
Environmental Drivers of Patterns of Plant Diversity Along a Wide Environmental Gradient in Korean Temperate Forests

Abstract
:1. Introduction
2. Materials and Methods
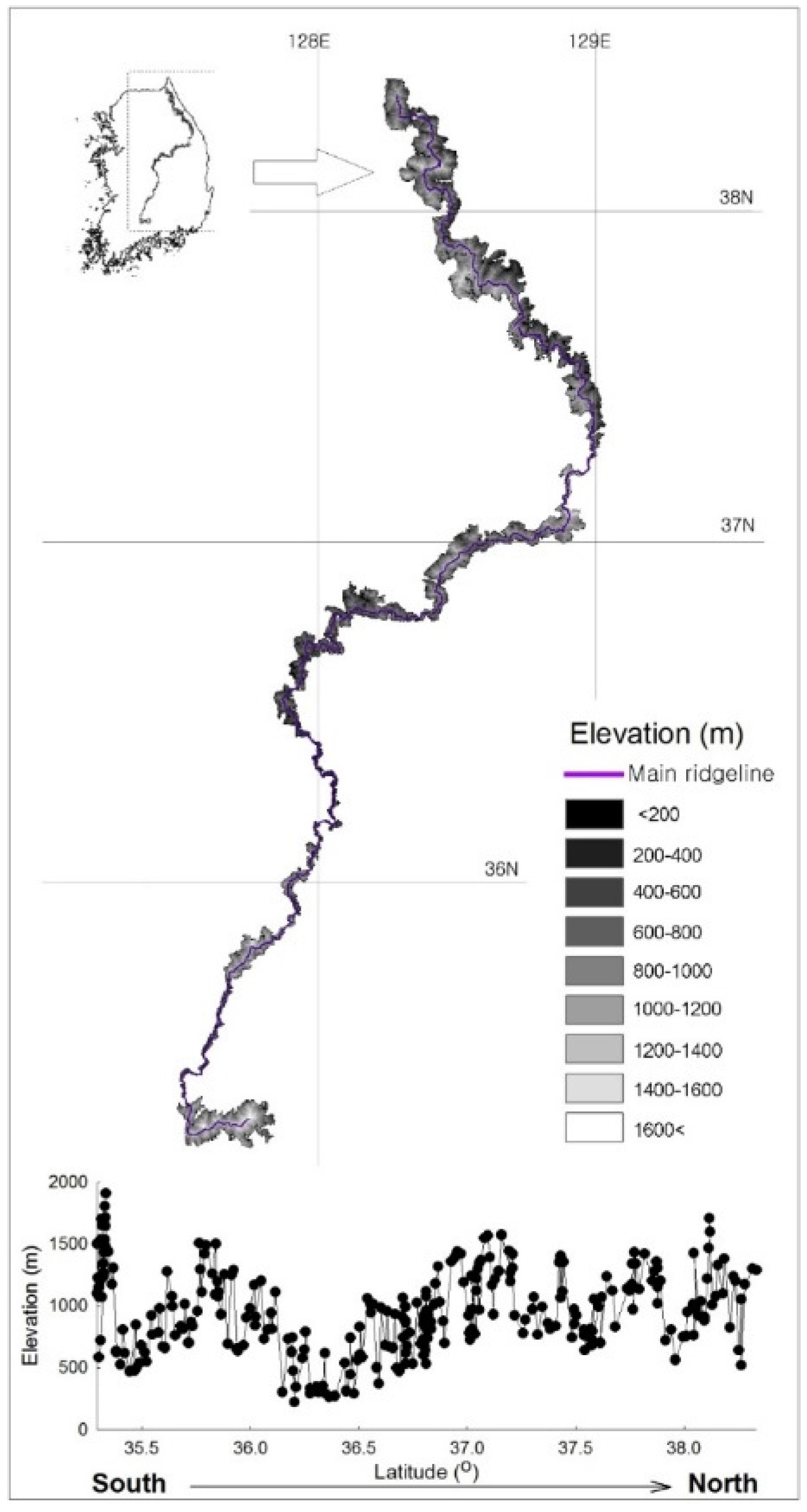
2.1. Study area

2.2. Plant Data and Diversity Indices
2.3. Explanatory Variables
2.4. Data Analysis
3. Results
3.1. General Description

{kind=link}
{kind=link}
{kind=link}
| Elevational Band (m) | Number of Plots | Total | Woody | Herbaceous |
|---|---|---|---|---|
| 200 | 36 | 188 | 86 | 102 |
| 300 | 64 | 193 | 93 | 100 |
| 400 | 64 | 240 | 106 | 134 |
| 500 | 46 | 195 | 88 | 107 |
| 600 | 61 | 218 | 101 | 117 |
| 700 | 93 | 284 | 126 | 158 |
| 800 | 117 | 372 | 138 | 234 |
| 900 | 113 | 364 | 127 | 237 |
| 1000 | 103 | 389 | 133 | 256 |
| 1100 | 72 | 355 | 118 | 237 |
| 1200 | 70 | 366 | 114 | 252 |
| 1300 | 68 | 337 | 111 | 226 |
| 1400 | 75 | 320 | 109 | 211 |
| 1500 | 54 | 239 | 86 | 153 |
| 1600 | 38 | 191 | 73 | 118 |
| 1700 | 26 | 110 | 39 | 71 |
| All bands pooled | 1100 | 802 | 248 | 554 |

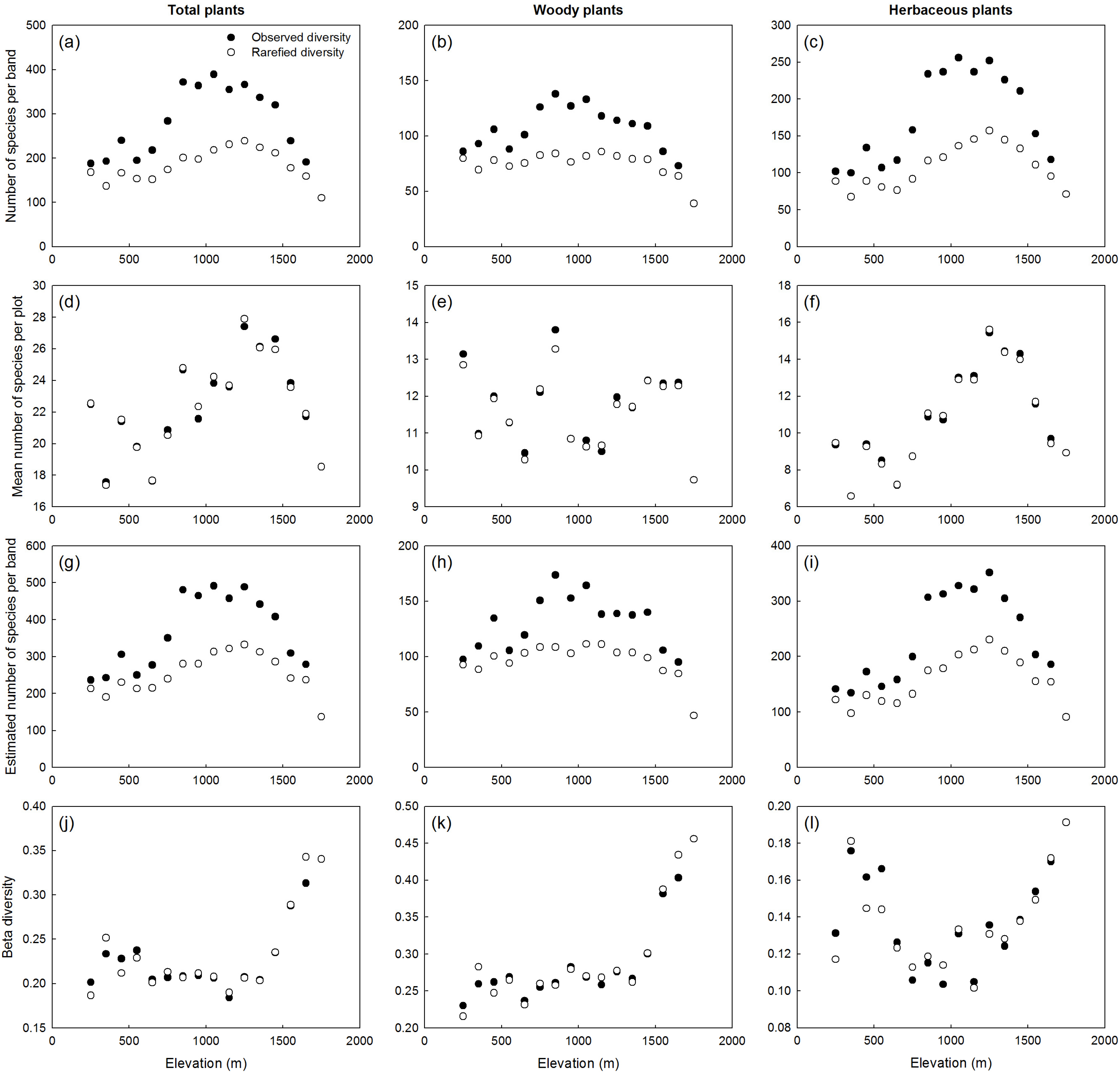
3.2. Elevational Plant Diversity Patterns
| Observed Diversity | Rarefied Diversity | |||
|---|---|---|---|---|
| Linear | Quadratic | Linear | Quadratic | |
| Total | ||||
| NSB | <0.01 | 0.77 *** | 0.04 | 0.57 ** |
| MNSP | 0.16 | 0.33 | 0.15 | 0.34 |
| ENSB | 0.02 | 0.76 *** | 0.04 | 0.64 ** |
| BETAD | 0.34 * | 0.78 *** | 0.38 * | 0.74 *** |
| Woody | ||||
| NSB | 0.08 | 0.84 *** | 0.2 | 0.67 *** |
| MNSP | 0.02 | 0.02 | 0.01 | 0.01 |
| ENSB | 0.05 | 0.79 *** | 0.14 | 0.79 *** |
| BETAD | 0.61 *** | 0.85 *** | 0.61 *** | 0.83 *** |
| Herbaceous | ||||
| NSB | 0.06 | 0.74 *** | 0.17 | 0.59 ** |
| MNSP | 0.28 * | 0.50 ** | 0.27 * | 0.50 * |
| ENSB | 0.07 | 0.72 *** | 0.15 | 0.62 ** |
| BETAD | 0.03 | 0.61 ** | 0.09 | 0.59 ** |

3.3. Plant Diversity Patterns with Explanatory Variables
| logRA | PC1topo | PC1hetero | PC1climate | EVI | PC1vegest | VTD | |
|---|---|---|---|---|---|---|---|
| Observed diversity | |||||||
| Total | |||||||
| NSB | 0.30 * | 0.03 | 0.61 *** | 0.03 | 0.03 | 0.02 | <0.01 |
| MNSP | <0.01 | 0.02 | 0.26 * | 0.04 | 0.10 | <0.01 | <0.01 |
| ENSB | 0.25 | 0.03 | 0.62 *** | 0.02 | 0.01 | 0.01 | <0.01 |
| BETAD | 0.63 *** | 0.14 | 0.30 * | 0.59 *** | 0.26 * | 0.26 * | <0.01 |
| Woody | |||||||
| NSB | 0.66 *** | 0.01 | 0.53 *** | 0.27 * | 0.19 | 0.12 | 0.08 |
| MNSP | <0.01 | 0.16 | <0.01 | 0.03 | 0.04 | 0.20 | 0.31 * |
| ENSB | 0.59 *** | <0.01 | 0.54 *** | 0.20 | 0.17 | 0.10 | 0.10 |
| BETAD | 0.65 *** | 0.31 * | 0.16 | 0.81 *** | 0.36 * | 0.42 ** | 0.03 |
| Herbaceous | |||||||
| NSB | 0.17 | 0.07 | 0.58 *** | <0.01 | <0.01 | <0.01 | 0.01 |
| MNSP | 0.01 | 0.11 | 0.30 * | 0.10 | 0.10 | 0.02 | 0.05 |
| ENSB | 0.13 | 0.06 | 0.58 *** | <0.01 | <0.01 | <0.01 | 0.01 |
| BETAD | 0.39 ** | <0.01 | 0.47 ** | 0.14 | 0.08 | 0.02 | 0.02 |
| Rarefied diversity | |||||||
| Total | |||||||
| NSB | 0.10 | 0.01 | 0.56 *** | <0.01 | <0.01 | 0.01 | <0.01 |
| MNSP | <0.01 | 0.02 | 0.26 * | 0.03 | 0.11 | <0.01 | <0.01 |
| ENSB | 0.13 | 0.02 | 0.63 *** | <0.01 | <0.01 | 0.01 | <0.01 |
| BETAD | 0.60 *** | 0.14 | 0.23 | 0.62 *** | 0.24 | 0.33 * | <0.01 |
| Woody | |||||||
| NSB | 0.52 ** | 0.14 | 0.43 ** | 0.40 ** | 0.10 | 0.28 * | 0.09 |
| MNSP | <0.01 | 0.16 | 0.01 | 0.02 | 0.04 | 0.18 | 0.32 * |
| ENSB | 0.62 *** | 0.06 | 0.58 *** | 0.33 * | 0.14 | 0.17 | 0.10 |
| BETAD | 0.64 *** | 0.28 * | 0.12 | 0.81 *** | 0.36 * | 0.44 ** | 0.01 |
| Herbaceous | |||||||
| NSB | 0.01 | 0.08 | 0.47 ** | 0.03 | 0.04 | 0.01 | 0.04 |
| MNSP | 0.01 | 0.11 | 0.29 * | 0.09 | 0.10 | 0.02 | 0.05 |
| ENSB | 0.03 | 0.07 | 0.53 *** | 0.02 | 0.04 | <0.01 | 0.02 |
| BETAD | 0.43 ** | 0.02 | 0.43 ** | 0.23 | 0.05 | 0.04 | 0.02 |
| Plant Group | Dependent Variable | Regression Equation | F | R2 | P |
|---|---|---|---|---|---|
| Observed diversity | |||||
| Total | NSB | y = 272.563 + 68.748 PC1hetero | 22.04 | 0.61 | <0.001 |
| MNSP | y = 22.352 + 1.572 PC1hetero | 4.81 | 0.26 | 0.046 | |
| ENSB | y = 351.195 + 89.798 PC1hetero | 23.26 | 0.62 | <0.001 | |
| BETAD | y = 0.465 − 0.026 logRA | 24.01 | 0.63 | <0.001 | |
| Woody | NSB | y = 3.793 + 10.891 logRA + 10.264 PC1hetero | 21.88 | 0.77 | <0.001 |
| MNSP | y = 5.603 + 3.229 VTD | 6.20 | 0.31 | 0.026 | |
| ENSB | y = 16.424 + 11.982 logRA + 14.314 PC1hetero | 17.33 | 0.73 | <0.001 | |
| BETAD | y = 0.292 − 0.022 PC1hetero + 0.0002 PC1climate | 89.29 | 0.93 | <0.001 | |
| Herbaceous | NSB | y = 169.563 + 50.018 PC1hetero | 19.15 | 0.58 | 0.001 |
| MNSP | y = 20.886 − 1.114 logRA + 2.345 PC1hetero | 7.15 | 0.52 | 0.008 | |
| ENSB | y = 226.663 + 66.164 PC1hetero | 19.69 | 0.58 | 0.001 | |
| BETAD | y = 0.140 − 0.019 PC1hetero | 12.19 | 0.47 | 0.004 | |
| Rarefied diversity | |||||
| Total | NSB | y = 182.574 + 28.012 PC1hetero | 17.92 | 0.56 | 0.001 |
| MNSP | y = 22.399 + 1.610 PC1hetero | 4.99 | 0.26 | 0.042 | |
| ENSB | y = 252.669 + 43.834 PC1hetero | 24.27 | 0.63 | <0.001 | |
| BETAD | y = 0.233 − 0.022 PC1hetero + 0.0001 PC1climate | 27.56 | 0.81 | <0.001 | |
| Woody | NSB | y = 20.258 + 5.982 logRA | 15.42 | 0.52 | 0.002 |
| MNSP | y = 5.899 + 3.025 VTD | 6.53 | 0.32 | 0.023 | |
| ENSB | y = 41.510 + 6.047 logRA + 7.518 PC1hetero | 22.47 | 0.78 | <0.001 | |
| BETAD | y = 0.294 − 0.022 PC1hetero + 0.0002 PC1climate | 61.67 | 0.91 | <0.001 | |
| Herbaceous | NSB | y = 107.844 + 20.365 PC1hetero | 12.31 | 0.47 | 0.003 |
| MNSP | y = 20.438 − 1.068 logRA + 2.289 PC1hetero | 6.49 | 0.50 | 0.011 | |
| ENSB | y = 157.311 + 32.458 PC1hetero | 15.89 | 0.53 | 0.001 | |
| BETAD | y = 0.137 − 0.016 PC1hetero + 0.00004 PC1climate | 10.92 | 0.63 | 0.002 |
4. Discussion
4.1. Elevational Plant Diversity Patterns
4.2. Determinants of Elevational Diversity Patterns
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Grytnes, J.A.; Vetaas, O.R. Species richness and altitude: A comparison between null models and interpolated plant species richness along the Himalayan altitudinal gradient, Nepal. Am. Nat. 2002, 159, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Ma, K.M.; Anand, M.; Fu, B.J. Do generalized scaling laws exist for species abundance distribution in mountains? Oikos 2006, 115, 81–88. [Google Scholar] [CrossRef]
- Acharya, B.K.; Chettri, B.; Vijayan, L. Distribution pattern of trees along an elevational gradient of Eastern Himalaya, India. Acta Oecologica 2011, 37, 329–336. [Google Scholar] [CrossRef]
- Swenson, N.G.; Erickson, D.L.; Mi, X.; Bourg, N.A.; Forero-Montaña, J.; Ge, X.; Howe, R.; Lake, J.K.; Liu, X.; Ma, K.; et al. Phylogenetic and functional diversity alpha and beta diversity in temperate and tropical tree communities. Ecology 2012, 93, S112–S125. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Borcard, B.; Peres-Neto, P.R. Analyzing beta diversity: Partitioning the spatial variation of community composition data. Ecol. Monogr. 2005, 75, 435–450. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Field, R.; Hawkins, B.A.; Cornell, H.V.; Currie, D.J.; Diniz-Filho, J.A.F.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; et al. Spatial species-richness gradients across scales: A meta-analysis. J. Biogeogr. 2009, 36, 132–147. [Google Scholar] [CrossRef]
- Gentili, R.; Armiraglio, S.; Sgorbati, S.; Baroni, C. Geomorphological disturbance affects ecological driving forces and plant turnover along an altitudinal stress gradient on alpine slopes. Plant Ecol. 2013, 214, 571–586. [Google Scholar] [CrossRef]
- Legendre, P.; Dale, M.R.T.; Fortin, M.J.; Casgrain, P.; Gurevitch, J. Effects of spatial structures on the results of field experiments. Ecology 2004, 85, 3202–3214. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Grau, O.; Grytnes, J.A.; Birks, H.J.B. A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya. J. Biogeogr. 2007, 34, 1907–1915. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, Z.; Fang, J. Altitudinal patterns of seed plant richness in the Gaoligong Mountains, south-east Tibet, China. Divers. Distrib. 2007, 13, 845–854. [Google Scholar] [CrossRef]
- Rowe, R.J. Environmental and geometric drivers of small mammal diversity along elevational gradients in Utah. Ecography 2009, 32, 411–422. [Google Scholar] [CrossRef]
- Wu, Y.; Colwell, R.K.; Rahbek, C.; Zhang, C.; Quan, Q.; Wang, C.; Lei, F. Explaining the species richness of birds along a subtropical elevational gradient in the Hengduan Mountains. J. Biogeogr. 2013, 40, 2310–2323. [Google Scholar] [CrossRef]
- Sanders, N.J. Elevational gradients in ant species richness: Area, geometry, and Rapoport’s rule. Ecography 2002, 25, 25–32. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C. Geographic range size and determinants of avian species richness. Science 2002, 297, 1548–1551. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 280–338. [Google Scholar] [CrossRef]
- Sfenthourakis, S.; Panitsa, M. From plots to islands: Species diversity at different scales. J. Biogeogr. 2012, 39, 750–759. [Google Scholar] [CrossRef]
- Lomolino, M.V. Elevational gradients of species-density: Historical and prospective views. Glob. Ecol. Biogeogr. 2001, 10, 3–13. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.S. Biogeography of Korea Plants, 2nd ed.; GeoBook Publishing: Seoul, Korea, 2007. (In Korean) [Google Scholar]
- Korea Forest Research Institute. Ecological Aspects of Baekdu Mountains in Korea and Delineation of Their Management and Conservation area; Korea Forest Research Institute: Seoul, Korea, 2003. (In Korean) [Google Scholar]
- Yun, J.I. Agroclimatic maps augmented by a GIS technology. Korean J. Agric. For. Meteorol. 2010, 12, 63–73. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Plant Sociology; Hafner Publishing Company: New York, NY, USA, 1965. [Google Scholar]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef]
- Colwell, R.K. Estimate S: Statistical Estimation of Species Richness and Shared Species from Samples. Available online: http://purl.oclc.org/estimates (accessed on 20 September 2015).
- Speckmann, B.; Snoeyink, J. Easy triangle strips for TIN terrain models. Int. J. Geograph. Inf. Sci. 2001, 15, 379–386. [Google Scholar]
- Jenness, J.S. Calculating landscape surface area from digital elevation models. Wildl. Soc. Bull. 2004, 32, 829–839. [Google Scholar] [CrossRef]
- Jenness, J.S. Topogrphic Position Index (tpi_jen.avx) Extension for ArcView 3.x. Available online: http://www.jennessent.com/arcview/tpi.htm (accessed on 10 September 2015).
- Adhikari, D.; Barik, S.K.; Upadhaya, K. Habitat distribution modeling for reintroduction of Ilex khasiana Purk., a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef]
- Oommen, M.A.; Shanker, K. Elevational species richness patterns emerge from multiple local mechanisms in Himalayan woody plants. Ecology 2005, 86, 3039–3047. [Google Scholar] [CrossRef]
- Černý, T.; Doležal, J.; Janeček, Š.; Šrůtek, M.; Valachovič, M.; Petřík, P.; Altman, J.; Bartoš, M.; Song, J.S. Environmental correlates of plant diversity in Korean temperature forests. Acta Oecologica 2013, 47, 37–45. [Google Scholar] [CrossRef]
- Lee, C.B.; Cho, H.J.; Chun, J.H.; Song, H.K.; Kim, H.H. Variations in species and functional plant diversity among forest types on the ridge of the Baekdudaegan Mountains, South Korea. J. Agric. Life Sci. 2013, 47, 147–162. [Google Scholar]
- Sang, W. Plant diversity patterns and their relationships with soil and climatic factors along an altitudinal gradient in the middle Tianshan Mountain area, Xinjiang, China. Ecol. Res. 2009, 24, 303–314. [Google Scholar] [CrossRef]
- Lee, C.B.; Chun, J.H.; Ahn, H.H. Elevational patterns of plant richness and their drivers on an Asian mountain. Nord. J. Bot. 2014, 32, 347–357. [Google Scholar] [CrossRef]
- Koleff, P.; Lennon, J.J.; Gaston, K.J. Are there latitudinal gradients in species turnover? Glob. Ecol. Biogeogr. 2003, 12, 483–498. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. A latitudinal gradient in large-scale beta diversity for vascular plants in North America. Ecol. Lett. 2007, 10, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Begon, M.; Harper, J.L.; Townsend, C.R. Ecology: Individuals, Populations and Communities; Blackwell Science: Oxford, UK, 1990. [Google Scholar]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhou, G.; Yang, L.; Li, Z. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slope of Qilianshan Mountans, Gansu, China. Plant Ecol. 2002, 165, 169–181. [Google Scholar] [CrossRef]
- Peterken, G.F.; Game, M. Historical factors affecting the number and distribution of vascular plant species in the woodlands of central Lincolnshire. J. Ecol. 1984, 72, 155–182. [Google Scholar] [CrossRef]
- Singleton, R.; Gardescu, S.; Marks, P.L.; Geber, M.A. Forest herb colonization of postagricultural forests in central New York State, USA. J. Ecol. 2001, 89, 325–338. [Google Scholar] [CrossRef]
- Webb, C.O. Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Romdal, T.S.; Grytnes, J.A. An indirect area effect on elevational species richness patterns. Ecography 2007, 30, 440–448. [Google Scholar] [CrossRef]
- Moeslund, J.E.; Arge, L.; Bøcher, P.K.; Nygaard, B.; Svenning, J.C. Geographically comprehensive assessment of salt-meadow vegetation-elevation relations using LiDAR. Wetlands 2011, 31, 471–482. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. Ecological Niches: Linking Classical and Contemporary Approaches; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Vivian-Smith, G. Microtopographic heterogeneity and floristic diversity in experimental wetland communities. J. Ecol. 1997, 85, 71–82. [Google Scholar] [CrossRef]
- Laliberté, E.; Grace, J.B.; Huston, M.A.; Lambers, H.; Teste, F.P.; Turner, B.L.; Wardle, D.A. How does pedogenesis drive plant diversity? Trends Ecol. Evol. 2013, 28, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Moeslund, J.E.; Arge, L.; Bøcher, P.K.; Dalgaard, T.; Svenning, J.C. Topography as a driver of local terrestrial vascular plant diversity patterns. Nord. J. Bot. 2013, 31, 129–144. [Google Scholar] [CrossRef]
- Bledsoe, B.P.; Shear, T.H. Vegetation along hydrologic and edaphic gradients in a North Carolina coastal plain creek bottom and implications for restoration. Wetlands 2000, 20, 126–147. [Google Scholar] [CrossRef]
- Hofer, G.; Bunce, R.G.H.; Edwards, P.J.; Szerencsits, E.; Wagner, H.H.; Herzog, F. Use of topographica variability for assessing plant diversity in agricultural landscapes. Agric. Ecosyst. Environ. 2011, 142, 144–148. [Google Scholar] [CrossRef]
- Chiarucci, A.; de Dominicis, V.; Wilson, J.B. Structure and floristic diversity in permanent monitoring plots in forest ecosystems of Tuscany. For. Ecol. Manag. 2001, 141, 201–210. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Guisan, A.; Vittoz, P.; Wohlgemuth, T.; Zimmermann, N.E.; Dullinger, S.; Pauli, H.; Willner, W.; Grytnes, J.A.; et al. Cross-scale analysis of the region effect on vascular plant species diversity in southern and northern European mountain ranges. PLoS ONE 2010, 5, e15743. [Google Scholar] [CrossRef] [PubMed]
- Novillo, A.; Ojeda, R.A. Elevational patterns in rodent diversity in the dry Andes: Disentangling the role of environmental factors. J. Mammal. 2014, 95, 99–107. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- McCain, C.M. The mid-domain effect applied to elevational gradients: Species richness of small mammals in Costa Rica. J. Biogeogr. 2004, 31, 19–31. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.-B.; Chun, J.-H. Environmental Drivers of Patterns of Plant Diversity Along a Wide Environmental Gradient in Korean Temperate Forests. Forests 2016, 7, 19. https://doi.org/10.3390/f7010019
Lee C-B, Chun J-H. Environmental Drivers of Patterns of Plant Diversity Along a Wide Environmental Gradient in Korean Temperate Forests. Forests. 2016; 7(1):19. https://doi.org/10.3390/f7010019
Chicago/Turabian StyleLee, Chang-Bae, and Jung-Hwa Chun. 2016. "Environmental Drivers of Patterns of Plant Diversity Along a Wide Environmental Gradient in Korean Temperate Forests" Forests 7, no. 1: 19. https://doi.org/10.3390/f7010019