, G. Removal of bacteria, spores and somatic cells from milk by centrifugation and microfiltration techniques. In Improving the Safety and Quality of Milk: Milk Production and Processing; Griffiths, M.W., Ed.; Woodhead Publishing Ltd.: Cambridge, UK, 2010; Volume 1, pp. 349–372. ISBN 9781845694388. [Google Scholar]
- Wang, D.; Fritsch, J.; Moraru, C.I. Shelf life and quality of skim milk processed by cold microfiltration with a 1.4-μm pore size membrane, with or without heat treatment. J. Dairy Sci. 2019, 102, 8798–8806. [Google Scholar] [CrossRef]
- Cheryan, M. Ultrafiltration and Microfiltration Handbook; Technomic Publishing Co., Inc.: Lancaster, PA, USA, 1998. [Google Scholar]
- Strathmann, H.; Giorno, L.; Drioli, E. Introduction to Membrane Science and Technology; Wiley-VCH: Weinheim, Germany, 2011. [Google Scholar]
- Smith, K. Development of membrane processes. In Membrane Processing. Dairy and Beverage Applications, 1st ed.; Tamime, A.Y., Ed.; Wiley-Blackwell: West Sussex, UK, 2013; pp. 16–29. ISBN 9781118457023. [Google Scholar]
- Wang, H.; Zhou, H. Understand the basics of membrane filtration. Chem. Eng. Prog. 2013, 109, 33–40. [Google Scholar]
- Smith, K. Commercial membrane technology. In Membrane Processing. Dairy and Beverage Applications, 1st ed.; Tamime, A.Y., Ed.; Wiley-Blackwell: West Sussex, UK, 2013; pp. 61–79. ISBN 9781118457023. [Google Scholar]
- Espina, V.S.; Jaffrin, M.Y.; Ding, L.H. Comparison of rotating ceramic membranes and polymeric membranes in fractionation of milk proteins by microfiltration. Desalination 2009, 245, 714–722. [Google Scholar] [CrossRef]
- Karasu, K.; Glennon, N.; Lawrence, N.D.; Stevens, G.W.; O’Connor, A.J.; Barber, A.R.; Yoshikawa, S.; Kentish, S.E. A comparison between ceramic and polymeric membrane systems for casein concentrate manufacture. Int. J. Dairy Technol. 2010, 63, 284–289. [Google Scholar] [CrossRef]
- Zulewska, J.; Newbold, M.; Barbano, D.M. Efficiency of serum protein removal from skim milk with ceramic and polymeric membranes at 50 °C. J. Dairy Sci. 2009, 92, 1361–1377. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.; DiMarzo, L.; Pranata, J.; Barbano, D.M.; Drake, M. Efficiency of removal of whey protein from sweet whey using polymeric microfiltration membranes. J. Dairy Sci. 2021, 104, 8630–8643. [Google Scholar] [CrossRef] [PubMed]
- Baruah, G.L.; Nayak, A.; Belfort, G. Scale-up from laboratory microfiltration to a ceramic pilot plant: Design and performance. J. Memb. Sci. 2006, 274, 56–63. [Google Scholar] [CrossRef]
- Liu, F.; Hashim, N.A.; Liu, Y.; Abed, M.R.M.; Li, K. Progress in the production and modification of PVDF membranes. J. Memb. Sci. 2011, 375, 1–27. [Google Scholar] [CrossRef]
- Carter, B.G.; Cheng, N.; Kapoor, R.; Meletharayil, G.H.; Drake, M.A. Invited review: Microfiltration-derived casein and whey proteins from milk. J. Dairy Sci. 2021, 104, 2465–2479. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Ramchandran, L.; Vasiljevic, T. Lower ultrafiltration temperature improves membrane performance and emulsifying properties of milk protein concentrates. Dairy Sci. Technol. 2015, 95, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Steinhauer, T.; Hanély, S.; Bogendörfer, K.; Kulozik, U. Temperature dependent membrane fouling during filtration of whey and whey proteins. J. Memb. Sci. 2015, 492, 364–370. [Google Scholar] [CrossRef]
- McCarthy, N.A.; Wijayanti, H.B.; Crowley, S.V.; O’Mahony, J.A.; Fenelon, M.A. Pilot-scale ceramic membrane filtration of skim milk for the production of a protein base ingredient for use in infant milk formula. Int. Dairy J. 2017, 73, 57–62. [Google Scholar] [CrossRef]
- O’Mahony, J.A.; Smith, K.E.; Lucey, J.A. Purification of Beta Casein from Milk. U.S. Patent US8889208 B2, 18 November 2014. [Google Scholar]
- O’Mahony, J.A.; Tuohy, J.J. Further applications of membrane filtration in dairy processing. In Membrane Processing. Dairy and Beverage Applications, 1st ed.; Tamime, A.Y., Ed.; Wiley-Blackwell: West Sussex, UK, 2013; pp. 225–262. ISBN 9781118457023. [Google Scholar]
- Hurt, E.E.; Adams, M.C.; Barbano, D.M. Microfiltration of skim milk and modified skim milk using a 0.1-μm ceramic uniform transmembrane pressure system at temperatures of 50, 55, 60, and 65 °C. J. Dairy Sci. 2015, 98, 765–780. [Google Scholar] [CrossRef]
- Crowley, S.V.; Caldeo, V.; McCarthy, N.A.; Fenelon, M.A.; Kelly, A.L.; O’Mahony, J.A. Processing and protein-fractionation characteristics of different polymeric membranes during filtration of skim milk at refrigeration temperatures. Int. Dairy J. 2015, 48, 23–30. [Google Scholar] [CrossRef]
- Tan, T.J.; Wang, D.; Moraru, C.I. A physicochemical investigation of membrane fouling in cold microfiltration of skim milk. J. Dairy Sci. 2014, 97, 4759–4771. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, M.; Heidebrecht, H.-J.; Schiffer, S.; Dumpler, J.; Kulozik, U. Milk protein fractionation by means of spiral-wound microfiltration membranes: Effect of the pressure adjustment mode and temperature on flux and protein permeation. Foods 2019, 8, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, K.S.Y.; Dunstan, D.E.; Martin, G.J.O. Influence of processing temperature on flux decline during skim milk ultrafiltration. Sep. Purif. Technol. 2018, 195, 322–331. [Google Scholar] [CrossRef]
- Barukčić, I.; Božanić, R.; Kulozik, U. Effect of pore size and process temperature on flux, microbial reduction and fouling mechanisms during sweet whey cross-flow microfiltration by ceramic membranes. Int. Dairy J. 2014, 39, 8–15. [Google Scholar] [CrossRef]
- Schiffer, S.; Kulozik, U. Effect of temperature-dependent bacterial growth during milk protein fractionation by means of 0.1 µm microfiltration on the length of possible production cycle times. Membranes 2020, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- de Wit, J.N. Thermal behaviour of bovine β-lactoglobulin at temperatures up to 150 °C. A review. Trends Food Sci. Technol. 2009, 20, 27–34. [Google Scholar] [CrossRef]
- Beckman, S.L.; Barbano, D.M. Effect of microfiltration concentration factor on serum protein removal from skim milk using spiral-wound polymeric membranes. J. Dairy Sci. 2013, 96, 6199–6212. [Google Scholar] [CrossRef]
- Koh, L.L.A.; Ashokkumar, M. Kentish, S.E. Membrane fouling, cleaning and disinfection. In Membrane Processing. Dairy and Beverage Applications, 1st ed.; Tamime, A.Y., Ed.; Wiley-Blackwell: West Sussex, UK, 2013; pp. 208–224. ISBN 9781118457023. [Google Scholar]
- James, B.J.; Jing, Y.; Chen, X.D. Membrane fouling during filtration of milk—A microstructural study. J. Food Eng. 2003, 60, 431–437. [Google Scholar] [CrossRef]
- France, T.C.; Bot, F.; Kelly, A.L.; Crowley, S.V.; O’Mahony, J.A. The influence of temperature on filtration performance and fouling during cold microfiltration of skim milk. Sep. Purif. Technol. 2021, 262, 118256. [Google Scholar] [CrossRef]
- Schmidt, V.S.J.; Kaufmann, V.; Kulozik, U.; Scherer, S.; Wenning, M. Microbial biodiversity, quality and shelf life of microfiltered and pasteurized extended shelf life (ESL) milk from Germany, Austria and Switzerland. Int. J. Food Microbiol. 2012, 154, 1–9. [Google Scholar] [CrossRef]
- Li, F.; Hunt, K.; Van Hoorde, K.; Butler, F.; Jordan, K.; Tobin, J.T. Occurrence and identification of spore-forming bacteria in skim-milk powders. Int. Dairy J. 2019, 97, 176–184. [Google Scholar] [CrossRef]
- Burgess, S.A.; Lindsay, D.; Flint, S.H. Thermophilic bacilli and their importance in dairy processing. Int. J. Food Microbiol. 2010, 144, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Seale, B.; Bremer, P.; Flint, S.; Brooks, J.; Palmer, J. Overview of the problems resulting from biofilm contamination in the dairy industry. In Biofilms in the Dairy Industry; Teh, K.H., Flint, S., Brooks, J., Knight, G., Eds.; John Wiley & Sons: West Sussex, UK, 2015; pp. 49–64. ISBN 9781118876213. [Google Scholar]
- Seale, B.; Burgess, S.; Flint, S.; Brooks, J.; Bremer, P.; Parkar, S. Thermophilic spore-forming bacilli in the dairy industry. In Biofilms in the Dairy Industry; Teh, K.H., Flint, S., Brooks, J., Knight, G., Eds.; John Wiley & Sons: West Sussex, UK, 2015; pp. 112–137. ISBN 9781118876213. [Google Scholar]
- Chamberland, J.; Messier, T.; Dugat-Bony, E.; Lessard, M.-H.; Labrie, S.; Doyen, A.; Pouliot, Y. Influence of feed temperature to biofouling of ultrafiltration membrane during skim milk processing. Int. Dairy J. 2019, 93, 99–105. [Google Scholar] [CrossRef]
- Samaržija, D.; Zamberlin, Š.; Pogačić, T. Psychrotrophic bacteria and milk and dairy products quality. Mljekarstvo 2012, 62, 77–95. [Google Scholar] [CrossRef]
- Jarto, I.; Lucey, J.A.; Smith, K.E. Impact of processing temperature on production of milk protein permeate during microfiltration of skim or whole milk. Int. J. Dairy Technol. 2018, 71, 844–848. [Google Scholar] [CrossRef]
- Beckman, S.L.; Zulewska, J.; Newbold, M.; Barbano, D.M. Production efficiency of micellar casein concentrate using polymeric spiral-wound microfiltration membranes. J. Dairy Sci. 2010, 93, 4506–4517. [Google Scholar] [CrossRef] [Green Version]
- Fox, P.F.; Uniacke-Lowe, T.; McSweeney, P.L.H.; O’Mahony, J.A. Salts of Milk. In Dairy Chemistry and Biochemistry, 2nd ed.; Springer: Cham, Switzerland, 2015; pp. 241–270. ISBN 9783319148922. [Google Scholar]
- France, T.C.; Kelly, A.L.; Crowley, S.V.; O’Mahony, J.A. The effects of temperature and transmembrane pressure on protein, calcium and plasmin partitioning during microfiltration of skim milk. Int. Dairy J. 2021, 114, 104930. [Google Scholar] [CrossRef]
- Lin, M.J.; Grandison, A.S.; Lewis, M.J. Partitioning of calcium and magnesium (total divalent cations) during membrane filtration of milk. J. Food Eng. 2015, 149, 153–158. [Google Scholar] [CrossRef]
- Zulewska, J.; Kowalik, J.; Lobacz, A.; Dec, B. Short communication: Calcium partitioning during microfiltration of milk and its influence on rennet coagulation time. J. Dairy Sci. 2018, 101, 10860–10865. [Google Scholar] [CrossRef]
- Crowley, S.V.; Burlot, E.; Silva, J.V.C.; McCarthy, N.A.; Wijayanti, H.B.; Fenelon, M.A.; O’Mahony, J.A. Rehydration behaviour of spray-dried micellar casein concentrates produced using microfiltration of skim milk at cold or warm temperatures. Int. Dairy J. 2018, 81, 72–79. [Google Scholar] [CrossRef]
- Dauphas, S.; Mouhous-Riou, N.; Metro, B.; MacKie, A.R.; Wilde, P.J.; Anton, M.; Riaublanc, A. The supramolecular organisation of β-casein: Effect on interfacial properties. Food Hydrocoll. 2005, 19, 387–393. [Google Scholar] [CrossRef]
- Crowley, S.V.; Kelly, A.L.; O’Mahony, J.A.; Lucey, J.A. Colloidal properties of protein complexes formed in β-casein concentrate solutions as influenced by heating and cooling in the presence of different solutes. Colloids Surf. B Biointerfaces 2019, 174, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Auty, M.A.E.; Crowley, S.V.; Kelly, A.L.; O’Mahony, J.A.; Brodkorb, A. Self-association of bovine β-casein as influenced by calcium chloride, buffer type and temperature. Food Hydrocoll. 2019, 88, 190–198. [Google Scholar] [CrossRef]
- Huppertz, T. Chemistry of the caseins. In Advanced Dairy Chemistry. Vol. 1A Proteins: Basic Aspects, 4th ed.; Fox, P.F., McSweeney, P.L.H., Eds.; Springer: Boston, MA, USA, 2013; pp. 135–160. ISBN 978-1-4614-4714-6. [Google Scholar]
- Korycha-Dahl, M.; Dumas, B.R.; Chene, N.; Martal, J. Plasmin Activity in Milk. J. Dairy Sci. 1983, 66, 704–711. [Google Scholar] [CrossRef]
- Richardson, B. Proteinases of bovine milk and the effect of pasteurization on their activity. N. Z. J. Dairy Sci. 1983, 18, 233–245. [Google Scholar]
- Aaltonen, T.; Ollikainen, P. Effect of microfiltration of milk on plasmin activity. Int. Dairy J. 2011, 21, 193–197. [Google Scholar] [CrossRef]
- Levin, M.A.; Burrington, K.J.; Hartel, R.W. Whey protein phospholipid concentrate and delactosed permeate: Applications in caramel, ice cream, and cake. J. Dairy Sci. 2016, 99, 6948–6960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maubois, J.L. Membrane microfiltration; a tool for a new approach in dairy technology. Aust. J. Dairy Technol. 2002, 57, 92–96. [Google Scholar]
- Lindquist, A. A Method for the Production of Sterile Skimmed Milk. Patent PCT WO No. 57549, 23 December 1998. [Google Scholar]
- Fritsch, J.; Moraru, C.I. Development and optimization of a carbon dioxide-aided cold microfiltration process for the physical removal of microorganisms and somatic cells from skim milk. J. Dairy Sci. 2008, 91, 3744–3760. [Google Scholar] [CrossRef] [PubMed]
- Griep, E.R.; Cheng, Y.; Moraru, C.I. Efficient removal of spores from skim milk using cold microfiltration: Spore size and surface property considerations. J. Dairy Sci. 2018, 101, 9703–9713. [Google Scholar] [CrossRef] [Green Version]
- Hansen, S.F.; Hogan, S.A.; Tobin, J.; Rasmussen, J.T.; Larsen, L.B.; Wiking, L. Microfiltration of raw milk for production of high-purity milk fat globule membrane material. J. Food Eng. 2020, 276, 109887. [Google Scholar] [CrossRef]
- Heino, A.T.; Uusi-Rauva, J.O.; Rantamäki, P.R.; Tossavainen, O. Functional properties of native and cheese whey protein concentrate powders. Int. J. Dairy Technol. 2007, 60, 277–285. [Google Scholar] [CrossRef]
- Evans, J.; Zulewska, J.; Newbold, M.; Drake, M.A.; Barbano, D.M. Comparison of composition, sensory, and volatile components of thirty-four percent whey protein and milk serum protein concentrates. J. Dairy Sci. 2009, 92, 4773–4791. [Google Scholar] [CrossRef]
- Jäger, R.; Kerksick, C.M.; Campbell, B.I.; Cribb, P.J.; Wells, S.D.; Skwiat, T.M.; Purpura, M.; Ziegenfuss, T.N.; Ferrando, A.A.; Arent, S.M.; et al. International society of sports nutrition position stand: Protein and exercise. J. Int. Soc. Sports Nutr. 2017, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Soodam, K.; Guinee, T.P. The case for milk protein standardisation using membrane filtration for improving cheese consistency and quality. Int. J. Dairy Technol. 2018, 71, 277–291. [Google Scholar] [CrossRef]
- De Kruif, C.G.; Holt, C. Casein micelle structure, functions and interactions. In Advanced Dairy Chemistry-1 Proteins, 3rd ed.; Fox, P.F., McSweeney, P.L.H., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003; pp. 233–276. ISBN 978-1-4419-8602-3. [Google Scholar]
- Creamer, L.K.; Berry, G.P.; Mills, O.E. A study of the dissociation of β-casein from the bovine casein micelle at low temperature. N. Z. J. Dairy Sci. Technol. 1977, 12, 58–66. [Google Scholar]
- Payens, T.A.J.; Van Markwijk, B.W. Some features of the association of β-casein. BBA–Biochim. Biophys. Acta 1963, 71, 517–530. [Google Scholar] [CrossRef]
- Andrews, A.L.; Atkinson, D.; Evans, M.T.A.; Finer, E.G.; Green, J.P.; Phillips, M.C.; Robertson, R.N. The conformation and aggregation of bovine β-casein A. I. Molecular aspects of thermal aggregation. Biopolymers 1979, 18, 1105–1121. [Google Scholar] [CrossRef]
- Farrell, H.M., Jr.; Wickham, E.D.; Unruh, J.J.; Qi, P.X.; Hoagland, P.D. Secondary structural studies of bovine caseins: Temperature dependence of β-casein structure as analyzed by circular dichroism and FTIR spectroscopy and correlation with micellization. Food Hydrocoll. 2001, 15, 341–354. [Google Scholar] [CrossRef]
- Schiffer, S.; Vannieuwenhuyse, L.; Susianto, C.; Hartinger, M.; Kulozik, U. Influence of pH and calcium concentration on milk protein fractionation by 0.1 μm microfiltration at low temperatures. Int. Dairy J. 2021, 118, 105048. [Google Scholar] [CrossRef]
- Rose, D. Relation between Micellar and Serum Casein in Bovine Milk. J. Dairy Sci. 1968, 51, 1897–1902. [Google Scholar] [CrossRef]
- Philippe, M.; Gaucheron, F.; Le Graet, Y.; Michel, F.; Garem, A. Physicochemical characterization of calcium-supplemented skim milk. Lait 2003, 83, 45–59. [Google Scholar] [CrossRef]
- Le Berre, O.; Daufin, G. Fouling and selectivity of membranes during separation of β-casein. J. Memb. Sci. 1994, 88, 263–270. [Google Scholar] [CrossRef]
- Schäfer, J.; Schubert, T.; Atamer, Z. Pilot-scale β-casein depletion from micellar casein via cold microfiltration in the diafiltration mode. Int. Dairy J. 2019, 97, 222–229. [Google Scholar] [CrossRef]
- van der Schaaf, J.M.; Crowley, S.V.; Kelly, A.L.; O’Mahony, J.A. Manufacture of Milk and Whey Products: Fractionated Casein Ingredients—β-Casein. In Reference Module in Food Science; Elsevier, 2021. [Google Scholar] [CrossRef]
- Atamer, Z.; Post, A.E.; Schubert, T.; Holder, A.; Boom, R.M.; Hinrichs, J. Bovine β-casein: Isolation, properties and functionality. A review. Int. Dairy J. 2017, 66, 115–125. [Google Scholar] [CrossRef]
- Chen, C.W. Guidance on the conceptual design of sustainable product-service systems. Sustainability 2018, 10, 2452. [Google Scholar] [CrossRef] [Green Version]
- Kamali, M.; Suhas, D.P.; Costa, M.E.; Capela, I.; Aminabhavi, T.M. Sustainability considerations in membrane-based technologies for industrial effluents treatment. Chem. Eng. J. 2019, 368, 474–494. [Google Scholar] [CrossRef]
- Anand, S.; Singh, D. Resistance of the constitutive microflora of biofilms formed on whey reverse-osmosis membranes to individual cleaning steps of a typical clean-in-place protocol. J. Dairy Sci. 2013, 96, 6213–6222. [Google Scholar] [CrossRef] [Green Version]
- Méthot-Hains, S.; Benoit, S.; Bouchard, C.; Doyen, A.; Bazinet, L.; Pouliot, Y. Effect of transmembrane pressure control on energy efficiency during skim milk concentration by ultrafiltration at 10 and 50 °C. J. Dairy Sci. 2016, 99, 8655–8664. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic representation of the difference in structure of a symmetric (porous) and asymmetric membrane (modified from Smith [7], copyright 2013 Wiley-Blackwell).
Figure 1.
Schematic representation of the difference in structure of a symmetric (porous) and asymmetric membrane (modified from Smith [7], copyright 2013 Wiley-Blackwell).

Figure 2.
Schematic representation of permeate flux decline during warm (50 °C: ■) and cold (4 °C: □) microfiltration of skim milk (left; modified from [19], copyright 2017 International Dairy Journal) and differences in compactness of the deposit layer on the membrane surface after MF at the respective temperature (right).
Figure 2.
Schematic representation of permeate flux decline during warm (50 °C: ■) and cold (4 °C: □) microfiltration of skim milk (left; modified from [19], copyright 2017 International Dairy Journal) and differences in compactness of the deposit layer on the membrane surface after MF at the respective temperature (right).

Figure 3.
Schematic representation of some of the microbial considerations when selecting microfiltration processing temperature and the implications of such processing temperatures for processing efficiency and finished product quality. Information regarding examples of bacteria species sourced from [36,37,38,39].
Figure 3.
Schematic representation of some of the microbial considerations when selecting microfiltration processing temperature and the implications of such processing temperatures for processing efficiency and finished product quality. Information regarding examples of bacteria species sourced from [36,37,38,39].

Figure 4.
Schematic representation of the use of microfiltration (MF) to improve microbiological quality and generation of different milk protein-derived ingredients as influenced by processing temperature. MCC, micellar casein concentrate; MCI, micellar casein isolate; SMP, skim milk powder; WPC, whey protein concentrate; WPI, whey protein isolate; WPPC, whey protein phospholipid concentrate; P, permeate; R, retentate.
Figure 4.
Schematic representation of the use of microfiltration (MF) to improve microbiological quality and generation of different milk protein-derived ingredients as influenced by processing temperature. MCC, micellar casein concentrate; MCI, micellar casein isolate; SMP, skim milk powder; WPC, whey protein concentrate; WPI, whey protein isolate; WPPC, whey protein phospholipid concentrate; P, permeate; R, retentate.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
France, T.C.; Kelly, A.L.; Crowley, S.V.; O’Mahony, J.A. Cold Microfiltration as an Enabler of Sustainable Dairy Protein Ingredient Innovation. Foods 2021, 10, 2091. https://doi.org/10.3390/foods10092091
AMA Style
France TC, Kelly AL, Crowley SV, O’Mahony JA. Cold Microfiltration as an Enabler of Sustainable Dairy Protein Ingredient Innovation. Foods. 2021; 10(9):2091. https://doi.org/10.3390/foods10092091
Chicago/Turabian StyleFrance, Thomas C., Alan L. Kelly, Shane V. Crowley, and James A. O’Mahony. 2021. "Cold Microfiltration as an Enabler of Sustainable Dairy Protein Ingredient Innovation" Foods 10, no. 9: 2091. https://doi.org/10.3390/foods10092091
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.