
Improvements in Human Keratinocytes and Antimicrobial Effect Mediated by Cell-Free Supernatants Derived from Probiotics

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of CFS
2.2. Cell Culture and Viability
2.3. Quantification of HA Production
2.4. Quantitative Real Time-Polymerase Chain Reaction (qRT-PCR)
2.5. Antimicrobial Effect against Escherichia coli and Staphylococcus aureus
2.6. Adherence to HT-29 Colorectal Cells
2.7. Statistical Analysis
3. Results and Discussion
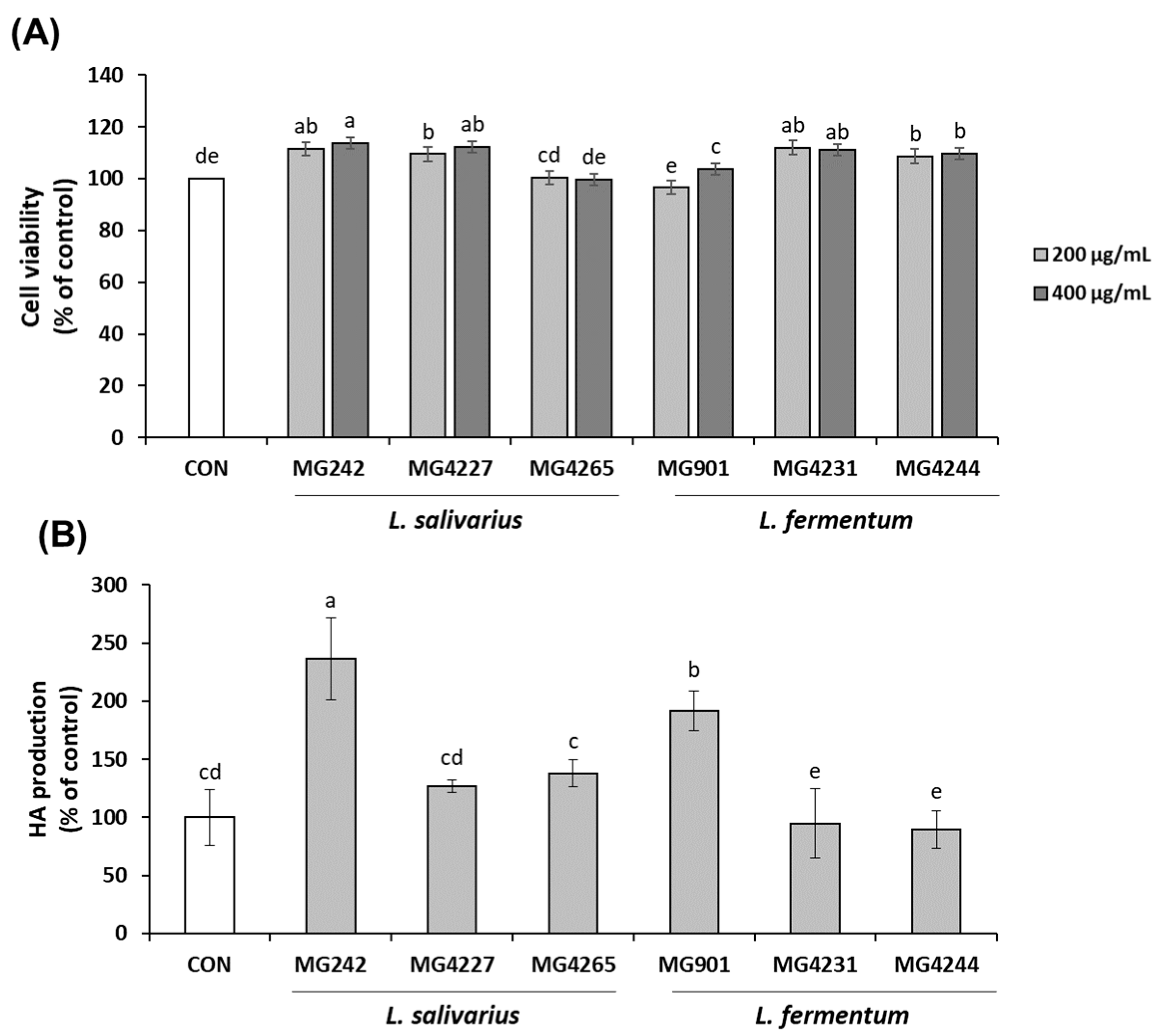
3.1. Effect of CFS Derived from L. salivarius and L. fermentum on Cell Viability and Hyaluronic Acid Production in HaCaT Keratinocytes
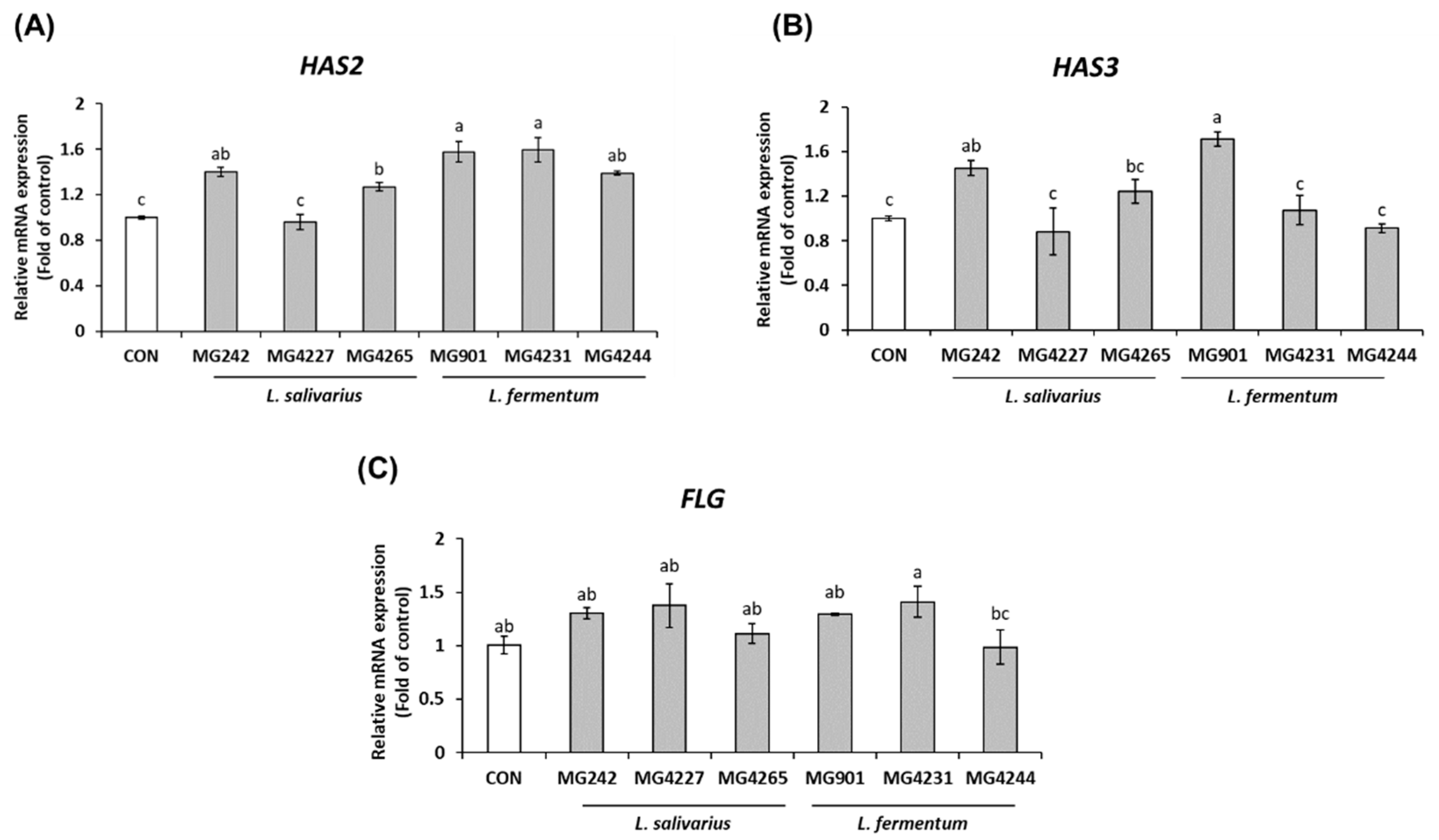
3.2. CFS Derived from L. salivarius and L. fermentum Affects mRNA Expression Associated with Skin Moisturization and Barrier Maintenance in HaCaT Keratinocytes
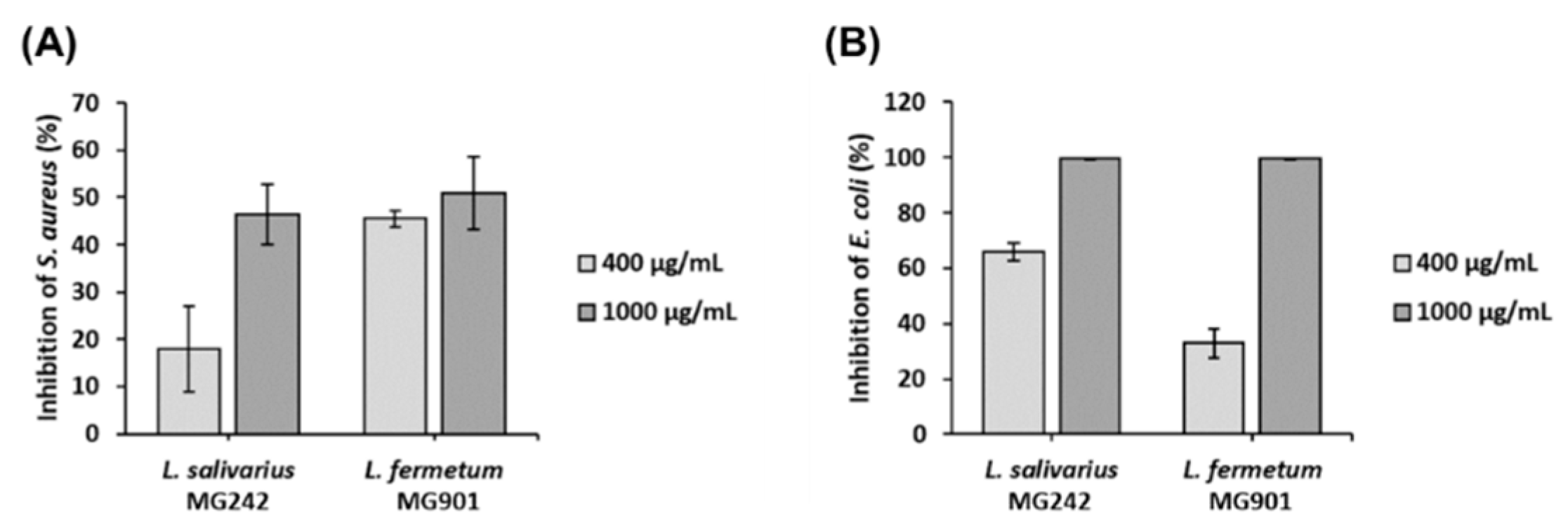
3.3. Antibacterial Effect of CFS Derived from L. salivarius MG242 and L. fermentum MG901
3.4. Adherence of Probiotics to HT-29 Colorectal Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kanitakis, J. Anatomy, histology and immunohistochemistry of normal human skin. Eur. J. Dermatol. 2002, 12, 390–399. [Google Scholar] [PubMed]
- Coates, M.; Blanchard, S.; MacLeod, A.S. Innate antimicrobial immunity in the skin: A protective barrier against bacteria, viruses, and fungi. PLoS Pathog. 2018, 14, e1007353. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Tsoi, L.C.; Billi, A.C.; Ward, N.L.; Harms, P.W.; Zeng, C.; Maverakis, E.; Kahlenberg, J.M.; Gudjonsson, J.E. Cytokinocytes: The diverse contribution of keratinocytes to immune responses in skin. JCI Insight 2020, 5, e142067. [Google Scholar] [CrossRef] [PubMed]
- Bitschar, K.; Wolz, C.; Krismer, B.; Peschel, A.; Schittek, B. Keratinocytes as sensors and central players in the immune defense against Staphylococcus aureus in the skin. J. Dermatol. Sci. 2017, 87, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Kakehi, K.; Kinoshita, M.; Yasueda, S.-i. Hyaluronic acid: Separation and biological implications. J. Chromatogr. B 2003, 797, 347–355. [Google Scholar] [CrossRef]
- Sanders, M.E. Probiotics: Definition, sources, selection, and uses. Clin. Infect. Dis. 2008, 46, S58–S61. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-S.; Lim, H.-S.; Oh, J.-S.; Lim, Y.-j.; Wuertz-Kozak, K.; Harro, J.M.; Shirtliff, M.E.; Achermann, Y. Antimicrobial activity of Lactobacillus salivarius and Lactobacillus fermentum against Staphylococcus aureus. Pathog. Dis. 2017, 75, ftx009. [Google Scholar] [CrossRef]
- Higuchi, T.; Suzuki, N.; Nakaya, S.; Omagari, S.; Yoneda, M.; Hanioka, T.; Hirofuji, T. Effects of Lactobacillus salivarius WB21 combined with green tea catechins on dental caries, periodontitis, and oral malodor. Arch. Oral Biol. 2019, 98, 243–247. [Google Scholar] [CrossRef]
- Niccoli, A.A.; Artesi, A.L.; Candio, F.; Ceccarelli, S.; Cozzali, R.; Ferraro, L.; Fiumana, D.; Mencacci, M.; Morlupo, M.; Pazzelli, P. Preliminary results on clinical effects of probiotic Lactobacillus salivarius LS01 in children affected by atopic dermatitis. J. Clin. Gastroenterol. 2014, 48, S34–S36. [Google Scholar] [CrossRef]
- Rodríguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Utrilla, M.P.; Chueca, N.; Garcia, F.; Olivares, M.; Rodríguez-Cabezas, M.E.; Gálvez, J. Differential intestinal anti-inflammatory effects of Lactobacillus fermentum and Lactobacillus salivarius in DSS mouse colitis: Impact on microRNAs expression and microbiota composition. Mol. Nutr. Food Res. 2017, 61, 1700144. [Google Scholar] [CrossRef]
- Kim, S.; Choi, S.-I.; Jang, M.; Jeong, Y.; Kang, C.-H.; Kim, G.-H. Anti-adipogenic effect of Lactobacillus fermentum MG4231 and MG4244 through AMPK pathway in 3T3-L1 preadipocytes. Food Sci. Biotechnol. 2020, 29, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3390–3415. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, Y.; Kang, C.-H. In vivo confirmation of the antimicrobial effect of probiotic candidates against Gardnerella vaginalis. Microorganisms 2021, 9, 1690. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-H.; Kim, J.-S.; Park, H.M.; Kim, S.; Paek, N.-S. Antioxidant activity and short-chain fatty acid production of lactic acid bacteria isolated from Korean individuals and fermented foods. 3 Biotech 2021, 11, 217. [Google Scholar] [CrossRef]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—a step beyond pre-and probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Kim, D.-W.; Chae, S.-J.; Kim, Y.-H.; Jung, H.-J.; Lee, S.-D.; Park, J.-C.; Cho, K.-H.; Sa, S.-J.; Kim, I.-C.; Kim, I.-H. Effects of prebiotics and probiotics on swine intestinal microflora and fermentation products in vitro fermentation. Korean J. Microbiol. 2013, 49, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.M.; Yosipovitch, G. Skin pH: From basic science to basic skin care. Acta Derm. Vener. 2013, 93, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Kang, C.-H. Probiotics alleviate oxidative stress in H2O2-exposed hepatocytes and t-BHP-induced C57BL/6 mice. Microorganisms 2022, 10, 234. [Google Scholar] [CrossRef]
- Kim, S.-G.; Lee, Y.-D.; Park, J.-H.; Moon, G.-S. Synergistic inhibition by bacteriocin and bacteriophage against Staphylococcus aureus. Food Sci. Anim. Resour. 2019, 39, 1015. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, J.Y.; Jeong, Y.; Kang, C.-H. Antioxidant activity and probiotic properties of lactic acid bacteria. Fermentation 2022, 8, 29. [Google Scholar] [CrossRef]
- Parolin, C.; Abruzzo, A.; Giordani, B.; Oliver, J.C.; Marangoni, A.; Luppi, B.; Vitali, B. Anti-candida activity of hyaluronic acid combined with Lactobacillus crispatus lyophilised supernatant: A new antifungal strategy. Antibiotics 2021, 10, 628. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Derm.-Endocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, G.; KV, K.; Prasad, S.B.; Bhaduri, A.; Jayaraman, G. Biosynthesis of hyaluronic acid polymer: Dissecting the role of sub structural elements of hyaluronan synthase. Sci. Rep. 2019, 9, 12510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spicer, A.P.; McDonald, J. Eukaryotic Hyaluronan Synthases. Available online: http://www.glycoforum.gr.jp/science/hyaluronan/HA07/HA07E.html (accessed on 20 May 2022).
- Kim, Y.; Lim, K.-M. Skin barrier dysfunction and filaggrin. Arch. Pharm. Res. 2021, 44, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Maeda, K. New functions of low-molecular-weight hyaluronic acid on epidermis filaggrin production and degradation. Cosmetics 2021, 8, 118. [Google Scholar] [CrossRef]
- Ryu, S.; Song, P.I.; Seo, C.H.; Cheong, H.; Park, Y. Colonization and infection of the skin by S. aureus: Immune system evasion and the response to cationic antimicrobial peptides. Int. J. Mol. Sci. 2014, 15, 8753–8772. [Google Scholar] [CrossRef] [Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Pessemier, B.D.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut–skin axis: Current knowledge of the interrelationship between microbial dysbiosis and skin conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef]
- Sinha, S.; Lin, G.; Ferenczi, K. The skin microbiome and the gut-skin axis. Clin. Dermatol. 2021, 39, 829–839. [Google Scholar] [CrossRef]
- Singh, Y.; Ahmad, J.; Musarrat, J.; Ehtesham, N.Z.; Hasnain, S.E. Emerging importance of holobionts in evolution and in probiotics. Gut Pathog. 2013, 5, 12. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Primer | Sequence (5′→3′) |
|---|---|---|
| HAS2 | Forward | ATTACCCAGTCCTGGCTTCG |
| Reverse | CCTGTGGAAGACTCAGCAGAA | |
| HAS3 | Forward | TGTCCAGATCCTCAACAAGTACGA |
| Reverse | CTGGAGGAGGCTGTTGC | |
| FLG | Forward | GGCTAAGTGAAAGACTTGAAGAGA |
| Reverse | AATAGACTATCAGTGGTGTCATAGG | |
| GAPDH | Forward | GTCTTCACCACCATGGAGAA |
| Reverse | AGGAGGCATTGCTGATGAT |
| Strains | Initial Counts (CFU/mL) | Adhesion Counts (CFU/mL) | Adherence (%) |
|---|---|---|---|
| L. salivarius MG242 | 8.54 ± 0.05 | 7.41 ± 0.07 | 85.79 ± 1.55% |
| L. fermentum MG901 | 8.62 ± 0.01 | 7.15 ± 0.02 | 82.69 ± 0.41% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.Y.; Kim, Y.; Kim, J.-I.; Lee, H.-Y.; Moon, G.-S.; Kang, C.-H. Improvements in Human Keratinocytes and Antimicrobial Effect Mediated by Cell-Free Supernatants Derived from Probiotics. Fermentation 2022, 8, 332. https://doi.org/10.3390/fermentation8070332
Lee JY, Kim Y, Kim J-I, Lee H-Y, Moon G-S, Kang C-H. Improvements in Human Keratinocytes and Antimicrobial Effect Mediated by Cell-Free Supernatants Derived from Probiotics. Fermentation. 2022; 8(7):332. https://doi.org/10.3390/fermentation8070332
Chicago/Turabian StyleLee, Ji Yeon, YongGyeong Kim, Ja-I Kim, Hyang-Yeol Lee, Gi-Seong Moon, and Chang-Ho Kang. 2022. "Improvements in Human Keratinocytes and Antimicrobial Effect Mediated by Cell-Free Supernatants Derived from Probiotics" Fermentation 8, no. 7: 332. https://doi.org/10.3390/fermentation8070332