Wood Quality and Growth Characterization across Intra- and Inter-Specific Hybrid Aspen Clones

Abstract
:1. Introduction
1.1. Populus tremuloides
1.2. Aspen Breeding
1.3. Heterosis
1.4. Study Objectives
2. Experimental Section
2.1. Sample Procurement

{kind=link}
| Genetics classification | Species | Abbreviation | Mother Origin | Father Origin |
|---|---|---|---|---|
| Intra-specific (OP) | P. tremuloides × P. tremuloides | NN | 55°25’ N, 113°21’ W | - |
| Intra-specific (FS) | P. tremula × P. tremula | EE | 60°32’ N, 24°29’ E | 60°37’ N, 24°31’ E |
| Intra-specific (OP) | P. davidiana × P. davidiana | CC | 47° N, 128° E | - |
| Inter-specific (FS) | P. tremula × P. tremuloides | EN | 60°32’ N, 24°15’ E | 54°50’ N, 119°32’ W |
| Inter-specific (FS) | P. tremuloides × P. davidiana | CN | 55°7’ N, 111°28’ W | 47° N, 128° E |
2.2. Growth Traits and Core Sampling

2.3. Wood Density
2.4. Microfibril Angle
2.5. Fiber Length, Width, and Coarseness
2.6. Cell Wall Chemistry
2.7. Syringyl-Guaiacyl Ratio
2.8. Data Analysis

3. Results and Discussion
3.1. Models
| Proc GLM | Kolmogorov-Smirnov | Bartlett | |||
|---|---|---|---|---|---|
| Trait | R-squared | RMSE | F-value | D-statistic | Chi-Square |
| (P-value) | (P-value) | (-value) | |||
| MFA | 0.12 | 1.07 | 5.18 | 0.07897 | 3.2338 |
| (0.001) | (0.058) | (0.520) | |||
| D | 0.10 | 29.25 | 4.86 | 0.056371 | 8.8251 |
| (0.001) | ( >0.150) | (0.066) | |||
| FL | 0.41 | 0.05 | 22.06 | 0.055896 | 2.1548 |
| (<0.001) | ( >0.150) | (0.707) | |||
| FW | 0.13 | 1.60 | 4.66 | 0.058263 | 8.3866 |
| (0.002) | (>0.150) | (0.078) | |||
| CS | 0.11 | 0.23 | 3.63 | 0.054734 | 3.1562 |
| (0.008) | ( >0.150) | (0.532) | |||
| S:G | 0.52 | 0.25 | 47.71 | 0.058548 | 6.7663 |
| (<0.001) | (0.132) | (0.149) | |||
| VOL | 0.47 | 0.000746 | 34.28 | 0.051991 | 14.84 |
| (<0.001) | (>0.150) | (0.005) | |||
| Trait | R-squared | RMSE | F-value | D-statistic | Chi-Square |
| (P-value) | (P-value) | (-value) | |||
| TC | 0.10 | 4.53 | 4.42 | 0.055846 | 9.9356 |
| (0.002) | (>0.150) | (0.042) | |||
| TL | 0.13 | 1.14 | 6.27 | 0.041626 | 3.9646 |
| (<0.001) | (>0.150) | (0.411) | |||
| SOL | 0.36 | 0.51 | 21.37 | 0.0556349 | 12.9529 |
| (<0.001) | (>0.150) | (0.012) | |||
| INSOL | 0.32 | 1.1459 | 18.04 | 0.047491 | 3.6516 |
| (<0.001) | (>0.150) | (0.455) | |||
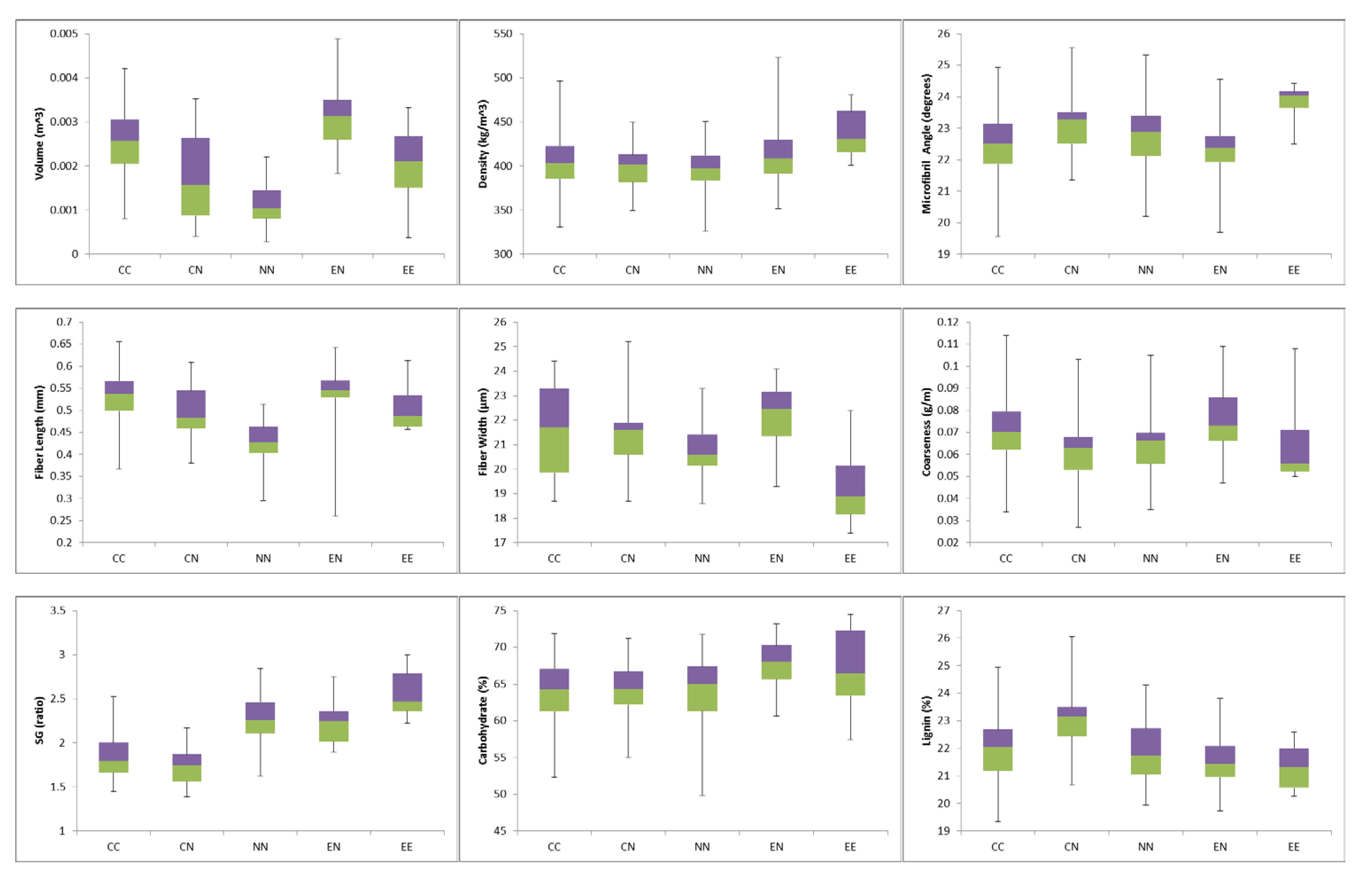
3.2. Least Squares Means Comparisons
3.2.1. Volume
| Cross Type 1 | D 2 | MFA | FL | FW | CS | S:G | VOL | TC | TL | SOL | INSOL |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | 407.10 | 22.5 | 0.531 | 21.7 | 0.072 | 1.85 | 0.00253 | 64.12 | 22.01 | - | - |
| (±3.78) | (±0.1) | (±0.007) | (±0.2) | (±0.002) | (±0.03) | (±0.00011) | (±0.61) | (±0.15) | - | - | |
| CN | 399.10 | 23.1 | 0.496 | 21.4 | 0.063 | 1.74 | 0.00173 | 64.50 | 22.93 | 2.51 | 20.37 |
| (±5.43) | (±0.2) | (±0.012) | (±0.4) | (±0.003) | (±0.05) | (±0.00015) | (±0.91) | (±0.23) | (±0.07) | (±0.23) | |
| EE | 436.22 | 23.7 | 0.511 | 19.4 | 0.068 | 2.56 | 0.00203 | 66.63 | 21.36 | - | - |
| (±8.44) | (±0.4) | (±0.027) | (±0.8) | (±0.008) | (±0.07) | (±0.00014) | (±1.31) | (±0.33) | - | - | |
| EN | 413.33 | 22.2 | 0.537 | 22.2 | 0.075 | 2.22 | 0.00311 | 67.69 | 21.59 | 3.42 | 18.12 |
| (±5.34) | (±0.2) | (±0.011) | (±0.3) | (±0.003) | (±0.05) | (±0.00014) | (±0.83) | (±0.21) | (±0.10) | (±0.19) | |
| NN | 398.72 | 22.8 | 0.428 | 20.7 | 0.065 | 2.29 | 0.00114 | 63.82 | 21.87 | 2.79 | 19.04 |
| (±4.14) | (±0.2) | (±0.010) | (±0.3) | (±0.003) | (±0.04) | (±0.00012) | (±0.66) | (±0.17) | (±0.09) | (±0.19) |
| Cross Type 1 | D 2 | MFA | FL | FW | CS | S:G | VOL | TC | TL | SOL | INSOL |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NN vs CC | −1.497 | 1.225 | −8.275 | −2.545 | −2.048 | 9.152 | −8.345 | −1.919 | −0.651 | - | - |
| (0.136) | (0.222) | (<0.001) | (0.012) | (0.043) | (<0.001) | (<0.001) | (0.057) | (0.516) | - | - | |
| NN vs EE | −3.988 | −2.471 | −2.904 | 1.539 | −0.085 | −3.302 | −4.740 | −0.336 | 1.379 | - | - |
| (<0.001) | (0.015) | (0.004) | (0.126) | (0.932) | (0.001) | (<0.001) | (0.737) | (0.170) | - | - | |
| CC vs EE | −3.148 | −3.282 | 0.759 | 2.738 | 0.840 | −8.898 | 2.707 | −1.742 | 1.806 | - | - |
| (0.002) | (0.001) | (0.449) | (0.007) | (0.403) | (<0.001) | (0.008) | (0.083) | (0.073) | - | - | |
| EN vs NN | 2.163 | −2.010 | 8.229 | 3.317 | 2.328 | −1.10323 | 10.766 | 3.659 | −1.032 | 4.922 | −3.245 |
| (0.032) | (0.046) | (<0.001) | (0.001) | (0.022) | (0.271) | (<0.001) | (<0.001) | (0.304) | (<0.001) | (<0.001) | |
| EN vs EE | −2.291 | −3.711 | 1.323 | 3.223 | 1.105 | −3.853 | 5.449 | 0.686 | 0.599 | - | - |
| (0.023) | (<0.001) | (0.188) | (0.002) | (0.272) | (0.002) | (<0.001) | (0.493) | (0.550) | - | - | |
| CN vs NN | 0.056 | 1.300 | 4.553 | 1.402 | −1.036 | −9.373 | 3.023 | 0.606 | 3.782 | −1.679 | 4.460 |
| (0.955) | (0.196) | (<0.001) | (0.163) | (0.302) | (<0.001) | (0.003) | (0.546) | (<0.001) | (0.095) | (<0.001) | |
| CN vs CC | −1.210 | 2.547 | −2.326 | −0.713 | −2.976 | −1.925 | −4.212 | 0.347 | 3.357 | - | - |
| (0.228) | (0.012) | (0.022) | (0.477) | (0.004) | (0.056) | (<0.001) | (0.729) | (0.001) | - | - |

3.2.2. Wood Density
3.2.3. Microfibril Angle
3.2.4. Fiber Length, Width, and Coarseness
3.2.5. Syringyl-Guaiacyl Ratio
3.2.6. Total Carbohydrate Concentration
3.2.7. Total Lignin Concentration
3.3. Correlations
| D | FL | FW | CS | S:G | TL | TC | SOL | INSOL | DBH | |
|---|---|---|---|---|---|---|---|---|---|---|
| MFA | 0.09 | −0.02 | −0.27 | −0.06 | −0.1 | −0.02 | −0.02 | - | - | - |
| (−0.26) | (0.83) | (0.04) | (0.55) | (0.2) | (0.77) | (0.81) | ||||
| D | 0.2 | −0.25 | 0.19 | 0.11 | −0.11 | 0.18 | - | - | - | |
| (0.02) | (0.04) | (0.03) | (0.14) | (0.14) | (0.02) | |||||
| FL | 0.4 | 0.45 | −0.11 | −0.2 | 0.12 | - | - | - | ||
| (<0.001) | (<0.001) | (0.2) | (0.03) | (0.18) | ||||||
| FW | 0.4 | (0.02) | −0.19 | 0.09 | - | - | - | |||
| (<0.001) | (0.83) | (0.03) | (0.34) | |||||||
| CS | 0.06 | −0.15 | 0.05 | - | - | - | ||||
| (0.48) | (0.09) | (0.59) | ||||||||
| S:G | −0.39 | 0.3 | 0.43 | −0.58 | - | |||||
| (<0.001) | (<0.001) | (<0.001) | (<0.001) | |||||||
| TL | −0.19 | - | - | - | ||||||
| (0.01) | ||||||||||
| HT | 0.88 | |||||||||
| (<0.001) |
3.4. Heterosis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Zsuffa, L.E.; Giordano, L.D.; Stettler, R.F. Trends in poplar culture: Some global and regional perspectives. In Biology of Populus and Its Implications for Management and Conservation; Stettler, R.F., Bradshaw, H.D., Heilman, P.E., Hinckley, T.M., Eds.; NRC Research Press: Ottawa, Canada, 1996; pp. 515–539. [Google Scholar]
- Peterson, E.B.; Peterson, N.M. Aspen [Populus tremuloides] managers’ handbook for British Columbia. FRDA Rep. Vic.BC 1995, 230, 1–36. [Google Scholar]
- Einspahr, D.W.; Wyckoff, G.W. North American aspen: Timber supply, utilization, and researc. North. J. Appl. For. 1990, 7, 168–171. [Google Scholar]
- Einspahr, D.W.; Benson, M.K. Fiber and pulp properties of triploid and triploid hybrid aspen. Tappi J. 1970, 53, 1853–1856. [Google Scholar]
- Peterson, E.B.; Peterson, N.M. Ecology, management, and use of aspen and balsam poplar in the prairie provinces, Canada. Can. For. Service 1992, 1, 252. [Google Scholar]
- Cisneros, N.; Belanger, L.; Gee, W.Y.; Watson, P.A.; Hatton, J.V. Wood and fiber properties of hybrid poplars from Southern British Columbia. Tappi. Proc. 2000, 83, 60. [Google Scholar]
- Ondro, W.J. Utilization and Market Potential of Poplar in Alberta; Northern Forestry Centre, Forestry Canada: Edmonton, Canada, 1989; pp. 271–274. [Google Scholar]
- Mansfield, S.D. Solutions for dissolution—Engineering cell walls for deconstruction. Curr. Opin.Biotechnol. 2009, 20, 286–294. [Google Scholar] [CrossRef]
- Einspahr, D.W. Production and utilization of triploid hybrid aspen. Iowa State J. Resour. 1984, 58, 401–409. [Google Scholar]
- Zsuffa, L. A summary review of interspecific breeding in the genus Populus L. In Proceedings of the 14th meeting of the Canadian Tree Improvement Association, Part 2, Fredericton, New Brunswick, Canada, 28–30 August 1973; pp. 107–123.
- Einspahr, D.W.; Winton, L.L. Genetics of quaking aspen. Aspen Bibliogr. 1976, Paper No. 5026, 1–23. [Google Scholar]
- Li, B.; Wyckoff, G.W.; Einspahr, D.W. Hybrid aspen performance and genetic gains. North. J. Appl. For. 1993, 10, 117–122. [Google Scholar]
- Melchior, G.H. Breeding of aspen and hybrid aspen and their importance for practical use. Allg. For. und Jagdztg. 1985, 156, 112–122. [Google Scholar]
- Kailong, L.; Fangchun, Z.; Guorong, B.; Jia, S. Research advances in genetics and breeding of Populus davidiana Dode in China. J. For. Res. 1999, 10, 25–30. [Google Scholar] [CrossRef]
- Yu, Q.; Tigerstedt, P.M.A.; Haapanen, M. Growth and phenology of hybrid aspen clones (Populustremula L. × Populus tremuloides Michx.). Silv. Fenn. 2001, 35, 15–25. [Google Scholar]
- Tullus, A.; Rytter, L.; Tullus, T.; Weih, M.; Tullus, H. Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scand. J. For. Res. 2012, 27, 10–29. [Google Scholar] [CrossRef]
- Aziz, S.; Wyckoff, G.W.; Wyckoff, J.L. Wood and Pulp Properties of Aspen and Its Hybrids,1996 Pulping Conference Proceedings, Tappi, Norcross, GA, USA; 1996; pp. 437–444.
- Farmer, R.E. Genetic-improvement of poplar in Western Canada-alternatives, opportunities, and pitfalls. In Aspen Management for the 21st Century; Navratil, S., Chapman, P.B., Eds.; Forestry Canada, Northwest Region and Poplar Council of Canada: Edmonton, Alberta, Canada, 1991; p. 174. [Google Scholar]
- Li, B. Aspen improvement strategies for Western Canada-Alberta and Saskatchewan. For. Chron. 1995, 71, 720–724. [Google Scholar]
- Heimburger, C. Report on poplar hybridization. For. Chron. 1936, 12, 285–290. [Google Scholar]
- Stettler, R.F.; Fenn, R.C.; Heilman, P.E.; Stanton, B.J. Populus trichocarpa × Populus deltoides hybrids for short rotation culture: Variation patterns and 4-year field performance. Can. J. For. Res. 1988, 18, 745–753. [Google Scholar] [CrossRef]
- Wu, R.L.; Wang, M.X.; Huang, M.R. Quantitative genetics of yield breeding for Populus short rotation culture. I. Dynamics of genetic control and selection model of yield traits. Can. J. For. Res. 1992, 22, 175–182. [Google Scholar] [CrossRef]
- Falconer, D.S. Introduction to Quantitative Genetics; Longman Scientific and Technical: London, UK, 1989; Volume 3. [Google Scholar]
- Baranwal, V.K.; Mikkilineni, V.; Zehr, U.B.; Tyagi, A.K.; Kapoor, S. Heterosis: Emerging ideas about hybrid vigour. J. Exp. Bot. 2012, 63, 6309–6314. [Google Scholar] [CrossRef]
- Li, B.; Wu, R. Genetic causes of heterosis in juvenile aspen: A quantitative comparison across intra- and inter-specific hybrids. Theor. Appl. Genet. 1996, 93, 380–391. [Google Scholar] [CrossRef]
- Li, B.; Howe, G.T.; Wu, R. Developmental factors responsible for heterosis in aspen hybrids (Populus tremuloides × P. tremula). Tree Physiol. 1998, 18, 37–43. [Google Scholar] [CrossRef]
- British Columbia Forest Inventory Division. Whole Stem Cubic Metre Volume Equations and Tables:Centimetre Diameter Class Merchantable Volume Factors; Forest Inventory Division, British Columbia Forest Service, Department of Forests: Victoria, British Columbia, Canada, 1976. [Google Scholar]
- Robinson, A.R.; Mansfield, S.D. Rapid analysis of poplar lignin monomer composition by a streamlined thioacidolysis procedure and near-infrared reflectance-based prediction modeling. Plant J. 2009, 58, 706–714. [Google Scholar] [CrossRef]
- Junkkonen, R.; Heräjärvi, H. Physical Properties of European and Hybrid Aspen Wood after Three Different Drying Treatments. In Proceedings of the 5th International Symposium of Wood Structure and Properties, Zvolen, Slovakia, 3–6 September 2006; pp. 257–263.
- Bjurhager, I.; Berglund, L.A.; Bardage, S.L.; Sundberg, B. Mechanical characterization of juvenile European aspen (Populus tremula) and hybrid aspen (Populus tremula × Populus tremuloides) using full-field strain measurements. J. Wood Sci. 2008, 54, 349–355. [Google Scholar] [CrossRef]
- Kärki, T. Variation of wood density and shrinkage in European aspen (Populus tremula). Eur. J. Wood Wood Prod. 2001, 59, 79–84. [Google Scholar] [CrossRef]
- Yanchuk, A.; Dancik, B.; Micko, M. Intraclonal variation in wood density of trembling aspen in Alberta. Wood Fiber Sci. 1983, 15, 387–394. [Google Scholar]
- Tsoumis, G. Science and Technology of Wood. Structure, Properties, Utilization; Van Nostrand Reinhold: New York, NY, USA, 1991; pp. xiii–494. [Google Scholar]
- Yanchuk, A.; Micko, M. Radial variation of wood density and fibre length in trembling aspen. IAWA Bull. 1990, 11, 211–215. [Google Scholar]
- Horvath, L.; Peralta, P.; Peszlen, I.; Csoka, L.; Horvath, B.; Jakes, J. Modeling hygroelastic properties of genetically modified aspen. Wood Fiber Sci. 2012, 44, 22–35. [Google Scholar]
- Maloney, V.J.; Mansfield, S.D. Characterization and varied expression of a membrane-bound endo-β-1,4-glucanase in hybrid poplar. Plant Biotechnol. J. 2010, 8, 294–307. [Google Scholar] [CrossRef]
- Barnett, J.R.; Bonham, V.A. Cellulose microfibril angle in the cell wall of wood fibres. Biolog. Rev. 2004, 79, 461–472. [Google Scholar] [CrossRef]
- Fang, S.; Yang, W.; Tian, Y. Clonal and within-tree variation in microfibril angle in poplar clones. New For. 2006, 31, 373–383. [Google Scholar] [CrossRef]
- Francis, R.C.; Hanna, R.B.; Shin, S.J.; Brown, A.F.; Riemenschneider, D.E. Papermaking characteristics of three Populus clones grown in the North-Central United States. Biomass Bioenerg. 2006, 30, 803–808. [Google Scholar] [CrossRef]
- Groover, A.T.; Nieminen, K.; Helariutta, Y.; Mansfield, S.D. Wood formation in Populus. Genet. Genomics Populus 2010, 8, 201–224. [Google Scholar]
- Yanchuk, A.D.; Dancik, B.P.; Micko, M.M. Variation and heritability of wood density and fiber length of trembling aspen in Alberta, Canada. Silvae Genet. 1984, 33, 11–16. [Google Scholar]
- Amidon, T.E. Effect of the wood properties of hardwoods on Kraft paper properties. Tappi J. 1981, 64, 123–126. [Google Scholar]
- Horn, R.A. Morphology of Pulp Fiber from Hardwoods and Influence on Paper Strength; Forest Products Lab: Madison, WI, USA, 1978; pp. 1–9. [Google Scholar]
- Paavilainen, L. Conformability, flexibility and collapsibility of sulphate pulp fibres. Paperi ja puu 1993, 75, 689–702. [Google Scholar]
- Via, B.K.; Stine, M.; Shupe, T.F.; So, C.L.; Groom, L. Genetic improvement of fiber length and coarseness based on paper product performance and material variability: A review. IAWA J. 2004, 25, 401–414. [Google Scholar] [CrossRef]
- Karaim, B.W.; Wengert, E.M.; Szabo, T.; Adams, R.D. Opportunities for aspen and balsam poplar utilization in Alberta. Aspen Bibliogr. 1990, Paper No. 2929, 1–27. [Google Scholar]
- Mansfield, S.D.; Weineisen, H. Wood fiber quality and Kraft pulping efficiencies of trembling aspen (Populus tremuloides Michx.) clones. J. Wood Chem. Technol. 2007, 27, 135–151. [Google Scholar] [CrossRef]
- Stewart, J.J.; Kadla, J.F.; Mansfield, S.D. The influence of lignin chemistry and ultrastructure on the pulping efficiency of clonal aspen (Populus tremuloides Michx.). Holzforschung 2006, 60, 111–122. [Google Scholar]
- Chang, H.M.; Sarkanen, K.V. Species variation in lignin: Effects of species on the rate of Kraft delignification. Tappi J. 1973, 56, 132–134. [Google Scholar]
- Mansfield, S.D.; Kang, K.Y.; Chapple, C. Designed for deconstruction-poplar trees altered in cell wall lignification improve the efficacy of bioethanol production. New Phytol. 2012, 194, 91–101. [Google Scholar] [CrossRef]
- Pettersen, R.C. The chemical composition of wood. Chem. Solid Wood 1984, 57–126. [Google Scholar] [CrossRef]
- Rowell, R.M. Handbook of Wood Chemistry and Wood Composites; CRC Press LLC: Boca Raton, FL, USA, 2005; pp. 1–659. [Google Scholar]
- Mansfield, S.D.; Mooney, C.; Saddler, J.N. Substrate and enzyme characteristics that limit cellulose hydrolysis. Biotechnol. Prog. 1999, 15, 804. [Google Scholar] [CrossRef]
- Baucher, M.; Halpin, C.; Petit-Conil, M.; Boerjan, W. Lignin: Genetic engineering and impact on pulping. Crit. Rev. Biochem. Mole. Biol. 2003, 38, 305–350. [Google Scholar] [CrossRef]
- Ukrainetz, N.K.; Kang, Y.K.; Aitken, S.N.; Stoehr, M.; Mansfield, S.D. Heritability and phenotypic and genetic correlations of coastal Douglas-fir (Pseudotsuga menziesii) wood quality traits. Can. J. For. Res. 2008, 38, 1536–1546. [Google Scholar] [CrossRef]
- Gerendiain, A.Z.; Peltola, H.; Pulkkinen, P.; Jaatinen, R.; Pappinen, A. Differences in fibre properties in cloned Norway spruce (Picea abies). Can. J. For. Res. 2008, 38, 1071–1082. [Google Scholar] [CrossRef]
- Kibblewhite, R.P.; Uprichard, J.M. Kraft pulp qualities of eleven radiata pine clones. Appita J. 1996, 49, 243–250. [Google Scholar]
- Bose, S.K.; Francis, R.C.; Govender, M.; Bush, T.; Spark, A. Lignin content versus syringyl to guaiacyl ratio amongst poplars. Bioresour. Technol. 2009, 100, 1628–1633. [Google Scholar] [CrossRef]
- Del Río, J.C.; Gutiérrez, A.; Hernando, M.; Landín, P.; Romero, J.; Martínez, Á.T. Determining the influence of eucalypt lignin composition in paper pulp yield using Py-GC/MS. J. Anal. Appl. Pyrolysis 2005, 74, 110–115. [Google Scholar] [CrossRef]
- Porth, I.; Klápště, J.; Skyba, O.; Lai, B.S.K.; Geraldes, A.; Muchero, W.; Tuskan, G.A.; Douglas, C.J.; El-Kassaby, Y.A.; Mansfield, S.D. Populus trichocarpa cell wall chemistry and ultrastructure trait variation, genetic control and genetic correlations. New Phytol. 2013, 197, 777–790. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hart, J.F.; De Araujo, F.; Thomas, B.R.; Mansfield, S.D. Wood Quality and Growth Characterization across Intra- and Inter-Specific Hybrid Aspen Clones. Forests 2013, 4, 786-807. https://doi.org/10.3390/f4040786
Hart JF, De Araujo F, Thomas BR, Mansfield SD. Wood Quality and Growth Characterization across Intra- and Inter-Specific Hybrid Aspen Clones. Forests. 2013; 4(4):786-807. https://doi.org/10.3390/f4040786
Chicago/Turabian StyleHart, James F., Francis De Araujo, Barb R. Thomas, and Shawn D. Mansfield. 2013. "Wood Quality and Growth Characterization across Intra- and Inter-Specific Hybrid Aspen Clones" Forests 4, no. 4: 786-807. https://doi.org/10.3390/f4040786