
Toxicological Evaluation of Piceatannol, Pterostilbene, and ε-Viniferin for Their Potential Use in the Food Industry: A Review


Abstract
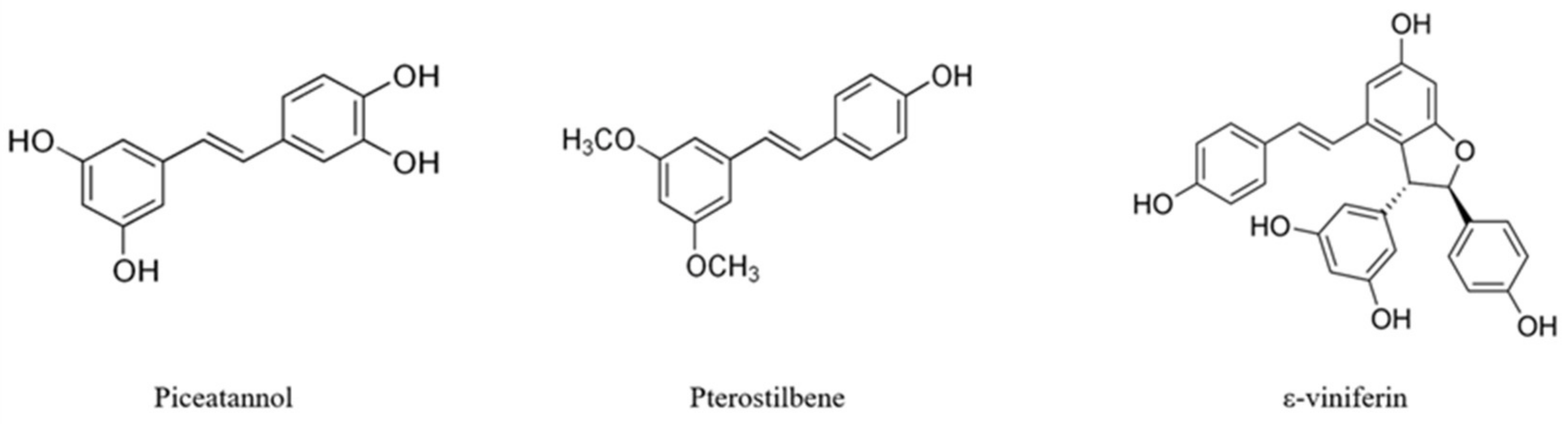
:1. Introduction
2. Cytotoxicity in In Vitro Studies Performed with Stilbenes
3. Genotoxicity in In Vitro Studies Performed with Stilbenes
4. Toxicological In Vivo Studies Performed with Stilbenes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Freyssin, A.; Page, G.; Fauconneau, B.; Rioux Bilan, A. Natural Stilbenes Effects in Animal Models of Alzheimer’s Disease. Neural Regen. Res. 2020, 15, 843–849. [Google Scholar] [CrossRef]
- Sirerol, J.A.; Rodríguez, M.L.; Mena, S.; Asensi, M.A.; Estrela, J.M.; Ortega, A.L. Role of Natural Stilbenes in the Prevention of Cancer. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutrim, C.S.; Cortez, M.A.S. A Review on Polyphenols: Classification, Beneficial Effects and Their Application in Dairy Products. Int. J. Dairy Technol. 2018, 71, 564–578. [Google Scholar] [CrossRef]
- Rivière, C.; Pawlus, A.D.; Mérillon, J.M. Natural Stilbenoids: Distribution in the Plant Kingdom and Chemotaxonomic Interest in Vitaceae. Nat. Prod. Rep. 2012, 29, 1317–1333. [Google Scholar] [CrossRef]
- Likhtenshtein, G.I. Stilbenes Synthesis and Applications. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2012. [Google Scholar] [CrossRef]
- Khan, Z.A.; Iqbal, A.; Shahzad, S.A. Synthetic Approaches toward Stilbenes and Their Related Structures. Mol. Divers. 2017, 21, 483–509. [Google Scholar] [CrossRef] [PubMed]
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A Review of Dietary Stilbenes: Sources and Bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Tresserra-Rimbau, A.; Lamuela-Raventos, R.M.; Moreno, J.J. Polyphenols, Food and Pharma. Current Knowledge and Directions for Future Research. Biochem. Pharmacol. 2018, 156, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Cicatiello, C.; Franco, S.; Pancino, B.; Blasi, E. The Value of Food Waste: An Exploratory Study on Retailing. J. Retail. Consum. Serv. 2016, 30, 96–104. [Google Scholar] [CrossRef]
- Singh, D.; Mendonsa, R.; Koli, M.; Subramanian, M.; Nayak, S.K. Antibacterial Activity of Resveratrol Structural Analogues: A Mechanistic Evaluation of the Structure-Activity Relationship. Toxicol. Appl. Pharmacol. 2019, 367, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.S.; Loh, Y.C.; Tew, W.Y.; Yam, M.F. Vasorelaxant Effect of 3, 5, 4′-Trihydroxy-Trans-Stilbene (Resveratrol) and Its Underlying Mechanism. Inflammopharmacology 2020, 28, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Sureda, A.; Belwal, T.; Çetinkaya, S.; Süntar, İ.; Tejada, S.; Devkota, H.P.; Ullah, H.; Aschner, M. Polyphenols in the Treatment of Autoimmune Diseases. Autoimmun. Rev. 2019, 18, 647–657. [Google Scholar] [CrossRef]
- Silva, P.; Sureda, A.; Tur, J.A.; Andreoletti, P.; Cherkaoui-Malki, M.; Latruffe, N. How Efficient Is Resveratrol as an Antioxidant of the Mediterranean Diet, towards Alterations during the Aging Process? Free Radic. Res. 2019, 53, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Schieber, A. Side Streams of Plant Food Processing As a Source of Valuable Compounds: Selected Examples. Annu. Rev. Food Sci. Technol. 2017, 8, 97–112. [Google Scholar] [CrossRef] [PubMed]
- EFSA Scientific Opinion. Guidance for Submission for Food Additive Evaluations. EFSA J. 2012, 10. [Google Scholar] [CrossRef]
- EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies). Safety of Synthetic Trans-resveratrol as a Novel Food Pursuant to Regulation (EC) No 258/97. EFSA J. 2016, 14, 4368. [Google Scholar] [CrossRef] [Green Version]
- Mankowski, R.T.; You, L.; Buford, T.W.; Leeuwenburgh, C.; Manini, T.M.; Schneider, S.; Qiu, P.; Anton, S.D. Higher Dose of Resveratrol Elevated Cardiovascular Disease Risk Biomarker Levels in Overweight Older Adults—A Pilot Study. Exp. Gerontol. 2020, 131, 110821. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-rahman, A.; Anyangwe, N.; Carlacci, L.; Casper, S.; Danam, R.P.; Enongene, E.; Erives, G.; Fabricant, D.; Gudi, R.; Hilmas, C.J.; et al. The Safety and Regulation of Natural Products Used as Foods and Food Ingredients. Toxicol. Sci. 2011, 123, 333–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbo, B.D.; André-Miral, C.; Nasre-Nasser, R.G.; Schimith, L.E.; Santos, M.G.; Costa-Silva, D.; Muccillo-Baisch, A.L.; Hort, M.A. Resveratrol Derivatives as Potential Treatments for Alzheimer’s and Parkinson’s Disease. Front. Aging Neurosci. 2020, 12. [Google Scholar] [CrossRef]
- Maisanaba, S.; Llana-Ruiz-Cabello, M.; Gutiérrez-Praena, D.; Pichardo, S.; Puerto, M.; Prieto, A.I.; Jos, A.; Cameán, A.M. New Advances in Active Packaging Incorporated with Essential Oils or Their Main Components for Food Preservation. Food Rev. Int. 2017, 33, 447–515. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [Green Version]
- Billack, B.; Radkar, V.; Adiabouah, C. In Vitro Evaluation of the Cytotoxic and Antiproliferative Properties of Resveratrol and Several of Its Analogs. Cell. Mol. Biol. Lett. 2008, 13, 553–569. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Haza, A.I. Selective Apoptotic Effects of Piceatannol and Myricetin in Human Cancer Cells. J. Appl. Toxicol. 2012, 32, 986–993. [Google Scholar] [CrossRef]
- Farrand, L.; Byun, S.; Kim, J.Y.; Im-Aram, A.; Lee, J.; Lim, S.; Lee, K.W.; Suh, J.Y.; Lee, H.J.; Tsang, B.K. Piceatannol Enhances Cisplatin Sensitivity in Ovarian Cancer via Modulation of P53, X-Linked Inhibitor of Apoptosis Protein (XIAP), and Mitochondrial Fission. J. Biol. Chem. 2013, 288, 23740–23750. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, M.J.; Lin, C.W.; Yang, S.F.; Sheu, G.T.; Yu, Y.Y.; Chen, M.K.; Chiou, H.L. A Combination of Pterostilbene with Autophagy Inhibitors Exerts Efficient Apoptotic Characteristics in Both Chemosensitive and Chemoresistant Lung Cancer Cells. Toxicol. Sci. 2014, 137, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalal, M.; Klinguer, A.; Echairi, A.; Meunier, P.; Vervandier-Fasseur, D.; Adrian, M. Antimicrobial Activity of Resveratrol Analogues. Molecules 2014, 19, 7679–7688. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.H.; Dai, D.K.; Zheng, B.Z.Y.; Duan, R.; Dong, T.T.X.; Qin, Q.W.; Tsim, K.W.K. Piceatannol, a Natural Analog of Resveratrol, Exerts Anti-Angiogenic Efficiencies by Blockage of Vascular Endothelial Growth Factor Binding to Its Receptor. Molecules 2020, 25, 3769. [Google Scholar] [CrossRef]
- Tolba, M.F.; Abdel-Rahman, S.Z. Pterostilbine, an Active Component of Blueberries, Sensitizes Colon Cancer Cells to 5-Fluorouracil Cytotoxicity. Sci. Rep. 2015, 5, 15239. [Google Scholar] [CrossRef] [Green Version]
- Zghonda, N.; Yoshida, S.; Araki, M.; Kusunoki, M.; Mliki, A.; Ghorbel, A.; Miyazaki, H. Greater Effectiveness of ε-Viniferin in Red Wine than Its Monomer Resveratrol for Inhibiting Vascular Smooth Muscle Cell Proliferation and Migration. Biosci. Biotechnol. Biochem. 2011, 75, 1259–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmazer, A. Cancer Cell Lines Involving Cancer Stem Cell Populations Respond to Oxidative Stress. Biotechnol. Rep. 2018, 17, 24–30. [Google Scholar] [CrossRef]
- Chowdhury, S.A.; Kishino, K.; Satoh, R.; Hashimoto, K.; Kikuchi, H.; Nishikawa, H.; Shirataki, Y.; Sakagami, H. Tumor-Specificity and Apoptosis-Inducing Activity of Stilbenes and Flavonoids. Anticancer Res. 2005, 25, 2055–2063. [Google Scholar]
- Fritzer-Szekeres, M.; Savinc, I.; Horvath, Z.; Saiko, P.; Pemberger, M.; Graser, G.; Bernhaus, A.; Ozsvar-Kozma, M.; Grusch, M.; Jaeger, W.; et al. Biochemical effects of piceatannol in human HL-60 promyelocytic leukemia cells—Synergism with Ara-C. Int. J. Oncol. 2008, 33, 887–892. [Google Scholar] [CrossRef]
- Kang, C.H.; Moon, D.O.; Choi, Y.H.; Choi, I.W.; Moon, S.K.; Kim, W.J.; Kim, G.Y. Piceatannol Enhances TRAIL-Induced Apoptosis in Human Leukemia THP-1 Cells through Sp1- and ERK-Dependent DR5 up-Regulation. Toxicol. Vitr. 2011, 25, 605–612. [Google Scholar] [CrossRef]
- Ovesná, Z.; Kozics, K.; Bader, Y.; Saiko, P.; Handler, N.; Erker, T.; Szekeres, T. Antioxidant Activity of Resveratrol, Piceatannol and 3,3′,4,4′,5,5′-Hexahydroxy-Trans-Stilbenein Three Leukemia Cell Lines. Oncol. Rep. 2006, 16, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Tolomeo, M.; Grimaudo, S.; Di Cristina, A.; Roberti, M.; Pizzirani, D.; Meli, M.; Dusonchet, L.; Gebbia, N.; Abbadessa, V.; Crosta, L.; et al. Pterostilbene and 3′-Hydroxypterostilbene Are Effective Apoptosis-Inducing Agents in MDR and BCR-ABL-Expressing Leukemia Cells. Int. J. Biochem. Cell Biol. 2005, 37, 1709–1726. [Google Scholar] [CrossRef]
- Ha, D.T.; Chen, Q.C.; Hung, T.M.; Youn, U.J.; Ngoc, T.M.; Thuong, P.T.; Kim, H.J.; Seong, Y.H.; Min, B.S.; Bae, K. Stilbenes and Oligostilbenes from Leaf and Stem of Vitis Amurensis and Their Cytotoxic Activity. Arch. Pharm. Res. 2009, 32, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Roslie, H.; Chan, K.M.; Rajab, N.F.; Velu, S.S.; Kadir, S.A.I.A.S.A.; Bunyamin, I.; Weber, J.F.F.; Thomas, N.F.; Majeed, A.B.A.; Myatt, G.; et al. 3,5-Dibenzyloxy-4′-Hydroxystilbene Induces Early Caspase-9 Activation during Apoptosis in Human K562 Chronic Myelogenous Leukemia Cells. J. Toxicol. Sci. 2012, 37, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, M.; Zhang, Z.; Gao, T. Piceatannol Induced Apoptosis through Up-Regulation of MicroRNA-181a in Melanoma Cells. Biol. Res. 2017, 50, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Tomás-Barberán, F.A.; Espín, J.C. Grape Polyphenol Resveratrol and the Related Molecule 4-Hydroxystilbene Induce Growth Inhibition, Apoptosis, S-Phase Arrest, and Upregulation of Cyclins A, E, and B1 in Human SK-Mel-28 Melanoma Cells. J. Agric. Food Chem. 2003, 51, 4576–4584. [Google Scholar] [CrossRef]
- Larrosa, M.; Tomás-Barberán, F.A.; Espín, J.C. The Grape and Wine Polyphenol Piceatannol Is a Potent Inducer of Apoptosis in Human SK-Mel-28 Melanoma Cells. Eur. J. Nutr. 2004, 43, 275–284. [Google Scholar] [CrossRef]
- Yokozawa, T.; Kim, Y.J. Piceatannol Inhibits Melanogenesis by Its Antioxidative Actions. Biol. Pharm. Bull. 2007, 30, 2007–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesołowska, O.; Wiśniewski, J.; Bielawska-Pohl, A.; Paprocka, M.; Duarte, N.; Ferreira, M.J.U.; Duś, D.; Michalak, K. Stilbenes as Multidrug Resistance Modulators and Apoptosis Inducers in Human Adenocarcinoma Cells. Anticancer Res. 2010, 30, 4587–4593. [Google Scholar] [PubMed]
- Wolter, F.; Clausnitzer, A.; Akoglu, B.; Stein, J. Piceatannol, a Natural Analog of Resveratrol, Inhibits Progression through the s Phase of the Cell Cycle in Colorectal Cancer Cell Lines. J. Nutr. 2002, 132, 298–302. [Google Scholar] [CrossRef] [Green Version]
- Dias, S.J.; Li, K.; Rimando, A.M.; Dhar, S.; Mizuno, C.S.; Penman, A.D.; Levenson, A.S. Trimethoxy-Resveratrol and Piceatannol Administered Orally Suppress and Inhibit Tumor Formation and Growth in Prostate Cancer Xenografts. Prostate 2013, 73, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Sáez, V.; Pastene, E.; Vergara, C.; Mardones, C.; Hermosín-Gutiérrez, I.; Gómez-Alonso, S.; Gómez, M.V.; Theoduloz, C.; Riquelme, S.; von Baer, D. Oligostilbenoids in Vitis Vinifera L. Pinot Noir Grape Cane Extract: Isolation, Characterization, in Vitro Antioxidant Capacity and Anti-Proliferative Effect on Cancer Cells. Food Chem. 2018, 265, 101–110. [Google Scholar] [CrossRef]
- Murias, M.; Jäger, W.; Handler, N.; Erker, T.; Horvath, Z.; Szekeres, T.; Nohl, H.; Gille, L. Antioxidant, Prooxidant and Cytotoxic Activity of Hydroxylated Resveratrol Analogues: Structure-Activity Relationship. Biochem. Pharmacol. 2005, 69, 903–912. [Google Scholar] [CrossRef]
- Alas, S.; Bonavida, B. Inhibition of Constitutive STAT3 Activity Sensitizes Resistant Non-Hodgkin’s Lymphoma and Multiple Myeloma to Chemotherapeutic Drug-Mediated Apoptosis. Clin. Cancer Res. 2003, 9, 316–326. [Google Scholar]
- Liu, Y.; Wang, L.; Wu, Y.; Lv, C.; Li, X.; Cao, X.; Yang, M.; Feng, D.; Luo, Z. Pterostilbene Exerts Antitumor Activity against Human Osteosarcoma Cells by Inhibiting the JAK2/STAT3 Signaling Pathway. Toxicology 2013, 304, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Hasiah, A.H.; Ghazali, A.R.; Weber, J.F.F.; Velu, S.; Thomas, N.F.; Inayat Hussain, S.H. Cytotoxic and Antioxidant Effects of Methoxylated Stilbene Analogues on HepG2 Hepatoma and Chang Liver Cells: Implications for Structure Activity Relationship. Hum. Exp. Toxicol. 2011, 30, 138–144. [Google Scholar] [CrossRef]
- Wieder, T.; Prokop, A.; Bagci, B.; Essmann, F.; Bernicke, D.; Schulze-Osthoff, K.; Dörken, B.; Schmalz, H.G.; Daniel, P.T.; Henze, G. Piceatannol, a Hydroxylated Analog of the Chemopreventive Agent Resveratrol, Is a Potent Inducer of Apoptosis in the Lymphoma Cell Line BJAB and in Primary, Leukemic Lymphoblasts. Leukemia 2001, 15, 1735–1742. [Google Scholar] [CrossRef] [Green Version]
- Radkar, V.; Hardej, D.; Lau-Cam, C.; Billack, B. Evaluation of Resveratrol and Piceatannol Cytotoxicity in Macrophages, T Cells, and Skin Cells. Arh. Hig. Rada Toksikol. 2007, 58, 293–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Park, C.; Lee, J.O.; Kim, G.Y.; Lee, W.H.; Choi, Y.H.; Ryu, C.H. Induction of Apoptosis by Piceatannol in Human Leukemic U937 Cells through Down-Regulation of Bcl-2 and Activation of Caspases. Oncol. Rep. 2008, 19, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Kuo, P.L.; Hsu, Y.L. The Grape and Wine Constituent Piceatannol Inhibits Proliferation of Human Bladder Cancer Cells via Blocking Cell Cycle Progression and Inducing Fas/Membrane Bound Fas Ligand-Mediated Apoptotic Pathway. Mol. Nutr. Food Res. 2008, 52, 408–418. [Google Scholar] [CrossRef]
- Rüweler, M.; Gülden, M.; Maser, E.; Murias, M.; Seibert, H. Cytotoxic, Cytoprotective and Antioxidant Activities of Resveratrol and Analogues in C6 Astroglioma Cells in Vitro. Chem. Biol. Interact. 2009, 182, 128–135. [Google Scholar] [CrossRef]
- Liu, W.H.; Chang, L. Sen. Piceatannol Induces Fas and FasL Up-Regulation in Human Leukemia U937 Cells via Ca2+/P38α MAPK-Mediated Activation of c-Jun and ATF-2 Pathways. Int. J. Biochem. Cell Biol. 2010, 42, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Arai, D.; Kataoka, R.; Otsuka, S.; Kawamura, M.; Maruki-Uchida, H.; Sai, M.; Ito, T.; Nakao, Y. Piceatannol Is Superior to Resveratrol in Promoting Neural Stem Cell Differentiation into Astrocytes. Food Funct. 2016, 7, 4432–4441. [Google Scholar] [CrossRef] [Green Version]
- Takasawa, R.; Akahane, H.; Tanaka, H.; Shimada, N.; Yamamoto, T.; Uchida-Maruki, H.; Sai, M.; Yoshimori, A.; Tanuma, S.-i. Piceatannol, a Natural Trans-Stilbene Compound, Inhibits Human Glyoxalase I. Bioorg. Med. Chem. Lett. 2017, 27, 1169–1174. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, Y.; Xie, J.; Hu, Y.; Zhang, Y. Anti-Tumor Effect of Piceatannol through Induction of Cell Apoptosis via up-Regulation of MicroRNA-125b Expression on Pancreatic Cancer. Int. J. Clin. Exp. Med. 2017, 10, 14495–14502. [Google Scholar]
- Siedlecka-Kroplewska, K.; Ślebioda, T.; Kmieć, Z. Induction of Autophagy, Apoptosis and Aquisition of Resistance in Response to Piceatannol Toxicity in MOLT-4 Human Leukemia Cells. Toxicol. Vitr. 2019, 59, 12–25. [Google Scholar] [CrossRef]
- Liu, T.; Liu, M.; Guo, Q.; Liu, Y.; Zhao, Y.; Wu, Y.; Sun, B.; Wang, Q.; Liu, J.; Han, J. Investigation of Binary and Ternary Systems of Human Serum Albumin with Oxyresveratrol/Piceatannol and/or Mitoxantrone by Multipectroscopy, Molecular Docking and Cytotoxicity Evaluation. J. Mol. Liq. 2020, 311, 113364. [Google Scholar] [CrossRef]
- Chakraborty, A.; Gupta, N.; Ghosh, K.; Roy, P. In Vitro Evaluation of the Cytotoxic, Anti-Proliferative and Anti-Oxidant Properties of Pterostilbene Isolated from Pterocarpus Marsupium. Toxicol. Vitr. 2010, 24, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.J.; Ho, C.T.; Wang, Y.J. Pterostilbene Induces Autophagy and Apoptosis in Sensitive and Chemoresistant Human Bladder Cancer Cells. Mol. Nutr. Food Res. 2010, 54, 1819–1832. [Google Scholar] [CrossRef]
- Nutakul, W.; Sobers, H.S.; Qiu, P.; Dong, P.; Decker, E.A.; McClements, D.J.; Xiao, H. Inhibitory Effects of Resveratrol and Pterostilbene on Human Colon Cancer Cells: A Side-by-Side Comparison. J. Agric. Food Chem. 2011, 59, 10964–10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobolev, V.S.; Khan, S.I.; Tabanca, N.; Wedge, D.E.; Manly, S.P.; Cutler, S.J.; Coy, M.R.; Becnel, J.J.; Neff, S.A.; Gloer, J.B. Biological Activity of Peanut (Arachis hypogaea) Phytoalexins and Selected Natural and Synthetic Stilbenoids. J. Agric. Food Chem. 2011, 59, 1673–1682. [Google Scholar] [CrossRef] [Green Version]
- Harun, Z.; Ghazali, A.R. Potential Chemoprevention Activity of Pterostilbene by Enhancing the Detoxifying Enzymes in the HT-29 Cell Line. Asian Pac. J. Cancer Prev. 2012, 13, 6403–6407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, G.; Prosperini, A.; Font, G.; Ruiz, M.J. Effect of Polyphenols on Enniatins-Induced Cytotoxic Effects in Mammalian Cells. Toxicol. Mech. Methods 2012, 22, 687–695. [Google Scholar] [CrossRef]
- Mena, S.; Rodríguez, M.L.; Ponsoda, X.; Estrela, J.M.; Jäättela, M.; Ortega, A.L. Pterostilbene-Induced Tumor Cytotoxicity: A Lysosomal Membrane Permeabilization-Dependent Mechanism. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Siedlecka-Kroplewska, K.; Jozwik, A.; Kaszubowska, L.; Kowalczyk, A.; Boguslawski, W. Pterostilbene Induces Cell Cycle Arrest and Apoptosis in MOLT4 Human Leukemia Cells. Folia Histochem. Cytobiol. 2012, 50, 574–580. [Google Scholar] [CrossRef]
- Wang, Y.; Ding, L.; Wang, X.; Zhang, J.; Han, W.; Feng, L.; Sun, J.; Jin, H.; Wang, X.J. Pterostilbene Simultaneously Induces Apoptosis, Cell Cycle Arrest and Cyto-Protective Autophagy in Breast Cancer Cells. Am. J. Transl. Res. 2012, 4, 44–51. [Google Scholar] [PubMed]
- Pino, M.A.; Pietka-Ottlik, M.; Billack, B. Ebselen Analogues Reduce 2-Chloroethyl Ethyl Sulphide Toxicity in A-431 Cells. Arh. Hig. Rada Toksikol. 2013, 64, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikhil, K.; Sharan, S.; Chakraborty, A.; Bodipati, N.; Krishna Peddinti, R.; Roy, P. Role of Isothiocyanate Conjugate of Pterostilbene on the Inhibition of MCF-7 Cell Proliferation and Tumor Growth in Ehrlich Ascitic Cell Induced Tumor Bearing Mice. Exp. Cell Res. 2014, 320, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Wawszczyk, J.; Kapral, M.; Hollek, A.; Węglarz, L. In Vitro Evaluation of Antiproliferative and Cytotoxic Properties of Pterostilbene against Human Colon Cancer Cells. Acta Pol. Pharm. Drug Res. 2014, 71, 1051–1055. [Google Scholar]
- Zhang, B.; Wang, X.Q.; Chen, H.Y.; Liu, B.H. Involvement of the Nrf2 Pathway in the Regulation of Pterostilbene-Induced Apoptosis in HeLa Cells via ER Stress. J. Pharmacol. Sci. 2014, 126, 216–229. [Google Scholar] [CrossRef] [Green Version]
- Ko, C.P.; Lin, C.W.; Chen, M.K.; Yang, S.F.; Chiou, H.L.; Hsieh, M.J. Pterostilbene Induce Autophagy on Human Oral Cancer Cells through Modulation of Akt and Mitogen-Activated Protein Kinase Pathway. Oral Oncol. 2015, 51, 593–601. [Google Scholar] [CrossRef]
- Wu, C.H.; Hong, B.H.; Ho, C.T.; Yen, G.C. Targeting Cancer Stem Cells in Breast Cancer: Potential Anticancer Properties of 6-Shogaol and Pterostilbene. J. Agric. Food Chem. 2015, 63, 2432–2441. [Google Scholar] [CrossRef]
- Kong, Y.; Chen, G.; Xu, Z.; Yang, G.; Li, B.; Wu, X.; Xiao, W.; Xie, B.; Hu, L.; Sun, X.; et al. Pterostilbene Induces Apoptosis and Cell Cycle Arrest in Diffuse Large B-Cell Lymphoma Cells. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xie, B.; Xu, Z.; Hu, L.; Chen, G.; Wei, R.; Yang, G.; Li, B.; Chang, G.; Sun, X.; Wu, H.; et al. Pterostilbene Inhibits Human Multiple Myeloma Cells via ERK1/2 and JNK Pathway in Vitro and in Vivo. Int. J. Mol. Sci. 2016, 17, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiabouah Achy-Brou, C.A.; Billack, B. A Comparative Assessment of the Cytotoxicity and Nitric Oxide Reducing Ability of Resveratrol, Pterostilbene and Piceatannol in Transformed and Normal Mouse Macrophages. Drug Chem. Toxicol. 2017, 40, 36–46. [Google Scholar] [CrossRef]
- Hung, C.M.; Liu, L.C.; Ho, C.T.; Lin, Y.C.; Way, T. Der. Pterostilbene Enhances TRAIL-Induced Apoptosis through the Induction of Death Receptors and Downregulation of Cell Survival Proteins in TRAIL-Resistance Triple Negative Breast Cancer Cells. J. Agric. Food Chem. 2017, 65, 11179–11191. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, Y.; Di, S.; Feng, X.; Liu, D.; Jiang, S.; Hu, W.; Qin, Z.; Li, Y.; Lv, J.; et al. Pterostilbene Exerts Anticancer Activity on Non-Small-Cell Lung Cancer via Activating Endoplasmic Reticulum Stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bin, W.H.; Da, L.H.; Xue, Y.; Jing, B.W. Pterostilbene (3′,5′-Dimethoxy-Resveratrol) Exerts Potent Antitumor Effects in HeLa Human Cervical Cancer Cells via Disruption of Mitochondrial Membrane Potential, Apoptosis Induction and Targeting m-TOR/PI3K/Akt Signalling Pathway. JBUON 2018, 23, 1384–1389. [Google Scholar]
- Chang, H.P.; Lu, C.C.; Chiang, J.H.; Tsai, F.J.; Juan, Y.N.; Tsao, J.W.; Chiu, H.Y.; Yang, J.S. Pterostilbene Modulates the Suppression of Multidrug Resistance Protein 1 and Triggers Autophagic and Apoptotic Mechanisms in Cisplatin-Resistant Human Oral Cancer CAR Cells via AKT Signaling. Int. J. Oncol. 2018, 52, 1504–1514. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, K.; AlSharif, D.; Mazza, C.; Syar, P.; Al Sharif, M.; Fata, J.E. Resveratrol and Pterostilbene Exhibit Anticancer Properties Involving the Downregulation of HPV Oncoprotein E6 in Cervical Cancer Cells. Nutrients 2018, 10, 243. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Chen, L.; Liu, X.; Li, X.; Cao, Y.; Bai, Y.; Qi, F. Pterostilbene Inhibits Amyloid-β-Induced Neuroinflammation in a Microglia Cell Line by Inactivating the NLRP3/Caspase-1 Inflammasome Pathway. J. Cell. Biochem. 2018, 119, 7053–7062. [Google Scholar] [CrossRef]
- Liu, K.F.; Liu, Y.X.; Dai, L.; Li, C.X.; Wang, L.; Liu, J.; Lei, J.D. A Novel Self-Assembled PH-Sensitive Targeted Nanoparticle Platform Based on Antibody-4arm-Polyethylene Glycol-Pterostilbene Conjugates for Co-Delivery of Anticancer Drugs. J. Mater. Chem. B 2018, 6, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, K.; Mukherjee, S.; Vanmanen, J.; Banerjee, P.; Fata, J.E. Dietary Polyphenols, Resveratrol and Pterostilbene Exhibit Antitumor Activity on an HPV E6-Positive Cervical Cancer Model: An in Vitro and in Vivo Analysis. Front. Oncol. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.T.; Chen, P.W.; Li, S.; Ke, T.M.; Lin, S.H.; Yang, C.C. Pterostilbene Inhibits Lung Squamous Cell Carcinoma Growth in Vitro and in Vivo by Inducing S Phase Arrest and Apoptosis. Oncol. Lett. 2019, 18, 1631–1640. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Shin, E.A.; Kim, J.H.; Sim, D.Y.; Lee, H.; Park, J.E.; Lee, H.J.; Kim, S.H. NEDD9 Inhibition by MiR-25-5p Activation Is Critically Involved in Co-Treatment of Melatonin-and Pterostilbene-Induced Apoptosis in Colorectal Cancer Cells. Cancers 2019, 11, 1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Ci, X.; Ma, X.; Yu, Q.; Cui, Y.; Zhen, Y.; Li, S. Pterostilbene Activates the Nrf2-Dependent Antioxidant Response to Ameliorate Arsenic-Induced Intracellular Damage and Apoptosis in Human Keratinocytes. Front. Pharmacol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Hsu, Y.H.; Chen, S.Y.; Wang, S.Y.; Lin, J.A.; Yen, G.C. Pterostilbene Enhances Cytotoxicity and Chemosensitivity in Human Pancreatic Cancer Cells. Biomolecules 2020, 10, 709. [Google Scholar] [CrossRef]
- Zhao, Y.; Ye, D.; Luo, Q.; Li, J.; Liu, J. Pterostilbene Inhibits Human Renal Cell Carcinoma Cells Growth and Induces DNA Damage. Biol. Pharm. Bull. 2020, 43, 258–265. [Google Scholar] [CrossRef]
- Tian, X.; Guo, S.; Zhang, S.; Li, P.; Wang, T.; Ho, C.T.; Pan, M.H.; Bai, N. Chemical Characterization of Main Bioactive Constituents in Paeonia Ostii Seed Meal and GC-MS Analysis of Seed Oil. J. Food Biochem. 2020, 44, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Chang, E.J.; Sung, H.C.; Chung, S.K.; Park, H.D.; Choi, W.C. Cytotoxic and Antimutagenic Stilbenes from Seeds of Paeonia lactiflora. Arch. Pharm. Res. 2002, 25, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Billard, C.; Izard, J.C.; Roman, V.; Kern, C.; Mathiot, C.; Mentz, F.; Kolb, J.P. Comparative Antiproliferative and Apoptotic Effects of Resveratrol, ε-Viniferin and Vine-Shots Derived Polyphenols (Vineatrols) on Chronic B Lymphocytic Leukemia Cells and Normal Human Lymphocytes. Leuk. Lymphoma 2002, 43, 1991–2002. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Park, Y.H.; Choi, S.W.; Yang, E.K.; Lee, W.J. Resveratrol Derivatives Potently Induce Apoptosis in Human Promyelocytic Leukemia Cells. Exp. Mol. Med. 2003, 35, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Muhtadi Hakim, E.H.; Juliawaty, L.D.; Syah, Y.M.; Achmad, S.A.; Latip, J.; Ghisalberti, E.L. Cytotoxic Resveratrol Oligomers from the Tree Bark of Dipterocarpus hasseltii. Fitoterapia 2006, 77, 550–555. [Google Scholar] [CrossRef]
- Colin, D.; Lancon, A.; Delmas, D.; Lizard, G.; Abrossinow, J.; Kahn, E.; Jannin, B.; Latruffe, N. Antiproliferative Activities of Resveratrol and Related Compounds in Human Hepatocyte Derived HepG2 Cells Are Associated with Biochemical Cell Disturbance Revealed by Fluorescence Analyses. Biochimie 2008, 90, 1674–1684. [Google Scholar] [CrossRef] [Green Version]
- Marel, A.K.; Lizard, G.; Izard, J.C.; Latruffe, N.; Delmas, D. Inhibitory Effects of Trans-Resveratrol Analogs Molecules on the Proliferation and the Cell Cycle Progression of Human Colon Tumoral Cells. Mol. Nutr. Food Res. 2008, 52, 538–548. [Google Scholar] [CrossRef] [Green Version]
- Richard, T.; Poupard, P.; Nassra, M.; Papastamoulis, Y.; Iglésias, M.L.; Krisa, S.; Waffo-Teguo, P.; Mérillon, J.M.; Monti, J.P. Protective Effect of ε-Viniferin on β-Amyloid Peptide Aggregation Investigated by Electrospray Ionization Mass Spectrometry. Bioorg. Med. Chem. 2011, 19, 3152–3155. [Google Scholar] [CrossRef]
- Rohaiza, S.; Yaacob, W.A.; Din, L.B.; Nazlina, I. Cytotoxic Oligostilbenes from Shorea Hopeifolia. Afr. J. Pharm. Pharmacol. 2011, 5, 1272–1277. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.I.; Chien, W.C.; Huang, K.X.; Hsu, J.L. Anti-Inflammatory Effects of Vitisinol A and Four Other Oligostilbenes from Ampelopsis Brevipedunculata Var. Hancei. Molecules 2017, 22, 1195. [Google Scholar] [CrossRef]
- Nivelle, L.; Aires, V.; Rioult, D.; Martiny, L.; Tarpin, M.; Delmas, D. Molecular Analysis of Differential Antiproliferative Activity of Resveratrol, Epsilon Viniferin and Labruscol on Melanoma Cells and Normal Dermal Cells. Food Chem. Toxicol. 2018, 116, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, F.; Apaydın, E.; Önder, N.İ.; Şen, M.; Ayrım, A.; Öğünç, Y.; İncesu, Z. Apoptotic Effects of ε-Viniferin in Combination with Cis-Platin in C6 Cells. Cytotechnology 2018, 70, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Vion, E.; Page, G.; Bourdeaud, E.; Paccalin, M.; Guillard, J.; Rioux Bilan, A. Trans ε-Viniferin Is an Amyloid-β Disaggregating and Anti-Inflammatory Drug in a Mouse Primary Cellular Model of Alzheimer’s Disease. Mol. Cell. Neurosci. 2018, 88, 1–6. [Google Scholar] [CrossRef]
- Courtois, A.; Garcia, M.; Krisa, S.; Atgié, C.; Sauvant, P.; Richard, T.; Faure, C. Encapsulation of ϵ-Viniferin in Onion-Type Multi-Lamellar Liposomes Increases Its Solubility and Its Photo-Stability and Decreases Its Cytotoxicity on Caco-2 Intestinal Cells. Food Funct. 2019, 10, 2573–2582. [Google Scholar] [CrossRef] [PubMed]
- Aja, I.; Begoña Ruiz-Larrea, M.; Courtois, A.; Krisa, S.; Richard, T.; Ruiz-Sanz, J.I. Screening of Natural Stilbene Oligomers from Vitis Vinifera for Anticancer Activity on Human Hepatocellular Carcinoma Cells. Antioxidants 2020, 9, 469. [Google Scholar] [CrossRef] [PubMed]
- Medrano-Padial, C.; Puerto, M.; del Mar Merchán-Gragero, M.; Moreno, F.J.; Richard, T.; Cantos-Villar, E.; Pichardo, S. Cytotoxicity Studies of a Stilbene Extract and Its Main Components Intended to Be Used as Preservative in the Wine Industry. Food Res. Int. 2020, 137. [Google Scholar] [CrossRef] [PubMed]
- Azmi, A.S.; Bhat, S.H.; Hadi, S.M. Resveratrol-Cu(II) Induced DNA Breakage in Human Peripheral Lymphocytes: Implications for Anticancer Properties. FEBS Lett. 2005, 579, 3131–3135. [Google Scholar] [CrossRef] [Green Version]
- Makena, P.S.; Chung, K.T. Effects of Various Plant Polyphenols on Bladder Carcinogen Benzidine-Induced Mutagenicity. Food Chem. Toxicol. 2007, 45, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-C.; Lin, C.-Y.; Lin, H.-Y.; Wu, J.M. AKT/MTOR as Novel Targets of Polyphenol Piceatannol Possibly Contributing to Inhibition of Proliferation of Cultured Prostate Cancer Cells. ISRN Urol. 2012, 2012, 272697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanapal, J.; Balaraman Ravindrran, M. Chitosan/Poly (Lactic Acid)-Coated Piceatannol Nanoparticles Exert an in Vitro Apoptosis Activity on Liver, Lung and Breast Cancer Cell Lines. Artif. Cells, Nanomed. Biotechnol. 2018, 46, 274–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.Y.; Molagoda, I.M.N.; Park, C.; Kwon, T.K.; Yun, S.J.; Kim, W.J.; Kim, G.Y.; Choi, Y.H. Piceatannol-Induced Apoptosis Is Reversed by N-Acetyl-L-Cysteine through Restoration of XIAP Expression. Biol. Pharm. Bull. 2018, 41, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.; Caruso, F.; Antonioletti, R.; Viglianti, A.; Traversi, G.; Leone, S.; Basso, E.; Cozzi, R. Scavenging of Hydroxyl Radical by Resveratrol and Related Natural Stilbenes after Hydrogen Peroxide Attack on DNA. Chem. Biol. Interact. 2013, 206, 175–185. [Google Scholar] [CrossRef]
- Baechler, S.A.; Schroeter, A.; Dicker, M.; Steinberg, P.; Marko, D. Topoisomerase II-Targeting Properties of a Grapevine-Shoot Extract and Resveratrol Oligomers. J. Agric. Food Chem. 2014, 62, 780–788. [Google Scholar] [CrossRef] [PubMed]
- EFSA Scientific Opinion. Scientific Opinion on Genotoxicity Testing Strategies Applicable to Food and Feed Safety Assessment. EFSA J. 2011, 9. [Google Scholar] [CrossRef]
- Caillaud, M.; Guillard, J.; Richard, D.; Milin, S.; Chassaing, D.; Paccalin, M.; Page, G.; Bilan, A.R. Trans ε Viniferin Decreases Amyloid Deposits and Inflammation in a Mouse Transgenic Alzheimer Model. PLoS ONE 2019, 14, e212663. [Google Scholar] [CrossRef]
- Kiliç, V. Piceatannol Mediated Modulation of Oxidative Stress and Regeneration in the Liver of Endotoxemic Mice. J. Med. Food 2019, 22, 594–601. [Google Scholar] [CrossRef]
- Kosuru, R.; Rai, U.; Prakash, S.; Singh, A.; Singh, S. Promising Therapeutic Potential of Pterostilbene and Its Mechanistic Insight Based on Preclinical Evidence. Eur. J. Pharmacol. 2016, 789, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Fu, L. Piceatannol Inhibits Oxidative Stress through Modification of Nrf2-Signaling Pathway in Testes and Attenuates Spermatogenesis and Steroidogenesis in Rats Exposed to Cadmium during Adulthood. Drug Des. Devel. Ther. 2019, 13, 2811–2824. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.J.; Fernández, M.; Picó, Y.; Mañes, J.; Asensi, M.; Carda, C.; Asensio, G.; Estrela, J.M. Dietary Administration of High Doses of Pterostilbene and Quercetin to Mice Is Not Toxic. J. Agric. Food Chem. 2009, 57, 3180–3186. [Google Scholar] [CrossRef]
- Riche, D.M.; McEwen, C.L.; Riche, K.D.; Sherman, J.J.; Wofford, M.R.; Deschamp, D.; Griswold, M. Analysis of Safety from a Human Clinical Trial with Pterostilbene. J. Toxicol. 2013, 2013. [Google Scholar] [CrossRef]


{kind=link}
| Pure Stilbene | Experimental Model | Assays Performed | Exposure Conditions | Main Results | Reference |
|---|---|---|---|---|---|
| Piceatannol | BJAB cells | LDH activity | 25, 50, 75, and 100 µM for 4 h | The stilbene at concentrations ≤ 100 µM did not reduce cell viability, indicating that the membrane disrupting effect does not play a role in their death-inducing potency. | [51] |
| Piceatannol | Caco-2 and HCT-116 cells | Crystal violet | 12.5, 25, 50, 100, and 200 µM for 24, 48, and 72 h | A steady decrease in cell number was observed in a dose- and time-dependent manner. After the exposure of 200 µM for 72 h, the growth rate of cells decreased 60% ± 3.2% in Caco-2 cells and 58.3 ± 3.1% in HCT-116 cells. | [44] |
| Piceatannol | U266 and 2F7 cells | XTT assay | 50 µM for 24 h | Piceatannol displayed no cytotoxicity in any of the cell lines. | [48] |
| Piceatannol | SK-Mel-28 cells | MTT assay | 25, 50, and 100 µM for 96 h | The stilbene was rendered unstable only 4 h after its addition without an apparent effect on the cell cycle after 48 h of assay. | [40] |
| Piceatannol | SK-Mel-28 cells | Cell viability by flow cytometry | 1–100 µM for 4–48 h | Cell viability decreased with increasing concentrations and incubation time. | [41] |
| Piceatannol | HGF, HPC, HPLF, HSC-2, HSC-3, HSG, and HL-60 cells | MTT assay | 0–1000 µM for 24 h | The four tumor cell lines (HSC-2, HSC-3, HSG, and HL-60) were more sensitive to the stilbene than the three normal cell lines (HGF, HGC, and HPLF). The CC50 values were 367 µM for HGF, 414 µM for HPC, >1000 µM for HPLF, 63 µM for HSC-2, 232 µM for HSC-3, 373 µM for HSG, and 11 µM for HL-60 cells. | [32] |
| Piceatannol | HL-60 cells | CC-108 microcellcounter | 0–100 µM for 72 h | Significant decrease of cell viability at 10 µM. Piceatannol was more cytotoxic than resveratrol. The IC50 was set at 9.1 µM ± 0.28 | [47] |
| Piceatannol | L1210, K562, andHL-60 cells | TBET | 0–500 µM for 24 h | <20 µM and <10 µM concentrations caused cytotoxicity in L1210 and HL60 cells. The cytotoxic effect was lower in K562 cells. | [35] |
| Piceatannol (purity > 99%) | RAW264.7 cells | MTT assay | 0–50 µM for 48 h, with and without stimulation of zymosan | Cytotoxic effect was significant starting at 30 µM. This effect was attenuated to a significant extent by a cotreatment with zymosan. | [52] |
| Piceatannol | B16 cells | MTT assay | 5, 100, 200, and 400 µM for 24 h | No cytotoxicity. Cell viability was 99.8, 98.7, 95.3, and 90.1% at 5, 100, 200, and 400 µM, respectively. | [42] |
| Piceatannol (purity > 99%) | RAW 264.7, A–431, 10ScNCr/23, and CCR-CEM cells | TBET Cell proliferation studies using a hemocytometer | 0–50 µM for 24 h 10, 30, and 50 µM for 200 h | RAW 264.7 cells were more sensitive to piceatannol than other stilbenes (trans-resveratrol, trans-pterostilbene, and trans-stilbene-oxide). The IC50 was set at 1.30 µM ± 0.12. In the cell proliferation studies, there was no affection of 10ScNCr/23 cells at ≤50 μM, whereas an inhibitory effect was observed in RAW 264.7 and A431 cells at 50 μM, and in CCR-CEM cells at ≥10 μM. | [23] |
| Piceatannol | HL-60 cells | CC-108 microcellcounter | 3.125, 6.25, 12.5, and 25 µM for 72 h | The IC50 value was 14 µM after 3 days of incubation. | [33] |
| Piceatannol | U937 cells | TBET | 0–80 µM for 48 h | The 48 h treatment reduced cell viability in a concentration-dependent manner. | [53] |
| Piceatannol | T24 and HT1376 cells | XTT assay | 0.5, 2.5, 5, and 10 µM for 48 h | Dose-dependent effect. At 48 h, the maximum effect on proliferation inhibition was observed at 10 µM in both cells.The IC50 values were 3.9 µM in T24 cells and 4.6 µM in HT1376 cells. | [54] |
| Piceatannol (from Vitis amurensis) | L1210, K362, and HCT116 cells | MTT assay | 0–50 µM for 48 h | No cytotoxic effect was observed. The IC50 was not found and set at >50 µM. | [37] |
| Piceatannol | C6 cells (proliferating and growth arrested) | Protein content (Lorry method) | 1–100 µM for 72 h in proliferating cells and 24 h in growth-arrested cells | A cytotoxic effect at low micromolar concentrations was recorded in growth-arrested cells. The IC50 value in proliferating cells was 28 µM ±4 and in growth-arrested cells was 20 µM ± 2. | [55] |
| Piceatannol | U937 cells | MTT assay | 20, 40, 60, and 100 µM for 24 h | The cells showed a concentration-dependent and time-dependent decrease in cell viability. A reduction in cell viability of approximately 48% was observed after treatment with 5 µM for 24 h. | [56] |
| Piceatannol | LoVo and LoVo/doxorubicincells | SRB assay | 20, 40, 60, 80, and 100 µM for 72 h | In sensitive cells (LoVo), the effect of piceatannol and its derivative (trans-3, 5, 3’, 4’-tetracetoxystilbene) was more toxic than in resistant cells. After exposure to 100 µM for 72 h, the reduction of viability was approximately 50% in LoVo cells and 15% in Lovo/doxorubicin-resistant colon cells. | [43] |
| Piceatannol | THP-1, HL-60, and U937 cells | MTT assay | 10, 20, 30, 40, and 50 µM for 24 h | Treatment with piceatannol resulted in a dose-dependent inhibition of cell viability. | [34] |
| Piceatannol | HL-60 and HepG2 cells | MTT assay | 10–200 µM for 24, 48, and 72 h | A high inhibition was found after treatment with 100–200 µM for 24, 48, and 72 h in HL-60 cells. No significant effect on HepG2 cell growth at the doses and times used. | [24] |
| Piceatannol | K562 cells | MTT assay | 0–100 µM for 48 h | No cytotoxicity was recorded for the concentrations assayed. | [38] |
| Piceatannol (purity > 99%) | LNCaP, Du145, andPC3M cells | MTT assay | 1–100 µM for 6 days | The growth inhibitory effects found were cell specific. The IC50 was 31.7 µM in LNCaP cells, 23.2 µM in Du145 cells, and 34.6 µM in PC3M cells. | [45] |
| Piceatannol | OV2008, C13, A2780s, A2780cp, OVCAR-432, and SkOV-3 cells | MTS assay | 10 µM alone and in combination with 10 µM cis-diamminedichloroplatinum for 24 h in all cell lines 10 µM alone and in combination with 10 µM cis-diamminedichloroplatinum for 48 h in OV2008 | Piceatannol reduced cell viability in all tested cell lines and enhanced the cytotoxic effects of diamminedichloroplatinum in OV2008, A2780s, and OVCAR-432, concluding that p53 status is a determinant of piceatannol action. A time dependent decrease was observed in OV2008 viability after 48 h of exposure. Moreover, an additive effect with cis-diamminedichloroplatinum was determined. The IC50 value of piceatannol for 48 h in OV2008 cells was 29.1 µM. | [25] |
| Piceatannol | Undifferentiated neural stem cells | MTT assay | 1–20 µM for 72 h | No effect was observed at 2.5 µM or less. The IC50 was 13.5 µM. | [57] |
| Piceatannol (purity > 99%) | WM266-4 and A2058 cells | MTT assay | 0–200 µM for 36 h | The growth of both cells was inhibited in a dose-dependent manner. The IC50 was 29.4 µM in WM266–4 and 15.6 µM in A2058 cells. | [39] |
| Piceatannol | NCIH-522 cells | WST-8 assay | 10, 30, 50, 80, and 100 µM for 24, 48, and 72 h | The stilbene suppressed proliferation in a dose- and time- dependent manner. The IC50 was set at 53, 23, and 17 µM for 24, 48, and 72 h, respectively. | [58] |
| Piceatannol (purity > 99%) | SW1990 and PANC-1 cells | CCK-8 assay | 1, 10, 20, 40, 100, and 200 µM for 72 h | Cell proliferation was inhibited in a dose-dependent manner.The IC50 values were 30.69 µM and 21.82 µM for SW1990 and PANC-1 cells, respectively. | [59] |
| Piceatannol (purity ≥ 98%) | MRC-5, AGS, SK-MES-1, and J82 cells | MTT assay | 0–100 µg/mL for 72 h | No cytotoxic effect was observed in non-cancerous cells. The IC50 was set at >100 µM in MRC-5 cells, 44.4 µM ± 3.2 in AGS, 31.3 µM ± 2.1 in SK-MES-1 cells, and 27.7 µM ± 1.4 in J82 cells. | [46] |
| Piceatannol | MOLT-4 cells | NRU assay | 0.05, 15, 25, 50, and 100 µM for 48 h | A reduction in cell viability was observed in a concentration-dependent manner. Incubation with piceatannol for 6–8 h led to a significant increase in the number of cells in the sub-G1 fraction, indicating apoptotic DNA degradation. The IC50 value was 24.8 µM. | [60] |
| Piceatannol | HUVEC cells | MTT assay and LDH activity | 3–100 µM for 48 h | No effect on cell viability was found up to 30 µM. | [28] |
| Piceatannol (purity ≥ 98%) | HeLa cells | MTT assay | 0–250 µM for 48 h | The stilbene inhibited cell proliferation in a dose- dependent manner. The IC50 was 375.20 µM. | [61] |
| Pure Stilbene | Experimental Model | Assays Performed | Exposure Conditions | Main Results | Reference |
|---|---|---|---|---|---|
| Piceatannol | SK-Mel-28 cells | Hoechst 33258 staining | 1 µM for 8–48 h | Bright spherical beads could be seen in apoptotic cells. Apoptotic cells increased about 6-fold with respect to the control after 48 h. | [41] |
| Piceatannol | HL-60 cells | Hoechst 33258 staining and PI double staining. | 5, 10, 20, and 40 µM for 24 h | A dose-dependent increase of apoptotic cells was observed. After incubation with 40 µM, 96% showed early signs of apoptosis. | [33] |
| Piceatannol | U937 cells | DAPI staining | 10, 20, 40, and 60 µM for 48 h | Nuclei with chromatin condensation and the formation of apoptotic bodies were observed in the cells treated with piceatannol in a concentration-dependent manner. | [53] |
| Piceatannol | THP-1 cells | Light microscopy | 10, 20, 30, 40, and 50 for 24 h | Cell shrinkage occurred at concentrations higher than 30 µM. | [34] |
| Piceatannol | HL-60 and HepG2 cells | Hoechst 33258 staining and PI double staining | 50–200 µM for 24–72 h | A time-dependent increase of apoptotic cells was observed, the HL-60 being cells more sensitive. | [24] |
| Piceatannol | OV2008 cells | Hoechst 33258 staining | 10 µM for 24 h | Induction of apoptosis causing nuclear condensation and fragmentation was found. | [25] |
| Pure Stilbene | Experimental Model | Assays Performed | Exposure Conditions | Main Results | Reference |
|---|---|---|---|---|---|
| Pterostilbene | HL-60, HL-60R, K562, K562-ADR, and HUT78 cells | TBET | 1–100 µM for 48 h | Pterostilbene exhibited a similar inhibiting effect and dose response curve in all cell lines. The IC50 values obtained were 35 µM ± 7 in HL-60 cells, 24 µM ± 3 in HUT78, 10 µM ± 3 in K562 cells, 40 µM ± 3 in HL60-R cells, and 12 µM ± 2 in K562-ADR cells. | [36] |
| Pterostilbene (purity > 97%) | RAW 264.7 cells | MTT assay, TBET, and cell proliferation studies with hemocytometer | 0–50 µM for 24 h in MTT and trypan blue assays 10, 20 and 30 µM for 200 h in cell proliferation studies | Maximum inhibition was found from 20 to 30 µM. Cells cultured from > 72 h with < 10 µM were significantly different from the controls. The IC50 for MTT was 8.33 µM ± 0.88 and for TBET was 4.03 µM ± 0.12. | [23] |
| Pterostilbene isolated from Pterocarpus marsupuim | MCF-7 and PC3 cells | MTT assay | 0–100 µM for 24 h | Inhibition of cell growth was clearly observed from 40–80 µM. The IC50 values were 65.6 µM in MCF-7 and 74.3 µM in PC3 cells. | [62] |
| Pterostilbene (purity > 96%) | T24 and T24R cells | MTT assay | 50, 75 and 100 µM for 72 h | Growth decreased in both cell lines in a concentration- and time-dependent manner. The IC50 values for 48 h were 66.58 µM ± 1.84 in T24 cells and 77.95 ± 0.44 µM in T24R cells. | [63] |
| Pterostilbene (purity > 98%) | HepG2 and Chang cells | MTT assay | 3.125, 6.25, 12.5, 25, 50, and 100 µM for 24 h | A concentration-dependent decrease of cell viability in both cell lines was observed. However, no IC50 could be obtained. | [50] |
| Pterostilbene | HCT116, HT-29 and Caco-2 cells | MTT assay | 0–100 µM for 48 h | Cancer cells were more sensitive to pterostilbene than resveratrol, Caco-2 being the least. The IC50 values were 12 µM in HCT116 cells, 15 µM in HT-29 cells, and 75 µM in Caco-2 cells. | [64] |
| Pterostilbene | SK-MEL, KB, BT-549, SK-OV-3, Vero, and LLC-PK11 cells | NRU assay for solid tumor cells and TBET for non-cancerous cells | 0–25 µg/mL for 48 h | Moderate cytotoxicity was observed. No IC50 value was obtained. | [65] |
| Pterostilbene | HT-29 cells | MTT assay | 0–100 µM for 24 h | No cytotoxic effect was observed. The highest concentration tested only reduced cell viability by 20.17% ± 0.82. | [66] |
| Pterostilbene (purity ≥ 90%) | CHO-K1 cells | MTT assay | 20, 40, 60, 80, and 100 µM for 24 h | No cytotoxicity was recorded at lower concentrations; however, the growth inhibitory effect on cells was significant at 100 µM. | [67] |
| Pterostilbene | A375, A549, HT-29, and MCF7 cells | Countess Automated Cell Counter | 0–100 µM for 24, 48, and 72 h | Exposure to pterostilbene reduced tumor cell number in a concentration-, time-, and in a cell line-dependent way. Pterostilbene was more cytotoxic than resveratrol. The IC50 was set at 60.3 µM in HT-29 cells, 44 µM in MCF7 cells, 14.7 µM in A375, and 28.6 µM in A549 cells. | [68] |
| Pterostilbene | K562 cells | MTT assay | 0–100 µM for 48 h | Pterostilbene exhibited significant cytotoxicity while other stilbenes had slight cytotoxic effects. The IC50 value was 67 µM. | [38] |
| Pterostilbene | MOLT4 cells | NRU assay | 0–100 µM for 48 h | A dramatic decrease in cell viability was shown. The estimated concentration required to inhibit cell growth by 90% was 44 µM. | [69] |
| Pterostilbene | MCF-7 andBcap-37 cells | MTT assay | 0–150 µM for 24 h, 48 h, and 72 h | Inhibition of cell proliferation was recorded in a time- and dose-dependent manner. The IC50 values ranged from 50–100 µM for both cell lines and exposure times (24, 48, and 72 h). | [70] |
| Pterostilbene (purity ≥ 99%) | LNCaP, Du145, and PC3M cells | MTT assay | 1–100 µM for 6 days | Growth inhibition was reported for all tested cells. Pterostilbene displayed the highest cytotoxicity among piceatannol, resveratrol and two of its derivatives in PC3M cells. The IC50 values were 22.8 µM in LNCaP cells, 20.8 µM in Du145, and 17 µM in PC3M cells. | [45] |
| Pterostilbene | SOSP-9607 cells | MTT assay | 1, 2 and 4 µM for 12 h, 24 h, and 36 h | Cell growth was inhibited in a dose- and time-dependent manner. The IC50 value at 24 h was 1.81 µM. | [49] |
| Pterostilbene | A431 cells | MTT assay | 15, 30, and 60 µM for 24 h | No cytotoxic effect was recorded. | [71] |
| Pterostilbene (purity ≥ 98%) | A549 and A549 docetaxel resistant cells | MTT assay | 50, 75, and 100 µM for 24 h, 48 h, and 72 h | A significant decrease in the growth of both cell lines in a concentration- and time-dependent manner was reported. | [26] |
| Pterostilbene | MCF-7, T47D, PC-3, NCIH-522, HepG2, PA-1, and LNCaP cells | MTT assay | 0–100 µM for 24 h | Dose-dependent inhibition was found. MCF7, T47D, and HepG2 were more sensitive to pterostilbene. The IC50 was 65 µM ± 0.42 in MCF-7, 69 µM ± 1.58 in T47D, 75 µM ±3.55 in PC-3, 85 µM ± 2.64 in NCIH-522, 73 µM ± 1.81 in HepG2, 120 µM ± 2 in PA-1, and 70.4 µM ± 4.39 in LNCaP cells. | [72] |
| Pterostilbene | Caco-2 cells | SRB assay and LDH activity | 5, 10, 25, 40, 50, 60, 75, and 100 µM for 48 h in both assays and 72 h in SRB assay | Cells exposed to concentrations from 40–100 µM for 48 h exhibited significantly decreased cellular density and an increase in LDH release. At 72 h, all concentrations tested showed significant inhibition of cell proliferation. | [73] |
| Pterostilbene | HeLa cells | MTT assay | 5–160 µM after 24 h, 48 h, and 72 h | Rapid increase in the inhibition rate showing an “S” shape curve. At 80 µM, inhibition was 53.1% after 48 h. | [74] |
| Pterostilbene (purity ≥ 98%) | SAS and OECM-1 cells | MTT assay | 0–40 µM for 24 h and 48 h | Cell viability substantially decreased in a time-dependent manner in both cell lines. | [75] |
| Pterostilbene | Caco-2, HCT116, and CRL-158 cells | SRB assay | 0–1000 µM for 72 h in cancer cells and 40 and 80 µM in CRL-158 cells | Both cells suffered significant inhibition of viability, the non-cancerous cells being the most sensitive. The IC50 values were 31.2 µM ± 0.42 in Caco-2 and 84.4 µM ± 1.14 in HCT116 cells. | [29] |
| Pterostilbene | MCF-7 and MCF-7 CD44+/CD24- cells | TBET | 0–75 µM for 72 h | The effect of pterostilbene was more potent in MCF-7 CD44+/CD24-. The IC50 recorded was 25 µM in MCF-7 CD44+/CD24-. | [76] |
| Pterostilbene | NU-DUL-1, OCI-LY8, U2932, SUDHL-4, DB, and TMD8 cells | CCK-8 assay | 12.5, 25, 50, 75, and 100 µM for 48 h | Cell proliferation was significantly inhibited in a dose-dependent manner but not in a time-dependent way in SUDHL-4, DB, and NU-DUL-1 cells from 12.5 to 100 µM for 24 h, 48 h, and 72 h. | [77] |
| Pterostilbene | H929, ARP-1, OCI-MY5, and RPMI-8226 cells | CCK-8 assay | 10, 20, 30, 40, and 50 µM for 24 h, 48 h, and 72 h | Decrease of cell viability in a dose- and time-dependent manner. The IC50 values obtained for 72 h were 15.37 µM ± 0.98 in H929 cells, 26.15 µM ± 3.6 in ARP-1 cells, 43.36 µM ± 4.46 in OCI-MY5, and 23.58 µM ± 0.41 in RPMI-8226. | [78] |
| Pterostilbene (purity > 97%) | RAW 264.7 cells | MTT assay and TBET | 3, 10, 20, and 30 µM for 24 h and 48 h | Concentration-dependent toxicity was observed. Among all the stilbenes studied, pterostilbene was the most cytotoxic followed by piceatannol and resveratrol. The IC50 values for the MTT assay were 20.7 µM for 24 h and 19 µM for 48 h. Moreover, the IC50 values for TBET were 4 µM for 24 h and 3.6 µM for 48 h. | [79] |
| Pterostilbene | BT-20 and MDA-MB-468 cells | MTT assay | 10, 20, 40, and 80 µM for 48 h | A dose-dependent inhibition of cell proliferation was consistently observed. | [80] |
| Pterostilbene | PC9 and A549 cells | CCK-8 assay | 20, 40, and 60 µM for 24 h and 48 h | Inhibition of cell viability in a dose- and time-dependent manner. The IC50 values were 50.9 µM for 24 h and 27.35 µM for 48 h in PC9 cells, and 52.01 µM for 24 h and 24.12 µM for 48 h in A549 cells. | [81] |
| Pterostilbene | HeLa cells | MTT assay | 5, 25, 50, 100, 200, and 400 µM for 24 h and 48 h | Dose- and time-dependent cytotoxic effects were recorded. The IC50 values were 101.2 µM for 24 h and 65.9 µM for 48 h. | [82] |
| Pterostilbene | Cisplatin-resistant CAL 27 cells | MTT assay | 5, 10, 25, 50, 75, and 100 µM for 24, 48, and 72 h | A time- and concentration-dependent decrease in cell number was reported. The IC50 values were 78.26 µM ± 4.33 for 24 h, 48.04 µM ± 3.68 for 48 h, and 20.65 µM ± 4.88 for 72 h. | [83] |
| Pterostilbene | HeLa cells | WST-1 assay | 10–20 µM for 24 h | A dose-dependent effect was shown. Pterostilbene exhibited higher cytotoxicity than resveratrol at the same concentrations. The IC50 value was 42.3 µM. | [84] |
| Pterostilbene | BV-2 cells | MTT assay | 1, 5, 10, and 20 µM for 24 h | Pterostilbene did not affect the viability of BV-2 cells. | [85] |
| Pterostilbene | Daudi and K562 cells | CCK-8 assay | 0–100 µg/mL for 24 h, 48 h, and 72 h | A time- and dose-dependent decrease in cell viability was observed. The IC50 was 6.87 µM ±1.02 in Daudi cells and 7.05 µM ±1.14 in K562 cells. | [86] |
| Pterostilbene | TC1 cells | WST-1 assay | 5–100 µM for 72 h | Concentration-dependent cytotoxicity was observed. Pterostilbene was more cytotoxic than resveratrol. The IC50 was 15.61 µM | [87] |
| Pterostilbene | NCIH-520 and NCIH-226 cells | MTT assay | 1.56, 3.13, 6.25, 12.5, 25, and 50 µM for 24 h and 48 h | Cytotoxicity was observed for all the cells in a dose-dependent manner. H520 cells were more sensitive than the H226 cells. The IC50 was 47.7 µM ± 5.3 for 24 h and 31.4 µM ± 4.6 for 48 h in H520 cells and >50 µM for 24 h and 44.3 µM ± 3.7 for 48 h in H226 cells. | [88] |
| Pterostilbene | CCD-18-Co, HCT116, SW480, and HT-29 cells | MTT assay | 10, 20, 40, 60, 80, and 100 µM for 24 h | Pterostilbene did not affect the viability of normal colon CCD-18-Co cells, but it reduced the viability of HT-29, SW480, and HCT116 cells. | [89] |
| Pterostilbene (purity > 98%) | HaCat and JB6 cells | MTT assay | 3.75, 7.5, 15, 30 µM and 60 µM for 24 h | Only an exposure of 60 µM decreased cell viability in a significant manner in both cell lines. | [90] |
| Pterostilbene | MIA PaCa-2 and gemcitabine-resistant MIA PaCa-2 cells | MTT assay | 5, 10, 25, 50, and 75 µM for 48 and 72 h | Suppression of cell proliferation in a time- and dose-response manner. Similar EC50 values were obtained after 72 h of exposure for both cell lines (41.8 µM in MIA PaCa-2 cells and 42 µM in gemcitabine-resistant MIA PaCa-2 cells). | [91] |
| Pterostilbene (purity ≥ 98%) | RAW 264.7 and HCEC cells | MTT assay | 2.5, 5, 7.5, 10, 12.5, 15, 17.5, and 20 µM for 24 h | Dose-dependent cytotoxic effects toward both cell lines were shown. No IC50 was reported. | [92] |
| Pterostilbene | A498, ACHN, and HK-2 cells | MTT assay and LDH assay | 5, 10, 20, 50, and 100 µM for 24 h, 48 h, and 72 h in the MTT assay10, 20, and 50 µM for 24 h for LDH | Potent cytotoxic effects were recorded in renal cancer cells but not in non-cancerous renal cells. The cytotoxicity followed a dose- and time-dependent pattern. | [93] |
| Pure Stilbene | Experimental Model | Assays Performed | Exposure Conditions | Main Results | Reference |
|---|---|---|---|---|---|
| Pterostilbene isolated from Pterocarpus marsupuim | MCF-7 cells and PC3 cells | AO and EB by fluorescence microscopy and scanning electron microscopy | MCF-7 cells were exposed to 65.6 µM for 24 h and PC3 cells to 74.3 µM for 24 h | Cells exposed to the stilbene showed chromatin condensation, distorted surface morphology, loss of intercellular attachments and membrane blebbing. | [62] |
| Pterostilbene (purity > 96%) | T24 and T24R cells | DAPI staining, AO, electron microscopy, and phase contrast microscopy | 100 µM for 24 h, 48 h, and 72 h | Cytoplasm and cell surface had granular appearances in exposed cells. Autophagic vacuoles and autolysosomes were also found. Induction of acidic vesicular organelles was also reported. | [63] |
| Pterostilbene | MCF-7 and Bcap-37 cells | DAPI staining and electron microscopy | 50 µM for 24 h | Treated cells showed a foamy appearance and nuclear condensation. Floating cells were also observed. | [70] |
| Pterostilbene | SOSP-9607 cells | Phase contrast microscopy | 1, 2, and 4 µM for 24 h | A decrease in cellular attachment was observed. | [49] |
| Pterostilbene (purity ≥ 98%) | A549 and A549 docetaxel resistant cells | DAPI staining and microscopic observation | 100 µM for 48 h using microscopic observation and 50, 75, and 100 µM for 48 h using DAPI staining | Formation of vacuoles, condensed and fragmented nuclei were found in both cell types after 48 h. | [26] |
| Pterostilbene | MCF-7 cells | Phase contrast microscopy and AO and EB by fluorescence microscopy | 5, 15, 30, and 50 µM for 24 h | Apoptosis induction was evidenced by cell shrinkage and loss of confluence in a dose- dependent manner. Chromatic condensation and loss of cytoplasmic membrane integrity were also found. | [72] |
| Pterostilbene | HeLa cells | Hoechst 33342 staining, AO, and EB by fluorescence microscopy | 80 µM for 48 h | Granular nuclei and nuclear fragmentation were also observed. | [74] |
| Pterostilbene (purity ≥ 98%) | SAS and OECM-1 cells | DAPI staining, AO, and microscopic observation | 0–40 µM for 24 h | Both cell lines treated with the stilbene induced the formation of vacuoles in the cytoplasm, condensed nuclei, and acidic vesicular organelles in a concentration- and time-dependent manner. | [75] |
| Pterostilbene | MCF-7 and MCF-7 CD44 +/CD24- cells | Microscopy observation (400x) | 100 µM for 6 h | Cell necrosis, such as membrane injury and bleb formation, were found. | [76] |
| Pterostilbene | PC9 and A549 cells | Phase contrast microscopy | 20, 40, and 60 µM for 24 h | Significant cell shrinkage and a decreased cellular attachment rate were reported. | [81] |
| Pterostilbene | HeLa cells | AO and EB by fluorescence microscopy | 25, 100, and 200 µM for 48 h | Pterostilbene-treated cells showed cellular shrinkage, detached from one another and from the substratum, membrane blebbing, nuclear fragmentation, and chromatin condensation. | [82] |
| Pterostilbene | Cisplatin-resistant CAL 27 cells | AO, Monodans yleadaverine LysoTracker Red, Cathepsin B, Hoechst 33342, and phase contrast microscopy | 5, 10, 25, 50, 75, and 100 µM for 24 h, 48 h, and 72 h for phase contrast microscopy 25, 50, and 75 µM for 24 h for the other tests | An increased number of acidic vesicular organelles, accumulation of the autophagic vacuole marker and suppressed lysosome activity were observed. Moreover, DNA condensation was shown to occur at 25, 50, and 75 µM. | [83] |
| Pterostilbene | HeLa cells | Brightfield analysis | 40 µM for 24 h | Apoptotic blebbing was observed. Pterostilbene reduced cell numbers more markedly than resveratrol when cells were exposed to the same concentration. | [84] |
| Pterostilbene | TC1 cells | Brightfield analysis | 5–100 µM for 72 h | Cytoplasmic blebbing was reported after 48 h at <10 µM. | [87] |
| Pterostilbene | HCT116, SW480, and HT29 cells | DAPI staining and phase contrast microscopy | 40 µM for 24 h | Apoptotic bodies, DNA fragmentation and cell shrinkage | [89] |
| Pterostilbene | NCIH-520 and NCIH-226 cells | Brightfield analysis | 1.56, 3.13, 6.25, 12.5, 25 and 50 µM for 24 h and 48 h | Apoptotic morphological changes, cell shrinkage and cytoplasmic blebbing. H520 cells were more sensitive than NCIH-226 cells. | [88] |
| Pterostilbene | A498 and ACHN cells | Phase contrast microscopy | 10, 20 and 50 µM for 24 h and 48 h | A decreased number of cells and a reduction in cell-cell contact were observed in cancer cells in a dose-dependent manner. | [92] |
| Pure Stilbene | Experimental Model | Assays Performed | Exposure Conditions | Main Results | Reference |
|---|---|---|---|---|---|
| Trans-ε-viniferin and cis-ε-viniferin (from Paeonia lactiflora) | C6, HepG2, HeLa, MCF-7, and HT-29 cells | MTT assay | 0–100 µM for 70 h | Cytotoxicity was reported in all cell lines tested, although trans- and cis- ε-viniferin were markedly cytotoxic in C6 and HeLa cells. The IC50 values for trans-ε-viniferin were: 18.4 µM in C6 cells, 74.3 µM in HepG2 cells, 20.4 µM in HeLa cells, 44.8 µM in MCF-7 cells, and 88.4 µM in HT-29 cells. The IC50 values for cis-ε-viniferin were: 20.1 µM in C6 cells, 76.2 µM in HepG2 cells, 21.5 µM in HeLa cells, and 47.2µM in MCF-7 cells, and 90.2 µM in HT-29 cells. | [94] |
| ε-viniferin (from vine-shoots) | WSU-CLL cells | TBET | 0–100 µM for 24, 48, and 72 h | A concentration- and time-dependent decrease in cell viability was observed, ε-viniferin˂ resveratrol. The inhibition of cell multiplication was paralleled by a decrease in DNA synthesis. The IC50 at 72 h was 60 µM. | [95] |
| ε-viniferin | HL-60 cells | MTT assay | 10–200 µM for 24 h | Cell viability decreased in a dose-dependent manner. The IC50 was 33 µM. | [96] |
| (-)-ε-viniferin | HCF, HPC, HPLF, HSC-2, HSC-3, HSG, and HL-60 cells | MTT assay in adherent cells and TBET in non-adherent cells | 0–1000 µM for 24 h | The four tumor cell lines (HSC-2, HSC-3, HSG, and HL-60) were more sensitive to the stilbene than the three normal cells (HCF, HPC, and HPLF). The CC50 values were 111 µM for HCF cells, 146 µM for HPC cells, 94 µM for HPLF cells, 42µM for HSC-2 cells, 84 µM for HSC-3 cells, 110 µM for HSG cells, and 31 µM for HL-60 cells. | [32] |
| (-)-ε-viniferin | P-388 cells | MTT assay | 0–100 µM for 48 h | ε-viniferin moderately inhibited the cells in comparison to hopeaphenol which exhibited a greater effect. The IC50 found was 18.1 µM ± 0.7. | [97] |
| ε-viniferin | HepG2 cells | TBET | 30 µM for 24, 48 and 72 h. 1, 5, 10, 30, 60, and 100 µM for 48 h | At 60 µM, ε-viniferin completely inhibits cell proliferation. After 48 h, the toxicity potential of ε-viniferin was lower than resveratrol. The IC50 for 48 h was 58.4 µM. | [98] |
| ε-viniferin | SW480 cells | TBET (Coulter Counter) and MTT assay | 30 µM for 24, 48, 72, and 96 h in trypan blue assay. 3, 30, 60, and 100 µM for 48 h in coulter counter | Cells exposed to the stilbene grew similarly to the control. Reduced growth rate and percentage of cell inhibition. In the MTT assay, no inhibition of cell proliferation was recorded. | [99] |
| Trans-ε-viniferin (from Vitis amurensis) | L1210, K562, and HCT116 cells | MTT assay | 0–50 µM for 48 h | No cytotoxic activity was recorded. Therefore, the IC50 was assumed to be above 50 µM. | [37] |
| ε-viniferin glucoside | PC12 cells | MTT assay | 0–10 µM for 24 h | Cell viability was not significantly affected in any exposure to the stilbene. | [100] |
| (-)-ε-viniferin | HepG2 and Chang cells | MTT assay | 1.56–200 µg/mL for 72 h | No cytotoxic effect was found in either cell. | [101] |
| ε-viniferin | VSMCs | MTS assay | 10, 20, and 30 µM for 48 h | The antiproliferative rate of ε-viniferin at 20 µM was significantly higher than that of resveratrol at both 20 and 30 µM. | [30] |
| (+)-ε-viniferin (from Ampelopsis brevipedunculata) | RAW264.7 cells | MTT assay | 1, 5, and 10 µM for 12 h | (+)-ε-viniferin dramatically reduced cell viability to 60% after the exposition of 10 µM. No IC50 was obtained. | [102] |
| ε-viniferin | HT-144 and SK-MEL-25 cells | MTT assay and TBET | 25–200 µM for 24, 48, and 72 h | A decrease in cell survival in a time- and dose-dependent manner was observed in both cell lines. The IC50 for 48 h was 60 µM. | [103] |
| ε-viniferin | C6 cells | WST-1 assay | 95 and 130 µM for 12, 24, and 48 h | A decrease in cell proliferation was reported. This reduction was significant at all concentrations and times tested. | [104] |
| Trans-ε-viniferin | MRC-5, AGS, SK-MES-1, and J82 cells | MTT assay | 0–100 µg/mL for 72 h | Cytotoxic effects were observed in all tested cell lines. The IC50 values were 49.9 µM ± 3 in MRC-5 cells, 42.6 µM ± 1.7 in AGS cells, 78.8 µM ± 3.3 in SK-MES-1 cells, and 56.7 µM ± 1.2 in J82 cells. | [46] |
| Trans-ε-viniferin | Mouse primary co-culture of astrocytes and neurons | CellTiter 96 ® Aqueous | 1, 5, 10, 20, 50, and 100 µM for 72 h | A significant decrease in cell viability was observed at exposures of 50 and 100 µM. | [105] |
| ε-viniferin | Caco-2 cells | MTT and NRU assays | 1.56, 3.12, 6.25, 12.5, 25, 50, and 100 µM for 24 h | A dose-dependent decrease in cell viability was observed at 25 µM. ε-viniferin was slightly more toxic than resveratrol in Caco-2 cells. | [106] |
| Trans-ε-viniferin | COLO 205, HT-29, HepG2, AGS, and HL-60 cells | MTT assay | 0–100 µg/mL for 48 h | Dose-dependent cytotoxicity was reported, with a potent effect observed in HL-60 cells. The IC50 values were: 85.5 µM ± 8.1 in COLO205 cells, 13.9 µM ± 0.1 in HT-29 cells, 7.7 µM ± 0.2 in HepG2 cells, 9.3 µM ± 0.3 in AGS cells, and 5.6 µM ± 1.4 in HL-60 cells. | [93] |
| Trans-ε-viniferin (from Vitis vinifera) | HepG2, Hep3B, and HH4 cells | Crystal violet assay | 0–200 µM for 24, 48, and 72 h | Cell number decreased in a dose- and time-dependent manner, being more cytotoxic in Hep3B cells. In HH4 cells, higher concentrations were needed to induce toxicity. The IC50 values obtained were the following: - HepG2 cells: 140 µM ± 39.7 (24 h), 103.8 µM ± 19.2 (48 h), 94.8 µM ± 28.3 (72 h) - Hep3B cells: 108.1 µM ± 31.8 (24 h), 73.9 µM ± 17.3 (48 h), 63.1 µM ± 10.8 (72 h) - HH4 cells: >200 µM (24 h), 192.7 µM ± 21.1 (48 h), 177.9 µM ± 20.5 (72 h). | [107] |
| Trans-ε-viniferin | HepG2 and Caco-2 cells | MTS assay, NRU, and protein content | 0–100 μg/mL for 24 and 48 h | Both cell lines exposed to ε-viniferin exhibited a time-dependent decrease for all the endpoints studied The EC50 values were: HepG2: 28.28 ± 2.15 24 h and 17.85 ± 3.03 for 48 h. Caco-2 cells: 36.72 ± 3.01 for 24 h and 20.63 ± 1.25 48 h. | [108] |
| Pure Stilbene | Experimental Model | Assays Performed | Exposure Conditions | Main Results | Reference |
|---|---|---|---|---|---|
| ε-viniferin | HL-60 cells | Hoechst 33342 staining | 100 µM for 24 h | The cell line suffered chromatin condensation, nuclear fragmentation and cell collapse into apoptotic bodies. | [96] |
| ε-viniferin | SW480 cells | Hoechst 33342 staining | 30 µM for 48 h | No apoptotic changes were observed. | [99] |
| ε-viniferin | HT-144 and SK-MEL-25 cells | Hoechst 33342 staining | 60 µM for 48 and 72 h | Increase in nucleus size, condensation, and fragmentation of nuclear chromatin in SKMel-28 cells, but not in HT144 cells. | [103] |
| ε-viniferin | C6 cells | Hoechst 33342 staining | 95 and 130 µM for 48 h | Condensation of chromatin structure and slight contraction in the nuclear membrane. | [104] |
| Pure Stilbene | Experimental Model | Assays Performed | Exposure Conditions | Main Results | Reference |
|---|---|---|---|---|---|
| Piceatannol | BJAB cells | Flow cytometric determination of hypodiploid DNA | 0, 25, 50, 75, 100, and 125 µM for 4 h | Dose-dependent DNA fragmentation was observed. | [51] |
| Piceatannol | Human peripheral lymphocytes | Comet assay | 0, 10, 20, and 50 µM for 30 min in the presence of Cu (II) | DNA breakage was detected. The effect exerted by piceatannol was more potent than that of resveratrol and trans-stilbene. | [109] |
| Piceatannol | HL-60 and HSC-2 cell lines | Electrophoresis | 10, 20, and 40 µM to HL-60 or 320 µM to HSC-2 for 6 or 4 h, respectively | Piceatannol induced DNA fragmentation in a dose-dependent manner. | [32] |
| Piceatannol | L1210, K562, and HL-60 cell lines | Comet assay | Pretreatment of 0.625, 1, 2.5, or 5 µM for 24 h | A decrease in DNA damage was produced by H2O2 after piceatannol exposure. | [35] |
| Piceatannol | Salmonella typhimurium TA102 strain | Ames test | 50 µg/plate 200, 100, and 50 µg/plate | No mutagenicity was reported in the presence and absence of rat liver S9 mix. The stilbene exhibited antimutagenic activity in the presence of rat liver S9 mix. Moreover, it moderately inhibited the effect of the mutations produced by benzidine. | [110] |
| Piceatannol | U937 cells | Electrophoresis | 0, 10, 20, 40, or 60 µM for 48 h | Dose-dependent DNA fragmentation was detected after piceatannol exposure. | [53] |
| Piceatannol | THP-1 cells | Electrophoresis | 10 µM for 24 h | Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). DNA breakage was reported only in the presence of 20 ng/mL. | [34] |
| Piceatannol | Du145 cells | Western blot | 0, 10, and 25 µM for 72 h | Piceatannol caused DNA damage supported by increased phosphorylated histone H2AX. | [111] |
| Piceatannol | A549, HepG2, and MCF7 cells lines | Electrophoresis | 20, 40, or 60 µg/mL of piceatannol encapsulated with nanoparticles (chitosan and poly lactic acid) for 24 h | DNA fragmentation was observed in all cell lines. In A549 and HepG2 cell lines, the effect was dose-dependent. | [112] |
| Piceatannol | THP-1, HL-60, U937 and K562 cell lines | Electrophoresis | 25 or 50 µM for 24 h | After the piceatannol exposure, DNA breaks were observed in all cell lines tested. | [113] |
| Piceatannol | MOLT-4 cells | Electrophoresis | 45.5 µM for 12, 24, and 48 h | DNA fragmentation was detected, with the most potent effect observed after 12 h of exposure. | [60] |
| Pterostilbene | MCF-7 and PC3 cell lines | Electrophoresis Comet assay | 65.6 µM in MCF-7 and 74.3 µM in PC3 for 24 h | DNA damage was observed with both techniques. The comet assay indicated that MCF-7 cells were more sensitive than the PC3 cell line. | [62] |
| Pterostilbene | MOLT4 cells | Electrophoresis | 44 µM for 12, 24, and 48 h | Pterostilbene induced DNA breakage at all exposure times. | [69] |
| Pterostilbene | CHO-K1 cells | Standard and modified alkaline comet assay. Cytokinesis block micronucleus assay | 40 and 80 µM for 3 h 40 and 80 µM for 3 h plus 20 min with H2O2 20, 40, and 80 µM for 3 h 20, 40, and 80 µM for 3 h plus 20 min with H2O2 | In non-treated cells, basal damage decreased in a dose-dependent manner. At 80 µM, pterostilbene exhibited a protective effect, reducing the DNA oxidative damage more significantly than trimethoxystilbene and resveratrol. No increase in micronuclei was observed. The stilbene did not protect against H2O2 oxidative damage. | [114] |
| Pterostilbene Pterostilbene isothiocyanate conjugate | MCF-7 cells | Electrophoresis | 20 µM for 24 h 60 µM for 24 h | DNA breakage was reported. The conjugate produced DNA fragmentation. | [72] |
| Pterostilbene | HeLa cells | Electrophoresis | 80 and 120 µM for 24 h | Pterostilbene induced DNA damage. | [74] |
| Pterostilbene | HepG2 cells | Comet assay | 50 µM plus 1 µM of 4NQO for 4 h in co-exposure Pre-exposure at 50 µM for 4 h, and plus 1 µM of 4NQO for 4 h | In both assays performed, pterostilbene exhibited antimutagenic effects. | [67] |
| Pterostilbene | H929 cells | Western blot | 10, 20, and 40 µM for 24 h | DNA breakage was indicated by the increase in the marker ƴ-H2AX. | [69] |
| Pterostilbene | HeLa cells | Electrophoresis | 25, 10, or 200 µM for 48 h | After pterostilbene exposure, DNA damage was detected in a dose-dependent manner. | [82] |
| Pterostilbene | CAR cells | TUNEL assay | 50, 75, and 100 µM for 48 h | The TUNEL assays indicated that DNA fragmentation was induced by pterostilbene. | [83] |
| Pterostilbene | ACHN and A498 cell lines | Immunofluorescence analysis Western blot | 10, 20, and 50 µM for 24 h | Pterostilbene increased ƴ-H2AX, indicating DNA damage. | [92] |
| ε-viniferin | Salmonella typhimurium TA100 strain | Ames test | 35.2 µg/plate | The stilbene exhibited an antimutagenic effect in a dose-dependent manner against the mutations produced by N-methyl-N-nitro-N-nitrosoguanidine. | [94] |
| ε-viniferin contained in grapevine-shoot extract | A431 cells | Standard and modified alkaline Comet assay | ≥25 µg/mL for 1 h | An increase in DNA strand breaks was reported. | [115] |
| ε-viniferin | C6 cells | TUNEL assay | 95 and 130 µM for 12, 24, and 48 h | ε-viniferin produced DNA damage in a dose-dependent manner. | [104] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medrano-Padial, C.; Prieto, A.I.; Puerto, M.; Pichardo, S. Toxicological Evaluation of Piceatannol, Pterostilbene, and ε-Viniferin for Their Potential Use in the Food Industry: A Review. Foods 2021, 10, 592. https://doi.org/10.3390/foods10030592
Medrano-Padial C, Prieto AI, Puerto M, Pichardo S. Toxicological Evaluation of Piceatannol, Pterostilbene, and ε-Viniferin for Their Potential Use in the Food Industry: A Review. Foods. 2021; 10(3):592. https://doi.org/10.3390/foods10030592
Chicago/Turabian StyleMedrano-Padial, Concepción, Ana Isabel Prieto, María Puerto, and Silvia Pichardo. 2021. "Toxicological Evaluation of Piceatannol, Pterostilbene, and ε-Viniferin for Their Potential Use in the Food Industry: A Review" Foods 10, no. 3: 592. https://doi.org/10.3390/foods10030592