Forest Structure and Fine Root Biomass Influence Soil CO2 Efflux in Temperate Forests under Drought

 , , ,
, , ,
Abstract
:1. Introduction
- Since soil respiration consists of an autotrophic and a heterotrophic component, both soil organic carbon (OC) and root biomass are positively associated with soil CO2 efflux.
- Both forest and soil properties are important drivers of soil CO2 efflux at the landscape scale.
- Forest structure and composition influence soil CO2 efflux through their impact on soil OC, fine root biomass and (soil) climate.
2. Methods
2.1. Study Region
2.2. Forests and Forest Properties
2.3. Soil Properties
2.4. Soil Temperature and Volumetric Water Content
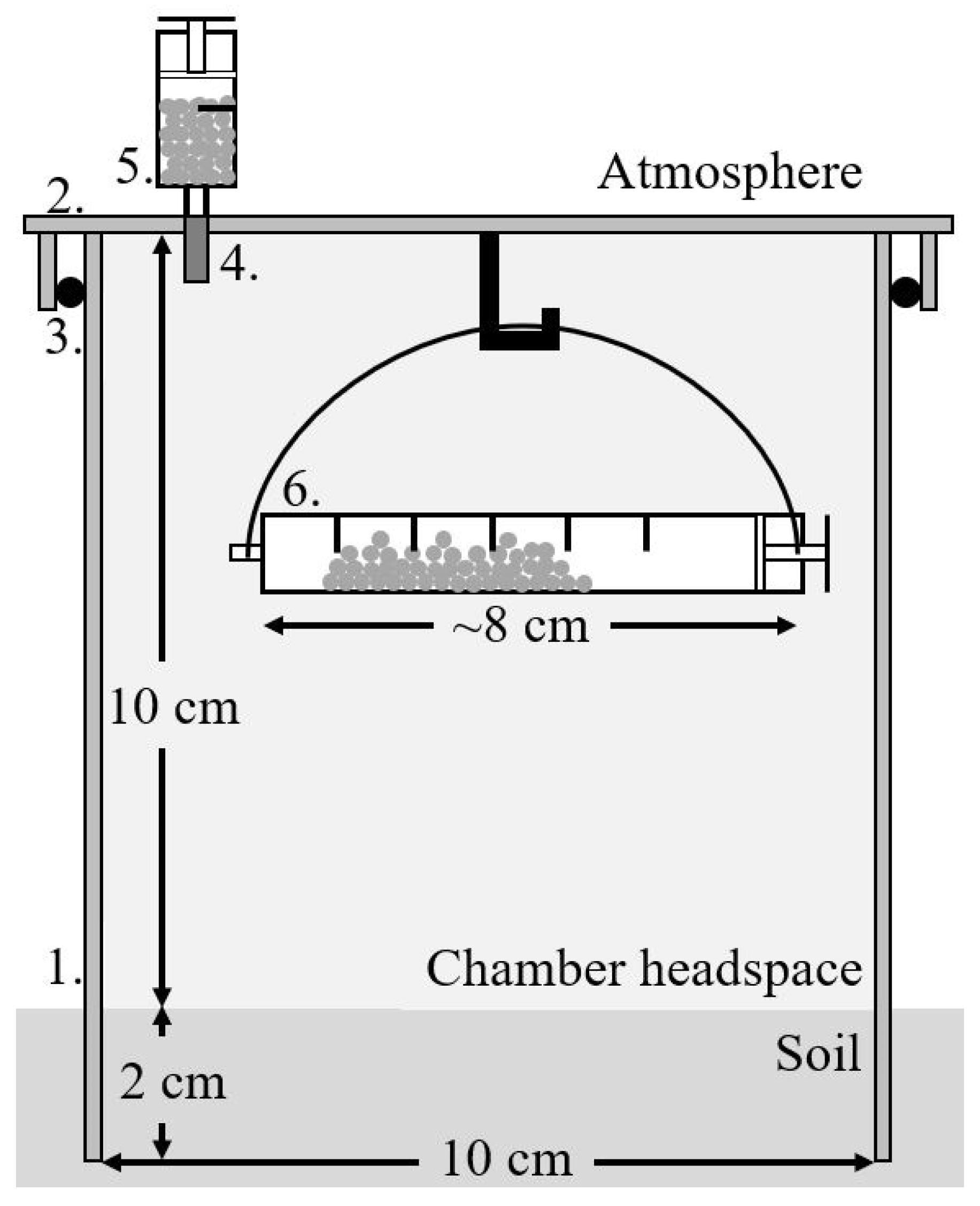
2.5. In Situ Soil CO2 Efflux
2.6. Data Analysis
3. Results
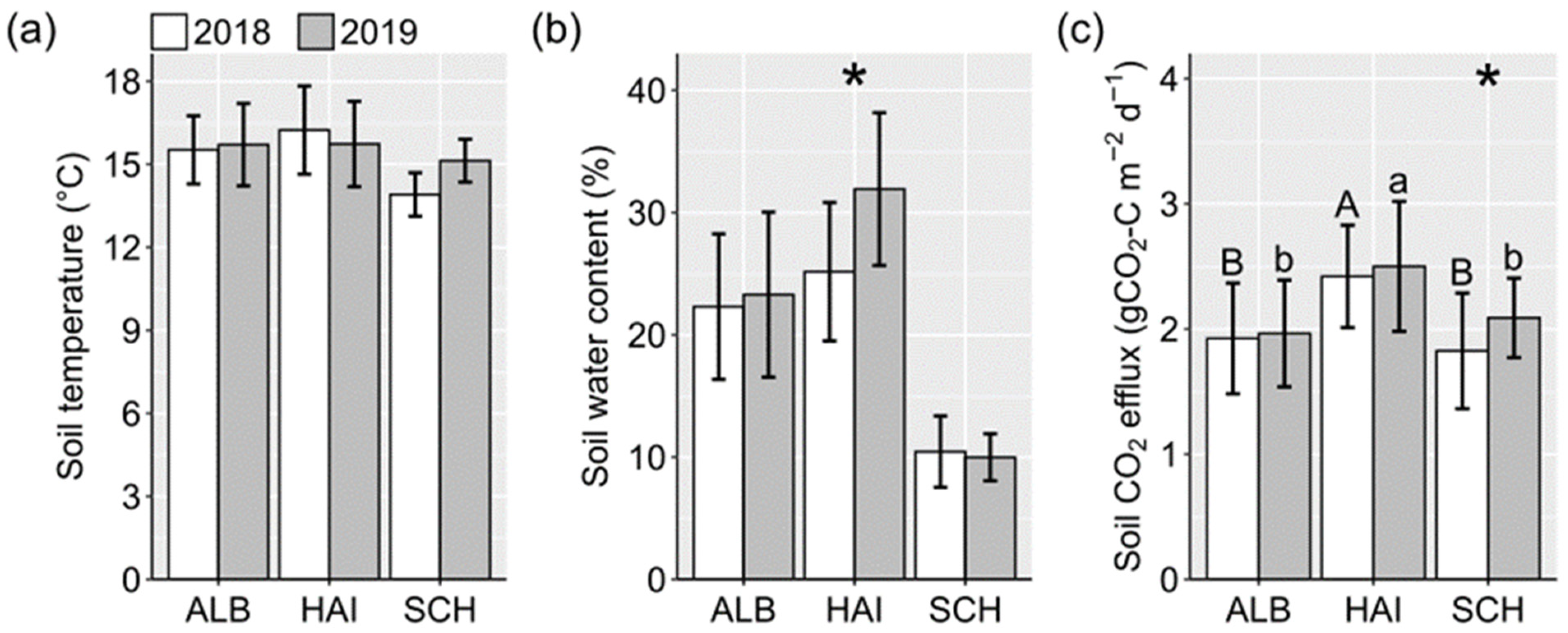
3.1. Forest Soil Temperature and Water Content
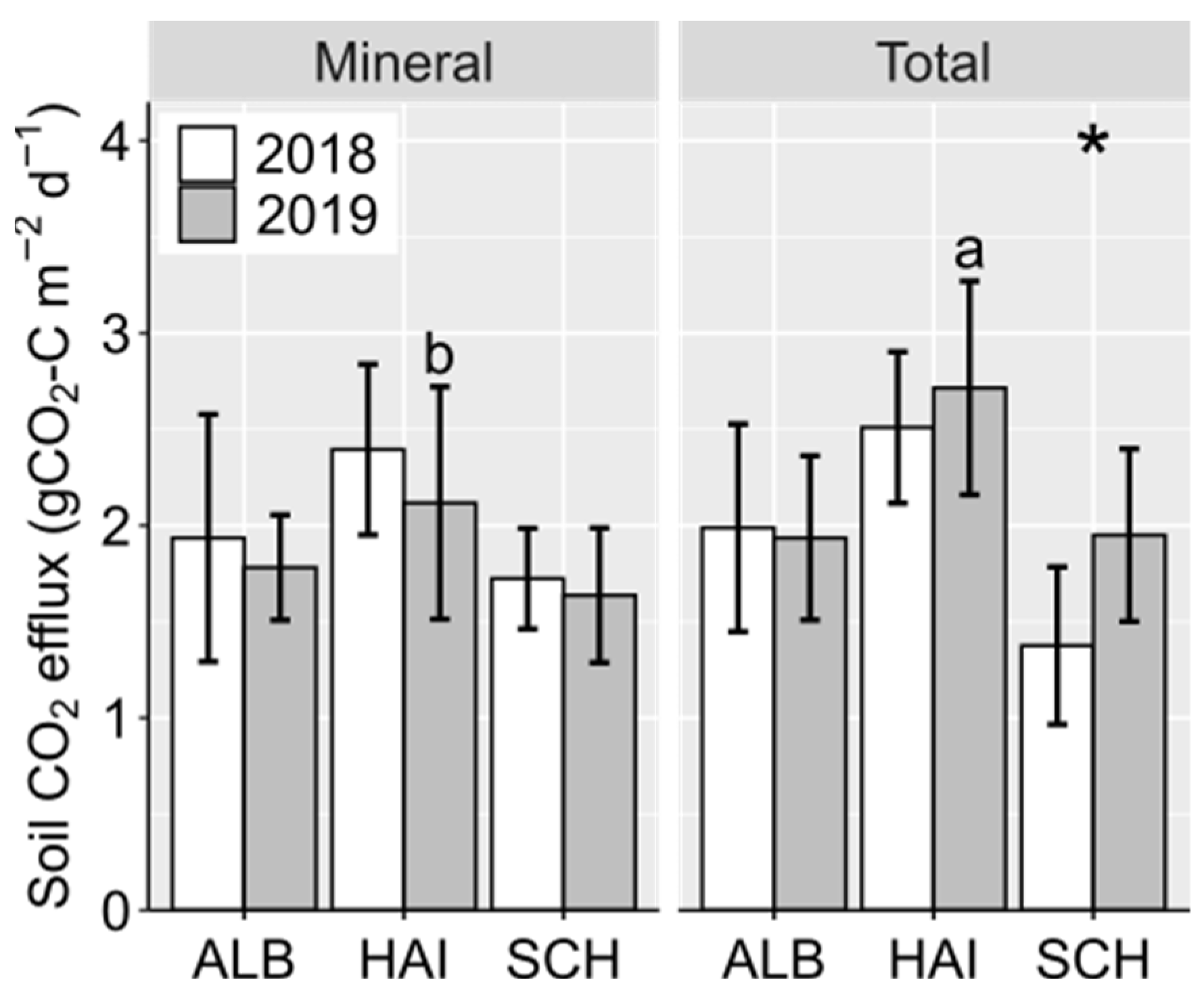
3.2. Forest Soil CO2 Efflux
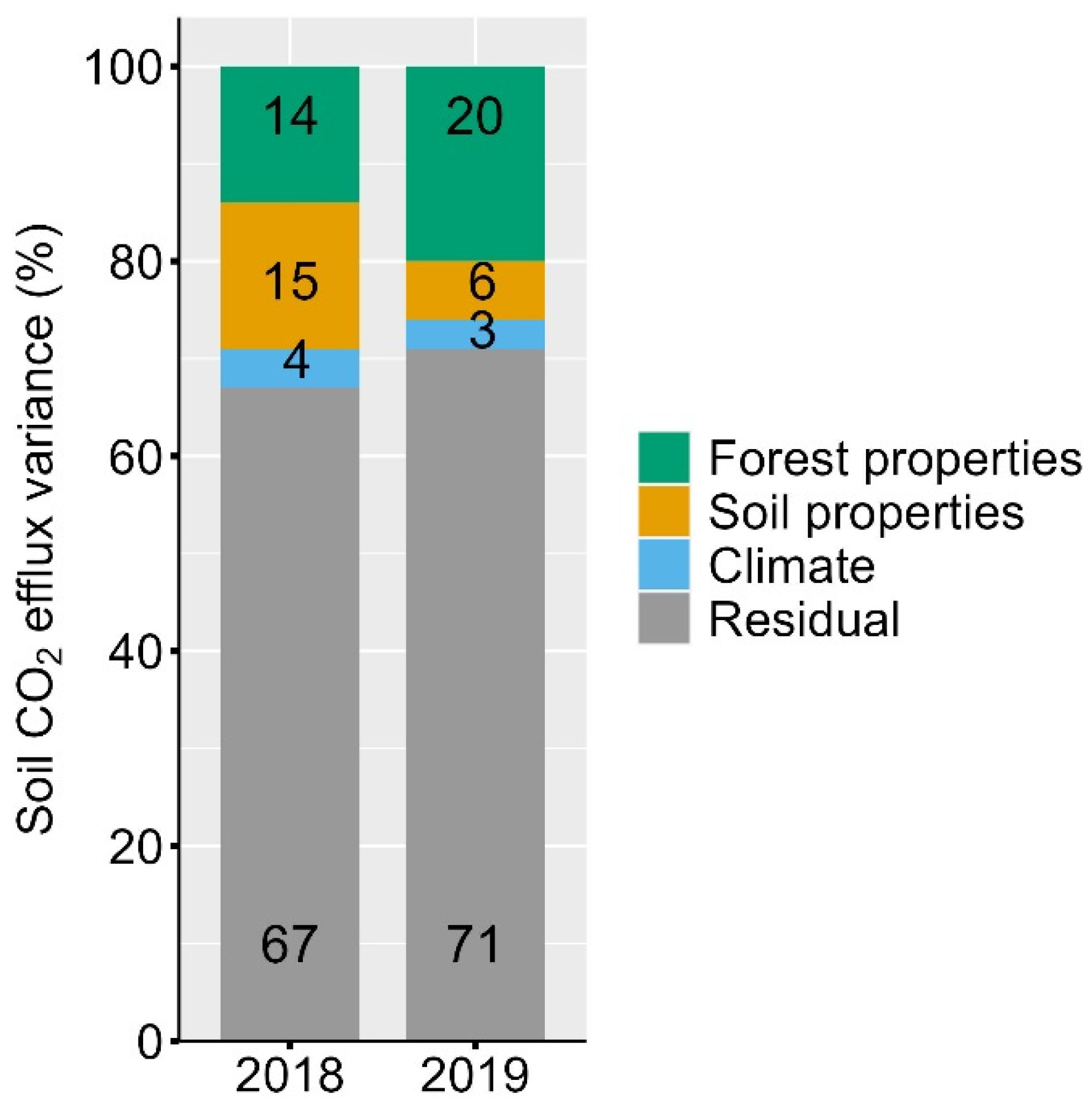
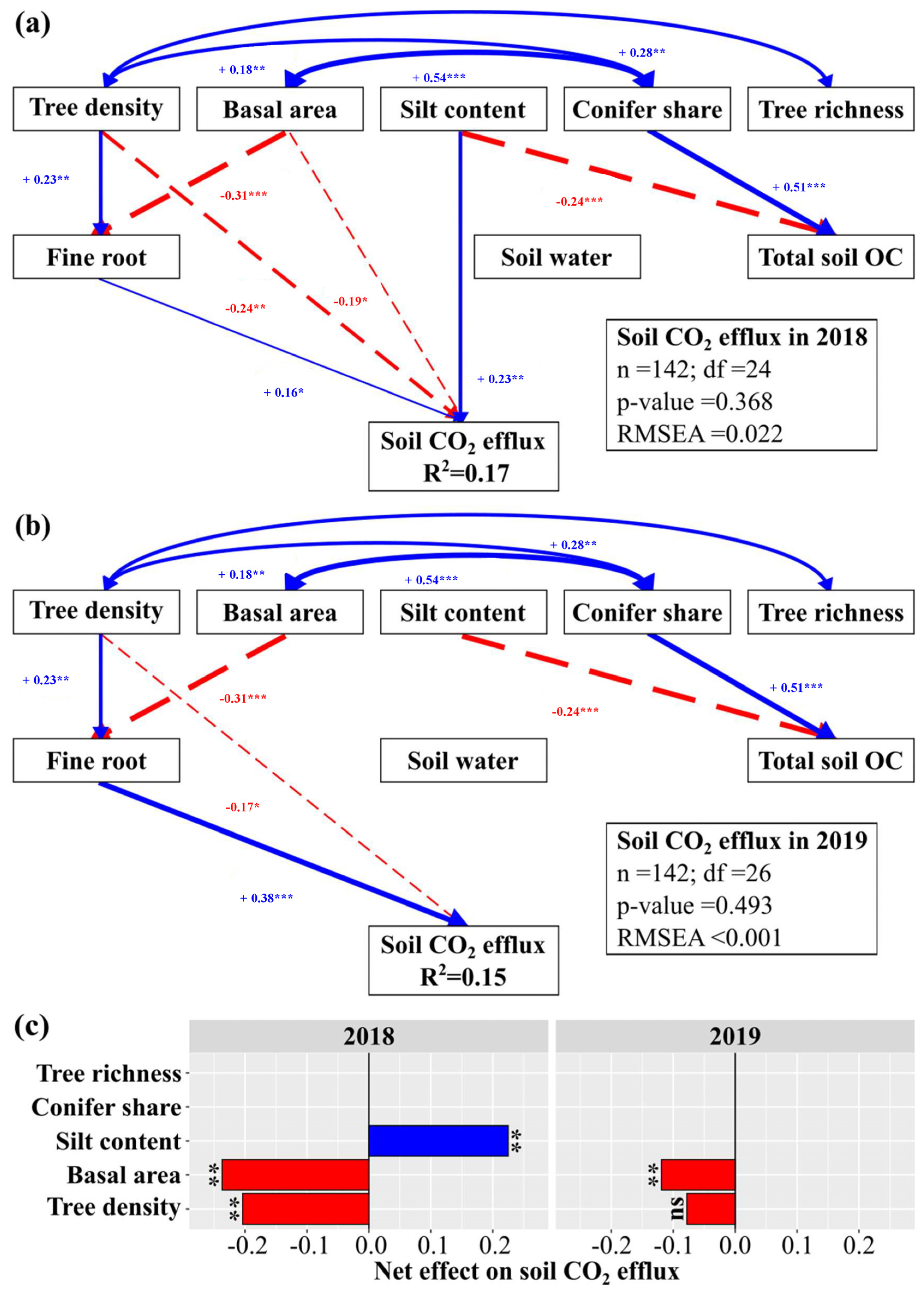
3.3. Forest Properties, Soil Properties and (Soil) Climate Effects on Soil CO2 Efflux
4. Discussion
4.1. Low Soil CO2 Efflux Due to Dry Conditions
4.2. Both Forest and Soil Properties Drive Soil CO2 Efflux at the Landscape under Drought
4.3. Forest Structure, but Not Composition, Influence Soil CO2 Efflux at the Landscape under Drought
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, Y.; Birdsay, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Ehbrecht, M.; Schall, P.; Ammer, C.; Fischer, M.; Seidel, D. Effects of structural heterogeneity on the diurnal temperature range in temperate forest ecosystems. For. Ecol. Manag. 2019, 432, 860–867. [Google Scholar] [CrossRef]
- Felipe-Lucia, M.R.; Soliveres, S.; Penone, C.; Manning, P.; van der Plas, F.; Boch, S.; Prati, D.; Ammer, C.; Schall, P.; Gossner, M.M.; et al. Multiple forest attributes underpin the supply of multiple ecosystem services. Nat. Commun. 2018, 9, 4839. [Google Scholar] [CrossRef] [PubMed]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Factors causing variation in fine root biomass in forest ecosystems. For. Ecol. Manag. 2011, 261, 265–277. [Google Scholar] [CrossRef]
- Rodeghiero, M.; Cescatti, A. Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps. Glob. Chang. Biol. 2005, 11, 1024–1041. [Google Scholar] [CrossRef]
- Kukumägi, M.; Ostonen, I.; Uri, V.; Helmisaari, H.S.; Kanal, A.; Kull, O.; Lõhmus, K. Variation of soil respiration and its components in hemiboreal Norway spruce stands of different ages. Plant Soil 2017, 414, 265–280. [Google Scholar] [CrossRef]
- Zhao, X.; Li, F.; Zhang, W.; Ai, Z.; Shen, H.; Liu, X.; Cao, J.; Manevski, K. Soil respiration at different stand ages (5, 10, and 20/30 years) in coniferous (Pinus tabulaeformis Carrière) and deciduous (Populus davidiana Dode) plantations in a sandstorm Source Area. Forests 2016, 7, 153. [Google Scholar] [CrossRef]
- Berger, T.W.; Inselsbacher, E.; Zechmeister-Boltenstern, S. Carbon dioxide emissions of soils under pure and mixed stands of beech and spruce, affected by decomposing foliage litter mixtures. Soil Biol. Biochem. 2010, 42, 986–997. [Google Scholar] [CrossRef]
- Borken, W.; Xu, Y.J.; Davidson, E.A.; Beese, F. Site and temporal variation of soil respiration in European beech, Norway spruce, and Scots pine forests. Glob. Chang. Biol. 2002, 8, 1205–1216. [Google Scholar] [CrossRef]
- Vesterdal, L.; Elberling, B.; Christiansen, J.R.; Callesen, I.; Schmidt, I.K. Soil respiration and rates of soil carbon turnover differ among six common European tree species. For. Ecol. Manag. 2012, 264, 185–196. [Google Scholar] [CrossRef]
- Jewell, M.D.; Shipley, B.; Low-Décarie, E.; Tobner, C.M.; Paquette, A.; Messier, C.; Reich, P.B. Partitioning the effect of composition and diversity of tree communities on leaf litter decomposition and soil respiration. Oikos 2017, 126, 959–971. [Google Scholar] [CrossRef]
- Schall, P.; Schulze, E.D.; Fischer, M.; Ayasse, M.; Ammer, C. Relations between forest management, stand structure and productivity across different types of Central European forests. Basic Appl. Ecol. 2018, 32, 39–52. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Wang, C.; Gower, S.T. A global relationship between the heterotrophic and autotrophic components of soil respiration? Glob. Chang. Biol. 2004, 10, 1756–1766. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Seperating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Borken, W.; Beese, F. Soil respiration in pure and mixed stands of European beech and Norway spruce following removal of organic horizons. Can. J. For. Res. Can. Rech. For. 2005, 35, 2756–2764. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Ogdahl, M.; Chorover, J.; Chadwick, O.A.; Oleksyn, J.; Zytkowiak, R.; Reich, P.B. Tree species effects on soil organic matter dynamics: The role of soil cation composition. Ecosystems 2007, 10, 999–1018. [Google Scholar] [CrossRef]
- Søe, A.R.B.; Buchmann, N. Spatial and temporal variations in soil respiration in relation to stand structure and soil parameters in an unmanaged beech forest. Tree Physiol. 2005, 25, 1427–1436. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps 2014; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Fischer, M.; Bossdorf, O.; Gockel, S.; Hänsel, F.; Hemp, A.; Hessenmöller, D.; Korte, G.; Nieschulze, J.; Pfeiffer, S.; Prati, D.; et al. Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 2010, 11, 473–485. [Google Scholar] [CrossRef]
- Herold, N.; Schöning, I.; Berner, D.; Haslwimmer, H.; Kandeler, E.; Michalzik, B.; Schrumpf, M. Vertical gradients of potential enzyme activities in soil profiles of European beech, Norway spruce and Scots pine dominated forest sites. Pedobiologia 2014, 57, 181–189. [Google Scholar] [CrossRef]
- Solly, E.F.; Schöning, I.; Boch, S.; Kandeler, E.; Marhan, S.; Michalzik, B.; Müller, J.; Zscheischler, J.; Trumbore, S.E.; Schrumpf, M. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. Plant Soil 2014, 382, 203–218. [Google Scholar] [CrossRef]
- DIN ISO 11277; Soil Quality—Determination of Particle Size Distribution in Mineral Soil Material—Method by Sieving and Sedimentation. Beuth: Berlin, Germany, 2002.
- Grüneberg, E.; Schöning, I.; Hessenmöller, D.; Schulze, E.D.; Weisser, W.W. Organic layer and clay content control soil organic carbon stocks in density fractions of differently managed German beech forests. For. Ecol. Manag. 2013, 303, 1–10. [Google Scholar] [CrossRef]
- Salter, P.J.; Williams, J.B. The influence of texture on the moisture characteristics of soils. II. Available-water capacity and moisture release characteristics. J. Soil Sci. 1965, 16, 310–317. [Google Scholar] [CrossRef]
- Keith, H.; Wong, S.C. Measurement of soil CO2 efflux using soda lime absorption: Both quantitative and reliable. Soil Biol. Biochem. 2006, 38, 1121–1131. [Google Scholar] [CrossRef]
- Grogan, P. CO2 flux measurement using soda lime: Correction for water formed during CO2 adsorption. Ecology 1998, 79, 1467–1468. [Google Scholar] [CrossRef]
- Janssens, I.; Ceulemans, R. Spatial variability in forest soil CO2 efflux assessed with a calibrated soda lime technique. Ecol. Lett. 1998, 1, 95–98. [Google Scholar] [CrossRef]
- McCoy, K.; Zimbron, J.; Sale, T.; Lyverse, M. Measurement of Natural Losses of LNAPL Using CO2 Traps. Groundwater 2015, 53, 658–667. [Google Scholar] [CrossRef]
- Bierbaß, P.; Gutknecht, J.L.M.; Michalzik, B. Nest-mounds of the yellow meadow ant (Lasius flavus) at the “Alter Gleisberg”, Central Germany: Hot or cold spots in nutrient cycling? Soil Biol. Biochem. 2015, 80, 209–217. [Google Scholar] [CrossRef]
- Näthe, K.; Levia, D.F.; Tischer, A.; Michalzik, B. Low-intensity surface fire effects on carbon and nitrogen cycling in soil and soil solution of a Scots pine forest in central Germany. Catena 2018, 162, 360–375. [Google Scholar] [CrossRef]
- Apostolakis, A.; Schöning, I.; Michalzik, B.; Klaus, V.H.; Boeddinghaus, R.S.; Kandeler, E.; Marhan, S.; Bolliger, R.; Fischer, M.; Prati, D.; et al. Drivers of soil respiration across a management intensity gradient in temperate grasslands under drought. Nutr. Cycl. Agroecosyst 2022, 124, 101–116. [Google Scholar] [CrossRef]
- Janssens, I.; Kowalski, A.S.; Longdoz, B.; Ceulemans, R. Assessing forest soil CO2 efflux: An In Situ comparison of four techniques. Tree Physiol. 2000, 20, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Book Series 2009: Statistics for Biology and Health; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Apostolakis, A.; Schöning, I.; Klaus, V.H.; Michalzik, B.; Bischoff, W.-A.; Boeddinghaus, R.S.; Bolliger, R.; Fischer, M.; Hölzel, N.; Kandeler, E.; et al. Direct and plant community mediated effects of management intensity on annual nutrient leaching risk in temperate grasslands. Nutr. Cycl. Agroecosystems 2022, 123, 83–104. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 15 December 2019).
- Faraway, J. Faraway: Functions and Datasets for Books by Julian Faraway, R package version 1.0.7; 2016. Available online: https://CRAN.R-project.org/package=faraway (accessed on 15 December 2019).
- Groemping, U. Relative Importance for Linear Regression in R: The Package relaimpo. J. Stat. Softw. 2006, 17, 1–27. [Google Scholar] [CrossRef]
- Rosseel, Y. lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Nikolova, P.S.; Raspe, S.; Andersen, C.P.; Mainiero, R.; Blaschke, H.; Matyssek, R.; Häberle, K.H. Effects of the extreme drought in 2003 on soil respiration in a mixed forest. Eur. J. For. Res. 2009, 128, 87–98. [Google Scholar] [CrossRef]
- Schindlbacher, A.; Wunderlich, S.; Borken, W.; Kitzler, B.; Zechmeister-Boltenstern, S.; Jandl, R. Soil respiration under climate change: Prolonged summer drought offsets soil warming effects. Glob. Chang. Biol. 2012, 18, 2270–2279. [Google Scholar] [CrossRef]
- Borken, W.; Savage, K.; Davidson, E.A.; Trumbore, S.E. Effects of experimental drought on soil respiration and radiocarbon efflux from a temperate forest soil. Glob. Chang. Biol. 2006, 12, 177–193. [Google Scholar] [CrossRef]
- Muhr, J.; Borken, W. Delayed recovery of soil respiration after wetting of dry soil further reduces C losses from a Norway spruce forest soil. J. Geophys. Res. Biogeosciences 2009, 114, 1–11. [Google Scholar] [CrossRef]
- Shabaga, J.A.; Basiliko, N.; Caspersen, J.P.; Jones, T.A. Seasonal controls on patterns of soil respiration and temperature sensitivity in a northern mixed deciduous forest following partial-harvesting. For. Ecol. Manag. 2015, 348, 208–219. [Google Scholar] [CrossRef]
- Luan, J.; Liu, S.; Wang, J.; Chang, S.X.; Liu, X.; Lu, H.; Wang, Y. Tree species diversity promotes soil carbon stability by depressing the temperature sensitivity of soil respiration in temperate forests. Sci. Total Environ. 2018, 645, 623–629. [Google Scholar] [CrossRef]
- Gan, H.Y.; Schöning, I.; Schall, P.; Ammer, C.; Schrumpf, M. Soil Organic Matter Mineralization as Driven by Nutrient Stoichiometry in Soils Under Differently Managed Forest Stands. Front. For. Glob. Chang. 2020, 3, 1–15. [Google Scholar] [CrossRef]
- Reichstein, M.; Rey, A.; Freibauer, A.; Tenhunen, J.; Valentini, R.; Banza, J.; Casals, P.; Cheng, Y.; Grünzweig, J.M.; Irvine, J.; et al. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Glob. Biogeochem. Cycles 2003, 17, 1104. [Google Scholar] [CrossRef]
- Grüneberg, E.; Schöning, I.; Riek, W.; Ziche, D.; Evers, J. Carbon Stocks and Carbon Stock Changes in German Forest Soils. In Status and Dynamics of Forests in Germany. Ecological Studies (Analysis and Synthesis); Wellbrock, N., Bolte, A., Eds.; Springer: Cham, Switzerland, 2019; Volume 237. [Google Scholar] [CrossRef]
- Saiz, G.; Byrne, K.A.; Butterbach-Bahl, K.; Kiese, R.; Blujdea, V.; Farrell, E.P. Stand age-related effects on soil respiration in a first rotation Sitka spruce chronosequence in central Ireland. Glob. Chang. Biol. 2006, 12, 1007–1020. [Google Scholar] [CrossRef]
- Ammer, C.; Wagner, S. An approach for modelling the mean fine-root biomass of Norway spruce stands. Trees—Struct. Funct. 2005, 19, 145–153. [Google Scholar] [CrossRef]
- Jagodzinski, A.M.; Ziolkowski, J.; Warnkowska, A.; Prais, H. Tree age effects on fine root biomass and morphology over chronosequences of fagus sylvatica, quercus robur and alnus glutinosa stands. PLoS ONE 2016, 11, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Finér, L.; Helmisaari, H.S.; Lõhmus, K.; Majdi, H.; Brunner, I.; Børja, I.; Eldhuset, T.; Godbold, D.; Grebenc, T.; Konôpka, B.; et al. Variation in fine root biomass of three European tree species: Beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosyst. 2007, 141, 394–405. [Google Scholar] [CrossRef]
- Vesterdal, L.; Clarke, N.; Sigurdsson, B.D.; Gundersen, P. Do tree species influence soil carbon stocks in temperate and boreal forests? For. Ecol. Manag. 2013, 309, 4–18. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Schwäbische-Alb | Hainich-Dün | Schorfheide-Chorin |
|---|---|---|---|
| Size (km2) | ~422 | ~1300 | ~1300 |
| Geology | Calcareous bedrock with karst phenomena | Calcareous bedrock | Young glacial landscape |
| Altitude (m asl) | 460–860 | 285–550 | 3–140 |
| AMT (°C) | 6.0–7.0 | 6.5–8.0 | 8.0–8.5 |
| AMP (mm) | 700–1000 | 500–800 | 500–600 |
| Study Region | Schwäbische-Alb | Hainich-Dün | Schorfheide-Chorin |
|---|---|---|---|
| Forest properties | |||
| Stand age (y) | 87 ± 49 b | 133 ± 48 a | 107 ± 40 b |
| Basal area (m2 ha−1) | 30.0 ± 10.0 | 30.5 ± 9.1 | 30.5 ± 7.5 |
| Mean diameter at breast height (cm) | 25.4 ± 10.0 b | 26.6 ± 10.0 b | 33.3 ± 11.3 a |
| Stand density (trees ha−1) | 701 ± 550 a | 568 ± 448 ab | 442 ± 492 b |
| Conifer share (%) | 30.1 ± 40.7 a | 6.6 ± 22.3 b | 37.4 ± 44.2 a |
| Tree species richness (species ha−1) | 6.2 ± 3.1 a | 5.6 ± 2.1 a | 3.7 ± 1.7 b |
| Soil properties | |||
| Organic layer OC stock (kg m−2) | 0.95 ± 0.45 b | 0.59 ± 0.18 b | 1.8 ± 1.3 a |
| Mineral soil OC stock (kg m−2) | 3.8 ± 0.06 a | 3.3 ± 0.07 b | 2.4 ± 0.05 c |
| pH | 5.3 ± 0.8 a | 4.8 ± 0.9 b | 3.5 ± 0.1 c |
| Silt content (g kg−1 soil) | 444.5 ± 107.6 b | 646.0 ± 94.4 a | 84.8 ± 49.2 c |
| Clay content (g kg−1 soil) | 496.1 ± 104.8 a | 301.0 ± 100.3 b | 44.8 ± 18.9 c |
| Total soil depth (cm) | 36.4 ± 21.2 c | 54.5 ± 13.1 b | 88.8 ± 6.3 a |
| Soil CO2 Efflux | 2018 | 2019 | ||
|---|---|---|---|---|
| df | =133 | df | =132 | |
| adj. R2 | =0.39 | adj. R2 | =0.42 | |
| Model p | <0.001 | Model p | <0.001 | |
| Explanatory variable | t-value | p-value | t-value | p-value |
| Intercept | 5.68 | <0.001 | 1.41 | 0.160 |
| Region Hainich-Dün | 1.76 | 0.080 | 4.42 | <0.001 |
| Region Schorfheide-Chorin | 1.12 | 0.267 | 0.77 | 0.443 |
| Soil temperature | - | - | 1.52 | 0.130 |
| Soil water content | - | - | 1.78 | 0.077 |
| Tree density | −2.27 | 0.025 | −3.10 | 0.002 |
| Basal area | −2.31 | 0.023 | - | - |
| Conifer share | - | - | 1.92 | 0.057 |
| Tree species richness | - | - | - | - |
| Fine root biomass | 1.58 | 0.116 | 4.87 | <0.001 |
| Organic layer OC stock | - | - | −1.73 | 0.085 |
| Soil C:N ratio | −1.62 | 0.107 | - | - |
| Silt content | 2.41 | 0.017 | - | - |
| Soil pH | 1.77 | 0.080 | 1.66 | 0.100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apostolakis, A.; Schöning, I.; Michalzik, B.; Ammer, C.; Schall, P.; Hänsel, F.; Nauss, T.; Trumbore, S.; Schrumpf, M. Forest Structure and Fine Root Biomass Influence Soil CO2 Efflux in Temperate Forests under Drought. Forests 2023, 14, 411. https://doi.org/10.3390/f14020411
Apostolakis A, Schöning I, Michalzik B, Ammer C, Schall P, Hänsel F, Nauss T, Trumbore S, Schrumpf M. Forest Structure and Fine Root Biomass Influence Soil CO2 Efflux in Temperate Forests under Drought. Forests. 2023; 14(2):411. https://doi.org/10.3390/f14020411
Chicago/Turabian StyleApostolakis, Antonios, Ingo Schöning, Beate Michalzik, Christian Ammer, Peter Schall, Falk Hänsel, Thomas Nauss, Susan Trumbore, and Marion Schrumpf. 2023. "Forest Structure and Fine Root Biomass Influence Soil CO2 Efflux in Temperate Forests under Drought" Forests 14, no. 2: 411. https://doi.org/10.3390/f14020411