
Changes in Phenolic Profiles and Inhibition Potential of Macrophage Foam Cell Formation during Noni (Morinda citrifolia Linn.) Fruit Juice Fermentation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Sample Preparation
2.3. Extraction of Phenolic Compounds
2.4. Phenolic Composition Analysis by UPLC–Q-TOF–MS/MS
2.5. Quantitative Analysis by UPLC
2.6. Antioxidant Activity Assay
2.6.1. DPPH Radical Scavenging Activity Assay
2.6.2. ABTS+ Radical Scavenging Activity Assay
2.6.3. OH∙(Hydroxyl) Radical Scavenging Activity Assay
2.6.4. Ferric Reducing Antioxidant Power (FRAP) Assay
2.7. Cell Experiments
2.7.1. Cell Culture and Treatment
2.7.2. Oil Red O Staining and Determination of the Intracellular lipid
2.7.3. Determination of the Cholesterol Content
2.8. Statistical Analysis
3. Results
3.1. Identification of Phenolic Compositions
3.2. Quantification of Predominant Individual Phenolic Compounds
3.3. Changes in Antioxidant Activity during Fermentation
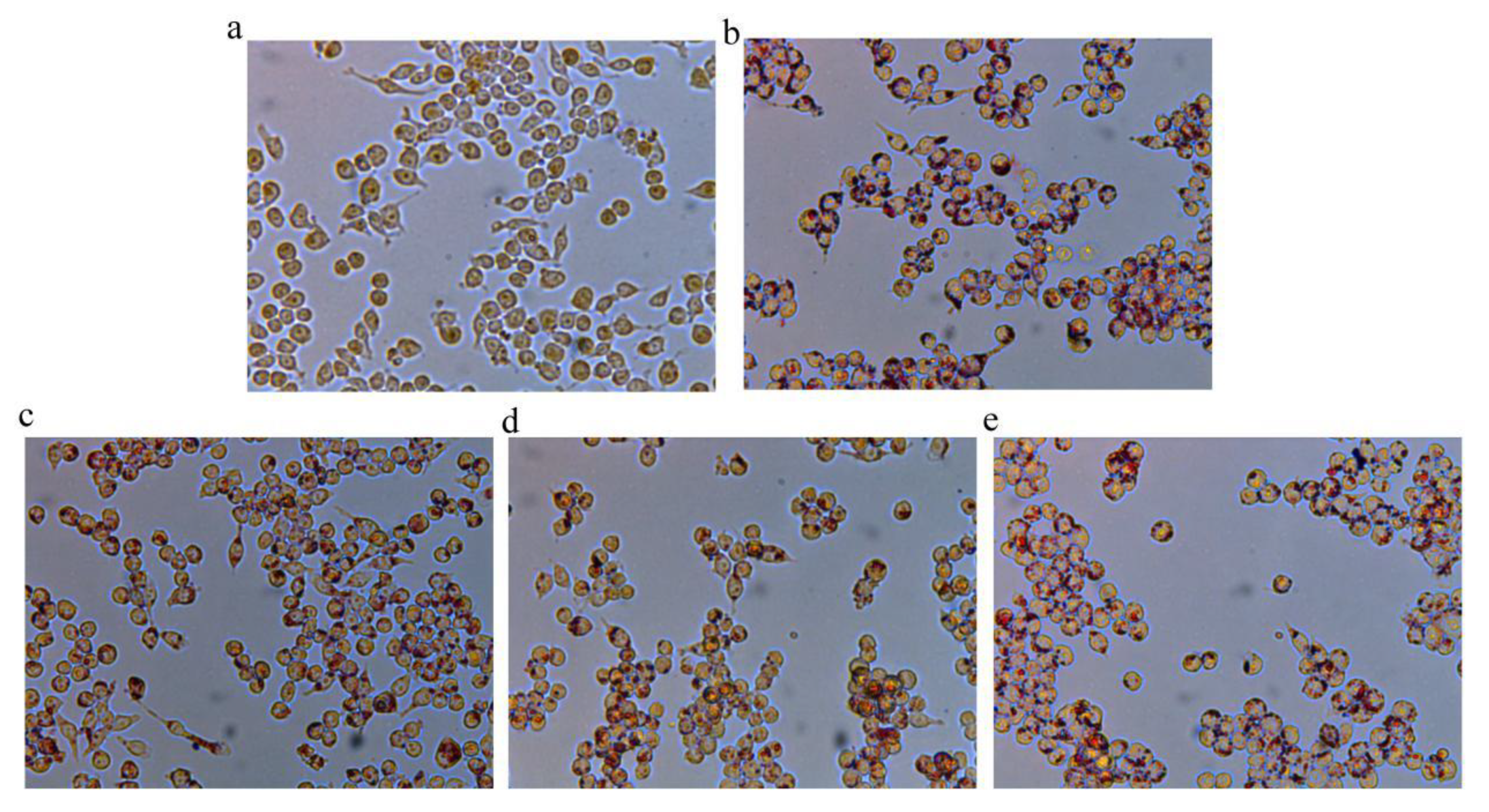
3.4. Oil Red O Staining and Analysis of Lipid Accumulation
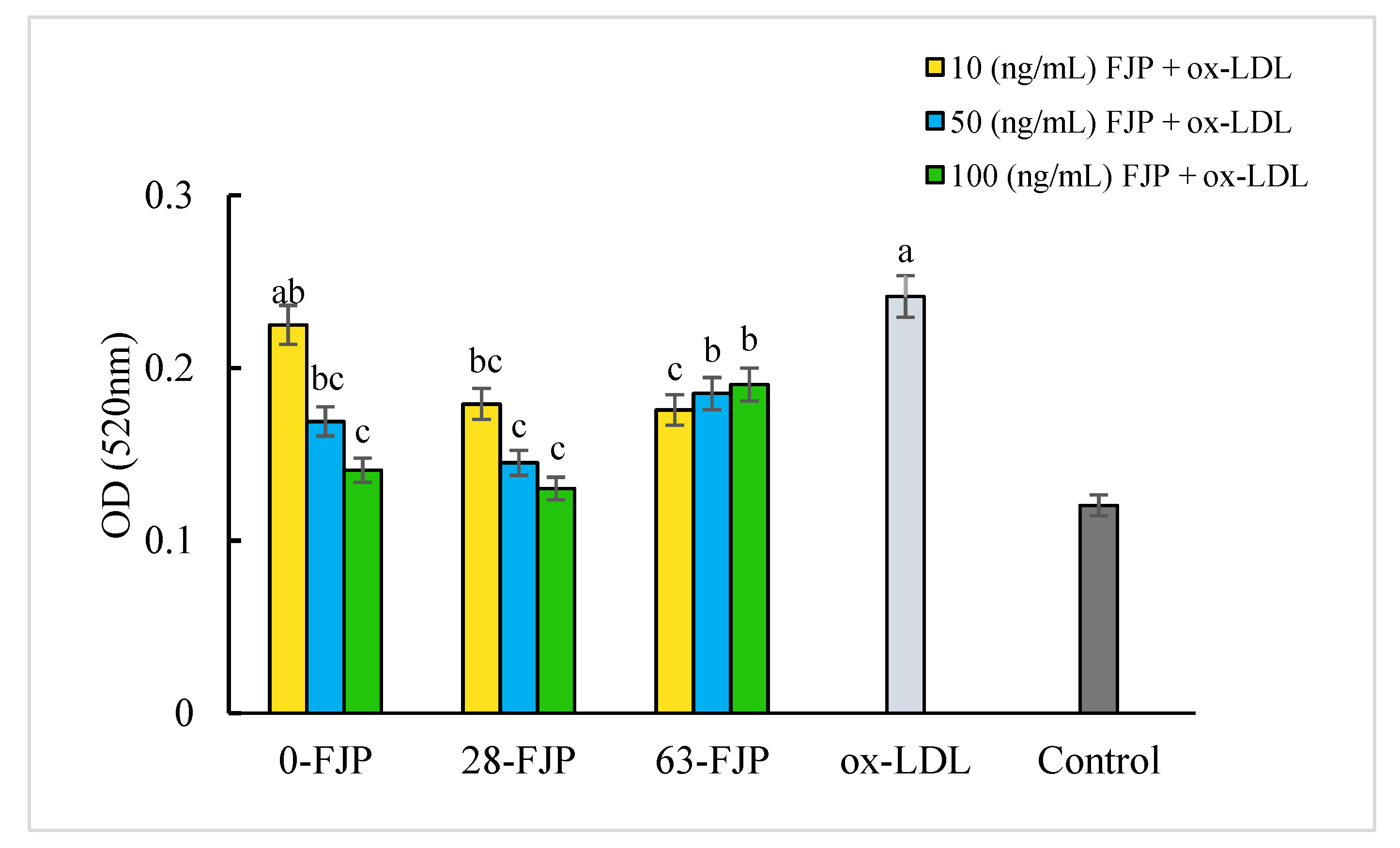
3.5. Effect of FJP on Intracellular Cholesterol Content in Foam Cells
3.6. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Abbasi-Parizad, P.; De Nisi, P.; Adani, F.; Pepe Sciarria, T.; Squillace, P.; Scarafoni, A.; Iametti, S.; Scaglia, B. Antioxidant and Anti-Inflammatory Activities of the Crude Extracts of Raw and Fermented Tomato Pomace and Their Correlations with Aglycate-Polyphenols. Antioxidants 2020, 9, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, M.; Arnaez, E.; Moreira, I.; Quesada, S.; Azofeifa, G.; Wilhelm, K.; Vargas, F.; Chen, P. Polyphenolic Characterization, Antioxidant, and Cytotoxic Activities of Mangifera indica Cultivars from Costa Rica. Foods 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Li, W.; You, B.; Tang, W.; Gan, T.; Feng, C.; Li, C.; Yang, R. Serum Metabonomic Study on the Antidepressant-like Effects of Ellagic Acid in a Chronic Unpredictable Mild Stress-Induced Mouse Model. J. Agric. Food Chem. 2020, 68, 9546–9556. [Google Scholar] [CrossRef]
- Lira, S.M.; Dionisio, A.P.; Holanda, M.O.; Marques, C.G.; Silva, G.S.D.; Correa, L.C.; Santos, G.B.M.; de Abreu, F.A.P.; Magalhaes, F.E.A.; Reboucas, E.L.; et al. Metabolic profile of pitaya (Hylocereus polyrhizus (F.A.C. Weber) Britton & Rose) by UPLC-QTOF-MS(E) and assessment of its toxicity and anxiolytic-like effect in adult zebrafish. Food Res. Int. 2020, 127, 108701. [Google Scholar] [CrossRef]
- Adriouch, S.; Lampure, A.; Nechba, A.; Baudry, J.; Assmann, K.; Kesse-Guyot, E.; Hercberg, S.; Scalbert, A.; Touvier, M.; Fezeu, L.K. Prospective Association between Total and Specific Dietary Polyphenol Intakes and Cardiovascular Disease Risk in the Nutrinet-Sante French Cohort. Nutrients 2018, 10, 1587. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Tresserra-Rimbau, A.; Estruch, R.; Martinez-Gonzalez, M.A.; Medina-Remon, A.; Fito, M.; Corella, D.; Salas-Salvado, J.; Portillo, M.P.; Moreno, J.J.; et al. Polyphenol Levels Are Inversely Correlated with Body Weight and Obesity in an Elderly Population after 5 Years of Follow Up (The Randomised PREDIMED Study). Nutrients 2017, 9, 452. [Google Scholar] [CrossRef] [Green Version]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Red Raspberries and Their Bioactive Polyphenols: Cardiometabolic and Neuronal Health Links. Adv. Nutr. 2016, 7, 44–65. [Google Scholar] [CrossRef] [Green Version]
- Mattera, R.; Benvenuto, M.; Giganti, M.G.; Tresoldi, I.; Pluchinotta, F.R.; Bergante, S.; Tettamanti, G.; Masuelli, L.; Manzari, V.; Modesti, A.; et al. Effects of Polyphenols on Oxidative Stress-Mediated Injury in Cardiomyocytes. Nutrients 2017, 9, 523. [Google Scholar] [CrossRef] [Green Version]
- Ravi, S.K.; Narasingappa, R.B.; Vincent, B. Neuro-nutrients as anti-alzheimer’s disease agents: A critical review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2999–3018. [Google Scholar] [CrossRef]
- Nowak, D.; Goslinski, M.; Przygonski, K.; Wojtowicz, E. The antioxidant properties of exotic fruit juices from acai, maqui berry and noni berries. Eur. Food Res. Technol. 2018, 244, 1897–1905. [Google Scholar] [CrossRef]
- Tresserra-Rimbau, A.; Lamuela-Raventos, R.M.; Moreno, J.J. Polyphenols, food and pharma. Current knowledge and directions for future research. Biochem. Pharmacol. 2018, 156, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Szwajgier, D.; Paduch, R.; Kukula-Koch, W.; Wasko, A.; Polak-Berecka, M. Fermented curly kale as a new source of gentisic and salicylic acids with antitumor potential. J. Funct. Foods 2020, 67, 103866. [Google Scholar] [CrossRef]
- Gong, X.; Jiang, S.; Tian, H.; Xiang, D.; Zhang, J. Polyphenols in the Fermentation Liquid of Dendrobium candidum Relieve Intestinal Inflammation in Zebrafish Through the Intestinal Microbiome-Mediated Immune Response. Front. Immunol. 2020, 11, 1542. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.R.; McMillen, H.; Etkin, N.L. Ferment this: The transformation of Noni, a traditional Polynesian medicine (Morinda citrifolia, Rubiaceae). Econ. Bot. 1999, 53, 51–68. [Google Scholar] [CrossRef]
- Wall, M.M.; Miller, S.; Siderhurst, M.S. Volatile changes in Hawaiian noni fruit, Morinda citrifolia L., during ripening and fermentation. J. Sci. Food Agric. 2018, 98, 3391–3399. [Google Scholar] [CrossRef] [PubMed]
- Pawlus, A.D.; Kinghorn, D.A. Review of the ethnobotany, chemistry, biological activity and safety of the botanical dietary supplement Morinda citrifolia (noni). J. Pharm. Pharmacol. 2007, 59, 1587–1609. [Google Scholar] [CrossRef] [PubMed]
- Mohd Zin, Z.; Abdul Hamid, A.; Osman, A.; Saari, N.; Misran, A. Isolation and Identification of Antioxidative Compound from Fruit of Mengkudu (Morinda citrifolia L.). Int. J. Food Prop. 2007, 10, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Chan-Blanco, Y.; Vaillant, F.; Mercedes Perez, A.; Reynes, M.; Brillouet, J.-M.; Brat, P. The noni fruit (Morinda citrifolia L.): A review of agricultural research, nutritional and therapeutic properties. J. Food Compos. Anal. 2006, 19, 645–654. [Google Scholar] [CrossRef]
- Yang, J.; Paulino, R.; Janke-Stedronsky, S.; Abawi, F. Free-radical-scavenging activity and total phenols of noni (Morinda citrifolia L.) juice and powder in processing and storage. Food Chem. 2007, 102, 302–308. [Google Scholar] [CrossRef]
- Cheng, Y.L.; Li, P.Z.; Hu, B.; Xu, L.; Liu, S.N.; Yu, H.; Guo, Y.H.; Xie, Y.F.; Yao, W.R.; Qian, H. Correlation analysis reveals the intensified fermentation via Lactobacillus plantarum improved the flavor of fermented noni juice. Food Biosci. 2021, 43, 101234. [Google Scholar] [CrossRef]
- Deng, S.X.; West, B.J.; Jensen, C.J. A quantitative comparison of phytochemical components in global noni fruits and their commercial products. Food Chem. 2010, 122, 267–270. [Google Scholar] [CrossRef]
- Motshakeri, M.; Ghazali, H.M. Nutritional, phytochemical and commercial quality of Noni fruit: A multi-beneficial gift from nature. Trends Food Sci. Technol. 2015, 45, 118–129. [Google Scholar] [CrossRef]
- Wang, R.M.; Yao, L.L.; Lin, X.; Hu, X.P.; Wang, L. Exploring the potential mechanism of Rhodomyrtus tomentosa (Ait.) Hassk fruit phenolic rich extract on ameliorating nonalcoholic fatty liver disease by integration of transcriptomics and metabolomics profiling. Food Res. Int. 2022, 151, 110824. [Google Scholar] [CrossRef]
- Wang, Z.; Feng, Y.; Yang, N.; Jiang, T.; Xu, H.; Lei, H. Fermentation of kiwifruit juice from two cultivars by probiotic bacteria: Bioactive phenolics, antioxidant activities and flavor volatiles. Food Chem. 2022, 373, 131455. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ren, B.B.; Chen, C.; Li, C.; Fu, X.; You, L.; Liu, R.H. Optimization of microwave-assisted extraction of Sargassum thunbergii polysaccharides and its antioxidant and hypoglycemic activities. Carbohyd. Polym. 2017, 173, 192–201. [Google Scholar] [CrossRef]
- Mueller, M.; Hobiger, S.; Jungbauer, A. Anti-inflammatory activity of extracts from fruits, herbs and spices. Food Chem. 2010, 122, 987–996. [Google Scholar] [CrossRef]
- Ma, C.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF/MS Characterization of Phenolic Compounds in Palm Fruits (Jelly and Fishtail Palm) and Their Potential Antioxidant Activities. Antioxidants 2019, 8, 483. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Patel, M.; Deshpande, S.; Alreshidi, M.; Siddiqui, A.J.; Reddy, M.N.; Emira, N.; De Feo, V. Effect of Adiantum philippense Extract on Biofilm Formation, Adhesion with Its Antibacterial Activities against Foodborne Pathogens, and Characterization of Bioactive Metabolites: An in vitro-in silico Approach. Front. Microbiol. 2020, 11, 823. [Google Scholar] [CrossRef]
- Li, W.; Yang, R.L.; Ying, D.Y.; Yu, J.W.; Sanguansri, L.; Augustin, M.A. Analysis of polyphenols in apple pomace: A comparative study of different extraction and hydrolysis procedures. Ind. Crops Prod. 2020, 147, 112250. [Google Scholar] [CrossRef]
- Kramberger, K.; Barlic-Maganja, D.; Bandelj, D.; Baruca Arbeiter, A.; Peeters, K.; Miklavcic Visnjevec, A.; Jenko Praznikar, Z. HPLC-DAD-ESI-QTOF-MS Determination of Bioactive Compounds and Antioxidant Activity Comparison of the Hydroalcoholic and Water Extracts from Two Helichrysum italicum Species. Metabolites 2020, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.F.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF/MS Characterization of Phenolic Compounds from Medicinal Plants (Hops and Juniper Berries) and Their Antioxidant Activity. Foods 2020, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Chou, O.; Ali, A.; Subbiah, V.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF-MS/MS Characterisation of Phenolics in Herbal Tea Infusion and Their Antioxidant Potential. Fermentation 2021, 7, 73. [Google Scholar] [CrossRef]
- Guzman, J.D. Natural Cinnamic Acids, Synthetic Derivatives and Hybrids with Antimicrobial Activity. Molecules 2014, 19, 19292–19349. [Google Scholar] [CrossRef]
- Zhong, B.M.; Robinson, N.A.; Warner, R.D.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF-MS/MS Characterization of Seaweed Phenolics and Their Antioxidant Potential. Mar. Drugs 2020, 18, 331. [Google Scholar] [CrossRef]
- Castro-Lopez, C.; Bautista-Hernandez, I.; Gonzalez-Hernandez, M.D.; Martinez-Avila, G.C.G.; Rojas, R.; Gutierrez-Diez, A.; Medina-Herrera, N.; Aguirre-Arzola, V.E. Polyphenolic Profile and Antioxidant Activity of Leaf Purified Hydroalcoholic Extracts from Seven Mexican Persea americana Cultivars. Molecules 2019, 24, 173. [Google Scholar] [CrossRef] [Green Version]
- Pierson, J.T.; Monteith, G.R.; Roberts-Thomson, S.J.; Dietzgen, R.G.; Gidley, M.J.; Shaw, P.N. Phytochemical extraction, characterisation and comparative distribution across four mango (Mangifera indica L.) fruit varieties. Food Chem. 2014, 149, 253–263. [Google Scholar] [CrossRef]
- Li, S.J.; Wang, R.M.; Hu, X.P.; Li, C.F.; Wang, L. Bio-affinity ultra-filtration combined with HPLC-ESI-qTOF-MS/MS for screening potential alpha-glucosidase inhibitors from Cerasus humilis (Bge.) Sok. leaf-tea and in silico analysis. Food Chem. 2022, 373, 131528. [Google Scholar] [CrossRef]
- Zhao, X.S.; Zhang, S.H.; Liu, D.; Yang, M.H.; Wei, J.H. Analysis of Flavonoids in Dalbergia odorifera by Ultra-Performance Liquid Chromatography with Tandem Mass Spectrometry. Molecules 2020, 25, 389. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.J.; Zhang, D.; Yu, C.; Yao, L.; Chen, Z.; Tao, Y.D.; Cao, W.G. Phytochemical Analysis, Antioxidant and Analgesic Activities of Incarvillea compacta Maxim from the Tibetan Plateau. Molecules 2019, 24, 1692. [Google Scholar] [CrossRef] [Green Version]
- Huynh, N.T.; Van Camp, J.; Smagghe, G.; Raes, K. Improved Release and Metabolism of Flavonoids by Steered Fermentation Processes: A Review. Int. J. Mol. Sci. 2014, 15, 19369–19388. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Wen, J.J.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Nie, S.P.; Xiong, T.; Xie, M.Y. Momordica charantia juice with Lactobacillus plantarum fermentation: Chemical composition, antioxidant properties and aroma profile. Food Biosci. 2019, 29, 62–72. [Google Scholar] [CrossRef]
- Li, Z.X.; Teng, J.; Lyu, Y.L.; Hu, X.Q.; Zhao, Y.L.; Wang, M.F. Enhanced Antioxidant Activity for Apple Juice Fermented with Lactobacillus plantarum ATCC14917. Molecules 2019, 24, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dueñas, M.; Fernández, D.; Hernández, T.; Estrella, I.; Muñoz, R. Bioactive phenolic compounds of cowpeas (Vigna sinensis L). Modifications by fermentation with natural microflora and with Lactobacillus plantarum ATCC 14917. J. Sci. Food Agric. 2005, 85, 297–304. [Google Scholar] [CrossRef]
- Yang, S.C.; Chen, T.I.; Li, K.Y.; Tsai, T.C. Change in Phenolic Compound Content, Reductive Capacity and ACE Inhibitory Activity in Noni Juice during Traditional Fermentation. J. Food Drug Anal. 2007, 15, 290–298. [Google Scholar] [CrossRef]
- Li, Q.; Chang, X.; Guo, R.; Wang, Q.; Guo, X. Dynamic effects of fermentation on phytochemical composition and antioxidant properties of wampee (Clausena lansium (Lour.) Skeel) leaves. Food Sci. Nutr. 2019, 7, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cheng, H.; Liu, H.; Ma, R.; Ma, J.; Fang, H. Fermentation by Multiple Bacterial Strains Improves the Production of Bioactive Compounds and Antioxidant Activity of Goji Juice. Molecules 2019, 24, 3519. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Fuhrman, B. Wine flavonoids protect against LDL oxidation and atherosclerosis. Ann. N.Y. Acad. Sci. 2002, 957, 146–161. [Google Scholar] [CrossRef]
- Tejada, S.; Pinya, S.; Bibiloni, M.D.; Tur, J.A.; Pons, A.; Sureda, A. Cardioprotective Effects of the Polyphenol Hydroxytyrosol from Olive Oil. Curr. Drug Targets 2017, 18, 1477–1486. [Google Scholar] [CrossRef]
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Benefits of Nut Consumption on Insulin Resistance and Cardiovascular Risk Factors: Multiple Potential Mechanisms of Actions. Nutrients 2017, 9, 1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terra, X.; Pallares, V.; Ardevol, A.; Blade, C.; Fernandez-Larrea, J.; Pujadas, G.; Salvado, J.; Arola, L.; Blay, M. Modulatory effect of grape-seed procyanidins on local and systemic inflammation in diet-induced obesity rats. J. Nutr. Biochem. 2011, 22, 380–387. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Li, X.L.; Li, T.Y.; Li, M.Y.; Huang, R.M.; Li, W.; Yang, R.L. 3-(4-Hydroxyphenyl)propionic acid, a major microbial metabolite of procyanidin A2, shows similar suppression of macrophage foam cell formation as its parent molecule. Rsc. Adv. 2018, 8, 6242–6250. [Google Scholar] [CrossRef] [Green Version]
- Belwal, T.; Pandey, A.; Bhatt, I.D.; Rawal, R.S.; Luo, Z. Trends of polyphenolics and anthocyanins accumulation along ripening stages of wild edible fruits of Indian Himalayan region. Sci. Rep. 2019, 9, 5894. [Google Scholar] [CrossRef]
- Afanas′ev, I.B.; Dorozhko, A.I.; Brodskii, A.V.; Kostyuk, V.A.; Potapovitch, A.I. Chelating and free radical scavenging mechanisms of inhibitory action of rutin and quercetin in lipid peroxidation. Biochem. Pharmacol. 1989, 38, 1763–1769. [Google Scholar] [CrossRef]
- Zhu, H.L.; Liu, S.X.; Yao, L.L.; Wang, L.; Li, C.F. Free and Bound Phenolics of Buckwheat Varieties: HPLC Characterization, Antioxidant Activity, and Inhibitory Potency towards alpha-Glucosidase with Molecular Docking Analysis. Antioxidants 2019, 8, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| No. | Proposed Compounds | Molecular Formula | RT (min) | m/z [M−H]− | m/z Fragments | FIor (ppm) | 0-FJP | 7-FJP | 28-FJP | 63-FJP |
|---|---|---|---|---|---|---|---|---|---|---|
| Phenolic acids (12) | ||||||||||
| 1 | 2-Hydroxy-2-phenylacetic acid | C8H8O3 | 1.84 | 151.03 | 91, 65 | −0.172 | − | − | + | − |
| 2 | Esculin | C15H16O9 | 2.67 | 339.07 | 177, 159, 133 | −0.06 | − | + | + | − |
| 3 | 4-Hydroxycoumarin | C9H6O3 | 7.99 | 161.02 | 133, 89 | −0.026 | − | − | + | + |
| 4 | Scopoletin | C10H8O4 | 9.42 | 191.03 | 120, 104 | −0.033 | + | − | − | + |
| 5 | 2,4-Dihydroxybenzoic acid | C7H6O4 | 3.13 | 153.01 | 153 | −0.006 | + | + | + | + |
| 6 | 3,4-Dihydroxybenzaldehyde | C7H6O3 | 4.80 | 137.02 | 109, 81 | 0.136 | − | + | − | + |
| 7 | 2-Hydroxy-4-methylbenzaldehyde | C8H8O2 | 4.06 | 135.04 | 135, 89 | 0.064 | − | + | + | − |
| 8 | 3-Hydroxyphenylpropionic acid | C9H10O3 | 3.22 | 165.05 | 119 | −0.851 | + | + | + | − |
| 9 | Cinnamic acid | C9H8O2 | 3.23 | 147.04 | 147, 103 | −0.008 | + | − | − | − |
| 10 | Caffeic acid | C9H8O4 | 3.9 | 179.04 | 1, 358, 979 | 0.076 | + | + | + | + |
| 11 | p-Coumaric acid | C9H8O3 | 6.308 | 163.03 | 133, 92 | −0.081 | + | + | − | − |
| 12 | 2,3-Dihydroxy-1-guaiacylpropanone | C10H12O5 | 4.834 | 211.06 | 137, 109 | −0.038 | − | + | + | + |
| Flavonols (7) | ||||||||||
| 13 | Quercetin 3-O-glucosyl-xyloside | C26H28O16 | 13.10 | 595.13 | 300, 271 | −0.112 | − | + | + | + |
| 14 | Rutin | C27H30O16 | 13.548 | 609.14 | 301, 151 | 0.021 | + | + | + | + |
| 15 | Luteolin 7-neohesperidoside | C27H30O15 | 15.92 | 593.15 | 285, 151 | −0.039 | − | + | + | − |
| 16 | Sativanone | C17H16O5 | 13.58 | 299.09 | 271, 255 | −0.091 | − | − | + | + |
| 17 | Isoquercitrin | C21H20O12 | 13.67 | 463.08 | 300, 271, 151 | −0.007 | + | + | + | + |
| 18 | Quercetin | C15H10O7 | 20.45 | 301.03 | 273, 229, 121 | 0.028 | + | + | + | + |
| 19 | 2’,7-Dihydroxy-4’,5’ -dimethoxyisoflavone | C17H14O6 | 34.84 | 313.07 | 270, 211 | 0.005 | + | + | − | + |
| Other phenolics (2) | ||||||||||
| 20 | Isoscutellarein 7-xyloside | C20H18O10 | 9.05 | 417.08 | 243, 175, 149 | −0.052 | − | − | − | + |
| 21 | Mucronulatol (±) | C17H18O5 | 14.34 | 301.10 | 149, 123 | −0.022 | − | + | + | + |
| Total | 21 | 10 | 15 | 15 | 14 |
| No. | Phenolic Compounds | 0-FJP (mg/L) | 7-FJP (mg/L) | 28-FJP (mg/L) | 63-FJP (mg/L) |
|---|---|---|---|---|---|
| 1 | caffeic acid | 0.96 ± 0.14 c | 1.21 ± 0.02 bc | 1.68 ± 0.04 a | 1.37 ± 0.18 b |
| 2 | 2,4-dihydroxybenzoic acid | 0.75 ± 0.13 c | 1.47 ± 0.11 ab | 1.7 ± 0.08 a | 1.26 ± 0.11 b |
| 3 | p-coumaric acid | 1.55 ± 0.06 c | 3.48 ± 0.02 ab | ND | ND |
| 4 | rutin | 17.67 ± 1.56 b | 20.45 ± 1.32 b | 26.69 ± 2.25 a | 9.25 ± 0.62 c |
| 5 | quercetin | 3.03 ± 0.35 c | 6.4 ± 0.8 b | 10.55 ± 1.36 a | 8.16 ± 1.04 ab |
| 6 | isoquercitrin | 8.25 ± 0.34 a | 6.57 ± 0.36 ab | 5.14 ± 2.10 b | 4.47 ± 0.05 b |
| Samples | DPPH Radical Scavenging Rate (%) | ABTS+ Radical Scavenging Rate(%) | OH∙ Radical Scavenging Rate (%) | FRAP Assay (μmol TE/100 mL) |
|---|---|---|---|---|
| 0-FJP | 60.11 ± 0.02 c | 66.72 ± 1.27 b | 45.22 ± 0.96 d | 877.7 ± 12.87 c |
| 7-FJP | 64.49 ± 0.22 b | 67.59 ± 1.05 b | 52.45 ± 0.95 b | 1013.33 ± 11.98 b |
| 28-FJP | 68 ± 0.29 a | 72.05 ± 1.67 a | 57.41 ± 0.63 a | 1144.01 ± 6.86 a |
| 63-FJP | 63.92 ± 0.53 b | 67.61 ± 1.06 b | 49.71 ± 0.57 c | 927.1 ± 51.14 c |
| Samples | Extract Concentration (ng/mL) | TC | FC | CE/TC (%) |
|---|---|---|---|---|
| Control | 0 | 124.69 ± 0.53 h | 71.98 ± 0.48 f | 42.27 ± 0.17 e |
| ox-LDL | 80 | 231.08 ± 0.94 a | 105.57 ± 1.26 a | 54.31 ± 0.43 a |
| 0-FJP | 10 | 158.31 ± 1.95 d | 91.29 ± 2.06 c | 42.33 ± 0.59 e |
| 50 | 148.79 ± 0.53 e | 89.13 ± 0.21 c | 40.09 ± 0.35 f | |
| 100 | 140.79 ± 0.37 f | 84.75 ± 0.48 d | 39.8 ± 0.29 f | |
| 28-FJP | 10 | 182.58 ± 0.47 b | 90.35 ± 0.58 c | 50.51 ± 0.37 b |
| 50 | 150.63 ± 0.56 e | 81.54 ± 0.42 e | 45.86 ± 0.46 c | |
| 100 | 130.56 ± 0.69 g | 72.82 ± 0.68 f | 44.22 ± 0.28 d | |
| 63-FJP | 10 | 158.58 ± 0.55 d | 94.53 ± 0.44 b | 40.38 ± 0.07 f |
| 50 | 160.33 ± 0.93 d | 86.31 ± 0.94 d | 46.16 ± 0.27 c | |
| 100 | 178.54 ± 0.66 c | 95.19 ± 0.5 b | 46.68 ± 0.09 c |
| Caffeic Acid | 2,4-Dihydroxybenzoic Acid | p-Coumaric Acid | Rutin | Quercetin | Isoquercitrin | |
|---|---|---|---|---|---|---|
| DPPH | 0.876 ** | 0.923 ** | 0.998 ** | 0.52 | 0.905 ** | −0.604 * |
| ABTS+ | 0.698 * | 0.675 * | 0.440 | 0.656 * | 0.783 ** | −0.284 |
| OH· | 0.860 ** | 0.951 ** | 0.973 ** | 0.637 * | 0.856 ** | −0.521 |
| FRAP | 0.831 ** | 0.872 ** | 0.984 ** | 0.747 ** | 0.774 ** | −0.402 |
| CE/TC | 0.684 * | 0.692 * | 0.662 | 0.252 | 0.827 ** | −0.342 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, K.; Dou, R.; Lin, X.; Hu, X.; Wang, Z.; Liu, S.; Li, C.; Li, W. Changes in Phenolic Profiles and Inhibition Potential of Macrophage Foam Cell Formation during Noni (Morinda citrifolia Linn.) Fruit Juice Fermentation. Fermentation 2022, 8, 201. https://doi.org/10.3390/fermentation8050201
Cai K, Dou R, Lin X, Hu X, Wang Z, Liu S, Li C, Li W. Changes in Phenolic Profiles and Inhibition Potential of Macrophage Foam Cell Formation during Noni (Morinda citrifolia Linn.) Fruit Juice Fermentation. Fermentation. 2022; 8(5):201. https://doi.org/10.3390/fermentation8050201
Chicago/Turabian StyleCai, Kun, Rong Dou, Xue Lin, Xiaoping Hu, Zhulin Wang, Sixin Liu, Congfa Li, and Wu Li. 2022. "Changes in Phenolic Profiles and Inhibition Potential of Macrophage Foam Cell Formation during Noni (Morinda citrifolia Linn.) Fruit Juice Fermentation" Fermentation 8, no. 5: 201. https://doi.org/10.3390/fermentation8050201