
Determination of Carbohydrate Composition in Mealworm (Tenebrio molitor L.) Larvae and Characterization of Mealworm Chitin and Chitosan



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Mealworm Powder
2.2. Total Soluble Sugar and Reducing Sugar Contents
2.3. Free Sugar Composition
2.4. Total Glucosamine Content
2.5. Extraction of Chitin and Preparation of Chitosan from Mealworms
2.6. Moisture and Crude Protein Contents
2.7. Color and Whiteness Index (WI)
2.8. Field-Emission Scanning Electron Microscopy (FESEM) Imaging and Energy Dispersive X-Ray Spectroscopy (EDS) Analysis
2.9. Nuclear Magnetic Resonance (NMR)
2.10. Fourier Transform Infrared Spectroscopy (FT-IR)
2.11. X-Ray Powder Diffraction (XRD)
2.12. Anti-Inflammatory Effect in Murine Macrophage Cell Line
2.13. Statistical Analysis
3. Results and Discussion
3.1. Carbohydrate Content and Its Composition in Mealworms
3.2. Yield, Moisture and Crude Protein Contents, and Color Traits of Chitin Extracted from Mealworms
3.3. Surface Microstructures of Chitin and Chitosan Derived from Mealworms
3.4. Determination of Precise DA and DDA of Mealworm Chitin and Chitosan Using NMR
3.5. FT-IR Spectral Analyses of Mealworm Chitin and Chitosan
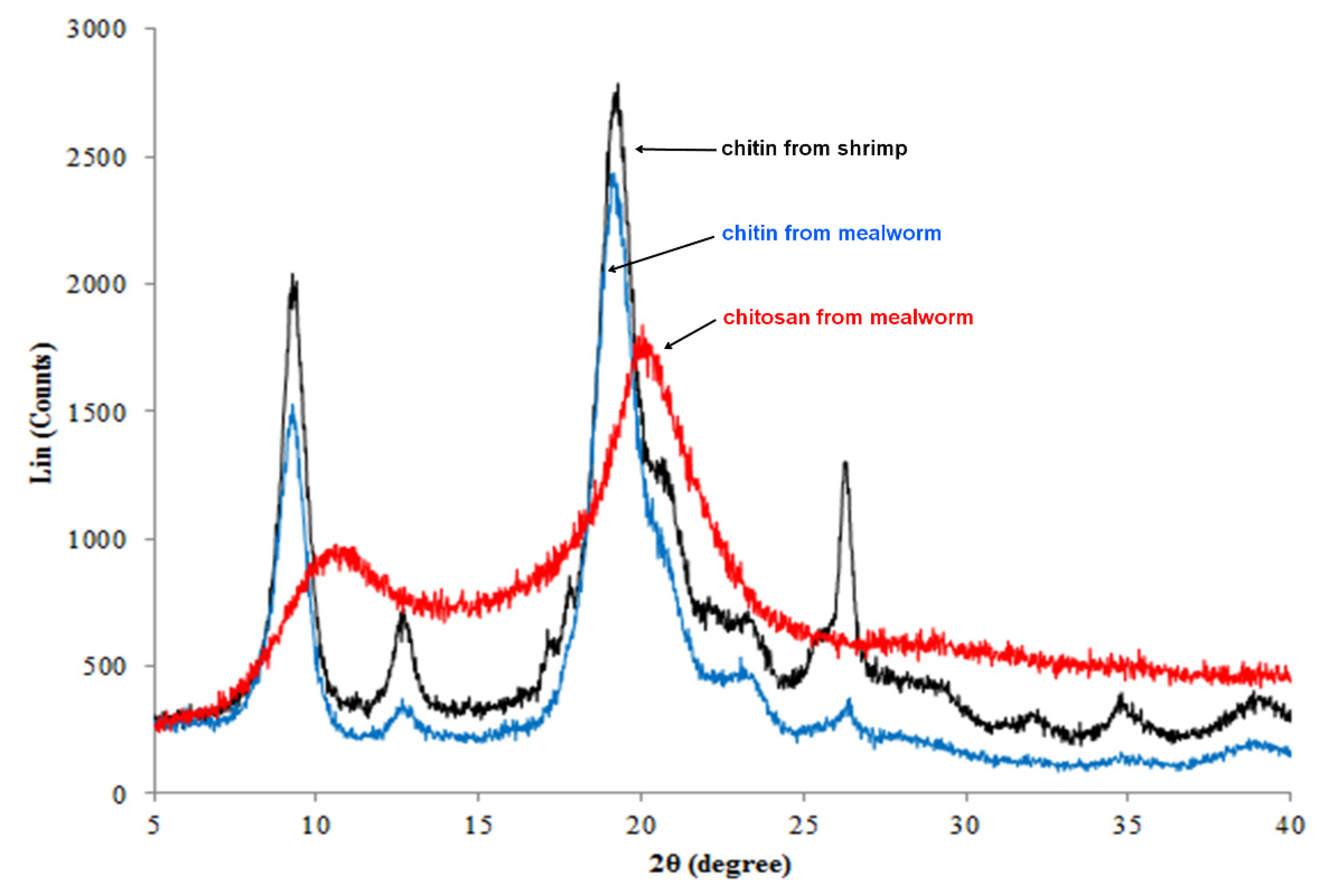
3.6. Determination of Crystalline Structures of Mealworm Chitin and Chitosan by XRD
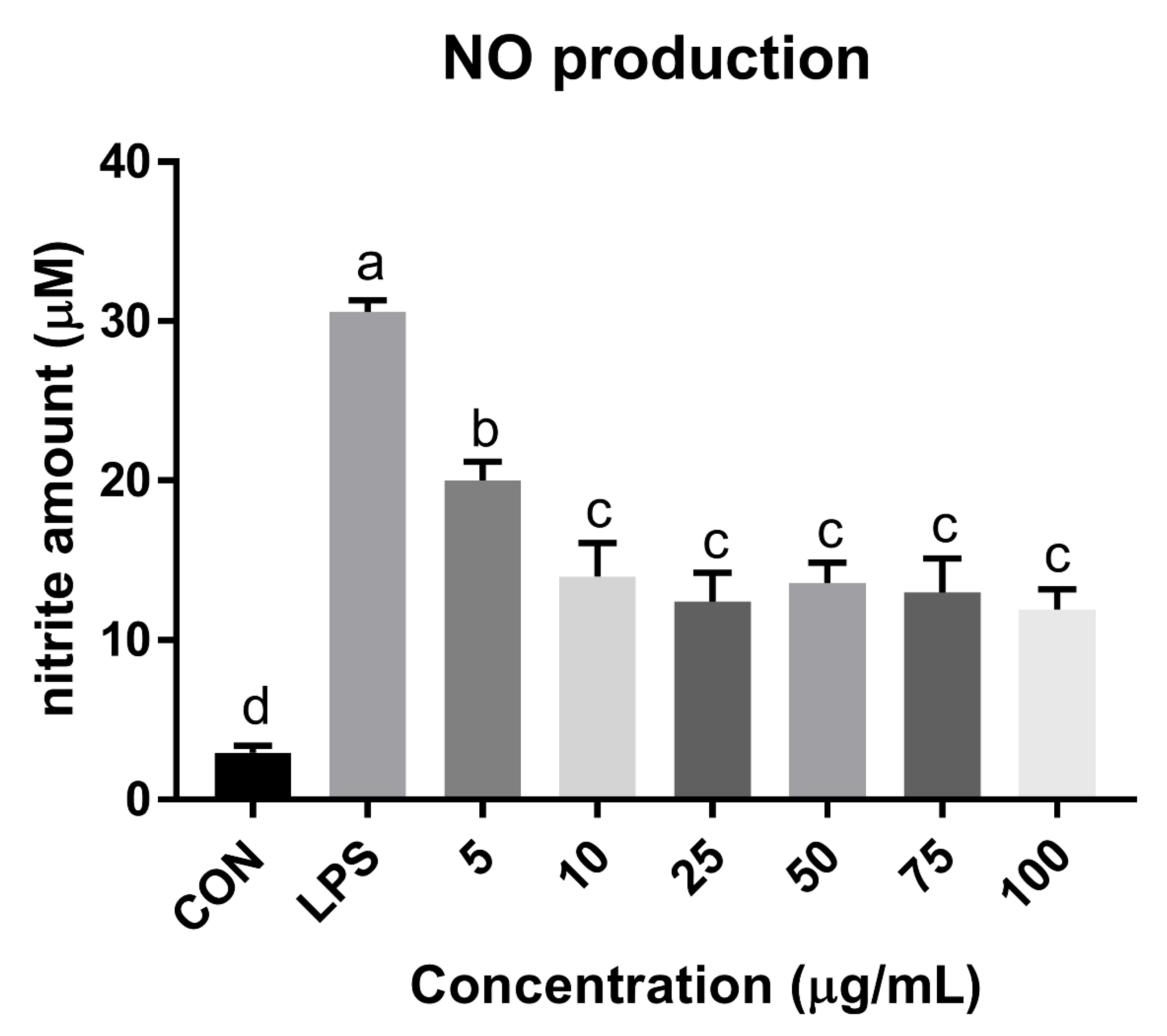
3.7. NO Reduction Activity of Mealworm Chitosan
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Boer, J.; Helms, M.; Aiking, H. Protein consumption and sustainability: Diet diversity in EU-15. Ecol. Econ. 2006, 59, 267–274. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- van Huis, A.; Oonincx, D.G. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Rochow, V.B. Can insects help to ease the problem of world food shortage. Search 1975, 6, 261–262. [Google Scholar]
- Rumpold, B.A.; Fröhling, A.; Reineke, K.; Knorr, D.; Boguslawski, S.; Ehlbeck, J.; Schlüter, O. Comparison of volumetric and surface decontamination techniques for innovative processing of mealworm larvae (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2014, 26, 232–241. [Google Scholar] [CrossRef]
- Liu, C.; Masri, J.; Perez, V.; Maya, C.; Zhao, J. Growth performance and nutrient composition of mealworms (Tenebrio molitor) fed on fresh plant materials-supplemented diets. Foods 2020, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Weaver, D.K.; McFarlane, J. The effect of larval density on growth and development of Tenebrio molitor. J. Insect Physiol. 1990, 36, 531–536. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor. Sci. Rep. 2020, 10, 1–10. [Google Scholar]
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary inclusion of Tenebrio molitor larvae meal: Effects on growth performance and final quality treats of blackspot sea bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49–58. [Google Scholar] [CrossRef]
- Bovera, F.; Loponte, R.; Marono, S.; Piccolo, G.; Parisi, G.; Iaconisi, V.; Gasco, L.; Nizza, A. Use of Tenebrio molitor larvae meal as protein source in broiler diet: Effect on growth performance, nutrient digestibility, and carcass and meat traits. J. Anim. Sci. 2016, 94, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.A.; Ding, Q.; Yin, L.; Chi, X.; Sun, N.; He, R.; Luo, L.; Ma, H.; Li, Z. Comparison of the nutritional value of mysore thorn borer (Anoplophora chinensis) and mealworm larva (Tenebrio molitor): Amino acid, fatty acid, and element profiles. Food Chem. 2020, 323, 126818. [Google Scholar] [CrossRef]
- Ghosh, S.; Lee, S.-M.; Jung, C.; Meyer-Rochow, V.B. Nutritional composition of five commercial edible insects in South Korea. J. Asia-Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- Schäufele, I.; Albores, E.B.; Hamm, U. The role of species for the acceptance of edible insects: Evidence from a consumer survey. Br. Food J. 2019, 121. [Google Scholar] [CrossRef]
- Kröncke, N.; Böschen, V.; Woyzichovski, J.; Demtröder, S.; Benning, R. Comparison of suitable drying processes for mealworms (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2018, 50, 20–25. [Google Scholar] [CrossRef]
- Son, Y.-J.; Lee, J.-C.; Hwang, I.-K.; Nho, C.W.; Kim, S.-H. Physicochemical properties of mealworm (Tenebrio molitor) powders manufactured by different industrial processes. LWT 2019, 116, 108514. [Google Scholar] [CrossRef]
- Son, Y.-J.; Choi, S.Y.; Hwang, I.-K.; Nho, C.W.; Kim, S.H. Could defatted mealworm (Tenebrio molitor) and mealworm oil be used as food ingredients? Foods 2020, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.t.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kim, K.-S.; Park, J.-B.; Kim, S.-A. Quality characteristics of kochujang prepared with Korean single-harvested pepper (Capsicum annuum L.). J. Korean Soc. Food Sci. Nutr. 2007, 36, 759–765. [Google Scholar] [CrossRef]
- Belcher, R.; Nutten, A.; Sambrook, C. The determination of glucosamine. Analyst 1954, 79, 201–208. [Google Scholar] [CrossRef]
- Elson, L.A.; Morgan, W.T.J. A colorimetric method for the determination of glucosamine and chondrosamine. Biochem. J. 1933, 27, 1824. [Google Scholar] [CrossRef] [Green Version]
- Manni, L.; Ghorbel-Bellaaj, O.; Jellouli, K.; Younes, I.; Nasri, M. Extraction and characterization of chitin, chitosan, and protein hydrolysates prepared from shrimp waste by treatment with crude protease from Bacillus cereus SV1. Appl. Biochem. Biotechnol. 2010, 162, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Al Sagheer, F.; Al-Sughayer, M.; Muslim, S.; Elsabee, M. Extraction and characterization of chitin and chitosan from marine sources in Arabian Gulf. Carbohydr. Polym. 2009, 77, 410–419. [Google Scholar] [CrossRef]
- AOAC, H.W. International A: Official Methods of Analysis of the AOAC International; The Association: Arlington County, VA, USA, 2000. [Google Scholar]
- Hsu, C.-L.; Chen, W.; Weng, Y.-M.; Tseng, C.-Y. Chemical composition, physical properties, and antioxidant activities of yam flours as affected by different drying methods. Food Chem. 2003, 83, 85–92. [Google Scholar] [CrossRef]
- Xu, J.; McCarthy, S.P.; Gross, R.A.; Kaplan, D.L. Chitosan film acylation and effects on biodegradability. Macromolecules 1996, 29, 3436–3440. [Google Scholar] [CrossRef]
- Kasaai, M.R.; Arul, J.; Charlet, G. Intrinsic viscosity–molecular weight relationship for chitosan. J. Polym. Sci. Part B: Polym. Phys. 2000, 38, 2591–2598. [Google Scholar] [CrossRef]
- Van de Velde, K.; Kiekens, P. Structure analysis and degree of substitution of chitin, chitosan and dibutyrylchitin by FT-IR spectroscopy and solid state 13C NMR. Carbohydr. Polym. 2004, 58, 409–416. [Google Scholar] [CrossRef]
- Ottøy, M.H.; Vårum, K.M.; Christensen, B.E.; Anthonsen, M.W.; Smidsrød, O. Preparative and analytical size-exclusion chromatography of chitosans. Carbohydr. Polym. 1996, 31, 253–261. [Google Scholar] [CrossRef]
- Lavertu, M.; Xia, Z.; Serreqi, A.; Berrada, M.; Rodrigues, A.; Wang, D.; Buschmann, M.; Gupta, A. A validated 1H NMR method for the determination of the degree of deacetylation of chitosan. J. Pharm. Biomed. Anal. 2003, 32, 1149–1158. [Google Scholar] [CrossRef]
- Baxter, A.; Dillon, M.; Taylor, K.A.; Roberts, G.A. Improved method for ir determination of the degree of N-acetylation of chitosan. Int. J. Biol. Macromol. 1992, 14, 166–169. [Google Scholar] [CrossRef]
- Yen, M.-T.; Yang, J.-H.; Mau, J.-L. Physicochemical characterization of chitin and chitosan from crab shells. Carbohydr. Polym. 2009, 75, 15–21. [Google Scholar] [CrossRef]
- Wan, Y.; Creber, K.A.M.; Peppley, B.; Bui, V.T. Ionic conductivity and related properties of crosslinked chitosan membranes. J. Appl. Polym. Sci. 2003, 89, 306–317. [Google Scholar] [CrossRef]
- Liao, G.Z.; Wang, G.Y.; Xu, X.L.; Zhou, G.H. Effect of cooking methods on the formation of heterocyclic aromatic amines in chicken and duck breast. Meat Sci. 2010, 85, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, S.; Albrecht, A.; Petersen, B.; Kreyenschmidt, J. Characterization and Comparison of Spoilage Processes in Fresh Pork and Poultry. J. Food Qual. 2012, 35, 372–382. [Google Scholar] [CrossRef]
- EnfÄLt, A.-C.; Hullberg, A. Glycogen, glucose and glucose-6-phosphate content in fresh and cooked meat and meat exudate from carriers and noncarriers of the RNˉ allele. J. Muscle Foods 2005, 16, 330–341. [Google Scholar] [CrossRef]
- Tarr, H. Postmortem degradation of glycogen and starch in fish muscle. J. Fish. Board Can. 1968, 25, 1539–1554. [Google Scholar] [CrossRef]
- Jakinovich Jr, W.; Sugarman, D. Sugar taste reception in mammals. Chem. Senses 1988, 13, 13–31. [Google Scholar] [CrossRef]
- Harish Prashanth, K.V.; Tharanathan, R.N. Chitin/chitosan: Modifications and their unlimited application potential—an overview. Trends Food Sci. Technol. 2007, 18, 117–131. [Google Scholar] [CrossRef]
- Song, Y.-S.; Kim, M.-W.; Moon, C.; Seo, D.-J.; Han, Y.S.; Jo, Y.H.; Noh, M.Y.; Park, Y.-K.; Kim, S.-A.; Kim, Y.W.; et al. Extraction of chitin and chitosan from larval exuvium and whole body of edible mealworm, Tenebrio molitor. Entomol. Res. 2018, 48, 227–233. [Google Scholar] [CrossRef]
- Siregar, H.C.; Suptijah, P. Mealworm (Tenebrio molitor) as Calcium, Phosphor, and Chitosan Source. Available online: https://repository.ipb.ac.id/handle/123456789/76877 (accessed on 11 January 2021).
- Jung, W.; Jo, G.; Kuk, J.; Kim, Y.; Oh, K.; Park, R. Production of chitin from red crab shell waste by successive fermentation with Lactobacillus paracasei KCTC-3074 and Serratia marcescens FS-3. Carbohydr. Polym. 2007, 68, 746–750. [Google Scholar] [CrossRef]
- Kaya, M.; Lelešius, E.; Nagrockaitė, R.; Sargin, I.; Arslan, G.; Mol, A.; Baran, T.; Can, E.; Bitim, B. Differentiations of chitin content and surface morphologies of chitins extracted from male and female grasshopper species. PLoS ONE 2015, 10, e0115531. [Google Scholar] [CrossRef]
- Wang, D.; Bai, Y.y.; Li, J.h.; Zhang, C.x. Nutritional value of the field cricket (Gryllus testaceus Walker). Insect Sci. 2004, 11, 275–283. [Google Scholar] [CrossRef]
- Paulino, A.T.; Simionato, J.I.; Garcia, J.C.; Nozaki, J. Characterization of chitosan and chitin produced from silkworm crysalides. Carbohydr. Polym. 2006, 64, 98–103. [Google Scholar] [CrossRef]
- Hong, N.; Huaixin, C.; Yanyan, Y.; Liang, T. Studies on Extraction and Preparation Technique of Silkworm Chrysalis (Bombyx mori L.) Pupa Chitin and Chitosan. J. Hubei Univ. (Nat. Sci. Ed.) 1998, 20, 94–96. [Google Scholar]
- Pesch, Y.-Y.; Riedel, D.; Behr, M. Drosophila Chitinase 2 is expressed in chitin producing organs for cuticle formation. Arthropod Struct. Dev. 2017, 46, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Heux, L.; Brugnerotto, J.; Desbrières, J.; Versali, M.F.; Rinaudo, M. Solid State NMR for Determination of Degree of Acetylation of Chitin and Chitosan. Biomacromolecules 2000, 1, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Haga, A.; Sekiguchi, H.; Hirano, S. Structure of insect chitin isolated from beetle larva cuticle and silkworm (Bombyx mori) pupa exuvia. Int. J. Biol. Macromol. 2000, 27, 99–105. [Google Scholar] [CrossRef]
- Suresh, S.; Srivastava, V.C.; Mishra, I.M. Adsorption of catechol, resorcinol, hydroquinone, and their derivatives: A review. Int. J. Energy Environ. Eng. 2012, 3, 32. [Google Scholar] [CrossRef]
- Kramer, K.J.; Kanost, M.R.; Hopkins, T.L.; Jiang, H.; Zhu, Y.C.; Xu, R.; Kerwin, J.L.; Turecek, F. Oxidative conjugation of catechols with proteins in insect skeletal systems. Tetrahedron 2001, 57, 385–392. [Google Scholar] [CrossRef]
- Foster, L.J.R.; Ho, S.; Hook, J.; Basuki, M.; Marçal, H. Chitosan as a Biomaterial: Influence of Degree of Deacetylation on Its Physiochemical, Material and Biological Properties. PLoS ONE 2015, 10, e0135153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussian, B. Chitin: Structure, Chemistry and Biology. In Targeting Chitin-containing Organisms; Yang, Q., Fukamizo, T., Eds.; Springer: Singapore, 2019; pp. 5–18. [Google Scholar] [CrossRef]
- Focher, B.; Naggi, A.; Torri, G.; Cosani, A.; Terbojevich, M. Structural differences between chitin polymorphs and their precipitates from solutions—evidence from CP-MAS 13C-NMR, FT-IR and FT-Raman spectroscopy. Carbohydr. Polym. 1992, 17, 97–102. [Google Scholar] [CrossRef]
- Lavall, R.L.; Assis, O.B.; Campana-Filho, S.P. β-Chitin from the pens of Loligo sp.: Extraction and characterization. Bioresour. Technol. 2007, 98, 2465–2472. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Blasinska, A.; Drobnik, J. Effects of nonwoven mats of Di-O-butyrylchitin and related polymers on the process of wound healing. Biomacromolecules 2008, 9, 776–782. [Google Scholar] [CrossRef]
- Szosland, L.; Krucinska, I.; Cislo, R.; Paluch, D.; Staniszewska-Kus, J.; Solski, L.; Szymonowicz, M. Synthesis of dibutyrylchitin and preparation of new textiles made from dibutyrylchitin and chitin for medical applications. Fibres Text. East. Eur. 2001, 9, 54–57. [Google Scholar]
- Badawy, R.M.; Mohamed, H.I. Chitin extration, composition of different six insect species and their comparable characteristics with that of the shrimp. J. Am. Sci. 2015, 11, 127–134. [Google Scholar]
- Sajomsang, W.; Gonil, P. Preparation and characterization of α-chitin from cicada sloughs. Mater. Sci. Eng. C 2010, 30, 357–363. [Google Scholar] [CrossRef]
- Muzzarelli, C.; Francescangeli, O.; Tosi, G.; Muzzarelli, R.A.A. Susceptibility of dibutyryl chitin and regenerated chitin fibres to deacylation and depolymerization by lipases. Carbohydr. Polym. 2004, 56, 137–146. [Google Scholar] [CrossRef]
- Hajji, S.; Younes, I.; Ghorbel-Bellaaj, O.; Hajji, R.; Rinaudo, M.; Nasri, M.; Jellouli, K. Structural differences between chitin and chitosan extracted from three different marine sources. Int. J. Biol. Macromol. 2014, 65, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, T.; Moores, A. Mechanochemical amorphization of chitin: Impact of apparatus material on performance and contamination. Beilstein J. Org. Chem. 2019, 15, 1217–1225. [Google Scholar] [CrossRef] [Green Version]
- Reginster, J.Y.; Deroisy, R.; Rovati, L.C.; Lee, R.L.; Lejeune, E.; Bruyere, O.; Giacovelli, G.; Henrotin, Y.; Dacre, J.E.; Gossett, C. Long-term effects of glucosamine sulphate on osteoarthritis progression: A randomised, placebo-controlled clinical trial. Lancet 2001, 357, 251–256. [Google Scholar] [CrossRef]
- Huang, B.; Xiao, D.; Tan, B.; Xiao, H.; Wang, J.; Yin, J.; Duan, J.; Huang, R.; Yang, C.; Yin, Y. Chitosan oligosaccharide reduces intestinal inflammation that involves calcium-sensing receptor (CaSR) activation in lipopolysaccharide (LPS)-challenged piglets. J. Agric. Food Chem. 2016, 64, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.-H.; Lin, Y.-Y.; Wu, G.-J.; Huang, C.-H.; Tsai, G.J. Effect of chitosan molecular weight on anti-inflammatory activity in the RAW 264.7 macrophage model. Int. J. Biol. Macromol. 2019, 131, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C.; Fu, E.; Shen, E.C. Chitosan inhibits prostaglandin E2 formation and cyclooxygenase-2 induction in lipopolysaccharide-treated RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2003, 308, 403–407. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dried Mealworm | |
|---|---|
| Crude carbohydrate (g/100 g) | 11.45 ± 0.38 |
| Total soluble sugar (g/100 g) | 3.22 ± 0.10 |
| Reducing sugar (g/100 g) | 0.19 ± 0.01 |
| Total glucosamine (g/100 g) | 4.84 ± 0.43 |
| Free sugar (mg/100 g) | |
| Glucose | 31.02 ± 1.95 |
| Fructose | 77.36 ± 0.35 |
| Sucrose | 12.46 ± 0.94 |
| Total | 120.84 ± 1.07 |
| Chitin from Mealworm | |
|---|---|
| Yield (g chitin/100 g dried mealworm powder) | 4.72 ± 0.21 |
| Moisture (g/100 g chitin) | 2.38 ± 0.05 |
| Crude protein (g/100 g chitin) | 9.96 ± 0.36 |
| Color | |
| L* (lightness) | 82.21 ± 0.72 |
| a* (redness) | 1.81 ± 0.08 |
| b* (yellowness) | 8.68 ± 0.37 |
| Whiteness index (WI) | 80.13 ± 0.81 |
| Chitin from Mealworm | Chitin from Shrimp | t-Value | |
|---|---|---|---|
| C/N † | 6.7743 ± 0.0038 | 6.7707 ± 0.0010 | −1.310 NS§ |
| DA ‡ (%) | 95.02 ± 0.22 | 94.81 ± 0.05 |
| Mealworm Chitin | Shrimp Chitin | t-Value | |
|---|---|---|---|
| Degree of crystallinity (%) | 57.85 ± 0.17 | 62.04 ± 0.10 | −37.053 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, Y.-J.; Hwang, I.-K.; Nho, C.W.; Kim, S.M.; Kim, S.H. Determination of Carbohydrate Composition in Mealworm (Tenebrio molitor L.) Larvae and Characterization of Mealworm Chitin and Chitosan. Foods 2021, 10, 640. https://doi.org/10.3390/foods10030640
Son Y-J, Hwang I-K, Nho CW, Kim SM, Kim SH. Determination of Carbohydrate Composition in Mealworm (Tenebrio molitor L.) Larvae and Characterization of Mealworm Chitin and Chitosan. Foods. 2021; 10(3):640. https://doi.org/10.3390/foods10030640
Chicago/Turabian StyleSon, Yang-Ju, In-Kyeong Hwang, Chu Won Nho, Sang Min Kim, and Soo Hee Kim. 2021. "Determination of Carbohydrate Composition in Mealworm (Tenebrio molitor L.) Larvae and Characterization of Mealworm Chitin and Chitosan" Foods 10, no. 3: 640. https://doi.org/10.3390/foods10030640