
Amino Acid Sequences of Lactoferrin from Red Deer (Cervus elaphus) Milk and Antimicrobial Activity of Its Derived Peptides Lactoferricin and Lactoferrampin



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
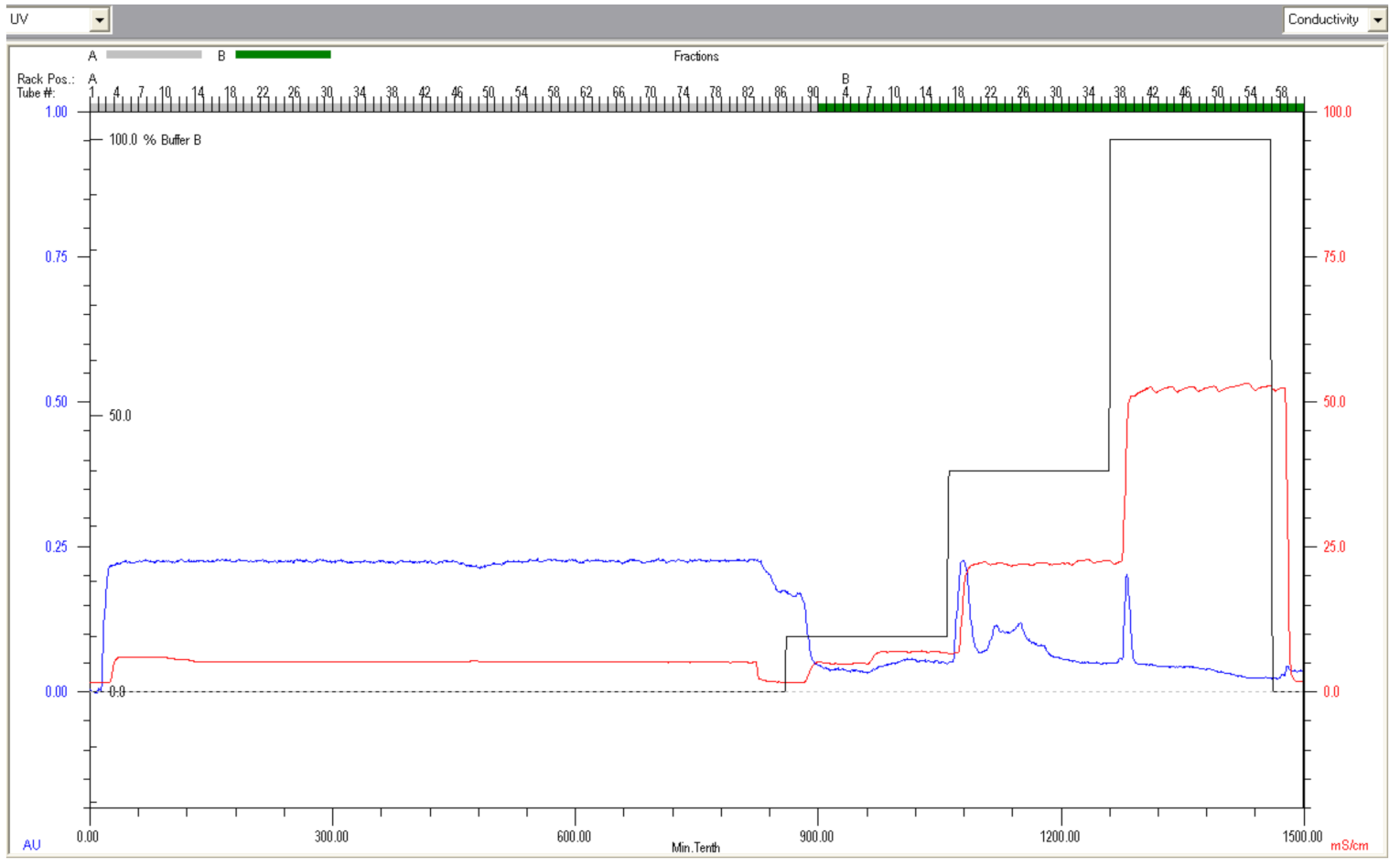
2.2. Purification of Deer Lactoferrin by Fast Protein Liquid Chromatography
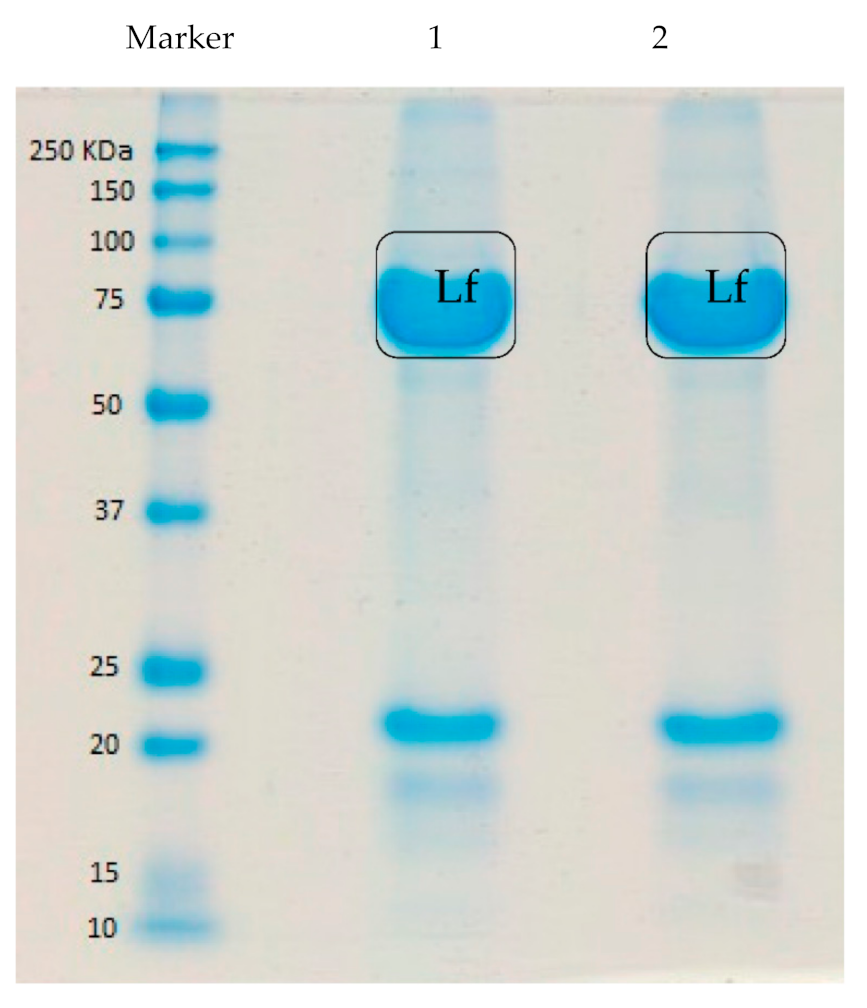
2.3. SDS-PAGE
2.4. Deer Lf Amino Acid Sequence Analysis
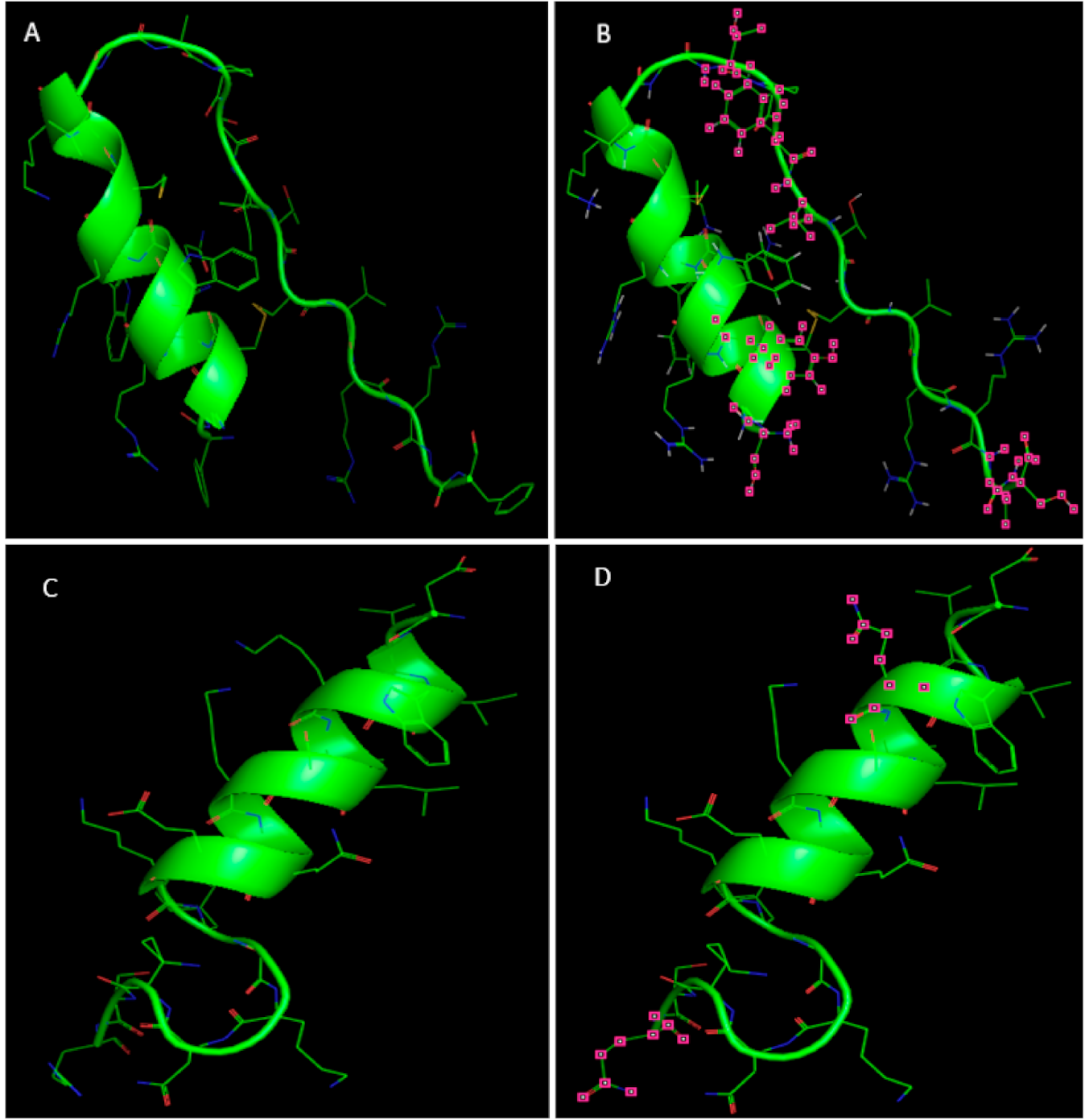
2.5. Three-Dimensional Structures of Deer Lfcin and Lfampin In Silico Analysis
2.6. Antibacterial Activity Assay for Deer Lfcin and Lfampin
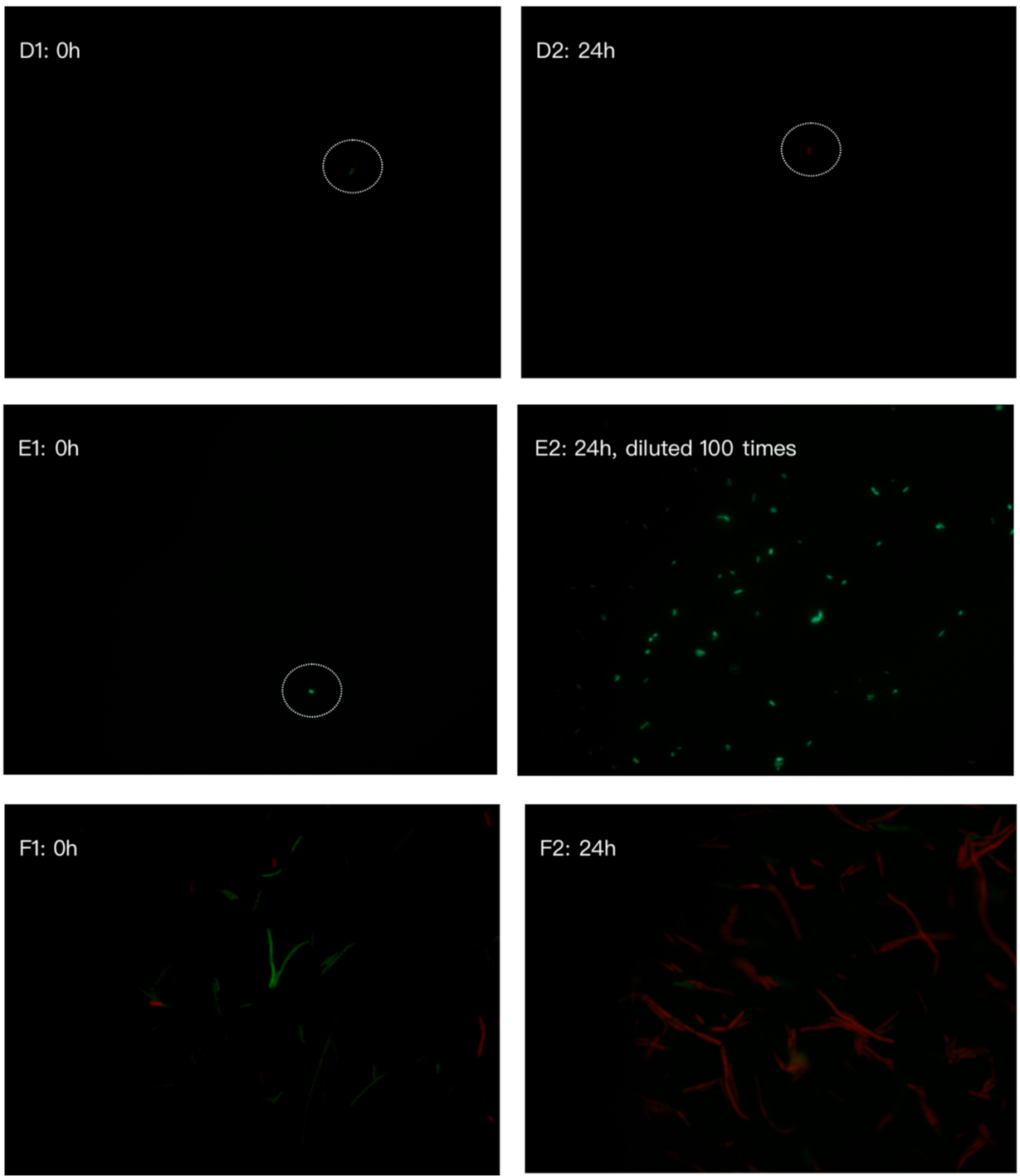
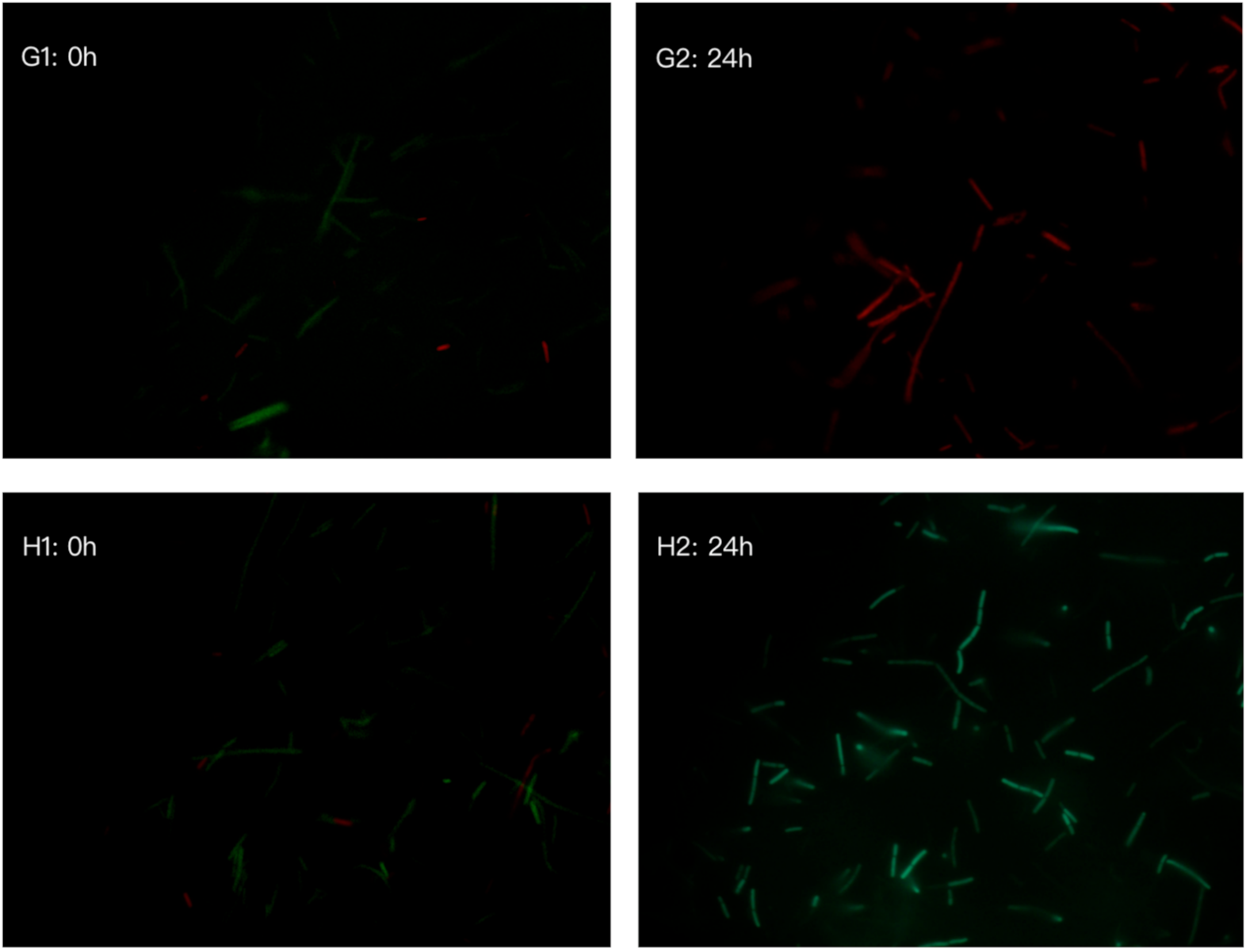
2.7. Bacterial Cell Viability by LIVE/DEAD BacLightTM Bacterial Viability Kit
2.8. Statistical Analysis
3. Results
3.1. Chromatography Fractionation and Electrophoresis of Lf from Deer Milk
3.2. Amino Acid Sequence of Deer Lactoferrin
Characteristics of Deer Lactoferricin and Lactoferrampin
3.3. Antibacterial Activities of Synthesized Peptides Lactoferricin and Lactoferrampin on Bacterial Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giacometti, A.; Cirioni, O.; Barchiesi, F.; Del Prete, M.S.; Scalise, G. Antimicrobial activity of polycationic peptides. Peptides 1999, 20, 1265–1273. [Google Scholar] [CrossRef]
- Bechinger, B.; Gorr, S.U. Antimicrobial Peptides: Mechanisms of Action and Resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, S.D.; Beverly, R.L.; Qu, Y.; Dallas, D.C. Milk bioactive peptide database: A comprehensive database of milk protein-derived bioactive peptides and novel visualization. Food Chem. 2017, 232, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bekhit, A.E.-D.A.; Mason, S.L.; Morton, J.D. Nutritional value of deer milk. In Nutrients in Dairy and Their Implications on Health and Disease; Watson, R.R., Collier, R.J., Preedy, V.R., Eds.; Elsevier BV: Amsterdam, The Netherlands, 2017; pp. 363–375. [Google Scholar]
- Wang, Y.; Bekhit, A.E.-D.A.; Mason, S.L.; Morton, J.D. Lactoferrin isolation and hydrolysis from red deer (Cervus elaphus) milk and the antibacterial activity of deer lactoferrin and it hydrolysates. Foods 2020, 9, 1711. [Google Scholar] [CrossRef] [PubMed]
- Franco, I.; Perez, M.D.; Conesa, C.; Calvo, M.; Sanchez, L. Effect of technological treatments on bovine lactoferrin: An overview. Food Res. Intern. 2018, 106, 173–182. [Google Scholar] [CrossRef]
- Goodman, R.E.; Schanbacher, F.L. Bovine lactoferrin mRNA: Sequence, analysis, and expression in the mammary gland. Biochem. Biophys. Res. Commun. 1991, 180, 75–84. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F.; Prignot, J.J.; Wauters, G. Immunohistochemical localization and bacteriostatic properties of an iron-binding protein from bronchial mucus. Thorax 1966, 21, 538–544. [Google Scholar] [CrossRef] [Green Version]
- Barbiroli, A.; Bonomi, F.; Capretti, G.; Iametti, S.; Manzoni, M.; Piergiovanni, L.; Rollini, M. Antimicrobial activity of lysozyme and lactoferrin incorporated in cellulose-based food packaging. Food Control 2012, 26, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence against microbial and viral attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef] [PubMed]
- Bruni, N.; Capucchio, M.T.; Biasibetti, E.; Pessione, E.; Cirrincione, S.; Giraudo, L.; Corona, A.; Dosio, F. Antimicrobial activity of lactoferrin-related peptides and applications in human and veterinary medicine. Molecules 2016, 21, 752. [Google Scholar] [CrossRef]
- Protein Data Bank. Available online: https://www.rcsb.org (accessed on 26 April 2021).
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identifification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- van der Kraan, M.I.A.; Groenink, J.; Nazmi, K.; Veerman, E.C.I.; Bolscher, J.G.M.; Amerongen, A.V.N. Lactoferrampin: A novel antimicrobial peptide in the N1-domain of bovine lactoferrin. Peptides 2004, 25, 177–183. [Google Scholar] [CrossRef]
- Pilbrow, J.; Bekhit, A.E.-D.A.; Carne, A. Fractionation of sheep cheese whey by a scalable method to sequentially isolate bioactive proteins. Food Chem. 2016, 203, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Jensen, O.N.; Podtelejnikov, A.V.; Sagliocco, F.; Wilm, M.; Vorm, O.; Mortensen, P.; Shevchenko, A.; Boucherie, H.; Mann, M. Linking genome and proteome by mass spectrometry: Large-scale identification of yeast proteins from two dimensional gels. Proc. Natl. Acad. Sci. USA 1996, 93, 14440–14445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Sabherwal, M.; Duncan, E.; Stevens, S.; Stockwell, P.; McConnell, M.; Bekhit, A.E.D.; Carne, A. In-depth characterization of sheep (Ovis aries) milk whey proteome and comparison with cow (Bos taurus). PLoS ONE 2015, 10, e0139774. [Google Scholar] [CrossRef] [Green Version]
- Mascot Search Engine. Available online: https://www.matrixscience.com (accessed on 26 April 2021).
- Hasselmann, C.; European Soc Clinical, M. Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, ix–xv. [Google Scholar]
- Conesa, C.; Sanchez, L.; Rota, C.; Perez, M.D.; Calvo, M.; Farnaud, S.; Evans, R.W. Isolation of lactoferrin from milk of different species: Calorimetric and antimicrobial studies. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 131–139. [Google Scholar] [CrossRef]
- Etzel, M.R. Manufacture and use of dairy protein fractions. J. Nutr. 2004, 134, 996S–1002S. [Google Scholar] [CrossRef]
- Strom, M.B.; Haug, B.E.; Rekdal, O.; Skar, M.L.; Stensen, W.; Svendsen, J.S. Important structural features of 15-residue lactoferricin derivatives and methods for improvement of antimicrobial activity. Biochem. Cell Biol. 2002, 80, 65–74. [Google Scholar] [CrossRef]
- Strom, M.B.; Rekdal, O.; Svendsen, J.S. The effects of charge and lipophilicity on the antibacterial activity of undecapeptides derived from bovine lactoferricin. J. Pept. Sci. 2002, 8, 36–43. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Wakabayashi, H.; Kawase, K.; Tomita, M. Antibacterial spectrum of lactoferricin B, a potent bactericidal peptide derived from the N-terminal region of bovine lactoferrin. J. Appl. Bact. 1992, 73, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Ulvatne, H.; Haukland, H.H.; Olsvik, O.; Vorland, L.H. Lactoferricin B causes depolarization of the cytoplasmic membrane of Escherichia coli ATCC 25922 and fusion of negatively charged liposomes. FEBS Lett. 2001, 492, 62. [Google Scholar] [CrossRef] [Green Version]
- Epand, R.M.; Vogel, H.J. Diversity of antimicrobial peptides and their mechanisms of action. Biochim. Biophys. Acta-Biomembr. 1999, 1462, 11–28. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.H.; Lee, M.K.; Kim, K.L.; Hahm, K.S. Structure-biological activity relationships of 11-residue highly basic peptide segment of bovine lactoferrin. Int. J. Pept. Protein Res. 1996, 48, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Lau, F.; Vogel, H.J. Solution structures and model membrane interactions of lactoferrampin, an antimicrobial peptide derived from bovine lactoferrin. Biochim. Biophys. Acta-Biomembr. 2007, 1768, 2355–2364. [Google Scholar] [CrossRef] [Green Version]
- Haney, E.F.; Nazmi, K.; Bolscher, J.G.M.; Vogel, H.J. Influence of specific amino acid side-chains on the antimicrobial activity and structure of bovine lactoferrampin. Biochem. Cell Biol. 2012, 90, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Vorland, L.H.; Ulvatne, H.; Andersen, J.; Haukland, H.H.; Rekdal, O.; Svendsen, J.S.; Gutteberg, T.J. Lactoferricin of bovine origin is more active than lactoferricins of human, murine and caprine origin. Scand. J. Infect. Dis. 1998, 30, 513–517. [Google Scholar]
- Chapple, D.S.; Mason, D.J.; Joannou, C.L.; Odell, E.W.; Gant, V.; Evans, R.W. Structure-function relationship of antibacterial synthetic peptides homologous to a helical surface region on human lactoferrin against Escherichia coli serotype O111. Infect. Immun. 1998, 66, 2434–2440. [Google Scholar] [CrossRef] [Green Version]
- Chaparro, S.C.V.; Salguero, J.T.V.; Baquero, D.A.M.; Perez, J.E.R. Effect of polyvalence on the antibacterial activity of a synthetic peptide derived from bovine lactoferricin against healthcare-Associated infectious pathogens. Biomed Res. Int. 2018, 2018, 5252891. [Google Scholar] [CrossRef] [PubMed]
- Mendez, N.D.H.; Casanova, Y.V.; Chimbi, A.K.G.; Hernandez, E.; Castro, A.L.L.; Diaz, J.M.M.; Monroy, Z.J.R.; Castaneda, J.E.G. Synthetic peptides derived from bovine lactoferricin exhibit antimicrobial activity against E. coli ATCC 11775, S. maltophilia ATCC 13636 and S. enteritidis ATCC 13076. Molecules 2017, 22, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.W.; Jheng, T.T.; Shyu, C.L.; Mao, F.C. Antimicrobial potential for the combination of bovine lactoferrin or its hydrolysate with lactoferrin-resistant probiotics against foodborne pathogens. J. Dairy Sci. 2013, 96, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Davey, H.M. Life, death, and in-between: Meanings and methods in microbiology. Appl. Environ. Microbiol. 2011, 77, 5571–5576. [Google Scholar] [CrossRef] [Green Version]
- Berney, M.; Hammes, F.; Bosshard, F.; Weilenmann, H.U.; Egli, T. Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight kit in combination with flow cytometry. Appl. Environ. Microbiol. 2007, 73, 3283–3290. [Google Scholar] [CrossRef] [Green Version]
- Stocks, S.M. Mechanism and use of the commercially available viability stain, BacLight. Cytometry Part A 2004, 61, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Ulvatne, H.; Samuelsen, O.; Haukland, H.H.; Kramer, M.; Vorland, L.H. Lactoferricin B inhibits bacterial macromolecular synthesis in Escherichia coli and Bacillus subtilis. FEMS Microbiol. Lett. 2004, 237, 377–384. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Deer Bovine 1 1–60 | MKLFV PALLA LGALG LCLAA PRKNV RWCAI SQPEW SKCYR WQWRM KKLGT PFVTC VRRTS 3 MKLFV PALLS LGALG LCLA 2 A PRKNV RWCTI SQPEW FKCRR WQWRM KKLGA PSITC VRRAF 4 |
| Deer Bovine 61–120 | ALECI RAIAA KKADA VTLDS GLVFE AGLDP YKLRP VAAEI YGTEK SPQTH YYAVA VVKKG ALECI RAIAE KKADA VTLDG GMVFE AGRDP YKLRP VAAEI YGTKE SPQTH YYAVA VVKKG |
| Deer Bovine 121–180 | SNFQL DQLQG RKSCH TGLGR SAGWN IPIGI LRPSL GWTES LEPLQ GAVAK FFSAS CVPCV SNFQL DQLQG RKSCH TGLGR SAGWI IPMGI LRPYL SWTES LEPLQ GAVAK FFSAS CVPCI |
| Deer Bovine 181–240 | DGKAY PNLCQ LCKGT GENQC ACSPR EPY_G YSGAF RCLQE GAGDV AFVKE TTVFE NLPEK DRQAY PNLCQ LCKGE GENQC ACSSR EPYFG YSGAF KCLQD GAGDV AFVKE TTVFE NLPEK |
| Deer Bovine 241–300 | ADRDQ YELLC LNNSR APVDA FKECH LAQVP SHAVV ARSVD GKEDL IWQLL SKAQE KFGKN ADRDQ YELLC LNNSR APVDA FKECH LAQVP SHAVV ARSVD GKEDL IWKLL SKAQE KFGKN |
| Deer Bovine 301–360 | KSQ 5 SF QLFGS PGSQR DLLFK DSALG FLRIP SKIDS ELYLG ARYLT ALKNL RETEE EVKAR KSR 6 SF QLFGS PPGQR DLLFK DSALG FLRIP SKVDS ALYLG SRYLT TLKNL RETAE EVKAR |
| Deer Bovine 361–420 | STRVV WCAVG PEEQK KCQQW SQQSD QSVTC ATAST TDDCI ALVLK GEADA LSLDG GYIYT YTRVV WCAVG PEEQK KCQQW SQQSG QNVTC ATAST TDDCI VLVLK GEADA LNLDG GYIYT |
| Deer Bovine 421–480 | AGKCG LVPVM AENRK SSKDS SLDCV LRPTE GYLAV AVVKK ANEGL TWNSL KGKKS CHTAV AGKCG LVPVL AENRK SSKHS SLDCV LRPTE GYLAV AVVKK ANEGL TWNSL KDKKS CHTAV |
| Deer Bovine 481–540 | DRTAG WNIPM GLIAN QTGSC KFDEF FSQSC APGAD PKSSL CALCA GDDQG LDKCV PNTKE DRTAG WNIPM GLIVN QTGSC AFDEF FSQSC APGAD PKSRL CALCA GDDQG LDKCV PNSKE |
| Deer Bovine 541–600 | KYYGY TGAFR CLAED VGDVA FVKND TVWEN TNGES SADWA KNLNR EDFRL LCLDG TRKPV KYYGY TGAFR CLAED VGDVA FVKND TVWEN TNGES TADWA KNLNR EDFRL LCLDG TRKPV |
| Deer Bovine 601–660 | TEAQS CHLAA APSHA VVSRS DRAAH VEQVL LHQQA LFGRN GKDCP DKFCL FKSET KNLLF TEAQS CHLAV APNHA VVSRS DRAAH VKQVL LHQQA LFGKN GKNCP DKFCL FKSET KNLLF |
| Deer Bovine 661–708 | NDNTR CLAKL GGRPT YEKYL GTEYV TAIAN LKKCS TSPLL EACAF LTR NDNTE CLAKL GGRPT YEEYL GTEYV TAIAN LKKCS TSPLL EACAF LTR |
| Lfcin | Lfampin | |||
|---|---|---|---|---|
| Deer | Bovine | Deer | Bovine | |
| E. coli ATCC 25922 | 240 b | 120 a | >960 d | 480 c |
| S. aureus ATCC 25923 | >960 | >960 | >960 | >960 |
| L. acidophilus ATCC 4356 | 480 a | >960 b | >960 b | >960 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Morton, J.D.; Bekhit, A.E.-D.A.; Carne, A.; Mason, S.L. Amino Acid Sequences of Lactoferrin from Red Deer (Cervus elaphus) Milk and Antimicrobial Activity of Its Derived Peptides Lactoferricin and Lactoferrampin. Foods 2021, 10, 1305. https://doi.org/10.3390/foods10061305
Wang Y, Morton JD, Bekhit AE-DA, Carne A, Mason SL. Amino Acid Sequences of Lactoferrin from Red Deer (Cervus elaphus) Milk and Antimicrobial Activity of Its Derived Peptides Lactoferricin and Lactoferrampin. Foods. 2021; 10(6):1305. https://doi.org/10.3390/foods10061305
Chicago/Turabian StyleWang, Ye, James D. Morton, Alaa EL-Din A. Bekhit, Alan Carne, and Susan L. Mason. 2021. "Amino Acid Sequences of Lactoferrin from Red Deer (Cervus elaphus) Milk and Antimicrobial Activity of Its Derived Peptides Lactoferricin and Lactoferrampin" Foods 10, no. 6: 1305. https://doi.org/10.3390/foods10061305