
Vulnerability of the Small-Scale Fishery to Climate Changes in the Northern-Central Adriatic Sea (Mediterranean Sea)
 , , ,
, , ,
Abstract
:1. Introduction
- (1)
- In the near future, would the Northern-Central Adriatic Sea be subjected to a meridionalization trend (sensu [41])?
- (2)
- Based on habitat suitability analysis, will SSF catch composition be affected by climate change and, if so, could warm-affinity species be considered a valid alternative option for the SSF in the area?
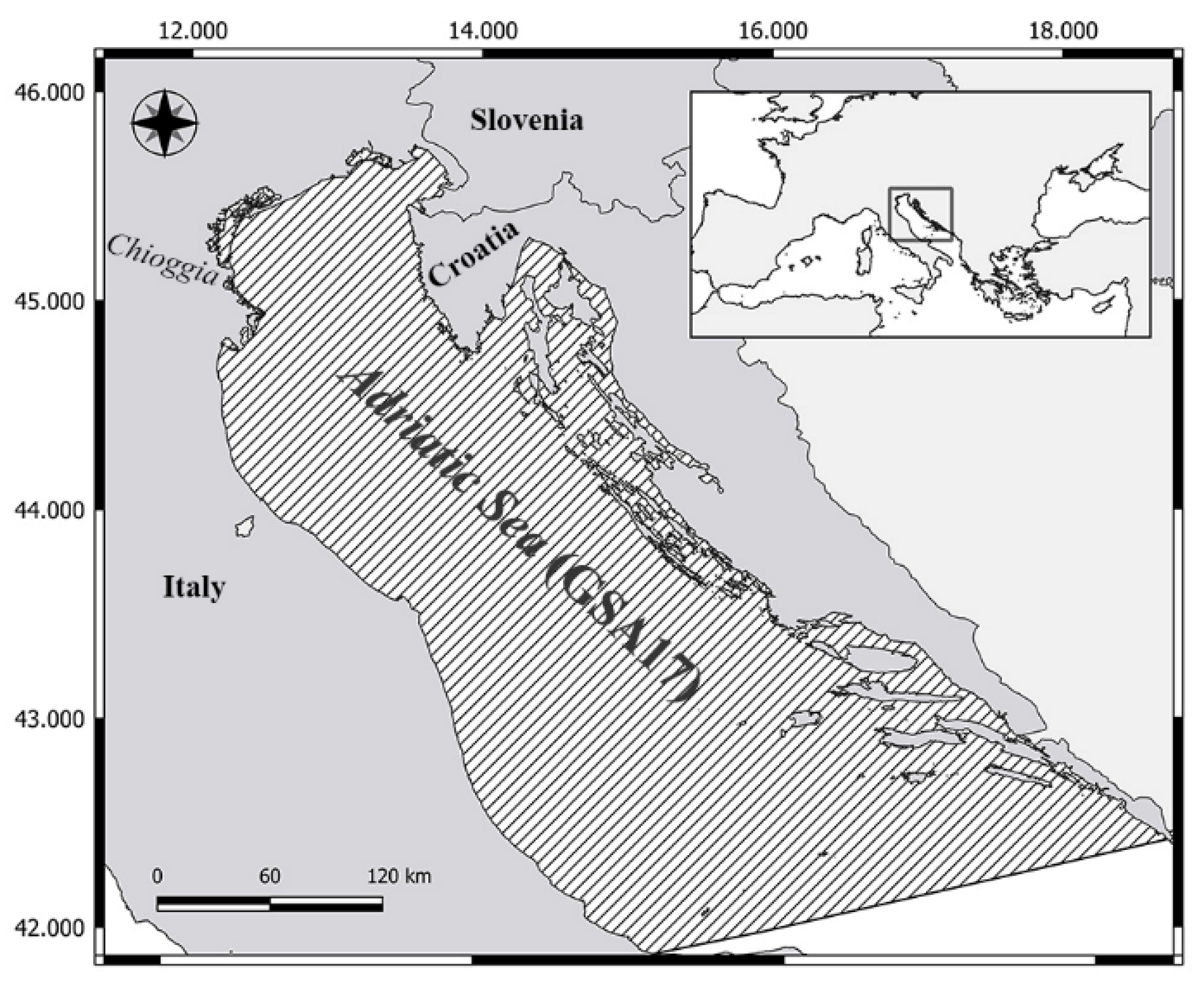
2. Materials and Method
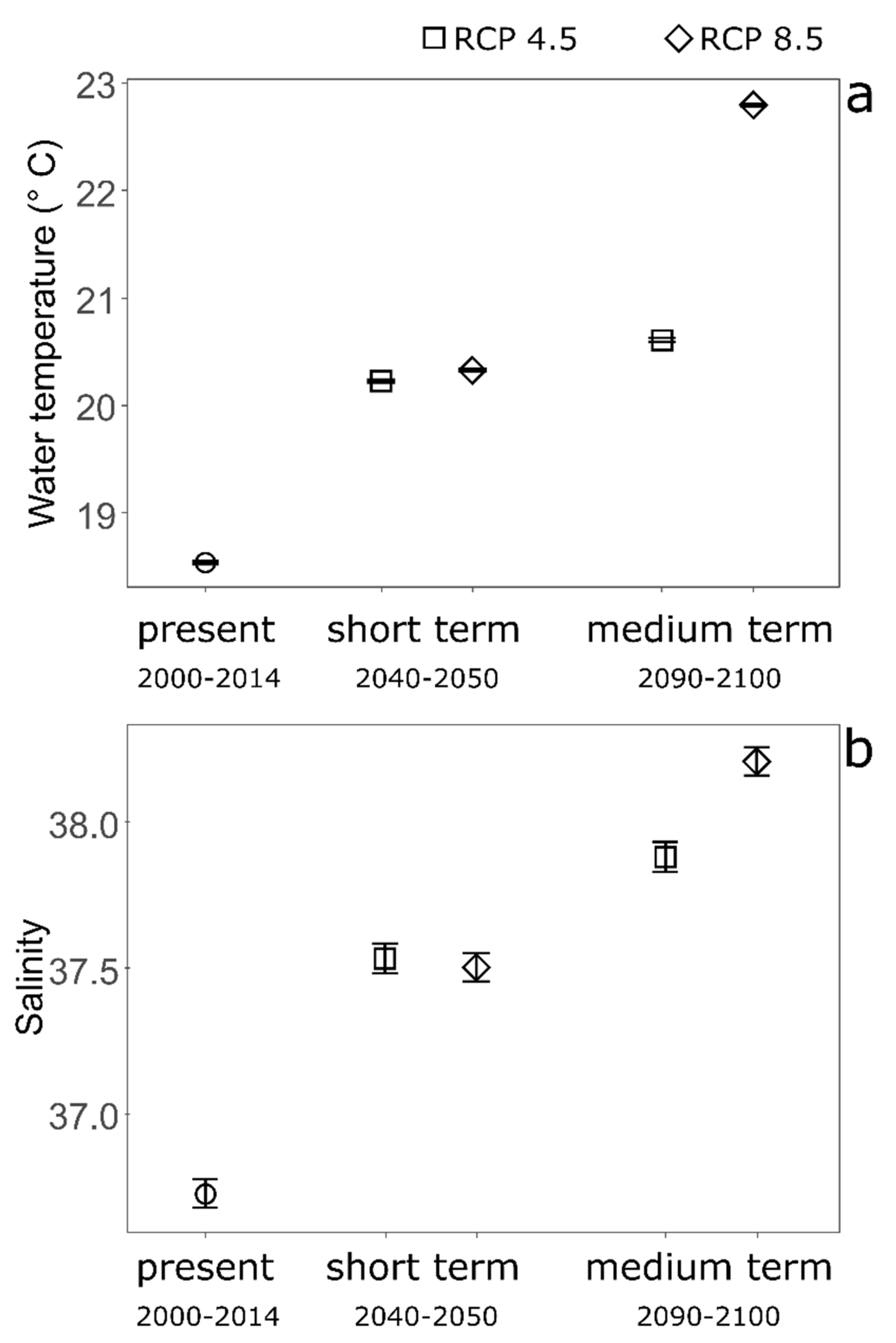
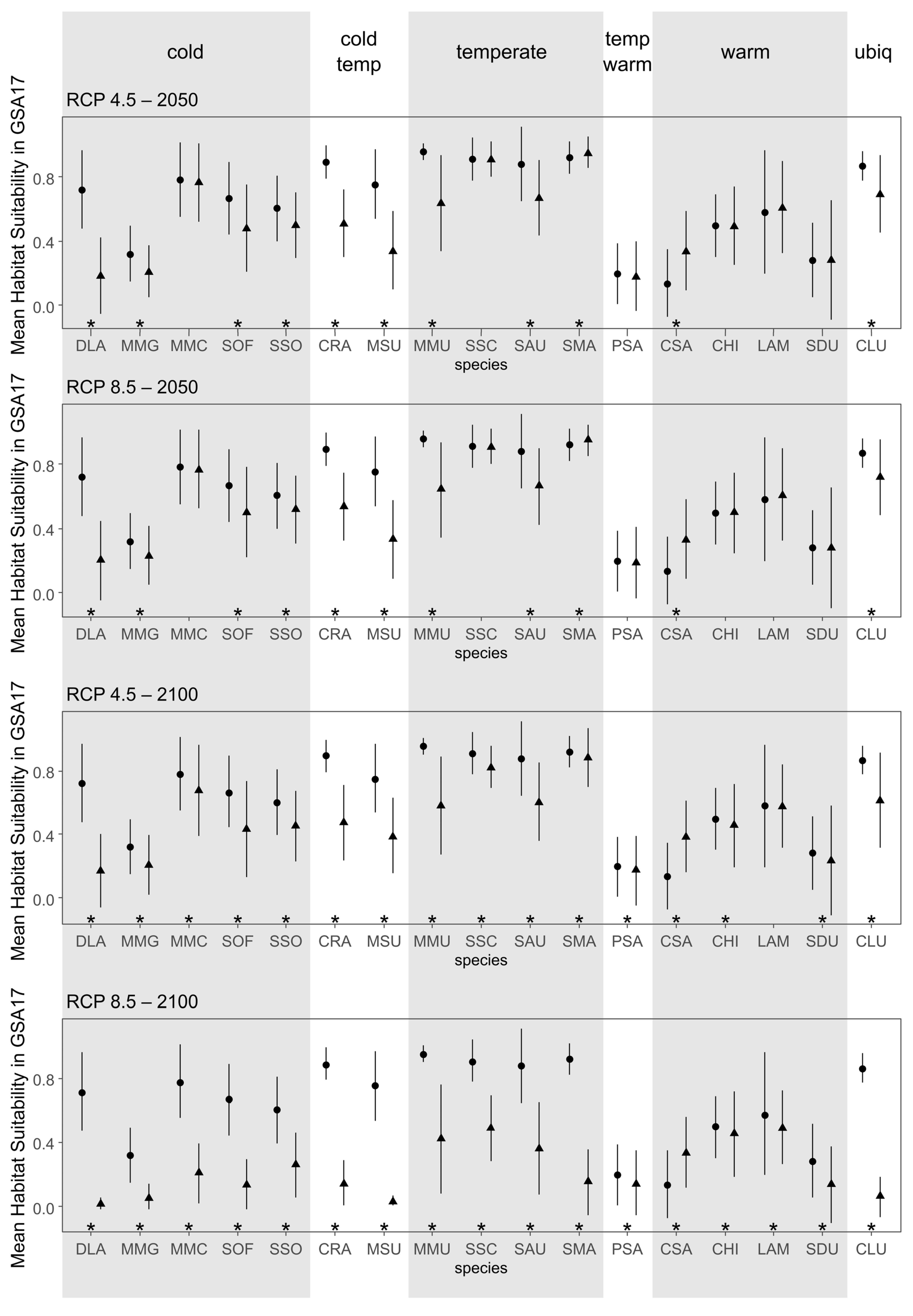
3. Results
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Barange, M.; Bahri, T.; Beveridge, M.C.M.; Cochrane, K.L.; Funge-Smith, S.; Poulain, F. Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options; FAO Fisheries and Aquaculture Technical Paper; FAO: Rome, Italy, 2018; 628p. [Google Scholar]
- Holsman, K.K.; Hazen, E.L.; Haynie, A.; Gourguet, S.; Hollowed, A.; Bograd, S.J.; Samhouri, J.F.; Aydin, K. Towards climate resiliency in fisheries management. ICES J. Mar. Sci. 2019, 76, 1368–1378. [Google Scholar] [CrossRef]
- Pinsky, M.L.; Fenichel, E.; Fogarty, M.; Levin, S.; McCay, B.; Martin KSt Selden, R.L.; Young, T. Fish and fisheries in hot water: What is happening and how do we adapt? Pop. Ecol. 2019, 63, 17–26. [Google Scholar] [CrossRef]
- Barange, M.; Cheung, W.W.L.; Merino, G.; Perry, R.I. Modelling the potential impacts of climate change and human activities on the sustainability of marine resources. Curr. Opin. Environ. Sustain. 2010, 2, 326–333. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Magnan, A.K.; Billé, R.; Cheung, W.W.L.; Howes, E.L.; Joos, F.; Allemand, D.; Bopp, L.; Cooley, S.R.; Eakin, C.M.; et al. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 2015, 349, 6243. [Google Scholar] [CrossRef]
- Hollowed, A.B.; Barange, M.; Beamish, R.J.; Brander, K.; Cochrane, K.L.; Drinkwater, K.F.; Foreman, M.G.G.; Hare, J.A.; Holt, J.; Ito, S.-I.; et al. Projected impacts of climate change on marine fish and fisheries. ICES J. Mar. Sci. 2013, 70, 1023–1037. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; p. 1132. [Google Scholar]
- Poloczanska, E.S.; Brown, C.J.; Sydeman, W.J.; Kiessling, W.; Schoeman, D.S.; Moore, P.J.; Brander, K.; Bruno, J.F.; Buckley, L.B.; Burrows, M.T.; et al. Global imprint of climate change on marine life. Nat. Clim. Chang. 2013, 3, 919–925. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Coll, M.; Piroddi, C.; Steenbeek, J.; Lasram, F.B.R.; Zenetos, A.; Cardoso, A.C. Invading the Mediterranean Sea: Biodiversity patterns shaped by human activities. Front. Mar. Sci. 2014, 1, 32. [Google Scholar] [CrossRef] [Green Version]
- Adloff, F.; Somot, S.; Sevault, F.; Jorda, G.; Aznar, R.; Déqué, M.; Herrmann, M.; Marcos, M.; Dubois, C.; Padorno, E.; et al. Mediterranean Sea response to climate change in an ensemble of twenty first century scenarios. Clim. Dyn. 2015, 45, 2775–2802. [Google Scholar] [CrossRef]
- Marbà, N.; Jordà, G.; Agusti, S.; Girard, C.; Duarte, C.M. Footprints of climate change on Mediterranean Sea biota. Front. Mar. Sci. 2015, 2, 56. [Google Scholar] [CrossRef]
- Boero, F.; Féral, J.P.; Azzurro, E.; Cardin, V.; Riedel, B.; Despalatovic, M.; Munda, I.; Moschella, P.; Zaouali, J.; Fonda Umani, S.; et al. Executive Summary of CIESM workshop climate warming and related changes in Mediterranean marine biota. CIESM Workshop Monogr. 2008, 35, 5–21. [Google Scholar]
- Edelist, D.; Rilov, G.; Golani, D.; Carlton, J.T.; Spanier, E. Restructuring the Sea: Profound shifts in the world’s most invaded marine ecosystem. Divers. Distrib. 2013, 19, 69–77. [Google Scholar] [CrossRef]
- Kovačić, M.; Lipej, L.; Dulčić, J.; Samuel, I.; Menachem, G. Evidence-based checklist of the Mediterranean Sea fishes. Zootaxa 2021, 4988, 7. [Google Scholar] [CrossRef]
- Lasram, F.; Guilhaumon, F.; Albouy, C.; Somot, S.; Thuiller, W.; Mouillo, D. The Mediterranean Sea as a ‘cul-de-sac’ for endemic fishes facing climate change. Glob. Chang. Biol. 2010, 16, 3233–3245. [Google Scholar] [CrossRef]
- Fortibuoni, T.; Libralato, S.; Raicevich, S.; Giovanardi, O.; Solidoro, C. Coding early naturalists’ accounts into long-term fish community changes in the Adriatic Sea (1800e2000). PLoS ONE 2010, 5, e15502. [Google Scholar] [CrossRef]
- Lotze, K.H.; Coll, M.; Dunne, J.A. Historical changes in marine resources, foodweb structure and ecosystem functioning in the Adriatic Sea, Mediterranean. Ecosystems 2011, 14, 198–222. [Google Scholar] [CrossRef]
- Coll, M.; Santojanni, A.; Palomera, I.; Arneri, E. Ecosystem assessment of the north-Central Adriatic Sea: Towards a multivariate reference framework. Mar. Ecol. Prog. Ser. 2010, 417, 93–210. [Google Scholar] [CrossRef]
- Ferretti, F.; Myers, R.A.; Serena, F.; Lotze, H.K. Loss of large predatory sharks from the Mediterranean Sea. Conserv. Biol. 2008, 22, 952–964. [Google Scholar] [CrossRef]
- Grbec, B.; Dulčić, J.; Morović, M. Long-term changes in landings of small pelagic fish in the eastern Adriatic and possible influence of climate oscillations over the Northern Hemisphere. Clim. Res. 2002, 20, 241–252. [Google Scholar] [CrossRef]
- Santojanni, A.; Arneri, E.; Bernardini, V.; Cingolani, N.; Di Marco, M.; Russo, A. Effects of environmental variables on recruitment of anchovy in the Adriatic Sea. Clim. Res. 2006, 31, 181–193. [Google Scholar] [CrossRef]
- Russo, A.; Rabitti, S.; Bastianini, M. Decadal climatic anomalies in the Northern Adriatic Sea inferred from a new oceanographic data set. Mar. Ecol. 2002, 23, 340–351. [Google Scholar] [CrossRef]
- Giani, M.; Djakovac, T.; Degobbis, D.; Cozzi, S.; Solidoro, C.; Fonda Umani, S. Recent changes in the marine ecosystems of the Northern Adriatic Sea. Est. Coast. Shelf Sci. 2012, 115, 1–13. [Google Scholar] [CrossRef]
- Zampieri, M.; Giorgi, F.; Lionello, P.; Nikulin, G. Regional climate change in the Northern Adriatic. Phys. Chem. Earth 2012, 40–41, 32–46. [Google Scholar] [CrossRef]
- Lionello, P. The climate of the Venetian and North Adriatic region: Variability, trends and future change. Phys. Chem. Earth 2012, 40–41, 1–8. [Google Scholar] [CrossRef]
- Azzurro, E.; Moschella, P.; Maynou, F. Tracking signal of change in Mediterranean fish diversity based on local ecological knowledge. PLoS ONE 2011, 6, e24885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulčić, J.; Pallaoro, A. First record of the marbled spinefoot Siganus rivulatus (Pisces: Siganidae) in the Adriatic Sea. J. Mar. Biol. Assoc. U. K. 2004, 84, 1087–1088. [Google Scholar] [CrossRef]
- Lloret, J.; Cowx, I.G.; Cabral, H.; Castro, M.; Font, T.; Gonçalves, J.M.; Gordoa, A.; Hoefnagel, E.; Matić-Skoko, S.; Mikkelsen, E.; et al. Small-scale coastal fisheries in European Seas are not what they were: Ecological, social and economic changes. Mar. Policy 2018, 98, 176–186. [Google Scholar] [CrossRef]
- Di Martino, V.; Stancanelli, B. The alien lionfish, Pterois miles (Bennett, 1828), enters the Adriatic Sea, Central Mediterranean Sea. J. Black Sea Mediterr. Environ. 2021, 27, 104–108. [Google Scholar]
- Tiralongo, F.; Crocetta, F.; Riginella, E.; Lillo, A.O.; Tondo, E.; Macali, A.; Mancini, E.; Russo, F.; Coco, S.; Paolillo, G.; et al. Snapshot of rare, exotic and overlooked fish species in the Italian seas: A citizen science survey. J. Sea Res. 2020, 164, 101930. [Google Scholar] [CrossRef]
- Coco, S.; Roncarati, A.; Tiralongo, F.; Felici, A. Meridionalization as a Possible Resource for Fisheries: The Case Study of Caranx rhonchus Geoffroy Saint-Hilaire, 1817, in Southern Italian Waters. J. Mar. Sci. Eng. 2022, 10, 274. [Google Scholar] [CrossRef]
- Hidalgo, M.; Mihneva, V.; Vasconcellos, M.; Bernal, M. Climate change impacts vulnerabilities adaptations: Mediterranean Sea the Black Sea marine fisheries. In Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options; Barange, M., Bahri, T., Beveridge, M.C., Cochrane, K.L., Funge-Smith, S., Poulain, F., Eds.; FAO: Kanagawa, Japan, 2018; 630p. [Google Scholar]
- Grati, F.; Aladzuz, A.; Azzurro, E.; Bolognini, L.; Carbonara, P.; Ҫobani, M.; Domenichetti, F.; Dragicevic, B.; Dulcic, J.; Ðurovic, M.; et al. Seasonal dynamics of small-scale fisheries in the Adriatic Sea. Med. Mar. Sci. 2018, 19, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Bradley, B.A.; Wilcove, D.S.; Oppenheimer, M. Climate change increases risk of plant invasion in the Eastern United States. Biol. Invasions 2010, 12, 1855–1872. [Google Scholar] [CrossRef]
- Fulton, E.A.; Bax, N.; Bustamante, R.H.; Dambacher, J.M.; Dichmont, C.M.; Dunstan, P.; Hayes, K.; Hobday, A.J.; Pitcher, R.; Plagányi, E.E.; et al. Modelling marine protected areas: Insights and hurdles. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140278. [Google Scholar] [CrossRef]
- Coro, G.; Vilas, L.G.; Magliozzi, C.; Ellenbroek, A.; Scarponi, P.; Pagano, P.F. Forecasting the ongoing invasion of Lagocephalus sceleratus in the Mediterranean Sea. Ecol. Model. 2018, 371, 37–49. [Google Scholar] [CrossRef]
- Marras, S.; Cucco, A.; Antognarelli, F.; Azzurro, E.; Milazzo, M.; Bariche, M.; Butenschön, M.; Kay, S.; Di Bitetto, M.; Quattrocchi, G.; et al. Predicting future thermal habitat suitability of competing native and invasive fish species: From metabolic scope to oceanographic modelling. Conserv. Physiol. 2015, 3, cou059. [Google Scholar] [CrossRef] [Green Version]
- Melo-Merino, S.M.; Reyes-Bonilla, H.; Lira-Noriega, A. Ecological niche models and species distribution models in marine environments: A literature review and spatial analysis of evidence. Ecol. Model. 2020, 415, 108837. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Cinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of marine invasive alien species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Giorgi, F. Climate change hotspot. Geophys. Res. Lett. 2006, 33, L08707. [Google Scholar] [CrossRef]
- Azzurro, E. The advance of thermophilic fishes in the Mediterranean Sea: Overview and methodological questions. In Climate Warming and Related Changes in Mediterranean Marine Biota. N° 35 in CIESM Workshop Monographs; Briand, F., Ed.; CIESM: Monte Carlo, Monaco, 2008; 152p. [Google Scholar]
- Béguinot, A. La Vita delle Piante Superiori Nella Laguna di Venezia; Ferrari, C., Ed.; Venezia, Italy, 1913; 348p. [Google Scholar]
- Marcello, A. Lacuna floristica del veneziano e sue condizioni bioclimatiche. Mem. Biogeogr. Adriat. 1960, 5, 51–118. [Google Scholar]
- Kourafalou, V.H. River plume development in semi-enclosed Mediterranean regions: North Adriatic Sea and northwestern Aegean Sea. J. Mar. Syst. 2001, 30, 181–205. [Google Scholar] [CrossRef]
- Sacchi, C.F. Il delta del Po come elemento disgiuntore nell’ecologia delle spiagge adriatiche—Considerazioni generali e conferme malacofaunistiche. Boll. Mus. Civ. Stor. Nat. Venezia 1978, 29, 43–73. [Google Scholar]
- Dulčić, J.; Soldo, A.; Jardas, I. Review of Croatian selected scientific literature on species mostly exploited by the national small-scale fisheries. AdriaMed. Adriat. Sea Small-Scale Fisheries. AdriaMed Tech. Consult. Adriat. Sea Small-Scale Fish 2005, 134–179. [Google Scholar]
- Grati, F.; Azzurro, E.; Scanu, M.; Tassetti, A.N.; Bolognini, L.; Guicciardi, S.; Vitale, S.; Scannella, D.; Carbonara, P.; Dragičević, B.; et al. Mapping small-scale fisheries through a coordinated participatory strategy. Fish Fish. 2022, 23, 773–785. [Google Scholar] [CrossRef]
- Matić-Skoko, S.; Ikica, Z.; Vrdoljak, D.; Peharda, M.; Tutman, P.; Dragićević, B.; Joksimović, A.; Dulčić, J.; Durović, M.; Mandić, M.; et al. A comparative approach to the Croatian and Montenegrin small-scale fisheries (SSF) in the coastal eastern Adriatic Sea: Fishing gears and target species. Acta Adriat. 2017, 58, 459–480. [Google Scholar] [CrossRef] [Green Version]
- Pranovi, F.; Caccin, A.; Franzoi, P.; Malavasi, S.; Zucchetta, M.; Torricelli, P. Vulnerability of artisanal fisheries to climate change in the Venice lagoon. J. Fish Biol. 2013, 83, 847–863. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Stevens, P.W.; Lamont, M.M.; Brusca, R.C.; Hart, K.M.; Waddle, J.H.; Langtimm, C.A.; Williams, C.M.; Keim, B.D.; Terando, A.J.; et al. Tropicalization of temperate ecosystems in North America: The northward range expansion of tropical organisms in response to warming winter temperatures. Glob. Chang. Biol. 2021, 27, 3009–3034. [Google Scholar] [CrossRef]
- Goren, M.; Aronov, A. First record of the Indo-Pacific parrot fish Scarus ghobban in the Eastern Mediterranean. Cybium 2002, 26, 239–240. [Google Scholar]
- Lipej, L.; Dulčić, J. The current status of Adriatic fish biodiversity. In Balkan Biodiversity; Springer: Dordrecht, The Netherlands, 2004; pp. 291–306. [Google Scholar]
- Parenti, P.; Bressi, N. First record of the orange-spotted grouper, Epinephelus coioides (Perciformes: Serranidae) in the Northern Adriatic Sea. Cybium 2001, 25, 281–284. [Google Scholar]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modelling. In Proceedings of the Twenty-First International Conference on Machine Learning, Alberta, Canada, 4–8 July 2004; Maximum entropy modeling of species geographic distributions; Phillips, S.J., Anderson, R.P., Schapire, R.E., Eds.; Scientific Research Publishing: Wuhan, China, 2004; Volume 190, pp. 231–259. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. MaxEnt modelling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with MaxEnt: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; Carvalheiro, L.G.; Polce, C.; van Loon, E.E.; Raes, N.; Reemer, M.; Biesmeijer, J.C. Fit-for-purpose: Species distribution model performance depends on evaluation criteria–Dutch hoverflies as a case study. PLoS ONE 2013, 8, e63708. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- van Proosdij, A.S.; Sosef, M.S.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2016, 39, 542–552. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Bradie, J.; Leung, B. A quantitative synthesis of the importance of variables used in MaxEnt species distribution models. J. Biogeogr. 2017, 44, 1344–1361. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Animal Physiology: Adaptation and Environment; Cambridge University Press: New York, NY, USA, 1997. [Google Scholar]
- Tyberghein, L.; Verbruggen, H.; Pauly, K.; Troupin, C.; Mineur, F.; De Clerck, O. Bio-ORACLE: A global environmental dataset for marine species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 272–281. [Google Scholar] [CrossRef]
- Gogina, M.; Zettler, M.L. Diversity and distribution of benthic macrofauna in the Baltic Sea. Data inventory and its use for species distribution modelling and prediction. J. Sea Res. 2010, 64, 313–321. [Google Scholar] [CrossRef]
- Olivar, M.P.; Emelianov, M.; Villate, F.; Uriarte, I.; Maynou, F.; Alvarez, I.; Morote, E. The role of oceanographic conditions and plankton availability in larval fish assemblages off the Catalan coast (NW Mediterranean). Fish. Oceanogr. 2010, 19, 209–229. [Google Scholar] [CrossRef]
- Dulčić, J.; Kraljević, M.; Grbec, B.; Pallaoro, A. Composition and temporal fluctuations of inshore juvenile fish populations in the Kornati Archipelago, eastern middle Adriatic. Mar. Biol. 1997, 129, 267–277. [Google Scholar] [CrossRef]
- Assis, J.; Tyberghein, L.; Bosch, S.; Verbruggen, H.; Serrão, E.A.; De Clerck, O. Bio-ORACLE v2. 0: Extending marine data layers for bioclimatic modelling. Glob. Ecol. Biogeogr. 2018, 27, 277–284. [Google Scholar] [CrossRef]
- Rilov, G.; Galil, B. Marine bioinvasions in the Mediterranean Sea–history, distribution and ecology. In Biological Invasions in Marine Ecosystems; Springer: Berlin/Heidelberg, Germany, 2009; pp. 549–575. [Google Scholar]
- Cheung, W.W.; Lam, V.W.; Sarmiento, J.L.; Kearney, K.; Watson, R.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Mannino, A.M.; Balistreri, P.; Deidun, A. The marine biodiversity of the Mediterranean Sea in a changing climate: The impact of biological invasions. In Mediterranean Identities-Environment, Society, Culture; INTECH: London, UK, 2017; pp. 101–127. [Google Scholar] [CrossRef] [Green Version]
- Occhipinti-Ambrogi, A. Global change and marine communities: Alien species and climate change. Mar. Poll. Bull. 2007, 55, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Astraldi, M.; Bianchi, C.N.; Gasparini, G.P.; Morri, C. Climatic fluctuations, current variability and marine species distribution: A case study in the Ligurian Sea (north-west Mediterranean). Oceanol. Acta 1995, 18, 139–149. [Google Scholar]
- Bruno, J.F.; Bates, A.E.; Cacciapaglia, C.; Pike, E.P.; Amstrup, S.C.; van Hooidonk, R.; Henson, S.A.; Aronson, R.B. Climate change threatens the world’s marine protected areas. Nat. Clim. Chang. 2018, 8, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Hiddink, J.G.; Ben Rais Lasram, F.; Cantrill, J.; Davies, A.J. Keeping pace with climate change: What can we learn from the spread of Lessepsian migrants? Glob. Chang. Biol. 2012, 18, 161–2171. [Google Scholar] [CrossRef]
- Pimental, D. Biological Invasions: Economic and Environmental Costs of Alien Plant, Animal and Microbe Species; CRC: New York, NY, USA, 2002; 369p. [Google Scholar]
- Bettoso, N.; Dulčić, J. First record of the oilfish Ruvettus pretiosus (Pisces: Gempylidae) in Northern Adriatic Sea. J. Mar. Biol. Assoc. U. K. 1999, 79, 1145–1146. [Google Scholar] [CrossRef]
- Dulčić, J.; Grbec, B.; Lipej, L.; Beg Paklar, G.; Supić, N.; Smirčić, A. The effect of the hemispheric climatic oscillations on the Adriatic ichthyofauna. Fresenius Environ. Bull. 2004, 13, 293–298. [Google Scholar]
- Dulčić, J.; Grbec, B.; Lipej, L. Information on the Adriatic ichthyofauna. Effect of water warming? Acta Adriat. 1999, 40, 33–43. [Google Scholar]
- Dulčić, J.; Grbec, B. Climate change and Adriatic ichthyofauna. Fish. Oceanogr. 2000, 9, 187–191. [Google Scholar] [CrossRef]
- Iveša, N.; Piria, M.; Gelli, M.; Trnski, T.; Špelić, I.; Radočaj, T.; Kljak, K.; Jug-Dujaković, J.; Gavrilović, A. Feeding habits of predatory thermophilic fish species and species with subtropical affinity from recently extended distributional range in northeast Adriatic Sea, Croatia. Diversity 2021, 13, 357. [Google Scholar] [CrossRef]
- Psomadakis, P.N.; Ceddia, P.; Vacchi, M. Additional record of Sphoeroides pachygaster (Pisces: Tetraodontidae) in the Tyrrhenian Sea and notes on the distribution of the species in the Mediterranean. JMBA2 Biodivers. Rec. 2006, 1, e18. [Google Scholar] [CrossRef]
- Sinovčić, G.; Franičević, M.; Keč, V. Unusual occurrence and some aspects of biology of juvenile gilt sardine (Sardinella aurita Valenciennes, 1847) in the Zrmanja River estuary (eastern Adriatic). J. Appl. Ichthyol. 2004, 20, 53–57. [Google Scholar] [CrossRef]
- CIESM. Climate warming and related changes in Mediterranean marine biota. In N° 35 in CIESM Workshop Monographs; Briand, F., Ed.; CIESM: Monte Carlo, Monaco, 2018; 152p. [Google Scholar]
- Hill, J.; DLFowler Van De Avyle, M.J. Species Profiles: Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates (Mid-Atlantic) Blue Crab; U.S. Army Corps of Engineers: Washington, DC, USA, 1989; 18p.
- Giordani Soika, A. Il Neptunus pelagicus (L.) nell’alto Adriatico. Natura 1951, 42, 18–20. [Google Scholar]
- Streftaris, N.; Zenetos, A. Alien Marine Species in the Mediterranean—the 100 ‘Worst Invasives’ and their Impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef] [Green Version]
- Tiralongo, F.; Villani, G.; Arciprete, R.; Mancini, E. Filling the gap on Italian records of an invasive species: First records of the Blue Crab, Callinectes sapidus Rathbun, 1896 (Decapoda: Brachyura: Portunidae), in Latium and Campania (Tyrrhenian Sea). Acta Adriat. 2012, 61, 99–104. [Google Scholar] [CrossRef]
- Gienapp, P.; Teplitsky, C.; Alho, J.; Mills, J.; Merila, J. Climate change and evolution: Disentangling environmental and genetic responses. Mol. Ecol. 2008, 17, 167–178. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- D’Amen, M.; Azzurro, E. Lessepsian fish invasion in Mediterranean marine protected areas: A risk assessment under climate change scenarios. ICES J. Mar. Sci. 2020, 77, 388–397. [Google Scholar] [CrossRef]
- Russo, A.; Artegiani, A. Adriatic Sea hydrography. Sci. Mar. 1996, 60, 33–43. [Google Scholar]
- Barausse, A.; Duci, A.; Mazzoldi, C.; Artioli, Y.; Palmeri, L. Trophic network model of the Northern Adriatic Sea: Analysis of an exploited and eutrophic ecosystem. Estuar. Coast Shelf Sci. 2009, 83, 77–590. [Google Scholar] [CrossRef]
- Fortibuoni, T.; Giovanardi, O.; Pranovi, F.; Raicevich, S.; Solidoro, C.; Libralato, S. Analysis of long-term changes in a Mediterranean marine ecosystem based on fishery landings. Front. Mar. Sci. 2017, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Pranovi, F.; Anelli Monti, M.; Caccin, A.; Brigolin, D.; Zucchetta, M. Permanent trawl fishery closures in the Mediterranean Sea: An effective management strategy? Mar. Pol. 2015, 60, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.; Monti, M.A.; Mangano, M.C.; Raffaetà, A.; Sarà, G.; Silvestri, C.; Pranovi, F. Temporal and spatial patterns of trawl fishing activities in the Adriatic Sea (Central Mediterranean Sea, GSA17). Ocean Coast. Manag. 2020, 192, 105231. [Google Scholar] [CrossRef]
- Russo, E.; Anelli Monti, M.; Toninato, G.; Silvestri, C.; Raffaetà, A.; Pranovi, F. Lockdown: How the COVID-19 Pandemic Affected the Fishing Activities in the Adriatic Sea (Central Mediterranean Sea). Front. Mar. Sci. 2021, 8, 685808. [Google Scholar] [CrossRef]
- Jacquemont, J.; Blasiak, R.; Le Cam, C.; Le Gouellec, M.; Claudet, J. Ocean conservation boosts climate change mitigation and adaptation. One Earth 2002, 5, 1126–1138. [Google Scholar] [CrossRef]
- Busch, D.S.; Griffis, R.; Link, J.; Abrams, K.; Baker, J.; Brainard, R.E.; Ford, M.; Hare, J.A.; Himes-Cornell, A.; Hollowed, A.; et al. Climate science strategy of the US National Marine Fisheries Service. Mar. Policy 2016, 74, 58–67. [Google Scholar] [CrossRef]
- Link, J.S.; Griffis, R.; Busch, S. NOAA Fisheries Climate Science Strategy; NOAA Technical Memorandum NMFS-F/SPO-155; U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service: Silver Spring, MD, USA, 2015; p. 70.
- Pinsky, M.L.; Mantua, N. Emerging adaptation approaches for climate-ready fisheries management. Oceanography 2014, 27, 146–159. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Acronym | Climatic Affinity |
|---|---|---|---|
| Callinectes sapidus | Blue crab | CSA | Warm |
| Chelidonichthys lucerna | Tub gurnard | CLU | Ubiquitous |
| Chelon ramada | Thinlip grey mullet | CRA | Cold/Temperate |
| Coryphaena hippurus | Common dolphinfish | CHI | Warm |
| Dicentrarchus labrax | European seabass | DLA | Cold |
| Lichia amia | Leerfish | LAM | Warm |
| Merlangius merlangus | Whiting | MMG | Cold |
| Merluccius merluccius | European hake | MMC | Cold |
| Mullus surmuletus | Surmullet | MSU | Cold/Temperate |
| Mustelus mustelus | Smooth-hound | MMU | Temperate |
| Pomatomus saltatrix | Bluefish | PSA | Temperate/Warm |
| Scorpaena scrofa | Red scorpionfish | SSC | Temperate |
| Sepia officinalis | Common cuttlefish | SOF | Cold |
| Seriola dumerili | Greater amberjack | SDU | Warm |
| Solea solea | Common sole | SSO | Cold |
| Sparus aurata | Gilthead seabream | SAU | Temperate |
| Squilla mantis | Mantis shrimp | SMA | Temperate |
| Species | AUC | 2050 | 2100 | ||||
|---|---|---|---|---|---|---|---|
| Latin Name | Common Name | Acronym | RCP4.5 | RCP8.5 | RCP4.5 | RCP8.5 | |
| Callinectes sapidus | Blue crab | CSA | 0.958 | 0.710 | 0.708 | 0.737 | 0.710 |
| Chelidonichthys lucerna | Tub gurnard | CLU | 0.963 | 0.445 | 0.453 | 0.415 | 0.061 |
| Chelon ramada | Thinlip grey mullet | CRA | 0.982 | 0.364 | 0.374 | 0.347 | 0.142 |
| Coryphaena hippurus | Common dolphinfish | CHI | 0.669 | 0.500 | 0.500 | 0.476 | 0.476 |
| Dicentrarchus labrax | European seabass | DLA | 0.977 | 0.204 | 0.216 | 0.189 | 0.025 |
| Lichia amia | Leerfish | LAM | 0.973 | 0.514 | 0.513 | 0.501 | 0.461 |
| Merlangius merlangus | Whiting | MMG | 0.973 | 0.395 | 0.420 | 0.393 | 0.159 |
| Merluccius merluccius | European hake | MMC | 0.979 | 0.494 | 0.496 | 0.463 | 0.208 |
| Mullus surmuletus | Surmullet | MSU | 0.980 | 0.312 | 0.305 | 0.344 | 0.046 |
| Mustelus mustelus | Smooth-hound | MMU | 0.931 | 0.400 | 0.401 | 0.377 | 0.305 |
| Pomatomus saltatrix | Bluefish | PSA | 0.908 | 0.482 | 0.490 | 0.467 | 0.431 |
| Scorpaena scrofa | Red scorpionfish | SSC | 0.928 | 0.500 | 0.500 | 0.476 | 0.350 |
| Sepia officinalis | Common cuttlefish | SOF | 0.973 | 0.419 | 0.428 | 0.392 | 0.170 |
| Seriola dumerili | Greater amberjack | SDU | 0.938 | 0.500 | 0.497 | 0.454 | 0.324 |
| Solea solea | Common sole | SSO | 0.977 | 0.452 | 0.461 | 0.427 | 0.301 |
| Sparus aurata | Gilthead seabream | SAU | 0.989 | 0.433 | 0.430 | 0.408 | 0.292 |
| Squilla mantis | Mantis shrimp | SMA | 0.980 | 0.508 | 0.508 | 0.490 | 0.141 |
| Thermal Affinity | Species | Percent Contribution | |||||
|---|---|---|---|---|---|---|---|
| ubiquitous | Chelidonichthys lucerna | Tmean | Tmin | Smax | Smin | Smean | Tmax |
| 39.7 | 27 | 14.8 | 13.9 | 3 | 1.6 | ||
| cold | Dicentrarchus labrax | Tmin | Tmean | Tmax | Smax | Smin | Smean |
| 48.7 | 28.1 | 12.9 | 9.9 | 0.3 | 0.1 | ||
| Merlangius merlangus | Tmin | Tmean | Tmax | Smax | Smin | Smean | |
| 50.7 | 41.5 | 4.2 | 3.1 | 0.5 | 0 | ||
| Merluccius merluccius | Smax | Tmean | Tmin | Smean | Smin | Tmax | |
| 52.7 | 34.7 | 9.8 | 1.9 | 0.4 | 0.4 | ||
| Sepia officinalis | Tmin | Tmean | Smax | Tmax | Smean | Smin | |
| 49 | 39.4 | 7.1 | 2.6 | 1.1 | 0.8 | ||
| Solea solea | Tmin | Tmean | Tmax | Smax | Smin | Smean | |
| 51.6 | 35.5 | 6.2 | 5.6 | 1.2 | 0.1 | ||
| temperate/cold | Chelon ramada | Tmin | Tmean | Smax | Smin | Tmax | Smean |
| 38.9 | 26.4 | 25.5 | 5.4 | 2.2 | 1.6 | ||
| Mullus surmuletus | Tmean | Smax | Tmin | Tmax | Smean | Smin | |
| 41.5 | 31.3 | 23.1 | 3.1 | 0.8 | 0.2 | ||
| temperate | Mustelus mustelus | Tmean | Tmin | Smax | Smin | Tmax | Smean |
| 37.2 | 30.1 | 21.5 | 10.5 | 0.5 | 0.2 | ||
| Scorpaena scrofa | Smin | Tmean | Tmin | Smax | Smean | Tmax | |
| 27.1 | 26.4 | 17.7 | 16.2 | 10.7 | 1.9 | ||
| Sparus aurata | Smax | Tmin | Tmean | Tmax | Smin | Smean | |
| 54.2 | 27.9 | 17.6 | 0.3 | 0.1 | 0 | ||
| Squilla mantis | Smax | Tmin | Tmax | Tmean | Smin | Smean | |
| 28.8 | 19.8 | 18.3 | 17.8 | 13.7 | 1.6 | ||
| warm/temperate | Pomatomus saltatrix | Tmin | Tmax | Tmean | Smin | Smax | Smean |
| 35.9 | 30.8 | 17.7 | 10.4 | 2.7 | 2.5 | ||
| warm | Callinectes sapidus | Tmax | Tmin | Smin | Smax | Tmean | Smean |
| 45.6 | 25.4 | 19.8 | 7.5 | 1.3 | 0.3 | ||
| Coryphaena hippurus | Tmax | Tmean | Smax | Tmin | Smean | Smin | |
| 48 | 37.7 | 9 | 3.6 | 1.7 | 0 | ||
| Lichia amia | Tmin | Smin | Smax | Tmean | Smean | Tmax | |
| 66.6 | 13.9 | 12.3 | 6 | 1.2 | 0 | ||
| Seriola dumerili | Tmax | Tmin | Tmean | Smax | Smin | Smean | |
| 43.4 | 28.7 | 19 | 6.3 | 2.5 | 0.1 | ||
| tropical | Epinephelus coioides | Tmin | Tmax | Smin | Tmean | Smax | Smean |
| 42.1 | 34.7 | 11.4 | 6.5 | 5.1 | 0.1 | ||
| Scarus ghobban | Tmin | Tmean | Tmax | Smax | Smin | Smean | |
| 58.6 | 14.9 | 13.6 | 12.6 | 0.2 | 0.1 | ||
| Terapon theraps | Tmin | Tmean | Smin | Tmax | Smax | Smean | |
| 38 | 21.9 | 20.9 | 16 | 2.6 | 0.5 | ||
| Tropical Species | Common Name | AUC | 2050 | 2100 | ||
|---|---|---|---|---|---|---|
| RCP4.5 | RCP8.5 | RCP4.5 | RCP8.5 | |||
| Epinephelus coioides | Orange-spotted grouper | 0.951 | 0.748 | 0.743 | 0.779 | 0.980 |
| Scarus ghobban | Blue-barred parrotfish | 0.900 | 0.975 | 0.975 | 0.978 | 0.992 |
| Terapon theraps | Large-scaled terapon | 0.936 | 0.881 | 0.875 | 0.898 | 0.984 |
| Time Frame | RCP Scenario | Coastal Waters | Transitional Waters | ||
|---|---|---|---|---|---|
| Landing Loss (%) | Economic Loss (EUR y−1) | Landing Loss (%) | Economic Loss (EUR y−1) | ||
| Short term (2040–2050) | 4.5 | −13.5 (1.4) | 65,869 (14,347) | −23.2 (2.2) | 11,824 (2750) |
| 8.5 | −15.4 (1.7) | 76,050 (16,797) | −26.8 (2.4) | 13,756 (3114) | |
| Medium term (2090–2100) | 4.5 | −23.1 (1.7) | 112,264 (18,313) | −35.8 (2.5) | 18,567 (3904) |
| 8.5 | −58.4 (2.9) | 284,607 (39,329) | −86.9 (1.5) | 46,044 (10,086) | |
| Scenarios | p adj Coastal Waters | p adj Transitional Waters |
|---|---|---|
| RCP4.5_2050–RCP8.5_2050 | n.s. | n.s. |
| RCP4.5_2100–RCP8.5_2100 | <0.001 | <0.001 |
| RCP4.5_2050–RCP4.5_2100 | <0.05 | <0.05 |
| RCP8.5_2050–RCP8.5_2100 | <0.001 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavraro, F.; Anelli Monti, M.; Matić-Skoko, S.; Caccin, A.; Pranovi, F. Vulnerability of the Small-Scale Fishery to Climate Changes in the Northern-Central Adriatic Sea (Mediterranean Sea). Fishes 2023, 8, 9. https://doi.org/10.3390/fishes8010009
Cavraro F, Anelli Monti M, Matić-Skoko S, Caccin A, Pranovi F. Vulnerability of the Small-Scale Fishery to Climate Changes in the Northern-Central Adriatic Sea (Mediterranean Sea). Fishes. 2023; 8(1):9. https://doi.org/10.3390/fishes8010009
Chicago/Turabian StyleCavraro, Francesco, Marco Anelli Monti, Sanja Matić-Skoko, Alberto Caccin, and Fabio Pranovi. 2023. "Vulnerability of the Small-Scale Fishery to Climate Changes in the Northern-Central Adriatic Sea (Mediterranean Sea)" Fishes 8, no. 1: 9. https://doi.org/10.3390/fishes8010009